Physiological Responses of Tomato and Cucumber Seedlings under Different Light–Dark Cycles

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant material and Precultivation
2.2. Experimental Setup and Light Quality Treatments
2.3. Biomass Parameters
2.4. Root, Stem and Leaf Nutrients
2.5. Pigment Concentrations
2.6. Physiological Parameters
2.7. Statistical Analysis
3. Results
3.1. Biomass Parameters
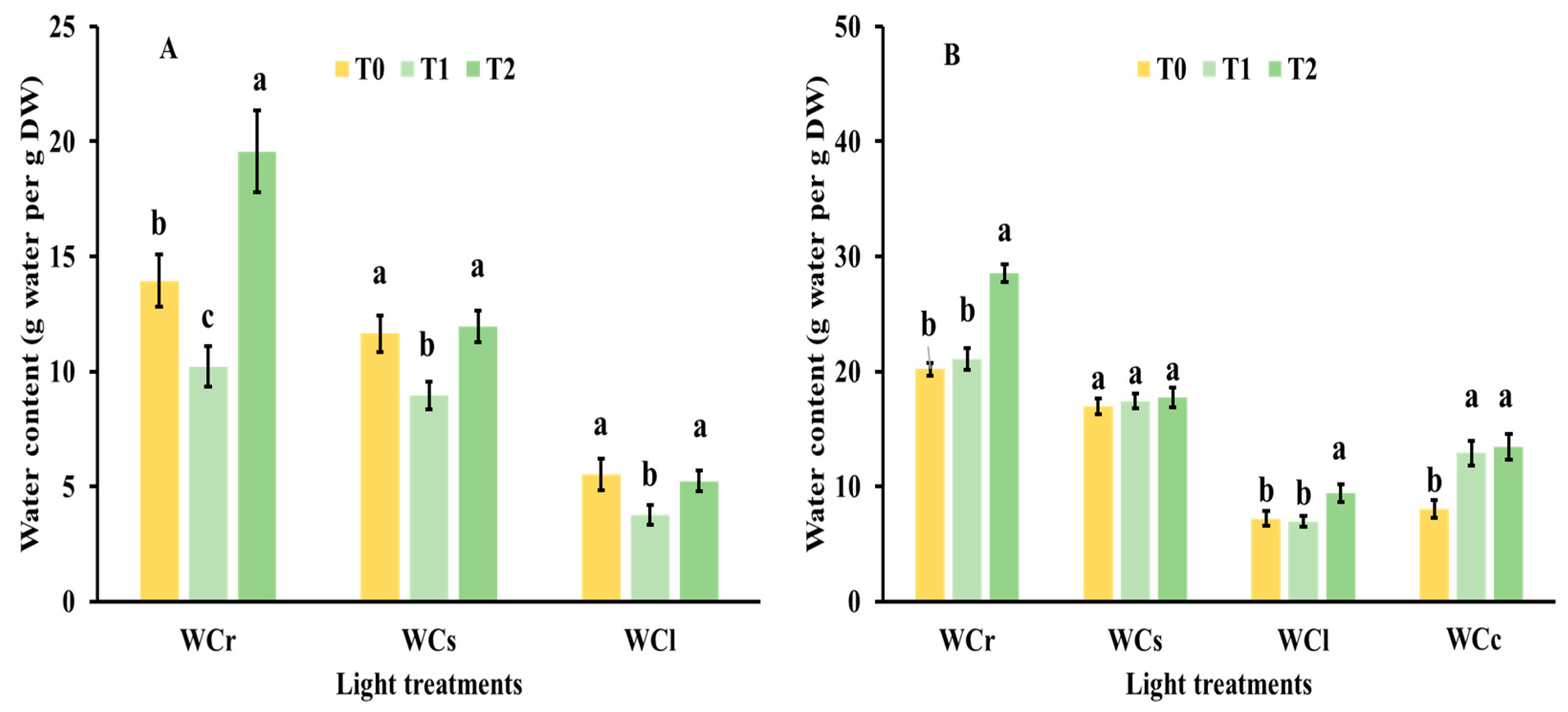
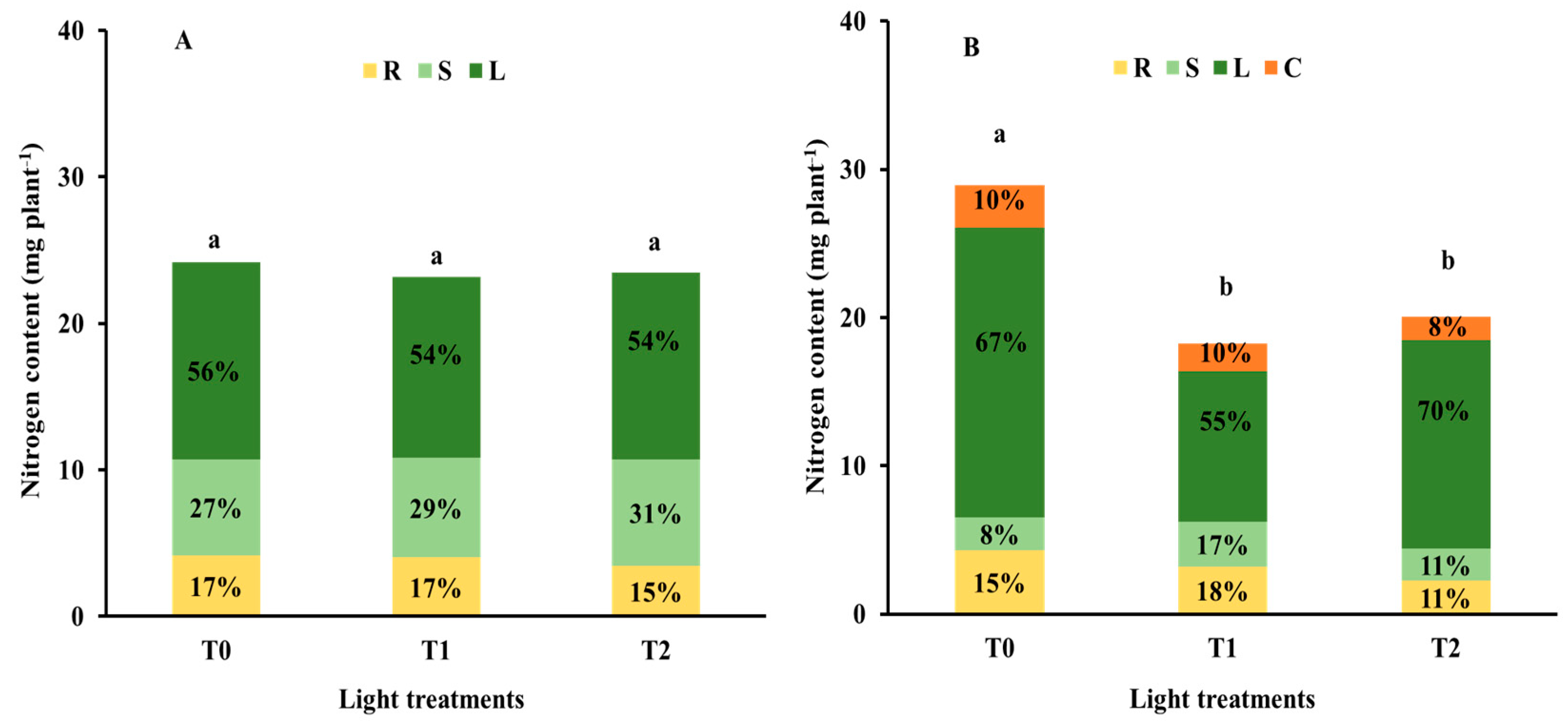
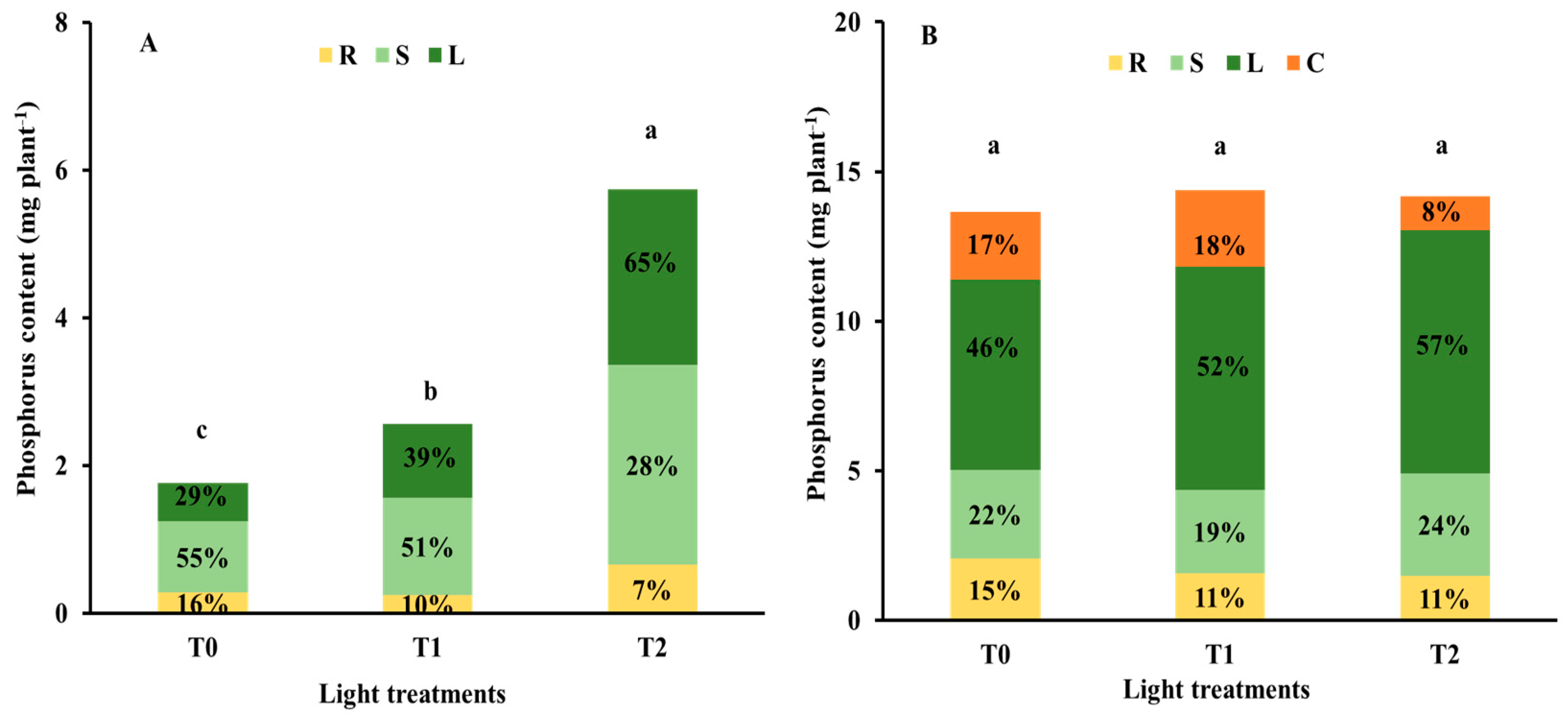
3.2. Nutrient Status
3.3. Pigment Concentrations
3.4. Physiological Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- García-Caparrós, P.; Almansa, E.M.; Chica, R.M.; Lao, M.T. Effects of Artificial Light Treatments on Growth, mineral composition, physiology, and pigment concentration in Dieffenbachia maculata “Compacta” plants. Sustainability 2019, 11, 2867. [Google Scholar] [CrossRef] [Green Version]
- García-Caparros, P.; Chica, R.M.; Almansa, E.M.; Rull, A.; Rivas, L.A.; García-Buendía, A.; Barbero, F.J.; Lao, M.T. Comparisons of different lighting systems for horticultural seedling production aimed at energy saving. Sustainability 2018, 10, 3351. [Google Scholar] [CrossRef] [Green Version]
- García-Caparros, P.; Almansa, E.M.; Barbero, F.J.; Chica, R.M.; Lao, M.T. Fittonia verschaffeltii response to artificial light treatments: Biomass, nutrient concentrations and physiological changes. Agronomy 2020, 10, 126. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.W.; Douwstra, P.; Trouwborst, G.; van Ieperen, W.; Harbinson, J. An artificial solar spectrum substantially alters plant development compared with usual climate room irradiance spectra. J. Exp. Bot. 2010, 61, 1267–1276. [Google Scholar] [CrossRef] [Green Version]
- Hernández, R.; Eguchi, T.; Deveci, M.; Kubota, C. Tomato seedling physiological responses under different percentages of blue and red photon flux ratios using LEDs and cool white fluorescent lamps. Sci. Hortic. 2016, 213, 270–280. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Lin, M.Y.; Mitchell, C.A. Light spectral and thermal properties govern biomass allocation in tomato through morphological and physiological changes. Env. Exp. Bot. 2019, 157, 228–240. [Google Scholar] [CrossRef]
