Appraisal of Salt Tolerance under Greenhouse Conditions of a Cucurbitaceae Genetic Repository of Potential Rootstocks and Scions

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions, Plant Material, and Salinity Treatments
2.2. Plant Growth Measurements
2.3. Photosynthetic Pigments Determination
2.4. Electrolyte Leakage Analysis
2.5. Statistical Analysis
3. Results
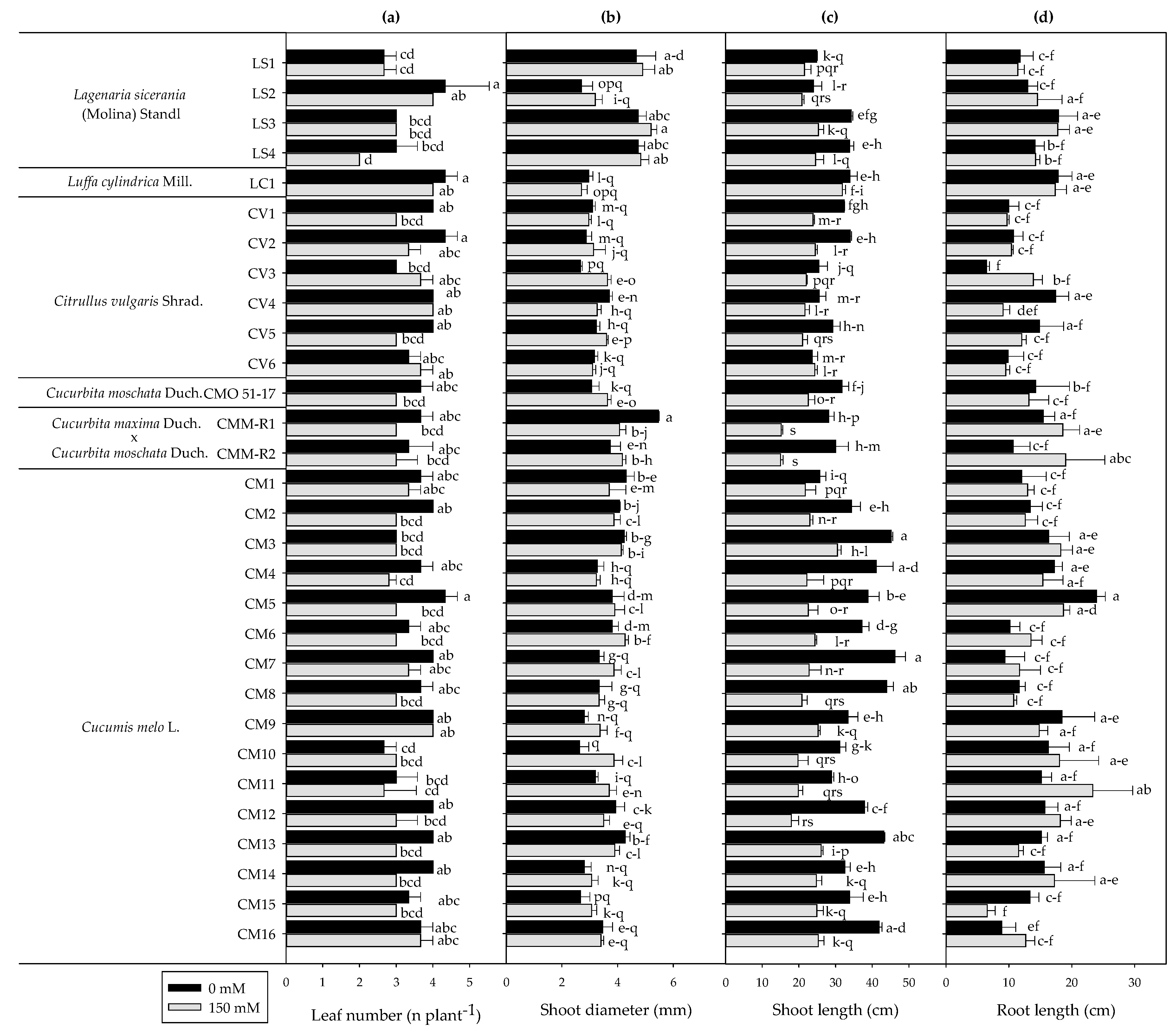
3.1. Plant Growth and Biomass Production
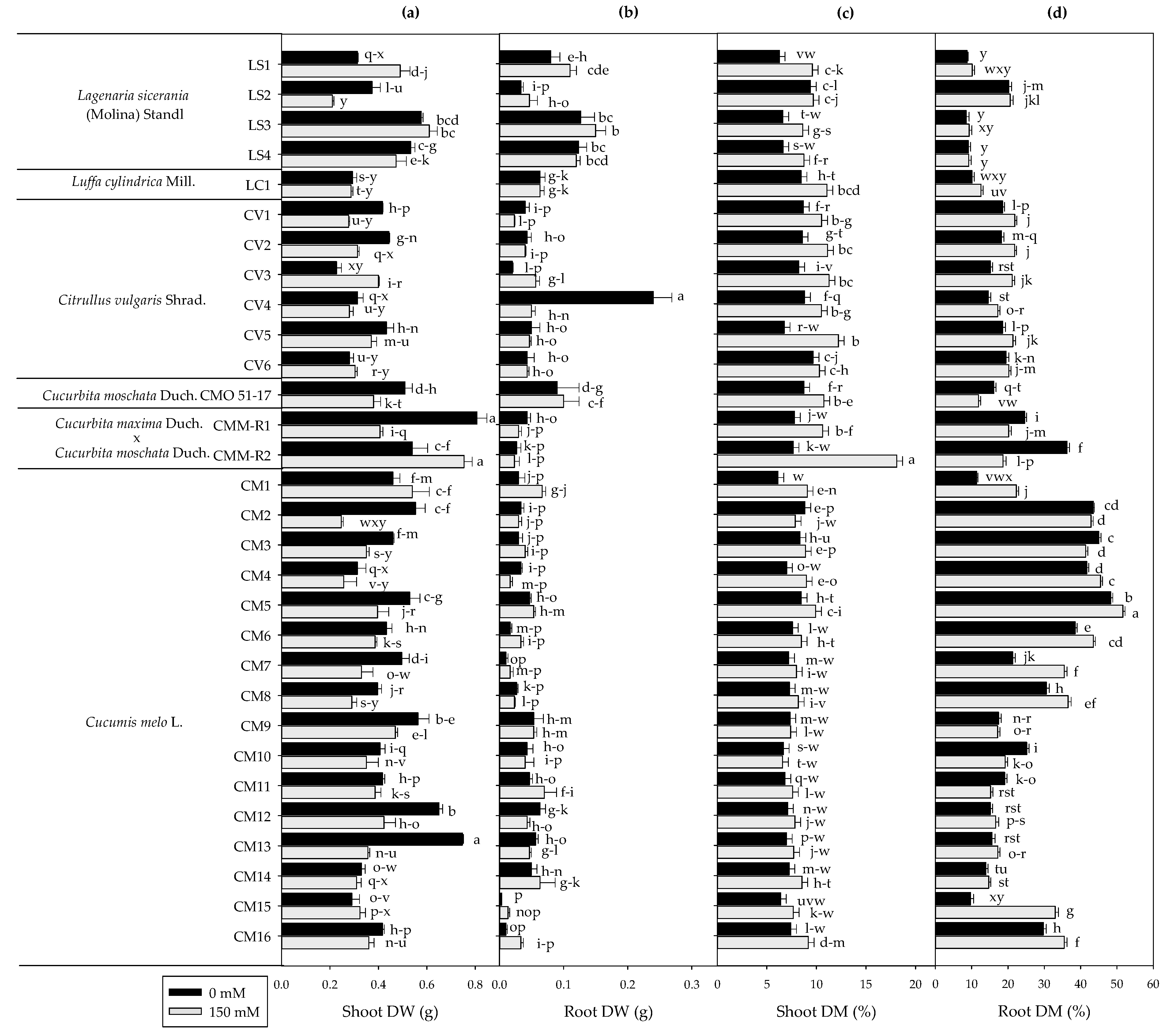
3.2. Plant Biomass Production
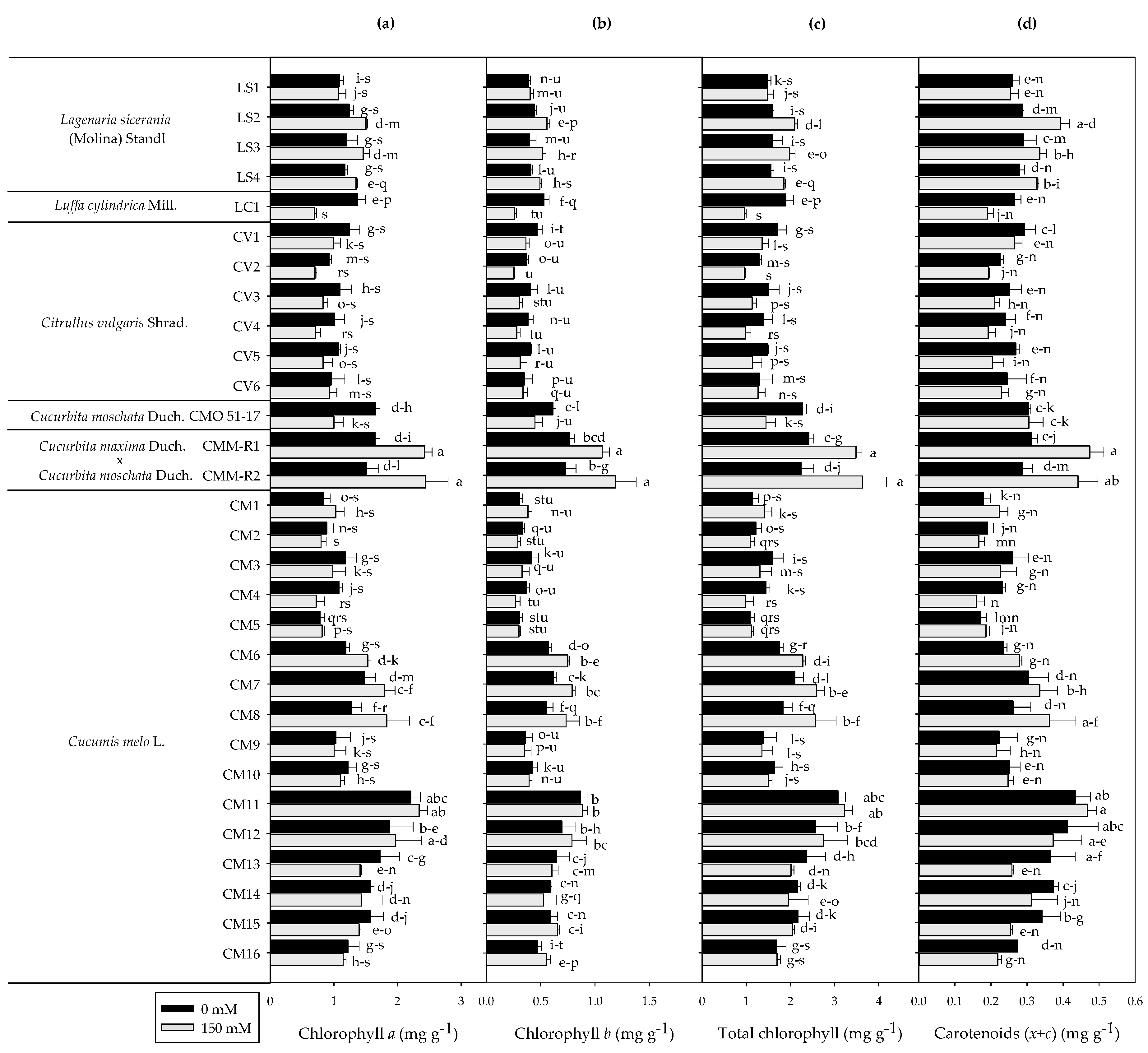
3.3. Photosynthetic Pigments
3.4. Electrolyte Leakage
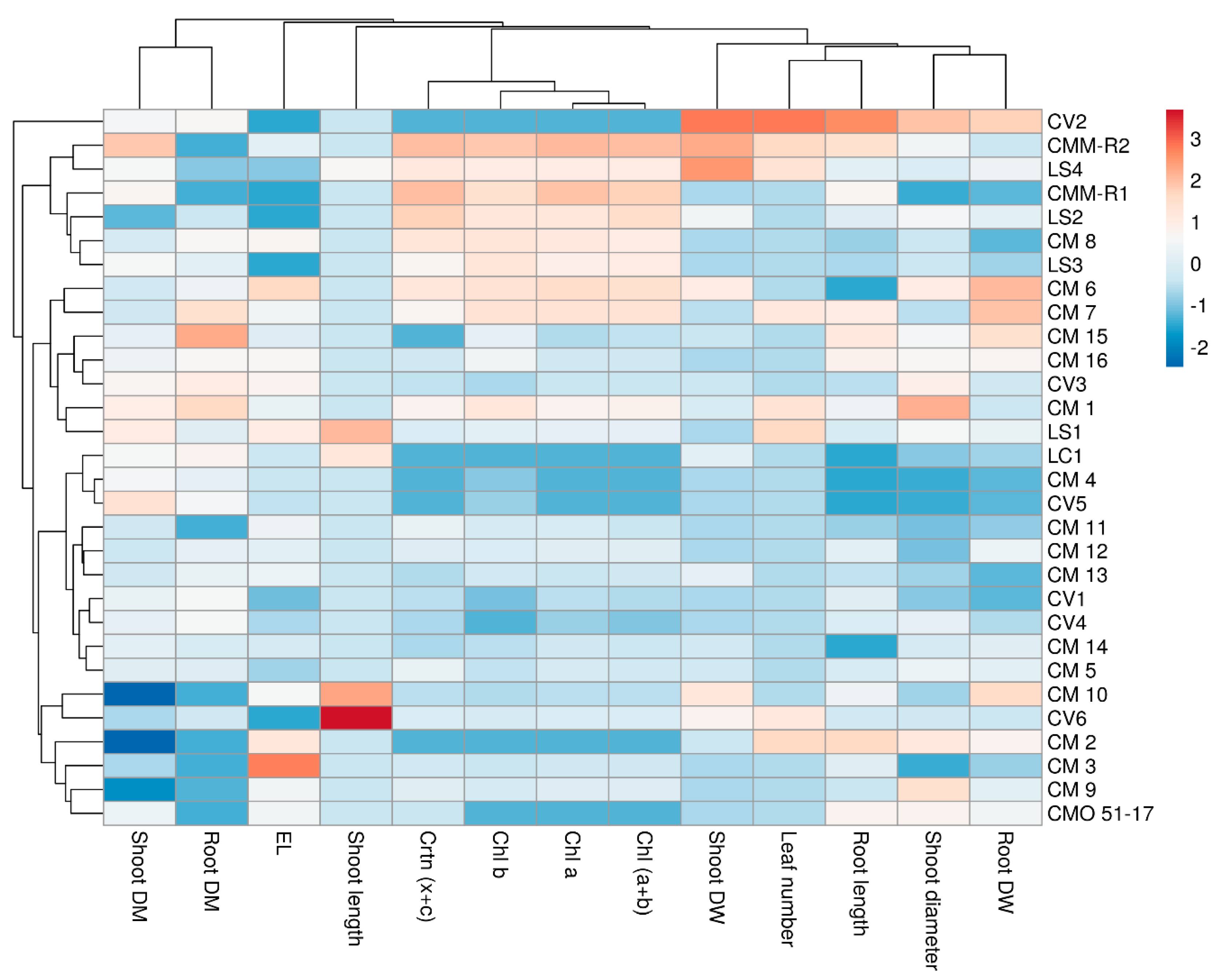
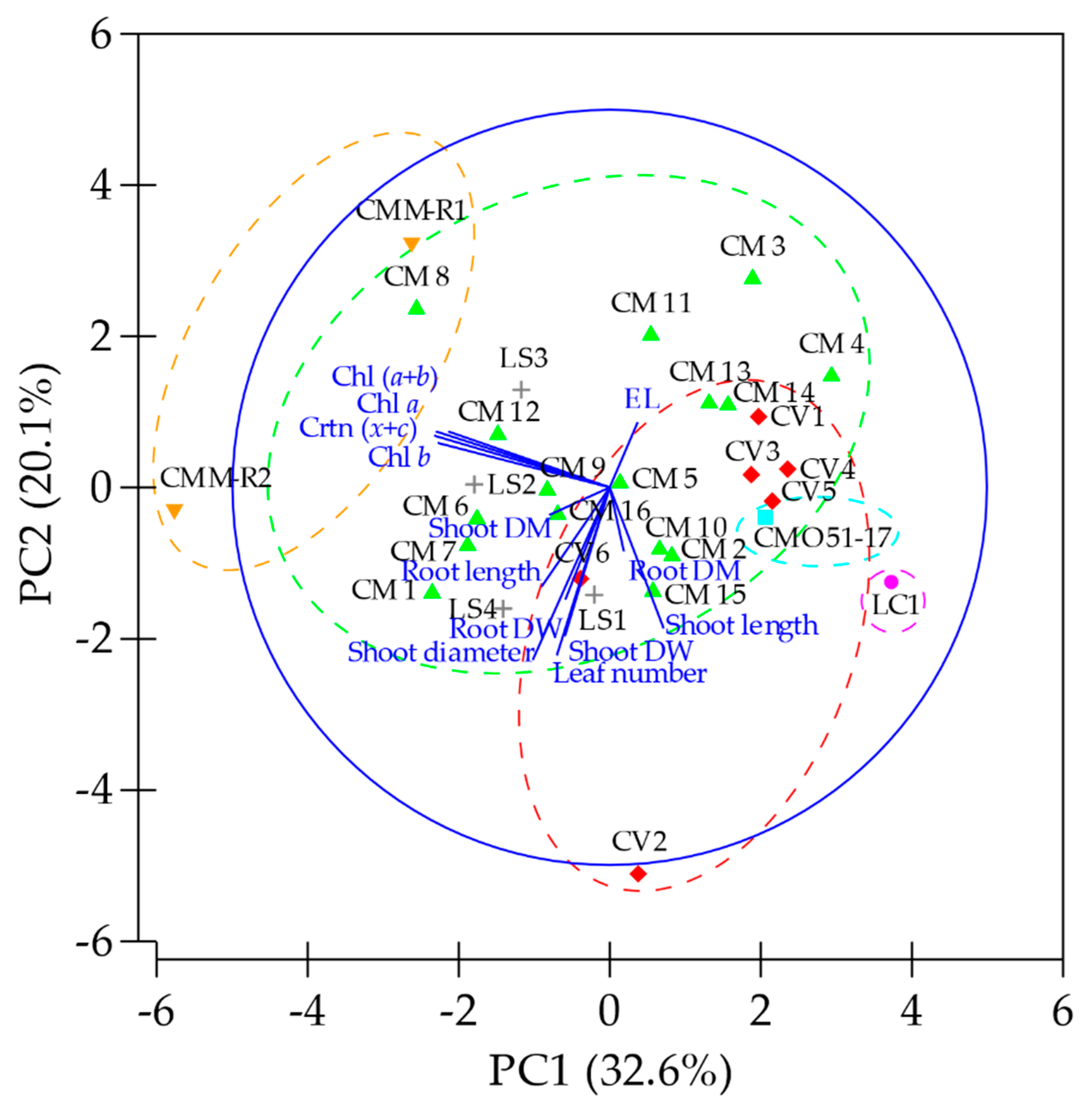
3.5. Cluster Heat Map and Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Colla, G.; Rouphael, Y.; Leonardi, C.; Bie, Z. Role of grafting in vegetable crops grown under saline conditions. Sci. Hortic. 2010, 127, 147–155. [Google Scholar] [CrossRef]
- Montanarella, L.; Badraoui, M.; Chude, V.; Baptista Costa, I.D.S.; Mamo, T.; Yemefack, M.; Singh Aulakh, M.; Yagi, K.; Young Hong, S.; Vijarnsorn, P.; et al. Status of the World’s Soil Resources: Main Report; FAO: Rome, Italy, 2015. [Google Scholar]
- Rouphael, Y.; Cardarelli, M.; Rea, E.; Colla, G. Improving melon and cucumber photosynthetic activity, mineral composition, and growth performance under salinity stress by grafting onto Cucurbita hybrid rootstocks. Photosynthetica 2012, 50, 180–188. [Google Scholar] [CrossRef]
- Balkaya, A.; Yildiz, S.; Horuz, A.; Doğru, S.M. Effects of Salt Stress on Vegetative Growth Parameters and Ion Accumulations in Cucurbit Rootstock Genotypes. J. Crop Breed. Genet. 2016, 2, 11–24. [Google Scholar]
