Continuous Monoculture of Alfalfa and Annual Crops Influence Soil Organic Matter and Microbial Communities in the Rainfed Loess Plateau of China

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Treatment Description
2.3. Soil Sampling
2.4. Soil C and N
2.5. Microbial Community-Level Physiological Profiling
2.6. Statistical Analysis
3. Results
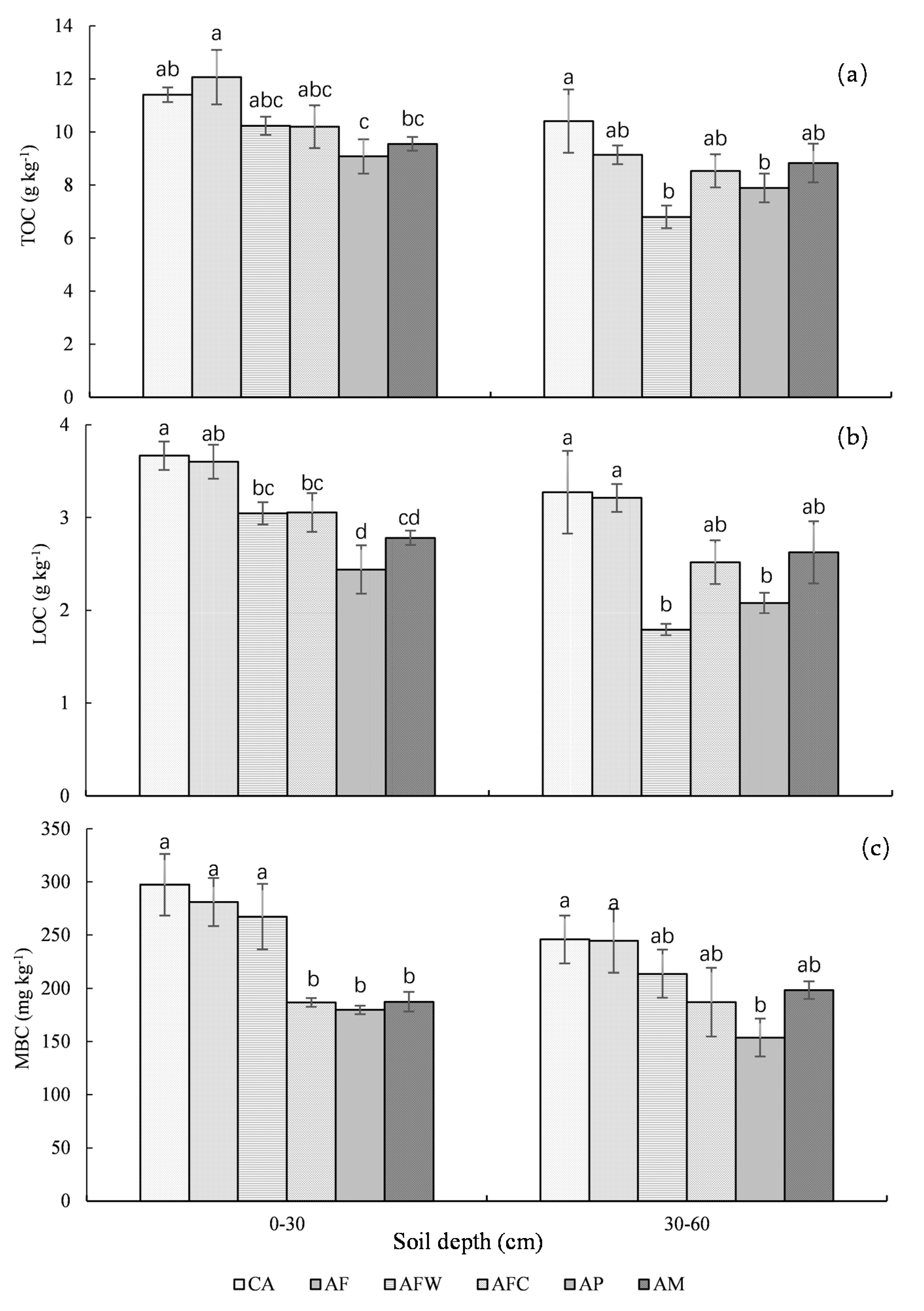
3.1. Soil Total Organic Carbon, Labile Organic Carbon, and Microbial Biomass Carbon
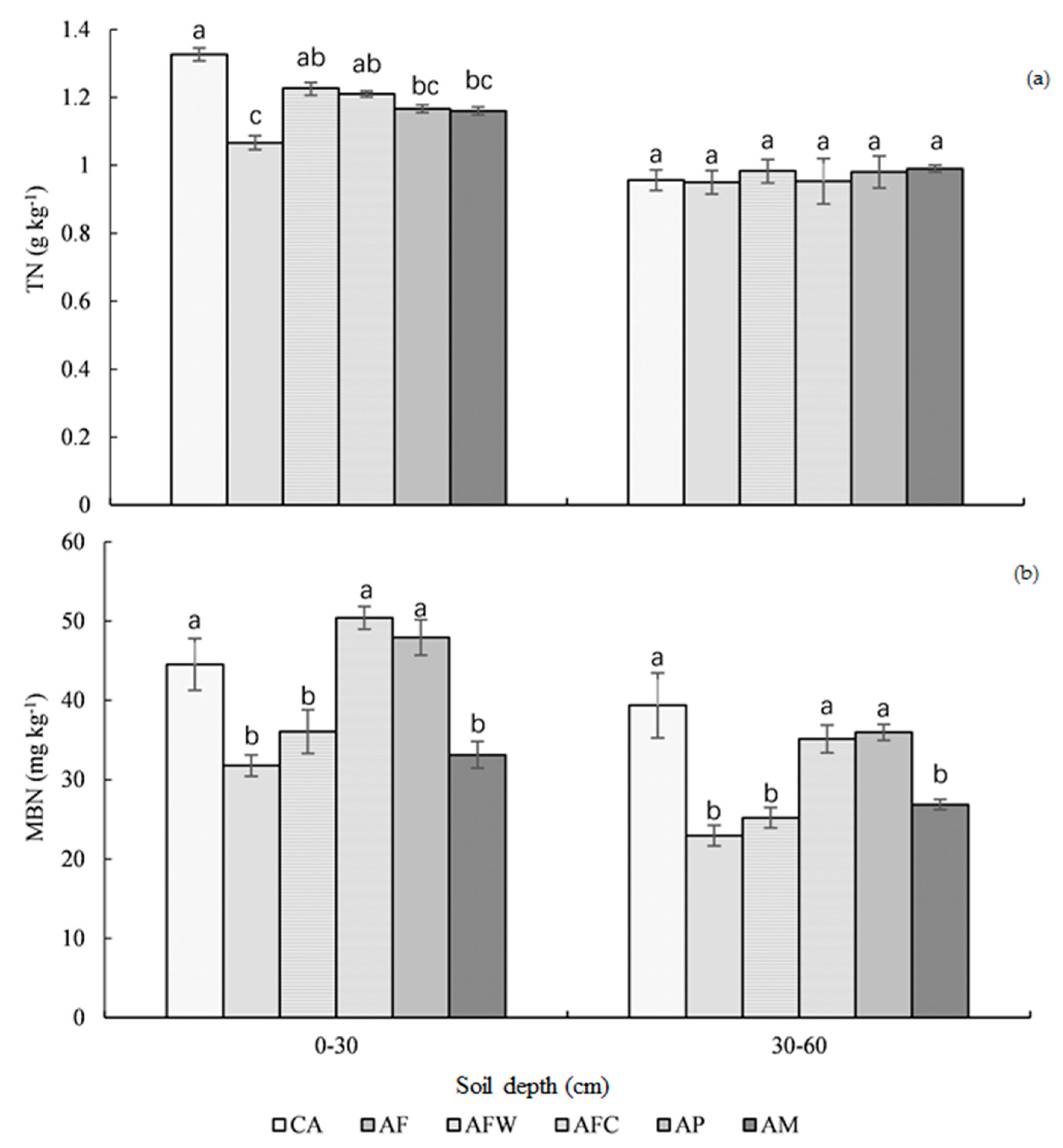
3.2. Soil Total Nitrogen and Microbial Biomass Nitrogen
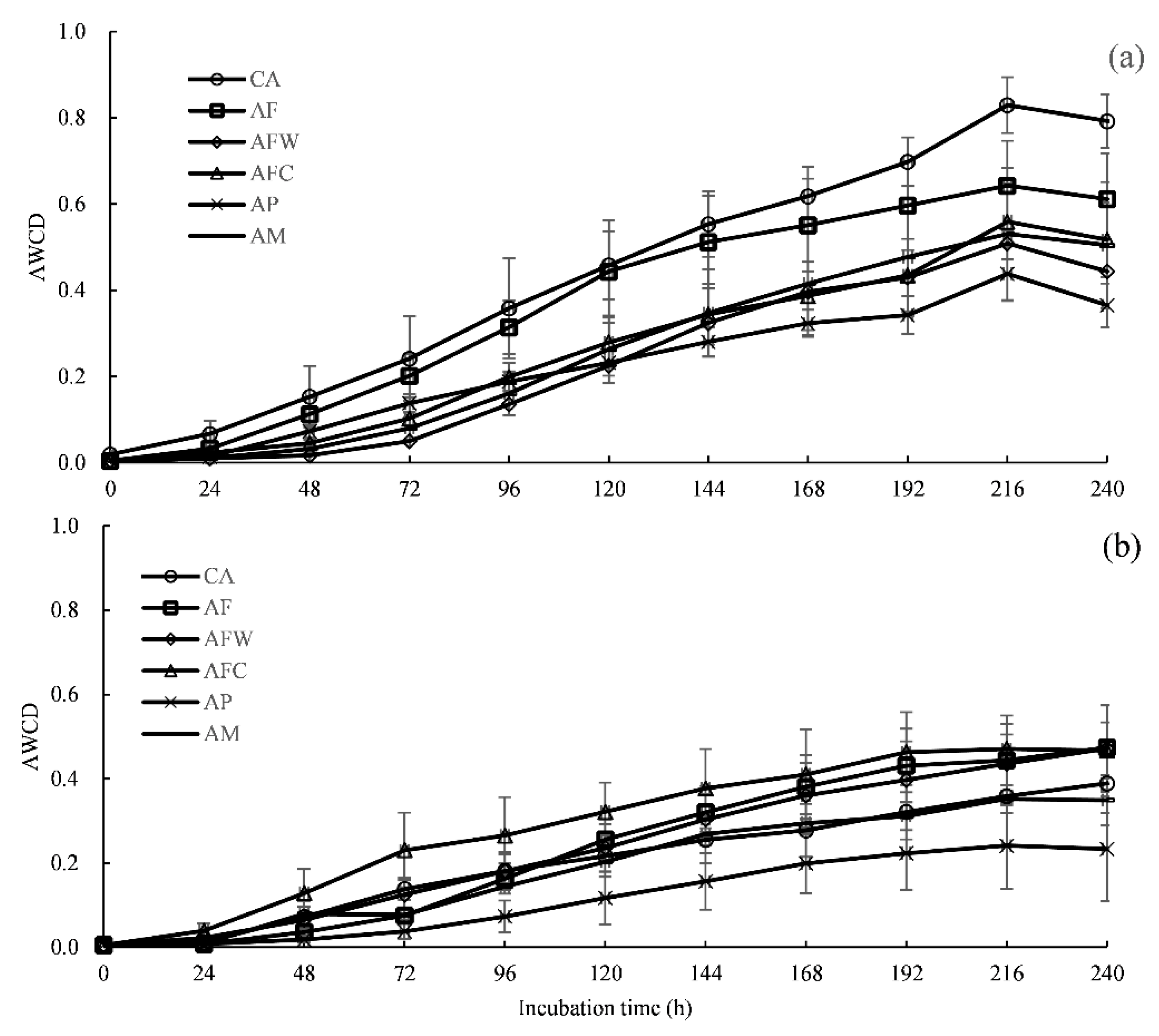
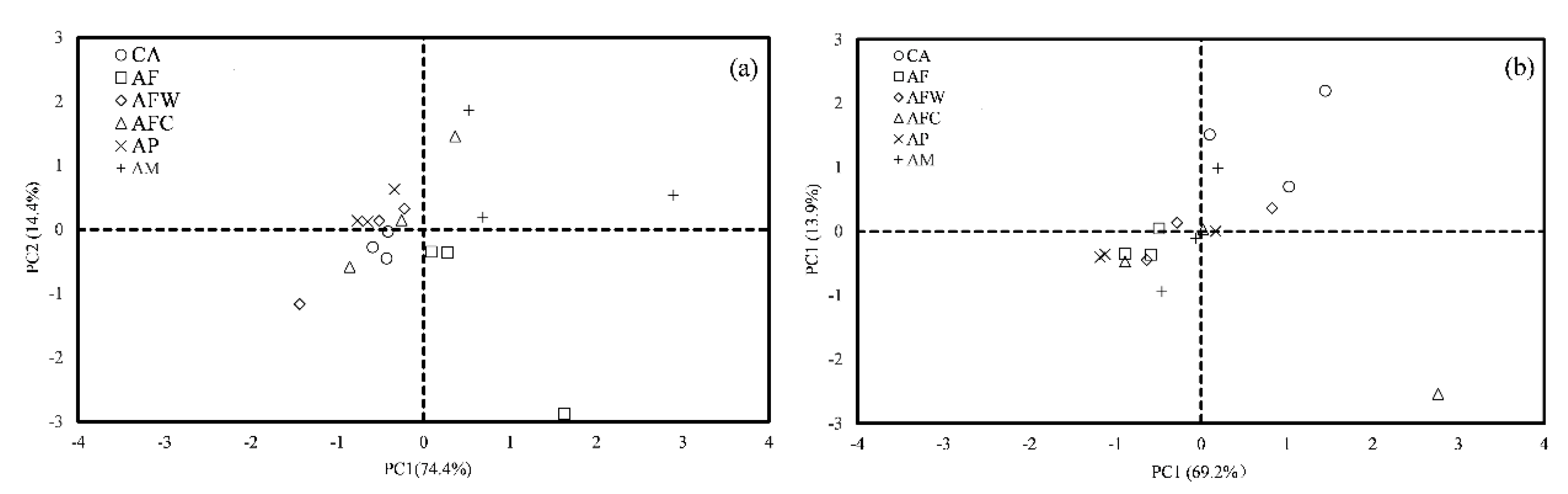
3.3. Community-Level Physiological Profiling
4. Discussion
4.1. Soil C and N
4.2. Community-Level Physiological Profiling
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gan, Y.; Hamel, C.; O’Donovan, J.T.; Cutforth, H.; Zentner, R.P.; Campbell, C.A.; Niu, Y.; Poppy, L. Diversifying crop rotations with pulses enhances system productivity. Sci. Rep. 2015, 5, 14625. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Bainard, L.D.; Bandara, M.; Hamel, C.; Gan, Y. Soil residual water and nutrients explain about 30% of the rotational effect in 4-yr pulse-intensified rotation systems. Can. J. Plant. Sci. 2017, 97, 852–864. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.M.; Kay, B.D. Rotation and tillage effects on soil organic carbon sequestration in a typical Hapludalf in Southern Ontario. Soil Tillage Res. 2001, 59, 107–114. [Google Scholar] [CrossRef]
- Whitbread, A.M.; Blair, G.J.; Lefroy, R.D.B. Managing legume leys, residues and fertilisers to enhance the sustainability of wheat cropping systems in Australia: 2. Soil physical fertility and carbon. Soil Tillage Res. 2000, 54, 77–89. [Google Scholar] [CrossRef]
- McCallum, M.H.; Connor, D.J.; O’Leary, G.L. Water use by lucerne and effect on crops in the Victorian Wimmera. Aust. J. Agric. Res. 2001, 52, 193–201. [Google Scholar] [CrossRef]
- Fan, J.; Shao, M.; Wang, Q.; Jones, S.B.; Fu, X. Toward sustainable soil and water resources use in China’s highly erodible semi-arid loess plateau. Geoderma 2010, 155, 93–100. [Google Scholar] [CrossRef]
- Luo, Z.; Gan, Y.; Niu, Y.; Zhang, R.; Li, L.; Cai, L.; Xie, J. Soil quality indicators and crop yield under long-term tillage systems. Exp. Agric. 2017, 53, 497–511. [Google Scholar] [CrossRef]
