Effect of Crop Residues on Weed Emergence




1
Department of Agronomy, Food, Natural Resources, Animals and Environment (DAFNAE), University of Padova, Viale dell’ Università 16, 35020 Legnaro, Italy
2
Institute for Sustainable Plant Protection, National Research Council of Italy, Viale dell’ Università 16, 35020 Legnaro, Italy
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(1), 163; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010163
Submission received: 11 December 2020
/
Revised: 9 January 2021
/
Accepted: 14 January 2021
/
Published: 16 January 2021
(This article belongs to the Special Issue Expanding the Knowledge on Weed Germination and Early Growth to Design Sustainable Management Strategies)
Abstract
:Weed behaviour in crop fields has been extensively studied; nevertheless, limited knowledge is available for particular cropping systems, such as no-till systems. Improving weed management under no-till conditions requires an understanding of the interaction between crop residues and the seedling emergence process. This study aimed to evaluate the influence of maize and wheat residues, applied in three different quantities (1, the field quantity, 0.5, and 1.5-fold amounts of the field quantity), on the emergence of eight weed species: Abutilon theophrasti, Amaranthus retroflexus, Chenopodium album, Digitaria sanguinalis, Echinochloa crus-galli, Setaria pumila, Sonchus oleraceus, and Sorghum halepense. The experiment was conducted over two consecutive years. The results showed that the quantities 1 and 1.5 could suppress seedling emergence by 20 and 44%, respectively, while the quantity 0.5 seems to promote emergence by 22% compared with the control without residues. Weed species showed different responses to crop residues, from C. album showing 56% less emergence to S. halepense showing a 44% higher emergence than the control without residues. Different meteorological conditions in the two-year experiment also exhibited a significant influence on weed species emergence.
1. Introduction
Weeds, being superior competitors than crops in contention for available resources, have become one of the major agricultural problems worldwide [1]. Different cropping systems experience this problem differently, depending on the soil characteristics from where weed seeds germinate and the obstacles they face in the early growth, such as crop residues present as post-harvest crop residues or used as mulches [2,3]. Crop residues are materials left on cultivated land after the crop has been harvested. Besides being used to suppress weed emergence, crop residues, coupled with good agricultural management practices, can also have many positive impacts on soil quality. They are considered an effective anti-erosion measure; they can improve soil structure, increase organic content in the soil, reduce evaporation, and help fix carbon dioxide (CO2). Therefore, they can also be an asset for seed germination [4]. Crop residues can also decrease evapotranspiration, reduce the soil temperature, and retain soil moisture, which can also be beneficial for germination and growth of both crops and weeds [5]. Seed germination is the passage from a dormant to a metabolically active embryo, accompanied by the use of natural resources, such as oxygen and water, and the release of CO2. Therefore, seeds need a favourable environment for germinating and emerging [6]. The fate of the seed may differ depending on the species and the dissemination period. Some may germinate immediately after dissemination, and others may have to pass a certain period of dormancy before germinating, caused by seed physical or physiological effects [7]. Seed germination and emergence are mainly influenced by four factors: Temperature, water and oxygen quantity and availability, and light. The effects of all these factors, even though essential, are species-specific, meaning that not all species have the same requirements. Some may germinate better with less light, some with depressed temperatures, and others only with high levels of humidity. Therefore, the local differences in these environmental factors may have a meaningful impact on seed germination and emergence [6,7]. Some crop residues may obstruct or prevent water infiltration, and may therefore, be detrimental to seed germination; the thickness of crop residues has been proposed as a possible cause for this phenomenon [8,9]. Crop residues can also inhibit light transmittance to the seeds underneath, which may also inhibit germination [10]. These findings confirm that crop residues can act as an obstacle for water and light penetration and can reduce temperature fluctuations, which can lead to a deceleration, reduction, or a complete absence of seed germination and plant emergence [5,11]. The effect of crop residues on weed plant emergence has been exploited to control weeds in almost all agricultural practices, using different mulches. Permanent soil cover is one of the three main pillars upon which conservation agriculture (CA) is built [12,13]. The three characteristics of CA are minimum soil disturbance, crop rotation, and maintenance of soil cover by crop residues and living mulch [13]. The influence of crop residues on seed germination and emergence is significant when no-till practices are applied because the absence of mechanical soil disturbance means seeds always remain on the soil surface in a micro-environment created by crop residues [11,14,15]. Even though positive effects on the reduction of weed emergence by covering the soil with crop residues have been demonstrated [16,17], some questions remain unanswered. Some of the most important questions include single weed species’ response to these measures and what effects different quantities of residues might have. The uniformity of residue distribution may also be critical, regardless of the amount used.
Some studies concerning the effect of different residue quantities on weed species emergence have been conducted with different crops and cover crop residues [18]. Nevertheless, there is a lack of information on the weed species-specific emergence response to different types and amounts of residues. This kind of knowledge could be useful for understanding the residue required to effectively reduce specific weed infestations.
