Augmenting the Sustainability of Vegetable Cropping Systems by Configuring Rootstock-Dependent Rhizomicrobiomes that Support Plant Protection

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Soil Microorganisms for Biological Control of Plant Diseases
3. How Plants Recruit Antagonist Microbes to Prevent the Infection by Pathogens
4. Defense Mechanisms in Grafted Plants against Soilborne Pathogens
5. Conclusion and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Tena, G. Recruiting microbial bodyguards. Nat. Plants 2018, 4, 857. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2018, 19, 29–37. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Salerno, A.; Rea, E. The effectiveness of grafting to improve alkalinity tolerance in watermelon. Environ. Exp. Bot. 2010, 68, 283–291. [Google Scholar] [CrossRef]
- Louws, F.J.; Rivard, C.L.; Kubota, C. Grafting fruiting vegetables to manage soilborne pathogens, foliar pathogens, arthropods and weeds. Sci. Hortic. 2010, 127, 127–146. [Google Scholar] [CrossRef]
- Poudel, R.; Jumpponen, A.; Kennelly, M.M.; Rivard, C.L.; Gomez-Montano, L.; Garrett, K.A. Rootstocks shape the rhizobiome: Rhizosphere and endosphere bacterial communities in the grafted tomato system. Appl. Environ. Microbiol. 2018, 85. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Bi, H.G.; Wei, Y.Y.; Li, T.; Wang, H.T.; Ai, X.Z. Effect of grafting on rhizosphere soil environment and its relationship with disease resistance and yield of pepper. Chin. J. Appl. Ecol. 2016, 27, 3539–3547. [Google Scholar]
- Zhou, B.L.; Xu, Y.; Yin, Y.L.; Ye, X.L. Effects of different years continuous cropping and grafting on the biological activities of eggplant soil. Chin. J. Appl. Ecol. 2010, 29, 290–294. [Google Scholar]
- Garbeva, P.; van Elsas, J.D.; Veen, J.A. Rhizosphere microbial community and its response to plant species and soil history. Plant Soil 2008, 302, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Viebahn, M.; Doornbos, R.; Wernars, K.; van Loon, L.C.; Smit, E.; Bakker, P.A.H.M. Ascomycete communities in the rhizosphere of fieldgrown wheat are not affected by introductions of genetically modified Pseudomonas putida WCS358r. Environ. Microbiol. 2005, 7, 1775–1785. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Assessment of the diversity and antagonism toward Rhizoctonia solani AG3 of Pseudomonas species in soil from different agricultural regimes. FEMS Microbiol. Ecol. 2004, 47, 51–64. [Google Scholar] [CrossRef]
- McSpadden Gardener, B.B. Diversity and ecology of biocontrol Pseudomonas spp. in agricultural systems. Phytopathology 2007, 97, 221–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reithner, B.; Ibarra-Laclette, E.; Mach, R.L.; Herrera-Estrella, A. Identification of mycoparasitism-related genes in Trichoderma atroviride. Appl. Environ. Microbiol. 2011, 73, 4361–4370. [Google Scholar] [CrossRef] [Green Version]
- Pinto, Z.V.; Cipriano, M.A.P.; dos Santos, A.S.; Pfenning, L.H.; Patrício, F.R.A. Control of lettuce bottom rot by isolates of Trichoderma spp. Summa Phytopathol. 2014, 40, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Salamone, A.L.; Gundersen, B.; Inglis, D.A. Clonostachys rosea, a potential biological control agent for Rhizoctonia solani AG-3 causing black scurf on potato. Biocontrol Sci. Technol. 2018, 28, 895–900. [Google Scholar] [CrossRef]