- Holford, I.C.R.; Schweitzer, B.E.; Crocker, G.J. Comparative effects of subterranean clover, medic, lucerne, and chickpea in wheat-rotations, on nitrogen, organic carbon, and moisture in two contrasting soils. Aust. J. Soil Res. 1998, 36, 57–72. [Google Scholar] [CrossRef]
- Mccallum, M.H.; Peoples, M.B.; Connor, D.J. Contributions of nitrogen by field pea (Pisum sativum L.) in a continuous cropping sequence compared with a lucerne (Medicago sativa L.)-based pasture ley in the Victorian Wimmera. Crop. Pasture Sci. 2000, 51, 13–22. [Google Scholar] [CrossRef]
- Li, L.L.; Huang, G.B.; Zhang, R.Z.; Cai, L.Q.; Luo, Z.Z.; Jin, X.J.; Zhang, E.H.; Bill, B.; Murray, U. Effects of lucerne removal time on soil water and productivity in a lucerne-wheat rotation on the western Loess Plateau. Acta Agron. Sin. 2011, 37, 686–693. [Google Scholar] [CrossRef]
- Li, Y.S. Productivity dynamic of alfalfa and its effects on water eco-environment. Acta Pedol. Sin. 2002, 39, 404–411. [Google Scholar] [CrossRef]
- Shen, Y.Y.; Li, L.L.; Chen, W.; Robertson, M.; Unkovich, M.; Bellotti, W.; Probert, M. Soil water, soil nitrogen and productivity of lucerne-wheat sequences on deep silt loams in a summer dominant rainfall environment. Field Crops Res. 2009, 111, 97–108. [Google Scholar] [CrossRef]
- Luo, Z.; Li, L.; Niu, Y.; Cai, L.; Zhang, R.; Xie, J. Soil dryness characteristics of alfalfa cropland and optimal growth years of alfalfa on the Loess Plateau of central Gansu, China. Chin. J. Appl. Ecol. 2015, 26, 3059–3065. [Google Scholar]
- Zhang, G.S.; Huang, G.B.; Zhang, R.Z.; Huang, P.; Jin, X.J.; Li, L.L. The effects of lucerne on top soil properties of Huangmian soil. Acta Prataculturae Sin. 2003, 12, 88–93. [Google Scholar]
- Li, J.; Chen, B.; Li, X.F.; Cheng, J.M.; Hao, M.D. Effects of deep soil desiccations on alfalfa grasslands in different rainfall areas of the Loess Plateau of China. Acta Ecol. Sin. 2007, 27, 75–89. [Google Scholar]
- Song, L.P.; Luo, Z.Z.; Li, L.L.; Cai, L.Q.; Zhang, R.Z.; Niu, Y.N. Effect of lucerne-crop rotations on soil physical properties in the semi-arid Loess Plateau of Central Gansu. Acta Prataculturae Sin. 2015, 24, 12–20. [Google Scholar]
- Kotroczó, Z.; Juhos, K.; Biró, B.; Kocsis, T.; Pabar, S.; Fekete, I.; Varga, C. Effect of detritus manipulation on different organic matter decompositions in temperate deciduous forest soils. Forests 2020, 11, 675. [Google Scholar] [CrossRef]