- King, S.R.; Davis, A.R.; Zhang, X.; Crosby, K. Genetics, breeding and selection of rootstocks for Solanaceae and Cucurbitaceae. Sci. Hortic. 2010, 127, 106–111. [Google Scholar] [CrossRef]
- Edelstein, M.; Ben-Hur, M.; Cohen, R.; Burger, Y.; Ravina, I. Boron and salinity effects on grafted and non-grafted melon plants. Plant Soil 2005, 269, 273–284. [Google Scholar] [CrossRef]
- Singh, H.; Kumar, P.; Kumar, A.; Kyriacou, M.C.; Colla, G.; Rouphael, Y. Kumar Grafting Tomato as a Tool to Improve Salt Tolerance. Agronomy 2020, 10, 263. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, D.; Öztekin, G.B.; Tüzel, Y.; Brückner, B.; Krumbein, A. Rootstocks can enhance tomato growth and quality characteristics at low potassium supply. Sci. Hortic. 2013, 149, 70–79. [Google Scholar] [CrossRef]
- Barbieri, G.; Vallone, S.; Orsini, F.; Paradiso, R.; De Pascale, S.; Zakharov, F.; Maggio, A. Stomatal density and metabolic determinants mediate salt stress adaptation and water use efficiency in basil (Ocimum basilicum L.). J. Plant Physiol. 2012, 169, 1737–1746. [Google Scholar] [CrossRef]
- Zhu, J.; Bie, Z.; Huang, Y.; Han, X. Effect of grafting on the growth and ion concentrations of cucumber seedlings under NaCl stress. Soil Sci. Plant Nutr. 2008, 54, 895–902. [Google Scholar] [CrossRef]
- Huang, Y.; Bie, Z.; Liu, P.; Niu, M.; Zhen, A.; Liu, Z.; Lei, B.; Gu, D.; Lu, C.; Wang, B. Reciprocal grafting between cucumber and pumpkin demonstrates the roles of the rootstock in the determination of cucumber salt tolerance and sodium accumulation. Sci. Hortic. 2013, 149, 47–54. [Google Scholar] [CrossRef]
- Xiong, M.; Zhang, X.; Shabala, S.; Shabala, L.; Chen, Y.; Xiang, C.; Nawaz, M.A.; Bie, Z.; Wu, H.; Yi, H.; et al. Evaluation of salt tolerance and contributing ionic mechanism in nine Hami melon landraces in Xinjiang, China. Sci. Hortic. 2018, 237, 277–286. [Google Scholar] [CrossRef]