- Jones, E.E.; Clarkson, J.P.; Mead, A.; Whipps, J.M. Effect of inoculum type and timing of application of Coniothyrium minitans on Sclerotinia sclerotiorum: Control of Sclerotinia disease in glasshouse lettuce. Plant Pathol. 2004, 53, 621–623. [Google Scholar] [CrossRef]
- Mischke, S. Mycoparasitism of selected sclerotia-forming fungi by Sporidesmium sclerotivorum. Can. J. Bot. 1998, 76, 460–466. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Vlami, M.; De Souza, T.J. Antibiotic production by bacterial biocontrol agents. Antonie Leeuwenhoek 2002, 81, 537–547. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; de Bruijn, I.; de Kock, M.J.D. Cyclic lipopeptide production by plant-associated Pseudomonas species: Diversity, activity, biosynthesis and regulation. Mol. Plant-Microbe Interact. 2006, 19, 699–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, S.J.; Kumar, T.; Chung, J.Y.; Ryu, M.; Park, G.; Shim, S.; Hong, S.B.; Seo, H.; Bae, D.W.; Bae, I.; et al. Trichoderma metabolites as biological control agents against Phytophthora pathogens. Biol. Control 2016, 92, 128–138. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C.; Steinberg, C. Biological control of plant diseases: The European situation. Eur. J. Plant Pathol. 2006, 114, 329–341. [Google Scholar] [CrossRef]
- Kumar, G.; Maharshi, A.; Patel, J.; Mukherjee, A.; Singh, H.B.; Sarma, B.K. Trichoderma: A potential fungal antagonist to control plant diseases. SATSA Mukhapatra 2017, 21, 206–218. [Google Scholar]
- Fitrianingsih, A.; Martanto, E.A.; Abbas, B. The effectiveness of fungi Gliocladium fimbriatum and Trichoderma viride to control Fusarium wilt disease of tomatoes (Lycopersicum esculentum). Indian J. Agric. Res. 2019, 53, 57–61. [Google Scholar]
- Li, Z.B.; Zhou, R.J.; Fu, J.F. Allelopathic effects of phenolic acids from ginseng rhizosphere soil on Cylindrocarpon destructans (Zinss) scholten. Allelopath. J. 2018, 43, 53–64. [Google Scholar] [CrossRef]
- Mwangi, M.W.; Muiru, W.M.; Narla, R.D.; Kimenju, J.W.; Kariuki, G.M. Management of Fusarium oxysporum f. sp. lycopersici and root-knot nematode disease complex in tomato by use of antagonistic fungi, plant resistance and neem. Biocontrol Sci. Technol. 2019, 29, 207–216. [Google Scholar] [CrossRef]
- Gava, C.A.T.; Pinto, J.M. Biocontrol of melon wilt caused by Fusarium oxysporum Schlect f. sp. melonis using seed treatment with Trichoderma spp. and liquid compost. Biol. Control 2016, 97, 13–20. [Google Scholar] [CrossRef]
- Małolepsza, U.; Nawrocka, J.; Szczech, M. Trichoderma virens 106 inoculation stimulates defence enzyme activities and enhances phenolic levels in tomato plants leading to lowered Rhizoctonia solani infection. Biocontrol Sci. Technol. 2017, 27, 180–199. [Google Scholar] [CrossRef]
- Salas-Marina, M.A.; Isordia-Jasso, M.I.; Islas-Osuna, M.A.; Delgado-Sánchez, P.; Jiménez-Bremont, J.F.; Rodríguez-Kessler, M.; Rosales-Saavedra, M.T.; Herrera-Estrella, A.; Casas-Flores, S. The Epl1 and Sm1 proteins from Trichoderma atroviride and Trichoderma virens differentially modulate systemic disease resistance against different life style pathogens in Solanum lycopersicum. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Martinez, C.; Blanc, F.; Le Claire, E.; Besnard, O.; Nicole, M.; Baccou, J.C. Salicylic acid and ethylene pathways are differentially activated in melon cotyledons by active or heat-denatured cellulase from Trichoderma longibrachiatum. Plant Physiol. 2001, 127, 334–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, E.V.; Ulhoa, U.J.; Cardoza, R.E.; Silva, R.N.; Gutiérrez, S. Involvement of Trichoderma harzianum Epl-1 protein in the regulation of Botrytis virulence- and tomato defense-related genes. Front. Plant Sci. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Nawrocka, J.; Szczech, M.; Małolepsza, U. Trichoderma atroviride enhances phenolic synthesis and cucumber protection against Rhizoctonia solani. Plant Prot. Sci. 2018, 54, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhao, H.; Zou, C.; Li, Y.; Chen, Y.; Wang, Z.; Jiang, Y.; Liu, A.; Zhao, P.; Wang, M.; et al. Combined inoculation with multiple arbuscular mycorrhizal fungi improves growth, nutrient uptake and photosynthesis in cucumber seedlings. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Umadevi, P.; Anandaraj, M. Proteomic analysis of the tripartite interaction between black pepper, Trichoderma harzianum and Phytophthora capsici provides insights into induced systemic resistance mediated by Trichoderma spp. Eur. J. Plant Pathol. 2019, 154, 607–620. [Google Scholar] [CrossRef]
- Kipngeno, P.; Losenge, T.; Maina, N.; Kahangi, E.; Juma, P. Efficacy of Bacillus subtilis and Trichoderma asperellum against Pythium aphanidermatum in tomatoes. Biol. Control 2015, 90, 92–95. [Google Scholar] [CrossRef]
- Akköprü, A.; Demir, S. Biological control of Fusarium wilt in tomato caused by Fusarium oxysporum f. sp. lycopersici by AMF Glomus intraradices and some Rhizobacteria. J. Phytopathol. 2005, 153, 544–550. [Google Scholar] [CrossRef]
- Wu, F.; Liu, B.; Zhou, X. Effects of root exudates of watermelon cultivars differing in resistance to Fusarium wilt on the growth and development of Fusarium oxysporum f.sp. niveum. Allelopath. J. 2010, 25, 403–414. [Google Scholar]
- Abdullah, M.T.; Ali, N.Y.; Suleman, P. Biological control of Sclerotinia sclerotiorum (Lib.) de Bary with Trichoderma harzianum and Bacillus amyloliquefaciens. Crop Prot. 2008, 27, 1354–1359. [Google Scholar] [CrossRef]
- Bidellaoui, B.; Segarra, G.; Hakkou, A.; Trillas, M.I. Beneficial effects of Rhizophagus irregularis and Trichoderma asperellum strain T34 on growth and Fusarium wilt in tomato plants. J. Plant Pathol. 2019, 101, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Medina, A.; Pascual, J.A.; Pérez-Alfocea, F.; lbacete, A.; Roldán, A. Trichoderma harzianum and Glomus intraradices modify the hormone disruption in duced by Fusarium oxysporum infection in melon plants. Phytopathology 2010, 100, 682–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbagh, S.K.; Roudini, M.; Panjehkeh, N. Systemic resistance induced by Trichoderma harzianum and Glomus mossea on cucumber damping-off disease caused by Phytophthora melonis. Arch. Phytopathol. Plant Prot. 2007, 50, 375–388. [Google Scholar] [CrossRef]
- Duc, N.H.; Mayer, Z.; Pék, Z.; Helyes, L.; Posta, K. Combined inoculation of arbuscular mycorrhizal fungi, Pseudomonas Fluorescens and Trichoderma spp. For enhancing defense enzymes and yield of three pepper cultivars. Appl. Ecol. Environ. Res. 2017, 15, 1815–1829. [Google Scholar] [CrossRef]