- Groher, T.; Röhlen-Schmittgen, S.; Fiebig, A.; Noga, G.; Hunsche, M. Influence of supplementary LED lighting on physiological and biochemical parameters of tomato (Solanum lycopersicum L.) leaves. Sci. Hortic. 2019, 250, 154–158. [Google Scholar] [CrossRef]
- Van Ieperen, W.; Savvides, A.; Fanourakis, D. Red and blue light effects during growth on hydraulic and stomatal conductance in leaves of young cucumber plants. Acta Hortic. 2012, 956, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, R.; Kubota, C. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using LEDs. Env. Exp. Bot. 2016, 121, 66–74. [Google Scholar] [CrossRef]
- Jackson, S.D. Plant responses to photoperiod. New Phytol. 2009, 181, 517–531. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Bueno, G.; Romero-Campero, F.J.; Lucas-Reina, E.; Romero, J.M.; Valverde, F. Evolution of photoperiod sensing in plants and algae. Curr. Opin. Plant Biol. 2017, 37, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Harmer, S.L.; Hogenesch, J.B.; Straume, M.; Chang, H.S.; Han, B.; Zhu, T.; Wang, X.; Kreps, J.A.; Kay, S.A. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science 2000, 290, 2110–2113. [Google Scholar] [CrossRef] [PubMed]
- Haydon, M.J.; Román, Á.; Arshad, W. Nutrient homeostasis within the plant circadian network. Front. Plant Sci. 2015, 6, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidaka, K.; Okamoto, A.; Araki, T.; Miyoshi, Y.; Dan, K.; Imamura, H.; Kitano, M.; Sameshima, K.; Okimura, M. Effect of photoperiod of supplemental lighting with light-emitting diodes on growth and yield of strawberry. Env. Control. Biol. 2014, 52, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Tsuruyama, J.; Shibuya, T. Growth and flowering responses of seed-propagated strawberry seedlings to different photoperiods in controlled environment chambers. Horttechnology 2018, 28, 453–458. [Google Scholar] [CrossRef]