This work tackles some of these issues to fill the gap and contribute more to the knowledge of the weed emergence processes under crop residues. The trials were conducted in Northern Italy by using maize and wheat residues, therefore representing the two major cereal crops grown in Europe [19], and eight weed species commonly found in the fields of Northern Italy.
2. Materials and Methods
The trials were conducted at the ‘Lucio Toniolo’ experimental farm of the University of Padova, located in Legnaro in the Veneto region, Northern Italy. For the trials, two types of crop residues were selected: Maize (Zea mays) and wheat (Triticum aestivum). In order to assess the quantities that are typically left in the field after harvest, maize and wheat residues from 1 m2 plots were collected where average yields were obtained and their dry matter was determined. The established amount was then set as quantity 1 and was used as a reference value for determining other quantities to be used. In order to simulate the amount of residues left by crops with increasing levels of productivity (low, average, high), three different quantities were used: 0.5, 1, and 1.5-fold of the amount collected per square meter. The weights of the three quantities for each crop residue are shown in Table 1.
The experiment was laid out in a factorial design with eight weed species, two crop residues, three residue quantities, and three blocks, with one plot without residues for each species and for each block, marked as 0. The seeds of each species were sown in a square area (plot) of 20 cm × 20 cm.
The eight weed species selected are commonly found in the fields of Northern Italy: Abutilon theophrasti Medik. (ABUTH), Amaranthus retroflexus L. (AMARE), Chenopodium album L. (CHEAL), Digitaria sanguinalis (L.) Scop. (DIGSA), Echinochloa crus-galli (L.) Beauv. (ECHCG), Setaria pumila (Poir.) Roem. & Schult. (SETPU), Sonchus oleraceus L. (SONOL), and Sorghum halepense (L.) Pers. (SORHA). Each species was sown in each block for each residue quantity used plus one control replicate (without residues), for both types of crop residue used. For each species, 200 seeds were sown per plot. Seeds were hand-harvested in summer 2018 from natural populations at the experimental farm. After being collected, the seeds were cleaned and dry stored at room temperature until used in the experiment.
Before sowing, the first 5 cm of soil were removed from the rows and heat sterilised at 105 °C for 48 h in a stove. Soil sterilisation was conducted to eliminate the seeds present in the natural seed bank. After sterilisation, the soil was redistributed in the rows. The seeds of each of the eight species were then sown in their respective plots on the soil and were not uniformly distributed, imitating the natural dispersal of seeds from the mother plant. After the sowing operations, the plots were covered with different residue types and quantities or left uncovered (control) according to the experimental plan. The appearance of the experimental site during and after the previously described operations is shown in Figure 1.
The seeds were sown in winter, creating conditions for vernalisation and dormancy release. The experiment was repeated for two consecutive years, following the same experimental design; the first experiment started on the 18 December 2018 and the second one on the 20 December 2019. After the beginning of seedling emergence, mid-March for the first experiment and the beginning of March for the second, emergence was monitored every two to three days. The emerged seedlings were removed and counted, and the number was recorded. The first experiment was concluded on the 26 June 2019, and the second on the 1 July 2020, following a period without emergence. For the same period, data concerning temperature and rainfall were recorded by the meteorological station at the Regional Agency for Environmental Protection (ARPA), located at the experimental farm.
Statistical Analysis
The total number of emerged seedlings was calculated for each replicate, and means ± standard error (SE) were determined for each treatment. Total emergence was calculated as the mean of replicates with residues and normalized with the means of control replicates without residues. Finally, the total emergence (expressed as a percentage of the total emergence observed in the control without residues), after verifying homoscedasticity and normality, was statistically analysed by a factorial ANOVA. Year (Y, two levels), Species (S, eight levels), Crop residue type (Cr, two levels), and residue quantities (Rq, three levels) were considered as experimental factors. Significance of differences between means was evaluated using Tukey’s HSD test (p < 0.05).
3. Results
The total number of emerged seedlings in the control plots (no residues) varied between species and years, with the highest percentage of emergence observed for E. crus-galli and S. pumila and the lowest for C. album and A. retroflexus. Data on emergence percentage for the different species in the different treatments are reported in Appendix A (Figure A1, Figure A2, Figure A3, Figure A4, Figure A5, Figure A6, Figure A7 and Figure A8). The results of the ANOVA on total emergence data showed that only the residue quantity (Rq) and weed species (S) were significant as factors as well as the interaction between the two years and weed species (Y × S) (Table 2). The effects of these two factors and the year × species interaction are explained in detail hereafter.