- Azcón-Aguilar, C.; Palenzuela, J.; Roldán, A.; Bautista, S.; Vallejo, R.; Barea, J.M. Analysis of the mycorrhizal potential in the rhizosphere of representative plant species from desertification-threatened Mediterranean shrublands. Appl. Soil Ecol. 2003, 22, 29–37. [Google Scholar] [CrossRef]
- Xavier, L.J.C.; Boyetchko, S.M. Arbuscular mycorrhizal fungi as biostimulants and bioprotectants of crops. Appl. Mycol. Biotechnol. 2002, 2, 311–340. [Google Scholar] [CrossRef]
- Zhou, B.L.; Zheng, J.D.; Bi, X.H.; Cai, L.L.; Guo, W.W. Effects of mycorrhizal fungi on eggplant Verticillium wilt and eggplant growth. Chin. J. Ecol. 2015, 34, 1026–1030. [Google Scholar]
- Panda, S.; Busatto, N.; Hussain, K.; Kamble, A. Piriformospora indica-primed transcriptional reprogramming induces defense response against early blight in tomato. Sci. Hortic. 2019, 255, 209–219. [Google Scholar] [CrossRef]
- Gao, P.; Qin, J.; Li, D.; Zhou, S. Inhibitory effect and possible mechanism of a Pseudomonas strain QBA5 against gray mold on tomato leaves and fruits caused by Botrytis cinerea. PLoS ONE 2018, 10, e0190932. [Google Scholar] [CrossRef] [Green Version]
- Patakioutas, G.; Dimou, D.; Yfanti, P.; Karras, G.; Ntatsi, G.; Savvas, D. Root inoculation with beneficial micro-organisms as a means to control Fusarium oxysporum f. sp. lycopersici in two Greek landraces of tomato grown on perlite. Acta Hortic. 2017, 1168, 277–286. [Google Scholar] [CrossRef]
- Zheng, N.; Feng, K.; Jingyan, J.; Xuefei, W.; Li, C.W.; Jiahui, L.; Yu, Z.; Zi, Y.; Bin, Z. Growth improvement and salt tolerance mechanisms of tomato seedlings mediated by plant growth-promoting rhizobacteria from contaminated soils. Chin. J. Appl. Environ. Biol. 2018, 24, 47–52. [Google Scholar]
- Lareen, A.; Burton, F.; Schäfer, P. Plant root-microbe communication in shaping root microbiomes. Plant Mol. Biol. 2016, 90, 575–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, G.; Opelt, K.; Zachow, C.; Lottmann, J.; Gotz, M.; Costa, R.; Smalla, K. The rhizosphere effect on bacteria antagonistic towards the pathogenic fungus Verticillium differs depending on plant species and site. FEMS Microbiol. Ecol. 2006, 56, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Miethling, R.; Wieland, G.; Backhaus, H.; Tebbe, C.C. Variation of microbial rhizosphere communities in response to crop species, soil origin, and inoculation with Sinorhizobium meliloti L33. Microb. Ecol. 2000, 40, 43–56. [Google Scholar] [CrossRef]
- Faure, D.; Dessaux, Y. Quorum sensing as a target for developing control strategies for the plant pathogen Pectobacterium. In New Perspectives and Approaches in Plant Growth-Promoting Rhizobacteria Research; Bakker, P.A.H.M., Raaijmakers, J.M., Bloemberg, G., Höfte, M., Lemanceau, P., Cooke, B.M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 353–365. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, S.; Chai, Y.; Clardy, J.; Kolter, R.; Guo, J.H.; Losick, R. A Bacillus subtilis sensor kinase involved in triggering biofilm formation on the roots of tomato plants. Mol. Microbiol. 2012, 85, 418–430. [Google Scholar] [CrossRef]
- Tan, S.; Yang, C.; Mei, X.; Shen, S.; Raza, W.; Shen, Q.; Xu, Y. The effect of organic acids from tomato root exudates on rhizosphere colonization of Bacillus amyloliquefaciens T-5. Appl. Soil Ecol. 2013, 64, 15–22. [Google Scholar] [CrossRef]