- Sarabi, B.; Bolandnazar, S.; Ghaderi, N.; Ghashghaie, J. Genotypic differences in physiological and biochemical responses to salinity stress in melon (Cucumis melo L.) plants: Prospects for selection of salt tolerant landraces. Plant Physiol. Biochem. 2017, 119, 294–311. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.R.; Perkins-Veazie, P.; Sakata, Y.; López-Galarza, S.; Maroto, J.V.; Lee, S.-G.; Huh, Y.-C.; Sun, Z.; Miguel, A.; King, S.R.; et al. Cucurbit Grafting. Crit. Rev. Plant Sci. 2008, 27, 50–74. [Google Scholar] [CrossRef]
- Sakata, Y.; Ohara, T.; Sugiyama, M. The hystory of melon and cucumber grafting in Japan. Acta Hortic. 2008, 767, 217–228. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell Membrane Stability as a Measure of Drought and Heat Tolerance in Wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Munns, R.; Passioura, J.; Colmer, T.D.; Byrt, C. Osmotic adjustment and energy limitations to plant growth in saline soil. New Phytol. 2019, 225, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Moradi, F.; Ismail, A.M. Responses of Photosynthesis, Chlorophyll Fluorescence and ROS-Scavenging Systems to Salt Stress During Seedling and Reproductive Stages in Rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sánchez-Blanco, M.J.; Hernández, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Yetişir, H.; Uygur, V. Plant growth and mineral element content of different gourd species and watermelon under salinity stress. Turk. J. Agric. For. 2009, 33, 65–77. [Google Scholar]
- Carillo, P.; Raimondi, G.; Kyriacou, M.C.; Pannico, A.; El-Nakhel, C.; Cirillo, V.; Colla, G.; De Pascale, S.; Rouphael, Y. Morpho-physiological and homeostatic adaptive responses triggered by omeprazole enhance lettuce tolerance to salt stress. Sci. Hortic. 2019, 249, 22–30. [Google Scholar] [CrossRef]