- Lehman, R.; Cambardella, C.; Stott, D.; Acosta-Martinez, V.; Manter, D.; Buyer, J.; Maul, J.; Smith, J.; Collins, H.; Halvorson, J.; et al. Understanding and enhancing soil biological health: The solution for reversing soil degradation. Sustainability 2015, 7, 988–1027. [Google Scholar] [CrossRef] [Green Version]
- Gessner, M.O.; Swan, C.M.; Dang, C.K.; McKie, B.G.; Bardgett, R.D.; Wall, D.H.; Hattenschwiler, S. Diversity meets decomposition. Trends Ecol. Evol. 2010, 25, 372–380. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, Q.; Hu, X.; Zhang, S.; Liu, Y.; Huang, W.; Ding, W. Soil microbial carbon metabolism reveals a disease suppression pattern in continuous ginger mono-cropping fields. Appl. Soil Ecol. 2019, 144, 165–169. [Google Scholar] [CrossRef]
- Nayyar, A.; Hamel, C.; Lafond, G.; Gossen, B.D.; Hanson, K.; Germida, J. Soil microbial quality associated with yield reduction in continuous-pea. Appl. Soil Ecol. 2009, 43, 115–121. [Google Scholar] [CrossRef]
- Yusuf, A.A.; Abaidoo, R.C.; Iwuafor, E.N.O.; Olufajo, O.O.; Sanginga, N. Rotation effects of grain legumes and fallow on maize yield, microbial biomass and chemical properties of an Alfisol in the Nigerian savanna. Agric. Ecosyst. Environ. 2009, 129, 325–331. [Google Scholar] [CrossRef]
- Niu, Y.; Bainard, L.D.; May, W.E.; Hossain, Z.; Hamel, C.; Gan, Y. Intensified pulse rotations buildup pea rhizosphere pathogens in cereal and pulse based cropping systems. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Wang, Y.X.; Chen, Z.R. The nature of soybean-cropping. Soybean Sci. 1991, 10, 31–36. [Google Scholar]
- Lamptey, S.; Li, L.; Xie, J.; Zhang, R.; Luo, Z.; Cai, L.; Liu, J. Soil respiration and net ecosystem production under different tillage practices in semi-arid Northwest China. Plant. Soil Environ. 2017, 63, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Lefroy, R.D.B.; Blair, G.J.; Strong, W.M. Changes in soil organic matter with cropping as measured by organic carbon fractions and 13C natural isotope abundance. Plant. Soil 1993, 155/156, 399–402. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [Green Version]