- Ling, N.; Huang, Q.; Guo, S.; Shen, Q. Paenibacillus polymyxa SQR-21systemically affects root exudates of watermelon to decrease the conidial germination of Fusarium oxysporum f.sp. niveum. Plant Soil 2011, 341, 485–493. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, Z.; Yang, X.; Ran, W.; Shen, Q. Trichoderma harzianum T-E5 significantly affects cucumber root exudates and fungal community in the cucumber rhizosphere. Appl. Soil Ecol. 2013, 72, 41–48. [Google Scholar] [CrossRef]
- Zhou, X.G.; Wang, J.; Jin, X.; Li, D.L.; Shi, Y.J.; Wu, F.Z. Effects of selected cucumber root exudates components on soil Trichoderma spp. communities. Allelopath. J. 2019, 47, 257–265. [Google Scholar] [CrossRef]
- Pozo, M.J.; Lopez-Raez, J.A.; Azcon-Aguilar, C.; Garcia-Garrido, J.M. Phytohormones as integrators of environmental signals in the regulation of mycorrhizal symbioses. New Phytol. 2015, 205, 1431–1436. [Google Scholar] [CrossRef]
- Muller, L.M.; Harrison, M.J. Phytohormones, miRNAs, and peptide signals integrate plant phosphorus status with arbuscular mycorrhizal symbiosis. Curr. Opin. Plant Biol. 2019, 50, 132–139. [Google Scholar] [CrossRef] [PubMed]
- López-Ráez, J.A.; Shirasu, K.; Foo, E. Strigolactones in plant interactions with beneficial and detrimental organisms. The Yin and Yang. Trends Plant Sci. 2017, 22, 527–537. [Google Scholar] [CrossRef]
- Hause, B. Jasmonates in arbuscular mycorrhizal interactions. Phytochemistry 2007, 68, 101–110. [Google Scholar] [CrossRef]
- Foo, E.; Ross, J.J.; Jones, W.T.; Reid, J.B. Plant hormones in arbuscular mycorrhizal symbioses: An emerging role for gibberellins. Ann. Bot. 2013, 111, 769–779. [Google Scholar] [CrossRef]
- De Los Santos, R.T.; Vierheilig, H.; Ocampo, J.A.; Garrido, J.M.G. Altered pattern of arbuscular mycorrhizal formation in tomato ethylene mutants. Plant Signal. Behav. 2011, 6, 755–758. [Google Scholar] [CrossRef] [Green Version]
- Etemadi, M.; Gutjahr, C.; Couzigou, J.M.; Zouine, M.; Lauressergues, D.; Timmers, A.; Audran, C.; Bouzayen, M.; Becard, G.; Combier, J.P. Auxin perception is required for arbuscule development in arbuscular mycorrhizal symbiosis. Plant Physiol. 2014, 166, 281–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillotin, B.; Etemadi, M.; Audran, C.; Bouzayen, M.; Becard, G.; Combier, J.P. Sl-IAA27 regulates strigolactone biosynthesis and mycorrhization in tomato (var. MicroTom). New Phytol. 2017, 213, 1124–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Medina, A.; Pescador, L.; Fernandez, I.; Rodrıguez-Serrano, M.; Garcıa, J.M.; Romero-Puertas, M.C.; Pozo, M.J. Nitric oxide and phytoglobin PHYTOGB1 are regulatory elements in the Solanum lycopersicum–Rhizophagus irregularis mycorrhizal symbiosis. New Phytol. 2019, 223, 1560–1574. [Google Scholar] [CrossRef] [PubMed]
- King, S.R.; Davis, A.R.; Liu, W.; Levi, A. Grafting for disease resistance. HortScience 2008, 43, 1673–1676. [Google Scholar] [CrossRef] [Green Version]
- Yetışır, H.; Sari, N.; Yücel, S. Rootstock resistance to Fusarium wilt and effect on watermelon fruit yield and quality. Phytoparasitica 2003, 31, 163. [Google Scholar] [CrossRef]
- Grimault, V.; Prior, P. Grafting tomato cultivars resistant or susceptible to bacterial wilt: Analysis of resistance mechanisms. J. Phytopathol. 1994, 141, 330–334. [Google Scholar] [CrossRef]