- Geilfus, C.-M.; Zörb, C.; Mühling, K.H. Salt stress differentially affects growth-mediating β-expansins in resistant and sensitive maize (Zea mays L.). Plant Physiol. Biochem. 2010, 48, 993–998. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Dai, H.; Zheng, W.; Cao, F.; Zhang, G.; Sun, D.; Wu, F. Genotypic differences in physiological characteristics in the tolerance to drought and salinity combined stress between Tibetan wild and cultivated barley. Plant Physiol. Biochem. 2013, 63, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Massa, D.; Salerno, A.; Rea, E. Yield, fruit quality and mineral composition of grafted melon plants grown under saline conditions. J. Hortic. Sci. Biotechnol. 2006, 81, 146–152. [Google Scholar] [CrossRef]
- Orsini, F.; Sanoubar, R.; Öztekin, G.B.; Kappel, N.; Tepecik, M.; Quacquarelli, C.; Tüzel, Y.; Bona, S.; Gianquinto, G. Improved stomatal regulation and ion partitioning boosts salt tolerance in grafted melon. Funct. Plant Boil. 2013, 40, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Sanoubar, R.; Orsini, F.; Gianquinto, G. Ionic partitioning and stomatal regulation. Plant Signal. Behav. 2013, 8, e27334. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Rea, E.; Cardarelli, M. Grafting cucumber plants enhance tolerance to sodium chloride and sulfate salinization. Sci. Hortic. 2012, 135, 177–185. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Rea, E. Effect of Salinity on Yield, Fruit Quality, Leaf Gas Exchange, and Mineral Composition of Grafted Watermelon Plants. HortScience 2006, 41, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Gong, B.; Wen, D.; Vandenlangenberg, K.; Wei, M.; Yang, F.; Shi, Q.; Wang, X. Comparative effects of NaCl and NaHCO3 stress on photosynthetic parameters, nutrient metabolism, and the antioxidant system in tomato leaves. Sci. Hortic. 