- Kang, J.H.; KrishnaKumar, S.; Atulba, S.L.S.; Jeong, B.R.; Hwang, S.J. Light intensity and photoperiod influence the growth and development of hydroponically grown leaf lettuce in a closed-type plant factory system. Hort. Env. Biotech. 2013, 54, 501–509. [Google Scholar] [CrossRef]
- Baille, A.; González-Real, M.M.; López, J.C.; Cabrera, J.; Pérez-Parra, J. Characterization of the solar diffuse component under “Parral” plastic greenhouses. Acta Hortic. 2003, 614, 341–346. [Google Scholar] [CrossRef]
- Ben Amor, N.; Ben Hamed, K.; Debez, A.; Grignon, C.; Abdelly, C. Physiological and antioxidant responses of the perennial halophyte Crithmum maritimum to salinity. Plant. Sci. 2005, 168, 889–899. [Google Scholar] [CrossRef]
- Krom, M.D. Spectrophotometric determination of ammonia: Study of a modified Berthelot reaction using salicylate and dicholoroisocyanurate. Analyst 1980, 105, 305–316. [Google Scholar] [CrossRef]
- Hogue, E.; Wilcow, G.E.; Cantliffe, D.J. Effect of soil P on phosphate fraction in tomato leaves. J. Am. Soc. Hortic. Sci. 1970, 95, 174–176. [Google Scholar]
- Lachica, M.; Aguilar, A.; Yanez, J. Análisis foliar: Métodos utilizados en la estación experimental del Zaidín. Anal. Edafol. Agrobiol. 1973, 32, 1033–1047. [Google Scholar]
- Wellburn, A. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvent with spectrophotometers of different resolution. J. Plant. Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Emerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Baret, F.; Guyot, G.; Major, D.J. Crop biomass evaluation using radiometric measurements. Photogrammetria 1989, 43, 241–256. [Google Scholar] [CrossRef]
- Koontz, H.V.; Prince, R.P. Effect of 16 and 24 hours daily radiation (light) on lettuce growth. Hortscience 1986, 21, 123–124. [Google Scholar] [PubMed]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kevei, E.; Toth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A. Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science 2005, 309, 630–633. [Google Scholar] [CrossRef] [Green Version]