3.1. Quantity of Residues
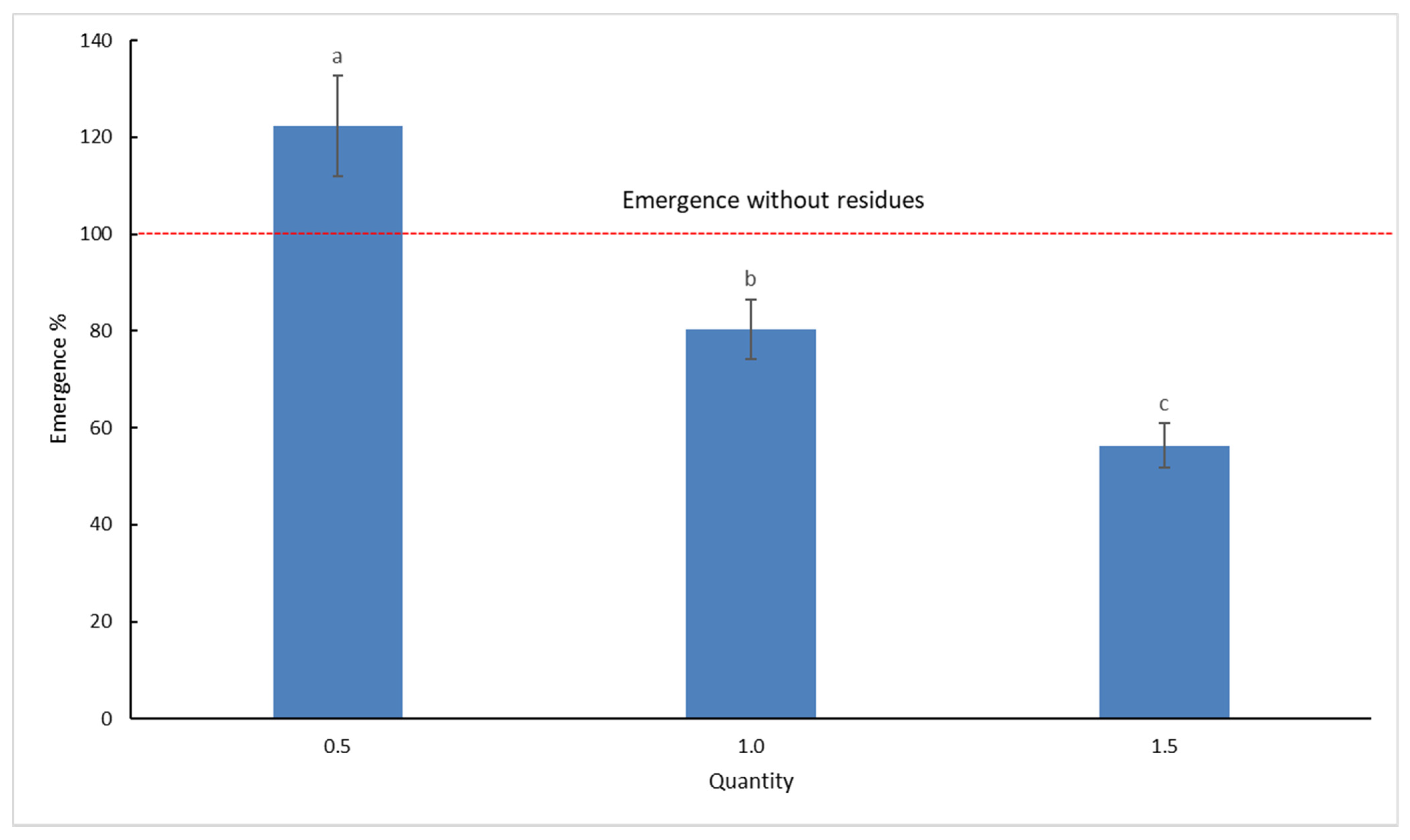
The effect of the quantity of crop residues on weed emergence is presented as percentages, relative to that of the control (Figure 2). Values above 100% indicate that under those residues, emergence was greater than without residues, while values below 100% indicate that the emergence was lower than in the control.
It is interesting to note that under quantity 0.5, weeds emerged better than they did without residues; in fact, the overall emergence was 22% higher than the control without residues. With quantity 1, the field quantity, the overall emergence was 20% lower than the emergence without residues. The quantity 1.5 produced the highest overall reduction (44%) of seedling emergence.