2013, 157, 1–12. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Canaguier, R.; Kumar, P.; Colla, G.; Lucini, L. The effect of a plant-derived biostimulant on metabolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic. 2015, 182, 124–133. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B.; Das, P. NaCl stress causes changes in photosynthetic pigments, proteins, and other metabolic components in the leaves of a true mangrove,Bruguiera parviflora, in hydroponic cultures. J. Plant Boil. 2002, 45, 28–36. [Google Scholar] [CrossRef]
- Munns, R. Plant Adaptations to Salt and Water Stress. Adv. Bot. Res. 2011, 57, 1–32. [Google Scholar]
- Mavrogianopoulos, G.; Spanakis, J.; Tsikalas, P. Effect of carbon dioxide enrichment and salinity on photosynthesis and yield in melon. Sci. Hortic. 1999, 79, 51–63. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, A.L.; Ashraf, M.; Altunlu, H. Improved salt tolerance of melon (Cucumis melo L.) by the addition of proline and potassium nitrate. Environ. Exp. Bot. 2007, 60, 397–403. [Google Scholar] [CrossRef]
- Sevengor, S.; Yasar, F.; Kusvuran, S.; Ellialtioglu, S. The effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidative enzymes of pumpkin seedling. Afr. J. Agric. Res. 2011, 6, 4920–4924. [Google Scholar]
- Tiwari, J.K.; Munshi, A.D.; Kumar, R.; Pandey, R.N.; Arora, A.; Bhat, J.S.; Sureja, A.K. Effect of salt stress on cucumber: Na+–K+ ratio, osmolyte concentration, phenols and chlorophyll content. Acta Physiol. Plant. 2009, 32, 103–114. [Google Scholar] [CrossRef]