- Graf, A.; Schlereth, A.; Stitt, M.; Smith, A.M. Circadian control of carbohydrate availability for growth in Arabidopsis plants at night. Proc. Natl. Acad. Sci. USA 2010, 107, 9458–9463. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.L.; Kay, S.A. Illuminating the mechanism of the circadian clock in plants. Trends Plant. Sci. 1996, 1, 51–57. [Google Scholar] [CrossRef]
- Holmes, M.G.; Smith, H. The function of phytochrome in the natural environment—II. The influence of vegetation canopies on the spectral energy distribution of natural daylight. Photochem. Photobiol. 1977, 25, 539–545. [Google Scholar] [CrossRef]
- Smith, H.; Whitelam, G.C. The shade avoidance syndrome: Multiple responses mediated by multiple phytochromes. Plant. Cell Env. 1997, 20, 840–844. [Google Scholar] [CrossRef] [Green Version]
- Raven, C.W. Chlorophyll formation and phytochrome. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, June 1973. [Google Scholar]
- Oakenfull, R.J.; Davis, S.J. Shining a light on the Arabidopsis circadian clock. Plant. Cell Env. 2017, 40, 2571–2582. [Google Scholar] [CrossRef] [Green Version]
- Webb, A.A. The physiology of circadian rhythms in plants. New Phytol. 2003, 160, 281–303. [Google Scholar] [CrossRef] [Green Version]
- Possart, A.; Fleck, C.; Hiltbrunner, A. Shedding (far-red) light on phytochrome mechanisms and responses in land plants. Plant. Sci. 2014, 217, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Kubota, C.; de Gelder, A.; Peet, M.M. Greenhouse tomato production. In Tomatoes; CABI: Wallingford, England, 2018; Volume 13, pp. 276–314. [Google Scholar]
- Federer, C.A. A soil-plant-atmosphere model for transpiration and availability of soil water. Water Resour. Res. 1979, 15, 555–562. [Google Scholar] [CrossRef]