- Classen, A.T.; Boyle, S.I.; Haskins, K.E.; Overby, S.T.; Hart, S.C. Community-level physiological profiles of bacteria and fungi: Plate type and incubation temperature influences on contrasting soils. FEMS Microbiol. Ecol. 2003, 44, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Kourtev, P.S.; Ehrenfeld, J.G.; Haggblom, M. Exotic plant species alter the microbial community structure and function in the soil. Ecology 2002, 83, 3152–3166. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Qu, L.Y.; Chen, L.D. An amendment on information extraction of Biolog EcoPlateTM. Microbiology 2009, 36, 1083–1091. [Google Scholar]
- Zak, J.C.; Willig, M.R.; Moorhead, D.L.; Wildman, H.G. Functional diversity of microbial communities: A quantitative approach. Soil Biol. Biochem. 1994, 26, 1101–1108. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, G.; Hai, X.; Li, J.; Shangguan, Z.; Peng, C.; Deng, L. Long-term forest succession improves plant diversity and soil quality but not significantly increase soil microbial diversity: Evidence from the Loess Plateau. Ecol. Eng. 2020, 142, 105631. [Google Scholar] [CrossRef]
- Song, M.Y.; Li, Z.P.; Wu, M.; Liu, M.; Jiang, C.Y. Changes in soil microbial biomass and community structure with cultivation chronosequence of greenhouse vegetables. Sci. Agric. Sin. 2015, 48, 3635–3644. [Google Scholar]
- Halvorson, A.D.; Peterson, G.A.; Reule, C.A. Tillage system and crop rotation effects on dryland crop yields and soil carbon in the central Great Plains. Agron. J. 2002, 94, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Halvorson, A.D.; Wienhold, B.J.; Black, A.L. Tillage, nitrogen, and cropping system effects on soil carbon sequestration. Soil Sci. Soc. Am. J. 2002, 66, 906–912. [Google Scholar] [CrossRef]
- Ortega, R.A.; Peterson, G.A.; Westfall, D.G. Residue accumulation and changes in soil organic matter as affected by cropping intensity in no-till dryland agroecosystem. Agron. J. 2002, 94, 944–954. [Google Scholar] [CrossRef]
- Sainju, U.M.; Lenssen, A.W.; Caesar-TonThat, T.; Waddell, J. Tillage and crop rotation effects on dryland soil and residue carbon and nitrogen. Soil Sci. Soc. Am. J. 2006, 70, 668–678. [Google Scholar] [CrossRef]
- Schomberg, H.H.; Jones, O.R. Carbon and nitrogen conservation in dryland tillage and cropping systems. Soil Sci. Soc. Am. J. 1999, 63, 1359–1366. [Google Scholar] [CrossRef]
- Andruschkewitsch, R.; Geisseler, D.; Koch, H.J.; Ludwig, B. Effects of tillage on contents of organic carbon, nitrogen, water-stable aggregates and light fraction for four different long-term trials. Geoderma 2013, 192, 368–377. [Google Scholar] [CrossRef]