- Shahbaz, M.; Noreen, N.; Perveen, S. Triacontanol modulates photosynthesis and osmoprotectants in canola (Brassica napus L.) under saline stress. J. Plant Interact. 2013, 8, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Shahbaz, M.; Ashraf, M.; Athar, H.-R. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul. 2008, 55, 51–64. [Google Scholar] [CrossRef]
- Romero, L.; Belakbir, A.; Ragala, L.; Ruiz, J.M. Response of plant yield and leaf pigments to saline conditions: Effectiveness of different rootstocks in melon plants (Cucumis melo L.). Soil Sci. Plant Nutr. 1997, 43, 855–862. [Google Scholar] [CrossRef] [Green Version]
- Akrami, M.; Arzani, A. Physiological alterations due to field salinity stress in melon (Cucumis melo L.). Acta Physiol. Plant. 2018, 40, 1–14. [Google Scholar] [CrossRef]
- Sivritepe, H. Özkan; Sivritepe, N.; Eris, A.; Turhan, E. The effects of NaCl pre-treatments on salt tolerance of melons grown under long-term salinity. Sci. Hortic. 2005, 106, 568–581. [Google Scholar] [CrossRef]
- Ziaf, K.; Amjad, M.; Pervez, M.A.; Iqbal, Q.; Rajwana, I.A.; Ayyub, M. Evaluation of different growth and physiological traits as indices of salt tolerance in hot pepper (Capsicum annuum L.). Pak. J. Bot. 2009, 41, 1797–1809. [Google Scholar]
- Rivelli, A.R.; De Maria, S.; Pizza, S.; Gherbin, P. Growth and physiological response of hydroponically-grown sunflower as affected by salinity and magnesium levels. J. Plant Nutr. 2010, 33, 1307–1323. [Google Scholar] [CrossRef]
- Bazrafshan, A.H.; Ehsanzadeh, P. Growth, photosynthesis and ion balance of sesame (Sesamum indicum L.) genotypes in response to NaCl concentration in hydroponic solutions. Photosynthetica 2014, 52, 134–147. [Google Scholar] [CrossRef]