3.2. Weed Species
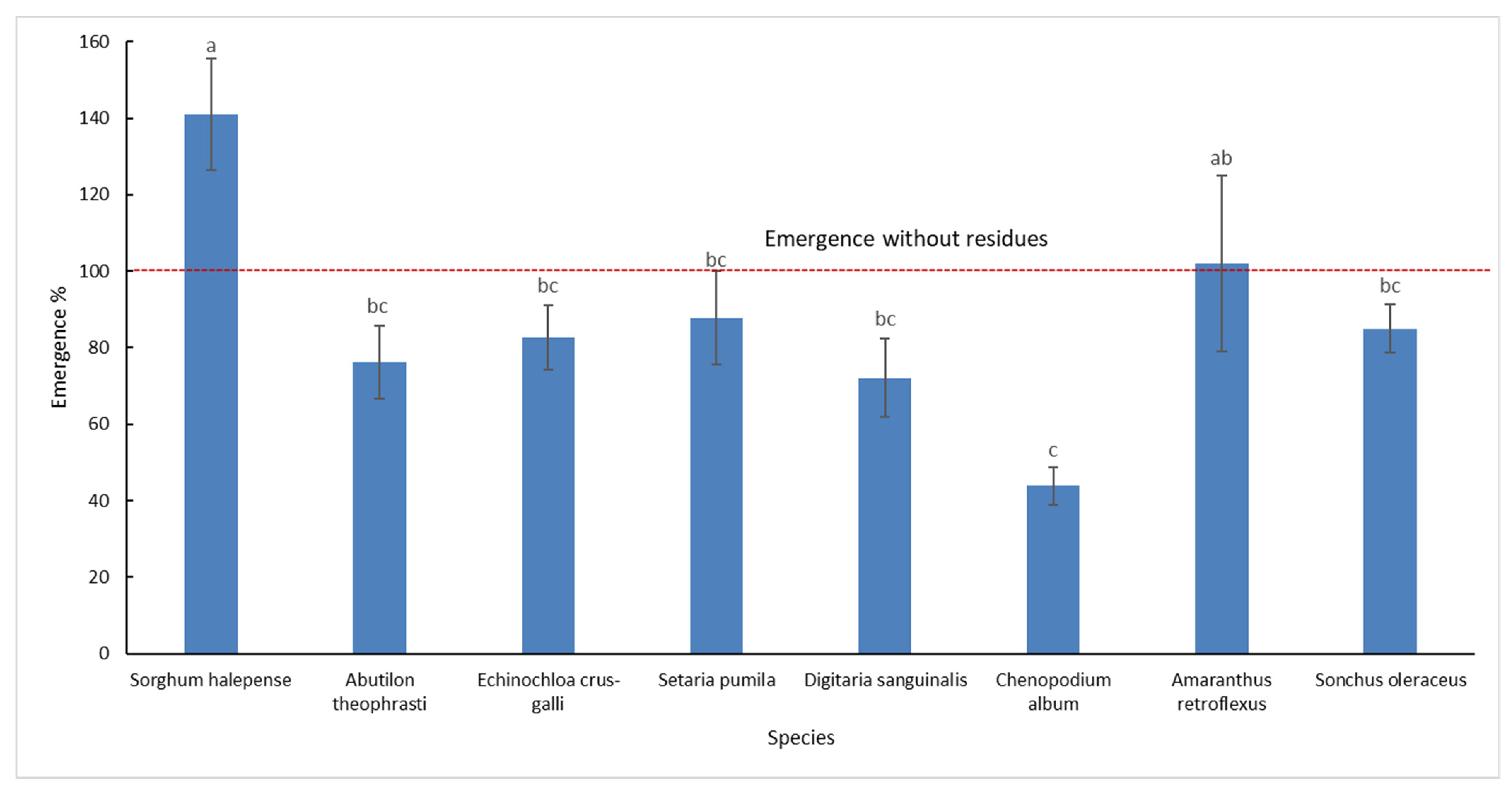
Emergence data for each of the eight species studied were analysed separately. As for the residue quantities, the emergence percentages were calculated based on the emergence of control plots-without residues. The results are shown in Figure 3.
The most interesting result was the emergence percentage of S. halepense which was 41% more with residues than without residues. A. retroflexus had approximately 2% more emergence in the presence of residues. All other species had an emergence percentage that was 12 to 56% lower with residues than without residues. The lowest emergence percentage observed was 44% which was exhibited by C. album.