- Hotta, C.T.; Gardner, M.J.; Hubbard, K.E.; Baek, S.J.; Dalchau, N.; Suhita, D.; Dodd, A.N.; Webb, A.A. Modulation of environmental responses of plants by circadian clocks. Plant. Cell Env. 2007, 30, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Noordally, Z.B.; Ishii, K.; Atkins, K.A.; Wetherill, S.J.; Kusakina, J.; Walton, E.J.; Kato, M.; Azuma, M.; Tanaka, K.; Hanaoka, M.; et al. Circadian control of chloroplast transcription by a nuclear-encoded timing signal. Science 2013, 339, 1316–1319. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Clarke, V.C. The nitrogen cost of photosynthesis. J. Exp. Bot. 2019, 70, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, R.A.; Stokes, T.L.; Thum, K.; Xu, X.; Obertello, M.; Katari, M.S.; Tanurdzic, M.; Dean, A.; Nero, D.C.; McClung, R.; et al. Systems approach identifies an organic nitrogen-responsive gene network that is regulated by the master clock control gene CCA1. Proc. Natl. Acad. Sci. USA 2008, 105, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydon, M.J.; Bell, L.J.; Webb, A.A.R. Interactions between plant circadian clocks and solute transport. J. Exp. Bot. 2011, 62, 2333–2348. [Google Scholar] [CrossRef]
- Voon, C.P.; Lim, B.L. ATP translocation and chloroplast biology. Nat. Sci. Rev. 2019, 6, 1073–1076. [Google Scholar] [CrossRef] [Green Version]
- Versaw, W.K.; Harrison, M.J.; Samuel, T.; Noble, R.; Parkway, S.N. A chloroplast phosphate transporter, PHT2;1, influences allocation of phosphate within the plant and phosphate-starvation responses. Plant. Cell 2002, 14, 1751–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; He, D.; Ji, F. Effects of light intensity and photoperiod on runner plant propagation of hydroponic strawberry transplants under LED lighting. Int. J. Agr. Biol. Eng. 2019, 12, 2631. [Google Scholar] [CrossRef]
- Yang, Z.; He, W.; Mou, S.; Wang, X.; Chen, D.; Hu, X.; Chen, L.; Bai, J. Plant growth and development of pepper seedlings under different photoperiods and photon flux ratios of red and blue LEDs. Trans. Chin. Soc. Agr. Eng. 2017, 33, 173–180. [Google Scholar]
- Takahashi, S.; Badger, M.R. Photoprotection in plants: A new light on photosystem II damage. Trends Plant. Sci. 2011, 16, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Casal, J.J. Phytochromes, cryptochromes, phototropin: Photoreceptor interactions in plants. Photochem. Photobiol. 2000, 71, 1–11. [Google Scholar] [CrossRef]