- Sanoubar, R.; Cellini, A.; Veroni, A.M.; Spinelli, F.; Masia, A.; Antisari, L.V.; Orsini, F.; Gianquinto, G. Salinity thresholds and genotypic variability of cabbage (Brassica oleracea L.) grown under saline stress. J. Sci. Food Agric. 2015, 96, 319–330. [Google Scholar] [CrossRef]
- García-Valenzuela, X.; Garcìa-Moya, E.; Rascón-Cruz, Q.; Herrera-Estrella, L.; Aguado-Santacruz, G. Chlorophyll accumulation is enhanced by osmotic stress in graminaceous chlorophyllic cells. J. Plant Physiol. 2005, 162, 650–661. [Google Scholar] [CrossRef]
- Wang, X.; Chang, L.; Wang, B.; Wang, D.; Li, P.; Wang, L.; Yi, X.; Huang, Q.; Peng, M.; Guo, A. Comparative proteomics of Thellungiella halophila leaves from plants subjected to salinity reveals the importance of chloroplastic starch and soluble sugars in halophyte salt tolerance. Mol. Cell. Proteomics 2013, 12, 2174–2195. [Google Scholar] [CrossRef] [Green Version]
- Shalata, A.; Neumann, P.M. Exogenous ascorbic acid (vitamin C) increases resistance to salt stress and reduces lipid peroxidation. J. Exp. Bot. 2001, 52, 2207–2211. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genotype | Species | Genotype | ||
|---|---|---|---|---|---|
| Original Code | Working Code | Original Code | Working Code | ||
| Cucumis melo L. | TR31586 | CM1 | Citrullus vulgaris Schrad. | TR40374 | CV1 |
| TR40563 | CM2 | TR64141 | CV2 | ||
| TR43722 | CM3 | TR43211 | CV3 | ||
| TR45883 | CM4 | TR66066 | CV4 | ||
| TR47822 | CM5 | TR43342 | CV5 | ||
| TR48527 | CM6 | TR80748 | CV6 | ||
| TR48611 | CM7 | Lagenaria sicerania (Molina) Standl. | Macis | LS1 | |
| TR49583 | CM8 | TR62066 | LS2 | ||
| TR51531 | CM9 | TR79616 | LS3 | ||
| TR51763 | CM10 | TR82049 | LS4 | ||
| TR61583 | CM11 | Cucurbita maxima Duch. × Cucurbita moschata Duch. | Nun 9075 | CMM-R1 | |
| TR61626 | CM12 | RS841 | CMM-R2 | ||
| TR61851 | CM13 | Cucurbita moschata Duch. | Plovdivski 51-17 | CMO 51-17 | |