In summary, the results indicated that the presence of crop residues favoured S. halepense, A. retroflexus emergence was not affected by residues, while for S. pumila, S. oleraceus, E. crus-galli, A. theophrasti, and D. sanguinalis, the presence of crop residues negatively affected emergence. C. album was subject to the most severe inhibition effect of the presence of crop residues on seedling emergence.
3.3. Year-Weed Species Interaction
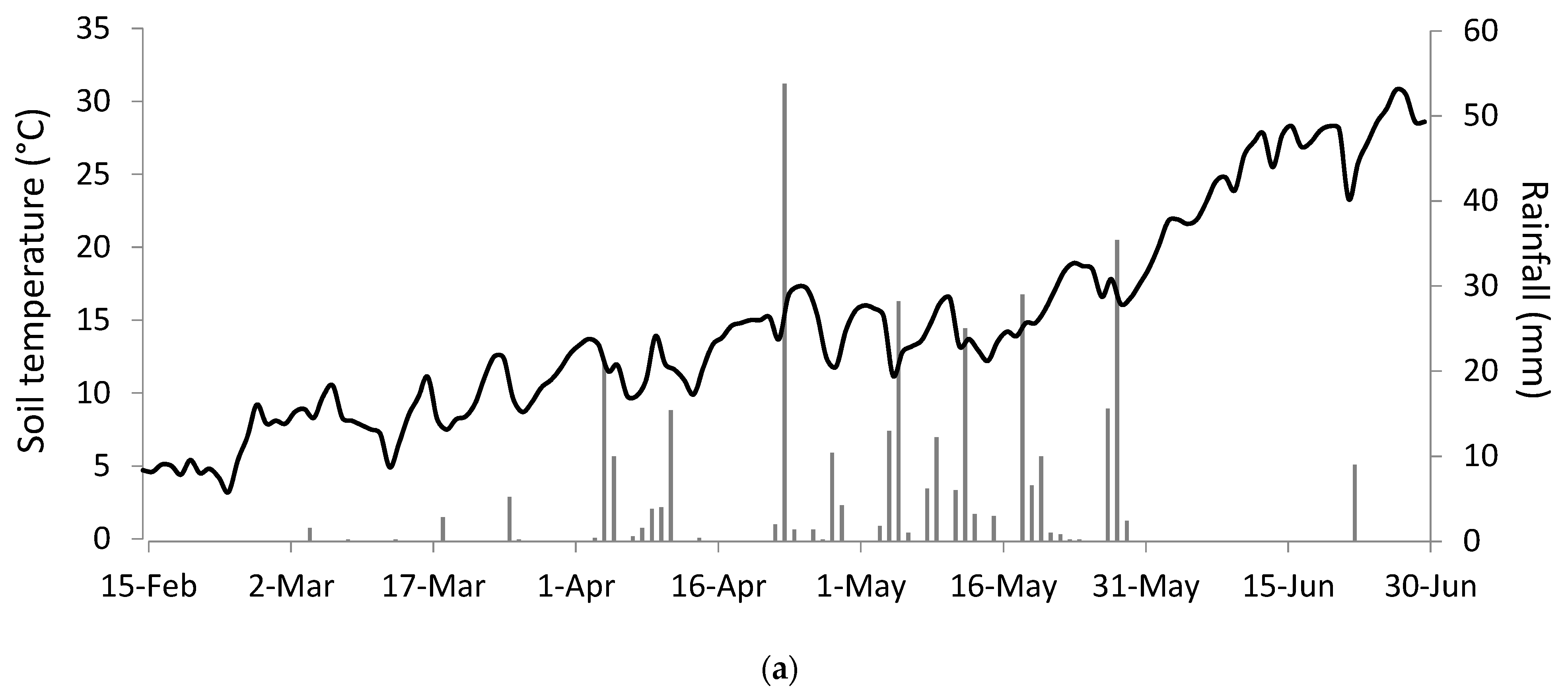
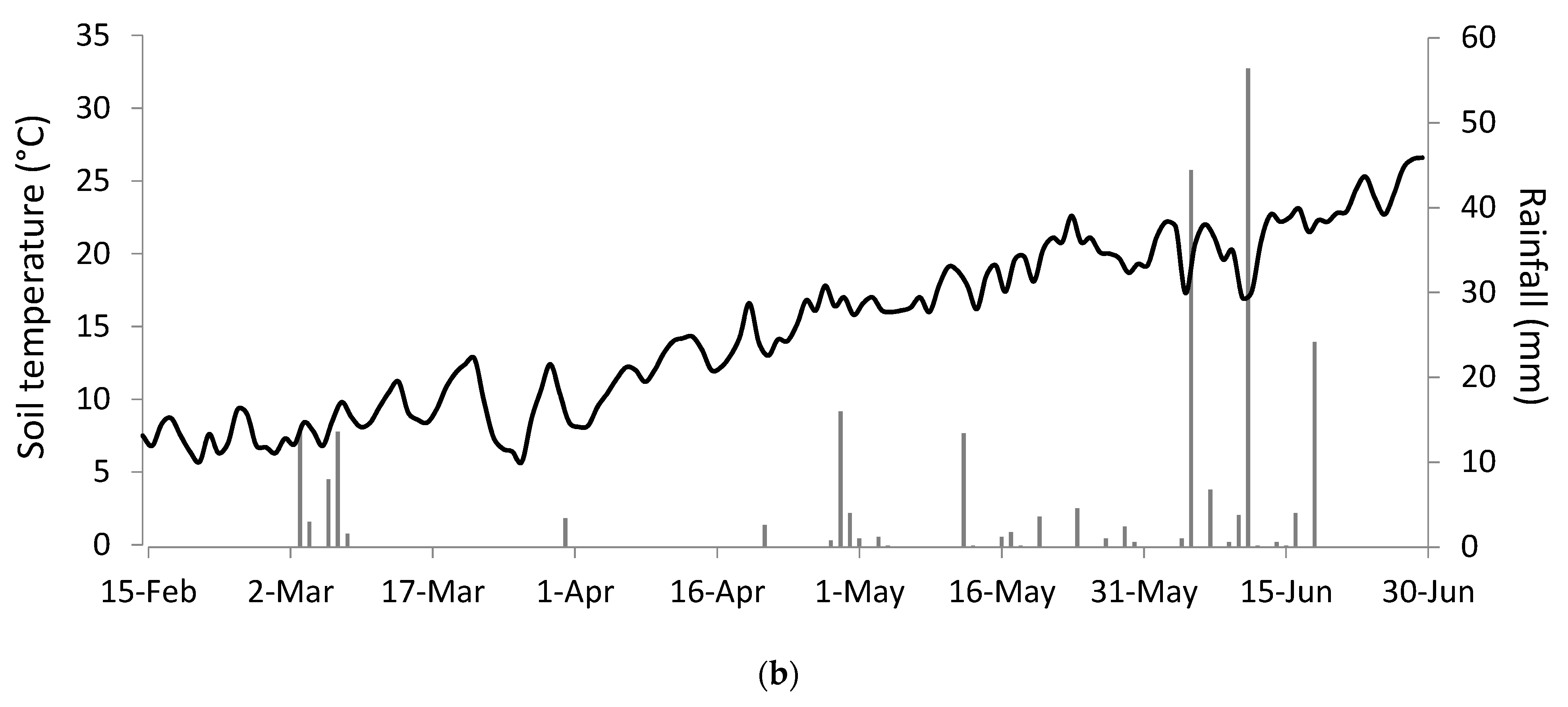
The same species showed differences in seedling emergence between the two years of study, likely resulting from the different meteorological conditions, as illustrated in Figure 4.