- Russell, A.E.; Laird, D.A.; Parkin, T.B.; Mallarino, A.P. Impact of nitrogen fertilization and cropping system on carbon sequestration in midwestern mollisols. Soil Sci. Soc. Am. J. 2005, 69, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Syswerda, S.P.; Corbin, A.T.; Mokma, D.J.; Kravchenko, A.N.; Robertson, G.P. Agricultural management and soil carbon storage in surface vs. deep layers. Soil Sci. Soc. Am. J. 2011, 75, 92–101. [Google Scholar] [CrossRef] [Green Version]
- VandenBygaart, A.J.; Bremer, E.; McConkey, B.G.; Ellert, B.H.; Angers, D.A.; Carter, M.R.; Drury, C.F.; Lafond, G.P.; McKenzie, R.H. Impact of sampling depth on differences in soil carbon stocks in long-term agroecosystem experiments. Soil Sci. Soc. Am. J. 2011, 75, 226–234. [Google Scholar] [CrossRef]
- Moore, J.M.; Susanne, K.; Tabatabai, M.A. Soil microbial biomass carbon and nitrogen as affected by cropping systems. Biol. Fert. Soils 2000, 31, 200–210. [Google Scholar] [CrossRef]
- Peoples, M.B.; Bowman, A.M.; Gault, R.R.; Herridge, D.F.; McCallum, M.H.; McCormick, K.M.; Norton, R.M.; Rochester, I.J.; Scammell, G.J.; Schwenke, G.D. Factors regulating the contributions of fixed nitrogen by pasture and crop legumes to different farming systems of eastern Australia. Plant Soil 2001, 228, 29–41. [Google Scholar] [CrossRef]
- Paustian, K.; Andren, O.; Janzen, H.H.; Lal, R.; Smith, P.; Tian, G.; Tiessen, H.; Van Noordwijk, M.; Woomer, P.L. Agricultural soils as a sink to mitigate CO2 emissions. Soil Use Manag. 1997, 13, 230–244. [Google Scholar] [CrossRef]
- Sainju, U.M.; Terrill, T.H.; Gelaye, S.; Singh, B.P. Soil aggregation and carbon and nitrogen pools under rhizoma peanut and perennial weeds. Soil Sci. Soc. Am. J. 2003, 67, 146–155. [Google Scholar] [CrossRef]
- Sainju, U.M.; Terrill, T.H.; Gelaye, S.; Singh, B.P. Soil carbon and nitrogen pools under long-term productivity of rhizoma peanut and perennial weeds management systems. Arch. Agron. Soil Sci. 2006, 52, 45–59. [Google Scholar] [CrossRef]
- Wu, T.Y.; Schoenau, J.J.; Li, F.M.; Qian, P.Y.; Malhi, S.S.; Shi, Y.C. Effect of tillage and rotation on organic carbon forms of chernozemic soils in Saskatchewan. J. Plant. Nutr. Soil Sci. 2003, 166, 328–335. [Google Scholar] [CrossRef]