- Albert, R.; Künstler, A.; Lantos, F.; Ádám, A.L.; Király, L. Graft-transmissible resistance of cherry pepper (Capsicum annuum var. cerasiforme) to powdery mildew (Leveillula taurica) is associated with elevated superoxide accumulation, NADPH oxidase activity and pathogenesis-related gene expression. Acta Physiol. Plant 2017, 39, 53. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Johnson, S.; Inglis, D.; Miles, C. Grafting effects on eggplant growth, yield, and Verticillium wilt incidence. Int. J. Veg. Sci. 2014, 20, 3–20. [Google Scholar] [CrossRef]
- Liu, N.; Zhou, B.; Zhao, X.; Lu, B.; Li, Y.; Hao, J. Grafting eggplant onto tomato rootstock to suppress Verticillium dahliae infection: The effect of root exudates. HortScience 2009, 44, 2058–2062. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Zhu, C.; Raza, W.; Wang, D.; Huang, Q.; Guo, S.; Ling, N.; Shen, Q. Coupling of the chemical niche and microbiome in the rhizosphere: Implications from watermelon grafting. Front. Agric. Sci. Eng. 2016, 3, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Cichy, K.A.; Snapp, S.S.; Kirk, W.W. Fusarium root rot incidence and root system architecture in grafted common bean lines. Plant Soil 2007, 300, 233–244. [Google Scholar] [CrossRef]
- Chitarra, W.; Perrone, I.; Avanzato, C.G.; Minio, A.; Boccacci, P.; Santini, D.; Gilardi, G.; Siciliano, I.; Gullino, M.L.; Delledonne, M.; et al. Grapevine grafting: Scion transcript profiling and defense-related metabolites induced by rootstocks. Front. Plant Sci. 2017, 8, 654. [Google Scholar] [CrossRef]
- Wallis, C.M.; Wallingford, A.K.; Chen, J. Grapevine rootstock effects on scion sap phenolic levels, resistance to Xylella fastidiosa infection, and progression of Pierce’s disease. Front. Plant Sci. 2013, 4, 502. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, T.; Itagaki, K.; Wang, Y.; Endo, R. Grafting transiently suppresses development of powdery mildew colonies, probably through a quantitative change in water relations of the host cucumber scions during graft healing. Sci. Hortic. 2015, 192, 197–199. [Google Scholar] [CrossRef]
- Song, Y.; Ling, N.; Ma, J.; Wang, J.; Zhu, C.; Raza, W.; Shen, Y.; Huang, Q.; Shen, Q. Grafting resulted in a distinct proteomic profile of watermelon root exudates relative to the un-grafted watermelon and the rootstock plant. J. Plant Growth Regul. 2016, 35, 778–791. [Google Scholar] [CrossRef]
- Ling, N.; Zhang, W.; Wang, D.; Mao, J.; Huang, Q.; Guo, S.; Shen, Q. Root exudates from grafted-root watermelon showed a certain contribution in inhibiting Fusarium oxysporum f. sp. niveum. PLoS ONE 2013, 8, e63383. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, S.; Zhao, J.; Zhou, X.; Wang, B.; Li, Y.; Zheng, G.; Zhang, J.; Cai, Z.; Huang, X. Watermelon planting is capable to restructure the soil microbiome that regulated by reductive soil disinfestation. Appl. Soil Ecol. 2018, 129, 52–60. [Google Scholar] [CrossRef]
- Yin, Y.; Luo, S.; Li, Y.; Zhou, J.; Tang, Y.; Liu, Y. Grafting shaping the microbial community structure to suppress Verticillium dahliae in the rhizosphere of eggplants. Am. J. Agric. For. 2018, 6, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Jaramillo, J.E.; Mendes, R.; Raaijmakers, J.M. Impact of plant domestication on rhizosphere microbiome assembly and functions. Plant Mol. Biol. 2016, 90, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Jousset, A. Plant breeding goes Microbial. Trends Plant Sci. 2017, 22, 555–558. [Google Scholar] [CrossRef]