| Kirkagac | CM14 | Luffa cylindrica Mill. | LC1 | ||
| Arava | CM15 | ||||
| Cesme | CM16 | ||||
| Leaf Number | Shoot Length | Shoot Diameter | Root Length | Shoot DW | Root DW | Shoot DM | Root DM | |
|---|---|---|---|---|---|---|---|---|
| Genotype (G) | ** | *** | ** | ** | ** | ** | ** | ** |
| Salt (NaCl) | ** | ** | ns | ns | ** | ns | ** | ns |
| G × NaCl | ** | ** | ** | *** | ** | ** | ** | ** |
| Chlorophyll a (mg g−1) | Chlorophyll b (mg g−1) | Total Chlorophyll (mg g−1) | Carotenoids (mg g−1) | Electrolyte Leakage (%) | |
|---|---|---|---|---|---|
| Genotype (G) | ** | ** | ** | ** | ** |
| Salt (NaCl) | ns | ns | ns | ns | ** |
| G × NaCl | ** | ** | ** | ** | ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modarelli, G.C.; Rouphael, Y.; De Pascale, S.; Öztekin, G.B.; Tüzel, Y.; Orsini, F.; Gianquinto, G. Appraisal of Salt Tolerance under Greenhouse Conditions of a Cucurbitaceae Genetic Repository of Potential Rootstocks and Scions. Agronomy 2020, 10, 967. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10070967
Modarelli GC, Rouphael Y, De Pascale S, Öztekin GB, Tüzel Y, Orsini F, Gianquinto G. Appraisal of Salt Tolerance under Greenhouse Conditions of a Cucurbitaceae Genetic Repository of Potential Rootstocks and Scions. Agronomy. 2020; 10(7):967. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10070967
Chicago/Turabian StyleModarelli, Giuseppe Carlo, Youssef Rouphael, Stefania De Pascale, Gölgen Bahar Öztekin, Yüksel Tüzel, Francesco Orsini, and Giorgio Gianquinto. 2020. "Appraisal of Salt Tolerance under Greenhouse Conditions of a Cucurbitaceae Genetic Repository of Potential Rootstocks and Scions" Agronomy 10, no. 7: 967. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10070967