- Chen, X.L.; Wang, L.C.; Li, T.; Yang, Q.C.; Guo, W.Z. Sugar accumulation and growth of lettuce exposed to different lighting modes of red and blue LED light. Sci. Rep. 2019, 9, 6926. [Google Scholar] [CrossRef] [Green Version]
- Almansa, E.M.; Espín, A.; Chica, R.M.; Lao, M.T. Bioassimilation behaviour of tomato seedling cultivars under different sources of artificial light. Aust. J. Crop. Sci. 2014, 8, 873. [Google Scholar]
- Graf, A.; Smith, A.M. Starch and the clock: The dark side of plant productivity. Trends Plant. Sci. 2011, 16, 169–175. [Google Scholar] [CrossRef]
- Jieun, P.; Yoogyeong, P.; Byoungryong, J.; Seungjae, H. Growth and anthocyanin content of lettuce as affected by artificial light source and photoperiod in a closedtype plant production system. Kor. J. Hortic. Sci. Technol. 2012, 30, 673–679. [Google Scholar]
- López, A.; Hellín, P.; Fenoll, J.; Flores, P. Effect of temperature and photoperiod on soluble sugar and organic acid concentrations in lettuce. Acta Hortic. 2012, 939, 227–230. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2002; p. 690. [Google Scholar]
- Hayashi, F.; Ichino, T.; Osanai, M.; Wada, K. Oscillation and regulation of proline content by P5CS and ProDH gene expressions in the light/dark cycles in Arabidopsis thaliana L. Plant. Cell Physiol. 2000, 41, 1096–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spectral region (nm) | Irradiance (µmol m−2 s−1) |
|---|---|
| UV (300–400) | 0.59 |
| B (400–500) | 201.83 |
| R (600–700) | 506.58 |
| FR (700–800) | 0.38 |
| PAR (400–700) | 731.41 |
| NIR (700–1100) | 0.37 |
| TOTAL (300–1100) | 732.32 |
| PAR:TOTAL | 1 |
| PAR:NIR | 1973.23 |
| B:R | 0.40 |
| B:FR | 528.61 |
| R:FR | 1326.76 |
| Treatments | T0 | T1 | T2 | |
|---|---|---|---|---|
| S. lycopersicum | Length | 27.38 ± 1.70 b | 31.13 ± 1.85 a | 32.75 ± 2.09 a |
| Internodes number | 4.00 ± 0.20 a | 3.75 ± 0.40 a | 4.25 ± 0.30 a | |
| Shoot diameter | 1.70 ± 0.10 a | 1.38 ± 0.09 b | 1.45 ± 0.10 b | |