- Marasco, R.; Rolli, E.; Fusi, M.; Michoud, G.; Daffonchio, D. Grapevine rootstocks shape underground bacterial microbiome and networking but not potential functionality. Microbiome 2018, 6, 3. [Google Scholar] [CrossRef]
- Nicola, L.; Insam, H.; Pertot, I.; Stres, B. Reanalysis of microbiomes in soils affected by apple replant disease (ARD): Old foes and novel suspects lead to the proposal of extended model of disease development. Appl. Soil Ecol. 2018, 129, 24–33. [Google Scholar] [CrossRef]


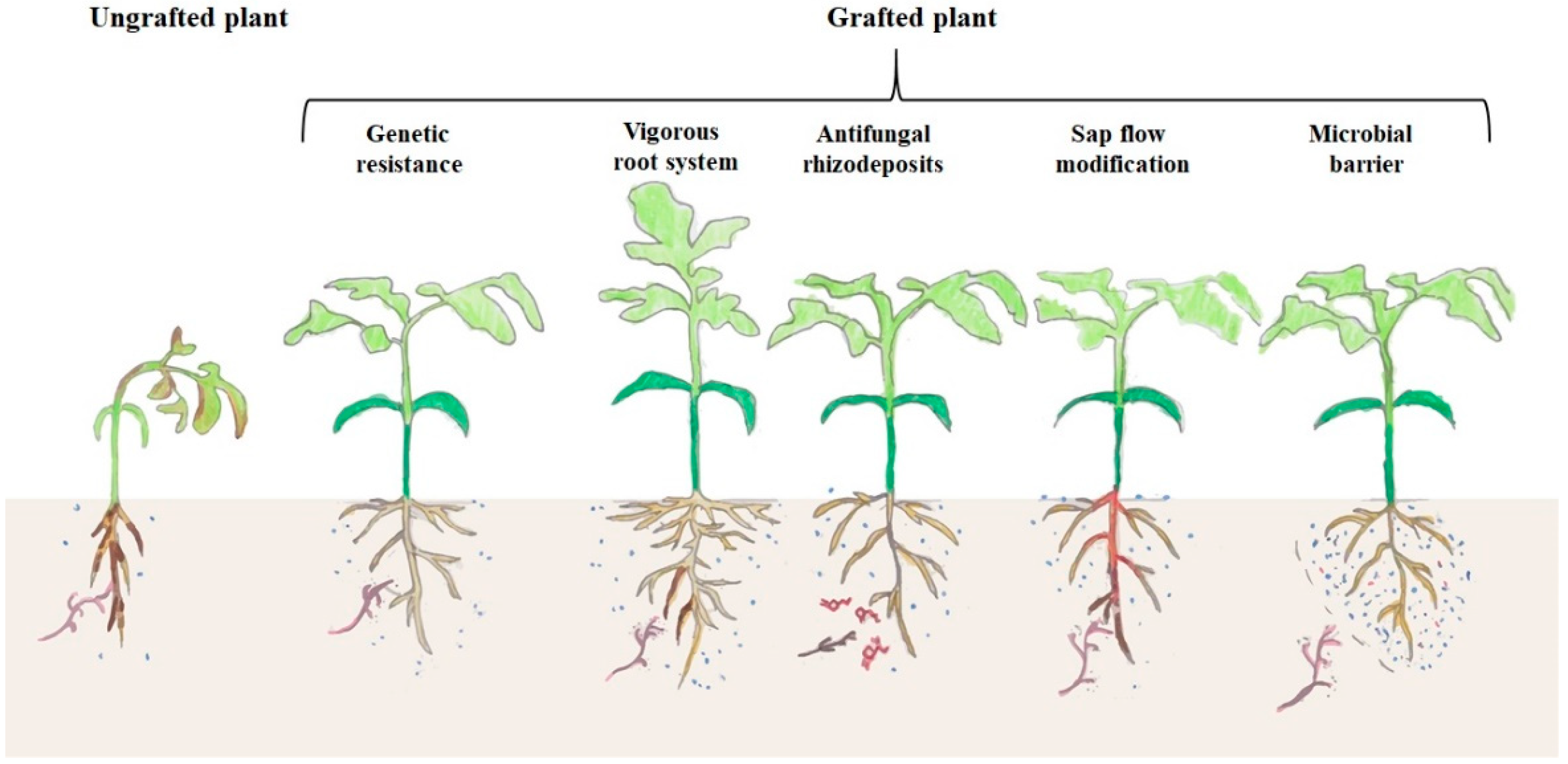{kind=link}
| To | Grafted Roots | Microbiota |
|---|---|---|
| Grafted roots | Allelopathy Competition for space, water and nutrients | Selection-filtering effects C-food providing Activation of the endophytism |
| Microbiota | Nutrients availability Growth promotion Induction of resistance to (a)biotic stresses | Antibiosis Hyperparassitism Niche occupying Essential food depletion |
| Management Goal | Ungrafted Plants | Grafted Plants |
|---|---|---|
| Reduction of disease risks | Cultivation of susceptible cultivars on healthy soils | Grafting of susceptible cultivars on disease resistant rootstocks |
| Genetic innovation regarding plant protection objectives | New cultivars resistant/tolerant to pathogens | New rootstock genotypes resistant/tolerant to pathogens New grafting combinations with enhanced root ability to recruit protective rhizobiome |
| Engineering soil microbiome for enhancing its antagonistic structure | Soil conditioning techniques mitigating soil sickness phenomena | Actions for maintenance of ecological bio-barriers proliferating on the rootstock roots |
| Increased effectiveness of the chemical control means | Wide-spectrum preventive and curative applications of fungicides | Targeted use of active molecules both in time and in dosage |
| Increased effectiveness of the biological control agents | Mass applications of the bio-based formulates | Coordinating application of the external microbial antagonists, with the naturally present rhizobiome Targeted use of specific microbial strains characterized by grafted-rhizosphere competence |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardarelli, M.; Rouphael, Y.; Kyriacou, M.C.; Colla, G.; Pane, C. Augmenting the Sustainability of Vegetable Cropping Systems by Configuring Rootstock-Dependent Rhizomicrobiomes that Support Plant Protection. Agronomy 2020, 10, 1185. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081185
Cardarelli M, Rouphael Y, Kyriacou MC, Colla G, Pane C. Augmenting the Sustainability of Vegetable Cropping Systems by Configuring Rootstock-Dependent Rhizomicrobiomes that Support Plant Protection. Agronomy. 2020; 10(8):1185. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081185
Chicago/Turabian StyleCardarelli, Mariateresa, Youssef Rouphael, Marios C. Kyriacou, Giuseppe Colla, and Catello Pane. 2020. "Augmenting the Sustainability of Vegetable Cropping Systems by Configuring Rootstock-Dependent Rhizomicrobiomes that Support Plant Protection" Agronomy 10, no. 8: 1185. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10081185