- Luo, Q.; Huang, B.L.; Tang, Z.X.; Lai, L.M.; Wei, W. Carbon source utilization of microbes in saline soil of three vegetation types in Xinjiang, China. Chin. J. Appl. Environ. Biol. 2013, 19, 96–104. [Google Scholar] [CrossRef]
- Buenemann, E.K.; Bossio, D.A.; Smithson, P.C.; Frossard, E.; Oberson, A. Microbial community composition and substrate use in a highly weathered soil as affected by crop rotation and P fertilization. Soil Biol. Biochem. 2004, 36, 889–901. [Google Scholar] [CrossRef]
- Bending, G.D.; Turner, M.K.; Jones, J.E. Interaction between crop residue and soil organic matter quality and the functional diversity of soil microbial communities. Soil Biol. Biochem. 2002, 34, 1073–1082. [Google Scholar] [CrossRef]
- Essel, E.; Li, L.; Deng, C.; Xie, J.; Zhang, R.; Luo, Z.; Cai, L. Evaluation of bacterial and fungal diversity in a long-term spring wheat–field pea rotation field under different tillage practices. Can. J. Soil Sci. 2018, 98, 619–637. [Google Scholar] [CrossRef]
- Silva, A.P.; Babujia, L.C.; Matsumoto, L.S.; Guimarães, M.F.; Hungri, M. Bacterial diversity under different tillage and crop rotation systems in an oxisol of Southern Brazil. Open Agric. J. 2013, 7, 40–47. [Google Scholar] [CrossRef]
- Meng, P.P.; Liu, X.; Qiu, H.Z.; Zhang, W.M.; Zhang, C.H. Fungal population structure and its biological effect in rhizosphere soil ofcontinuously cropped potato. Chin. J. Appl. Ecol. 2012, 23, 3079–3086. [Google Scholar]
- Qin, S.; Yeboah, S.; Xu, X.; Liu, Y.; Yu, B. Analysis on fungal diversity in rhizosphere soil of continuous cropping potato subjected to different furrow-ridge mulching managements. Front. Microbiol. 2017, 8, 845. [Google Scholar] [CrossRef]
- Garland, J.L. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbiol. Ecol. 1997, 24, 289–300. [Google Scholar] [CrossRef]
- Konopka, A.; Oliver, L.; Turco, R.F. The use of carbon substrate utilization patterns in environmental and ecological microbiology. Microb. Ecol. 1998, 35, 103–115. [Google Scholar] [CrossRef]
- Grayston, S.J.; Wang, S.Q.; Campbell, C.D.; Edwards, A.C. Selective influence of plant species on microbial diversity in the rhizosphere. Soil Biol. Biochem. 1998, 30, 369–378. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Abbrev | Description |
|---|---|---|
| Continuous alfalfa | CA | Alfalfa established in 2003 and continuously cropped to 2016 |