| C. sativus | Length | 7.15 ± 0.61 a | 7.12 ± 0.65 a | 7.37 ± 0.67 a |
| Internodes number | 4.00 ± 0.30 a | 3.75 ± 0.40 a | 4.00 ± 0.30 a | |
| Shoot diameter | 1.82 ± 0.11 a | 1.75 ± 0.10 a | 1.73 ± 0.10 a | |
| Light treatments | T0 | T1 | T2 | |
|---|---|---|---|---|
| S. lycopersicum | Chl a | 5.71 ± 0.51 a | 5.47 ± 0.61 a | 5.77 ± 0.47 a |
| Chl b | 3.82 ± 0.31 a | 2.17 ± 0.21 b | 2.11 ± 0.19 b | |
| Chl (a + b) | 9.50 ± 0.61 a | 7.60 ± 0.70 b | 7.85 ± 0.79 b | |
| Chl a/b | 1.50 ± 0.11 b | 2.52 ± 0.23 a | 2.70 ± 0.26 a | |
| Car | 0.01 ± 0.001 c | 0.24 ± 0.02 b | 0.47 ± 0.04 a | |
| C. sativus | Chl a | 3.64 ± 0.33 b | 3.36 ± 0.34 b | 4.45 ± 0.43 a |
| Chl b | 2.52 ± 0.30 a | 1.75 ± 0.18 b | 0.84 ± 0.09 b | |
| Chl (a + b) | 6.14 ± 0.56 a | 5.11 ± 0.40 b | 5.29 ± 0.50 b | |
| Chl a/b | 1.44 ± 0.12 c | 1.92 ± 0.16 b | 5.29 ± 0.50 a | |
| Car | 0.19 ± 0.02 a | 0.20 ± 0.02 a | 0.21 ± 0.02 a | |
| Light Treatments | T0 | T1 | T2 | |
|---|---|---|---|---|
| S. lycopersicum | TSS | 32.06 ± 3.21 a | 20.40 ± 1.94 b | 18.79 ± 1.83 b |
| Proline | 56.66 ± 5.55 a | 36.57 ± 3.48 b | 38.74 ± 3.75 b | |
| C. sativus | TSS | 21.86 ± 1.91 a | 15.48 ± 1.50 b | 16.26 ± 1.62 b |
| Proline | 78.72 ± 7.88 a | 47.26 ± 4.33 b | 35.84 ± 3.08 c | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Caparrós, P.; Sabio, F.; Barbero, F.J.; Chica, R.M.; Lao, M.T. Physiological Responses of Tomato and Cucumber Seedlings under Different Light–Dark Cycles. Agronomy 2020, 10, 945. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10070945
García-Caparrós P, Sabio F, Barbero FJ, Chica RM, Lao MT. Physiological Responses of Tomato and Cucumber Seedlings under Different Light–Dark Cycles. Agronomy. 2020; 10(7):945. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10070945
Chicago/Turabian StyleGarcía-Caparrós, Pedro, Francisco Sabio, Francisco Javier Barbero, Rosa María Chica, and María Teresa Lao. 2020. "Physiological Responses of Tomato and Cucumber Seedlings under Different Light–Dark Cycles" Agronomy 10, no. 7: 945. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10070945