In Figure 4, it can be observed that during the emergence period (from May to July), soil temperature and precipitation differed between years. The first year (2019) was characterised by a moderate and constant increase in temperatures in spring until the beginning of June when the temperature rose abruptly. The year was also characterised by low early spring precipitation, with most of the rainfall occurring during late spring. On the other hand, the second year (2020) suffered an extended dry period during most of spring with an important amount of rainfall during the late spring/early summer period. The temperature increased progressively from spring to summer.
Differences in weed species emergence between the two years are shown in Figure 5.
It is interesting to observe that some species (S. halepense, A. retroflexus, and S. oleraceous) emerged by approximately the same amount in both years. Other species, such as A. theophrasti and E. crus-galli, showed an emergence favoured by or indifferent to the presence of crop residues in 2019, respectively, and reduced by the presence of residues in 2020. On the contrary, the emergence of S. pumila and D. sanguinalis was negatively influenced by residues only in 2019.
4. Discussion
Residues covering seeds are known to prevent their germination by obstructing light penetration, decreasing soil temperature, affecting soil moisture, and reducing oxygen availability. Our results demonstrated significant suppression of seedling emergence with a higher amount of residues, but with fewer residues, there was a favourable effect on emergence for some species. This finding could be explained considering that with a low amount of residues, light and oxygen reach the seeds, the soil temperature is little affected, and residues can retain moisture which is then available for the seed germination process [11,20]. The results of our study agree with the findings of Liebman et al. and Ranaivoson et al., [21,22], who reported that low residue quantities stimulate weed emergence.
With high quantities of residues, weed emergence reduction is quite visible. Similar results were reported by Chauhan and colleagues [18]. In their study, rice (Oryze sativa) residues were used in two different quantities (0.3 kg/m2 and 0.6 kg/m2), similar to the wheat quantities used in our experiment. The authors found that both quantities negatively influenced weed emergence, which decreased with an increase in residue quantities, thus confirming the existence of a general trend of weed emergence suppression with an increase in crop residue quantities. However, it should be highlighted that Chauhan et al. [18] experiments used different types of residues and different weed species than our study, which could also influence the emergence.
There are different studies that argue that seed mass determines species emerging capacity under residues. Larger seeds have a higher germination rate than smaller seeds [23,24,25]. This is because larger seeds have more nutrient reserves than smaller seeds and can therefore, overcome obstacles for their germination and emergence more easily, giving young plants more time to adapt to the conditions of the environment [26,27,28]. Moreover, other studies indicated that large seeds need less light to germinate [29]. Furthermore, imbibition of large seeds can be difficult when they are on the soil surface because of poor seed–soil contact; the presence of crop residues might increase moisture on the soil surface and facilitate seed imbibition. This general rule could justify S. halepense behaviour (e.g., increase in emergence) as the presence of the residues favoured it, but in our study, this was not observed for all the other species with large seeds (e.g., A. theophrasti). Moreover, although C. album emergence, a species with small seeds, was inhibited by residue cover, A. retroflexus emerged even under residues. It seems clear in our findings that species do not follow the supposed rule on the positive relationship between seed size and emergence under residues. However, there are other authors who found that some of the small seed-size species, such as Sonchus asper (L.) Hill, Taraxacum officinale F. H. Wigg, Alopecurus myosuroides Huds., Veronica persica Poir., Plantago lanceolata L., and Portulaca oleracea L., could emerge under residues [30,31]. They proposed that these species have lower requirements for light and water to start germinating, compared to larger seeds. Moreover, given their thin seedling form and emergence speed, they are able to find a way and penetrate above the residue cover before exhausting nutrient reserves.
Since temperature and soil moisture are the most important factors for seed germination and seedling emergence [32], the absence of precipitation and too high or low temperatures may decrease germination, while rainfall during spring promotes it [33,34,35]. Therefore, weather conditions undoubtedly played a role in the observed differences in weed species total emergence between the two years of these experiments, and the emergence patterns of the species were different between 2019 and 2020 (emergence patterns of all eight species are in Appendix A). Seedling emergence was lower in 2020 than in 2019 due to drier conditions, with the exception of S. oleraceus. Although total emergence was lower in 2020, some species sensitive to drought, such as D. sanguinalis and S. pumila [36,37,38] in 2020 showed greater emergence under the residues compared to control, probably because the residues were able to retain some of the moisture in the upper soil layer. This level of moisture was probably enough to allow seed imbibition and germination for S. oleraceus, S. pumila, and D. sanguinalis but not for the remaining species. This could be related to inter-specific differences in base water potential for germination or in seed traits, such as size, coat or shape, involved in imbibition process.