| Alfalfa-fallow | AF | Alfalfa established in 2003 and removed in March 2012, field continuously fallowed to 2016 |
| Alfalfa-fallow-wheat | AFW | Alfalfa established in 2003 and removed in March 2012, field fallowed until spring wheat was sown in spring 2013, and spring wheat continuously cropped in 2014–2016 |
| Alfalfa-fallow-maize | AFC | Alfalfa established in 2003 and removed in March 2012, field fallowed until maize was sown in May 2013, and maize continuously cropped in 2014–2016 |
| Alfalfa-potato | AP | Alfalfa established in 2003 and removed in March 2012, potato was sown in May 2012 and continuously cropped in 2013–2016 |
| Alfalfa-millet | AM | Alfalfa established in 2003 and removed in March 2012, millet was sown in May 2012 and continuously cropped in 2013–2016 |
| Soil Depth | Treatment | Shannon’s Diversity | Substrate Evenness | Substrate Richness |
|---|---|---|---|---|
| 0–30 cm | CA | 3.14 ± 0.08a | 0.93 ± 0.02a | 29.33 ± 0.33a |
| AF | 2.95 ± 0.12ab | 0.91 ± 0.02a | 26.00 ± 3.05a | |
| AFW | 2.99 ± 0.03ab | 0.89 ± 0.02a | 28.33 ± 1.20a | |
| AFC | 2.75 ± 0.22bc | 0.84 ± 0.05a | 26.33 ± 2.33a | |
| AP | 2.55 ± 0.10c | 0.88 ± 0.02a | 18.33 ± 0.88b | |
| AM | 2.94 ± 0.02ab | 0.88 ± 0.01a | 28.00 ±0.58a | |
| 30–60 cm | CA | 2.67 ± 0.07a | 0.87 ± 0.01a | 21.33 ± 0.88a |
| AF | 2.55 ± 0.3a | 0.83 ± 0.11a | 21.33 ± 0.33a | |
| AFW | 2.56 ± 0.20a | 0.94 ± 0.04a | 15.33 ± 1.86b | |
| AFC | 2.53 ± 0.23a | 0.83 ± 0.06a | 20.67 ± 1.33a | |
| AP | 2.41 ± 0.25a | 0.88 ± 0.09a | 15.67 ± 0.33b | |
| AM | 2.69 ± 0.12a | 0.91 ± 0.01a | 19.67± 1.85a |
| Carbon Source | Substrate | 0–30 cm Soil Depth | 30–60 cm Soil Depth | ||
|---|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | ||
| Carbohydrates | β-methyl-d-glucoside | 0.897 | −0.162 | 0.659 | −0.574 |
| d-galactonic acid lactone | 0.833 | −0.338 | 0.832 | −0.371 | |
| d-xylose | 0.767 | −0.162 | 0.228 | 0.487 | |
| i-erythritol | 0.718 | 0.385 | 0.554 | 0.084 | |
| d-mannitol | 0.908 | 0.017 | 0.950 | −0.069 | |
| N-acetyl-d-glucosamine | 0.938 | −0.102 | 0.900 | −0.317 | |
| d-cellobiose | 0.874 | −0.032 | 0.810 | −0.454 | |
| α-d-glucose-1-phosphate | 0.849 | −0.250 | 0.788 | −0.489 | |
| α-d-lactose | 0.251 | 0.690 | −0.075 | −0.038 | |
| d,l-α-glycerol phosphate | 0.711 | −0.269 | 0.202 | 0.116 | |
| Amino acids | l-arginine | 0.663 | −0.009 | 0.419 | 0.584 |
| l-asparagine | 0.943 | 0.043 | 0.869 | 0.207 | |
| l-phenylalanine | 0.870 | −0.058 | 0.020 | 0.580 | |
| l-serine | 0.808 | −0.011 | 0.795 | 0.209 | |
| l-threonine | 0.317 | 0.278 | 0.316 | 0.466 | |
| Glycyl-l-glutamic acid | 0.566 | 0.166 | 0.471 | 0.701 | |
| Carboxylic acids | Pyruvic acid methyl ester | 0.437 | 0.532 | 0.567 | 0.481 |
| d-galacturonic acid | 0.685 | −0.306 | 0.740 | −0.112 | |
| γ-hydroxybutyric acid | 0.543 | 0.123 | 0.352 | 0.191 | |
| d-glucosaminic acid | 0.824 | −0.311 | 0.323 | 0.567 | |
| iItaconic acid | 0.614 | 0.133 | −0.076 | 0.430 | |
| α-ketobutyric acid | −0.044 | 0.700 | −0.047 | −0.190 | |
| d-malic acid | 0.803 | 0.025 | −0.007 | −0.077 | |
| Polymers | Tween 40 | 0.588 | 0.214 | 0.833 | 0.262 |
| Tween 80 | 0.695 | 0.356 | 0.748 | 0.214 | |
| α-cyclodextrin | 0.538 | 0.730 | 0.623 | −0.141 | |
| Glycogen | 0.549 | 0.503 | 0.752 | −0.014 | |
| Miscellaneous | 2-hydroxybenzoic acid | 0.756 | −0.046 | 0.228 | 0.495 |
| 4-hydroxybenzoic acid | 0.806 | 0.054 | 0.690 | 0.269 | |
| Amines | Phenylethylamine | 0.823 | −0.321 | 0.515 | −0.295 |
| Putrescine | 0.710 | −0.333 | 0.880 | −0.359 | |
| Measurement | Shannon’s Diversity Index | Substrate Evenness | Substrate Richness |
|---|---|---|---|
| TOC | 0.338 * | −0.158 | 0.535 ** |
| LOC | 0.337 * | −0.066 | 0.511 ** |
| MBC | 0.468 ** | 0.293 | 0.386 * |
| TN | 0.425 ** | 0.09 | 0.517 ** |
| MBN | 0.051 | −0.245 | 0.239 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Y.; Luo, Z.; Cai, L.; Coulter, J.A.; Zhang, Y.; Berti, M. Continuous Monoculture of Alfalfa and Annual Crops Influence Soil Organic Matter and Microbial Communities in the Rainfed Loess Plateau of China. Agronomy 2020, 10, 1054. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10071054
Niu Y, Luo Z, Cai L, Coulter JA, Zhang Y, Berti M. Continuous Monoculture of Alfalfa and Annual Crops Influence Soil Organic Matter and Microbial Communities in the Rainfed Loess Plateau of China. Agronomy. 2020; 10(7):1054. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10071054
Chicago/Turabian StyleNiu, Yining, Zhuzhu Luo, Liqun Cai, Jeffrey A. Coulter, Yaoquan Zhang, and Marisol Berti. 2020. "Continuous Monoculture of Alfalfa and Annual Crops Influence Soil Organic Matter and Microbial Communities in the Rainfed Loess Plateau of China" Agronomy 10, no. 7: 1054. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10071054