In general, it was observed that the presence of crop residues tended to delay weed emergence, with a few exceptions for some species in some years. These exceptions include the small seeded species such as S. oleraceus that showed no delay in emergence in both years, and C. album that showed no delay in emergence in 2019, and a slight delay in 2020. In 2019, emergence anticipation of S. halepense was observed, going from two weeks, with the smallest amount, to several days, with the highest number of residues, compared to control. However, this was not the case in 2020. Another interesting result was the emergence pattern of D. sanguinalis in 2020 where the amounts 0.5 and 1 of both residue types anticipated the emergence for more than a month, compared to control. Yet the quantity 1.5 of both residues delayed the emergence for almost two weeks. A. theophrasti emergence was delayed in both years from one to several weeks with the increasing amount of residues, and similar results were observed in the emergence patterns of E. crus-galli with emergence delay of few weeks up to a month, with the increasing amount of residues. These results are probably the product of the interaction between the specific microclimatic conditions created by the residues and the biology of these species that have different requirements of light, moisture, and temperature for dormancy breaking and germination [38,39,40,41,42,43]. However, we can comment on these results only hypothetically, considering that no measurements of microclimatic conditions underneath the residues were conducted, as it was not the goal of this work. These results are interesting because weeds emerging later are less competitive against crops and cause less yield loss. Moreover, these weeds could produce a lower amount of seeds or immature seeds; therefore, they could be less problematic in the future. In any case, the findings underline that, depending on the weather conditions, crop residue quantity, and weed species, the effects of crop residues on weed emergence dynamics could be very different. Additional research is also necessary to determine the exact impact of residues on altering the weather conditions and the species-specific response to that alteration.
From a practical point of view retaining crop residues on soil surface could lead to a lesser need for weed control treatments that would result in reduced production cost. Furthermore, sufficient residue quantity could improve soil properties and water retention and availability, that can influence the crop yield positively [21,43,44]. Some studies have found a significant yield increase by using mulches obtained from crop residues, compared to the scenarios without mulches [45,46]. In their study, Gill et al. [47] compared different scenarios for suppressing weeds in maize fields by also adding crop residue base mulches, also taking into account the effect on crop yield. Mulches consisted of grain legumes and were used in quantities of 5, 10, 15, and 20 t/ha, the authors determined that the residues had only a slightly positive effect on the yield, but the reduction of weed emergence was sufficient to justify the use of mulches. Nevertheless, for many farmers leaving crop residues on their fields is not economically convenient because they employ them for other uses, for example for livestock feed, compost, or bioenergy. It should be also underlined that high amounts of residues can interfere with crop emergence and yield. Thick layers of residues on soil surface can hinder sowing operations and, due to the same processes conferring their weed suppression ability, can cause delayed emergence of crop seedlings and a reduction of their number and biomass [44,45]. Moreover, the presence of crop residues can negatively affect crop growth immobilizing soil nitrogen and reducing nitrogen use efficiency of the crops [46,47]. However, several tactics and precautions can be adopted to minimize the negative impact on crop yield of retaining residues on soil surface. Specific tools and practices can facilitate sowing operation in presence of residues and improve crop emergence and establishment [48]. Crop cultivars with longer coleoptile can better tolerate during the germination-emergence phase the presence of residues on soil surface [45]. Finally, alternating cereals with legumes in crop rotation can provide residues with different chemical composition in order to reduce soil nitrogen immobilization and increase crop yield [46,47]. Farmers should therefore choose and implement according to their local conditions a set of tactics and management operations to maximise the beneficial effects, and to control the potential drawbacks, of retaining crop residues on soil surface.
5. Conclusions
Given the rise of agricultural land under conservation management and the increasing use of different mulches containing crop residues as part of integrated weed management, it becomes even more important to understand the behaviour of weed species under these conditions. From the results presented, it is clear that crop residues do not always inhibit the emergence of weed species. Specifically, it can be promoted by small amounts of residues, capable of maintaining the moisture and allowing enough light to reach the seeds underneath. Therefore, when using crop residues as mulches or when they are left on the soil after harvest as permanent cover, it is of extreme importance to spread residues uniformly on the soil surface to suppress or at least not favour weed emergence. However, for some agronomical practices such as false sowing, where the goal is to eliminate emerged weed seedlings before crop sowing, a quantity of residues that would promote weed emergence could be beneficial.
The general conclusion is that by increasing the quantity of residues, the emergence of all species is reduced, in what seems to be a synergy between the influence of soil moisture and temperature and the physical obstacle for seedling emergence. Given that low crop productivity reduces also the amount of residues left after the harvest, obtaining good crop establishment and grow is vital to achieve sufficient levels of residues for weed suppression. Using crop varieties with larger size can be another tactic to increase the amount of crop residues. Another important point to consider is that slowly decomposing crop residues could extend the weed suppression period. Residue cutting level could therefore be important, intact residues are decomposed slowly than shredded residues. Weed suppression caused by crop residues could be further increased with the introduction of cover crops during the inter-cropping periods. Cover crops, which produce high amounts of biomass, could indeed provide high amounts of suppressive residues in addition to crop residues.
It is important to remember that the experiments in this work were conducted on sterilised soil, and the situation in a no-till field could be more complex. In the surface layer of a mature no-till soil, crop residues interact with the soil and affect physical, chemical, and biotic soil properties. Therefore, weed seeds could also be affected by this interaction [3,49,50,51]. Given the complexity of the factors involved, more studies in no-till fields, with different weed species and different types and amounts of residues, are necessary to improve our knowledge of weed–residue cover interactions. Understanding whether crop residues retained on the soil surface can be a viable option for weed control would be of great utility for farmers and decision-makers in conservation agriculture and conventional agriculture when crop mulches are applied.
Author Contributions
Conceptualization, N.N., D.L., and R.M.; methodology, N.N. and D.L.; software, R.M.; validation, N.N.; investigation, N.N.; data curation, N.N. and R.M.; writing—original draft preparation, N.N.; writing—review and editing, D.L. and R.M.; supervision, R.M.; project administration, R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to their future usage in different projects.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
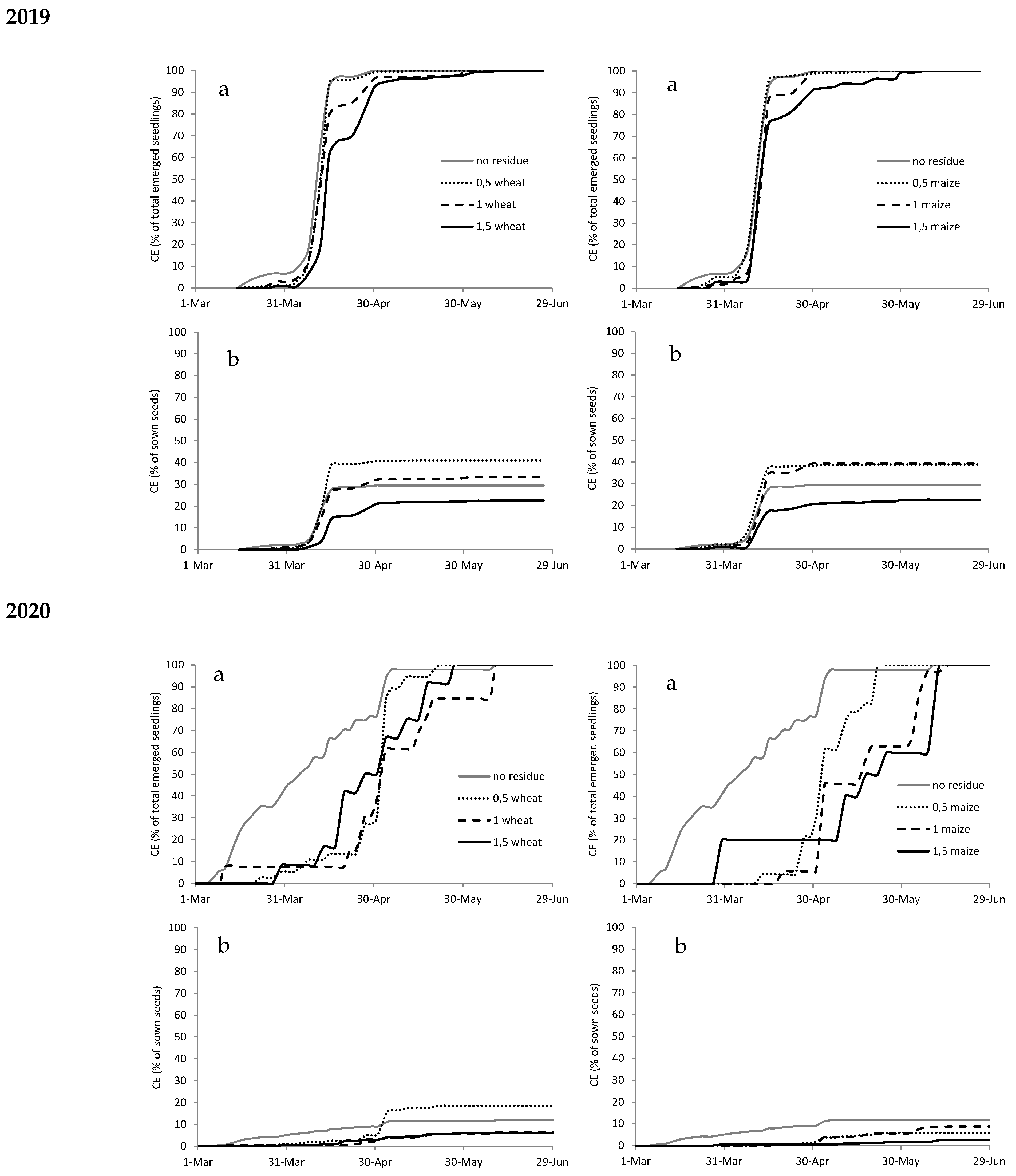
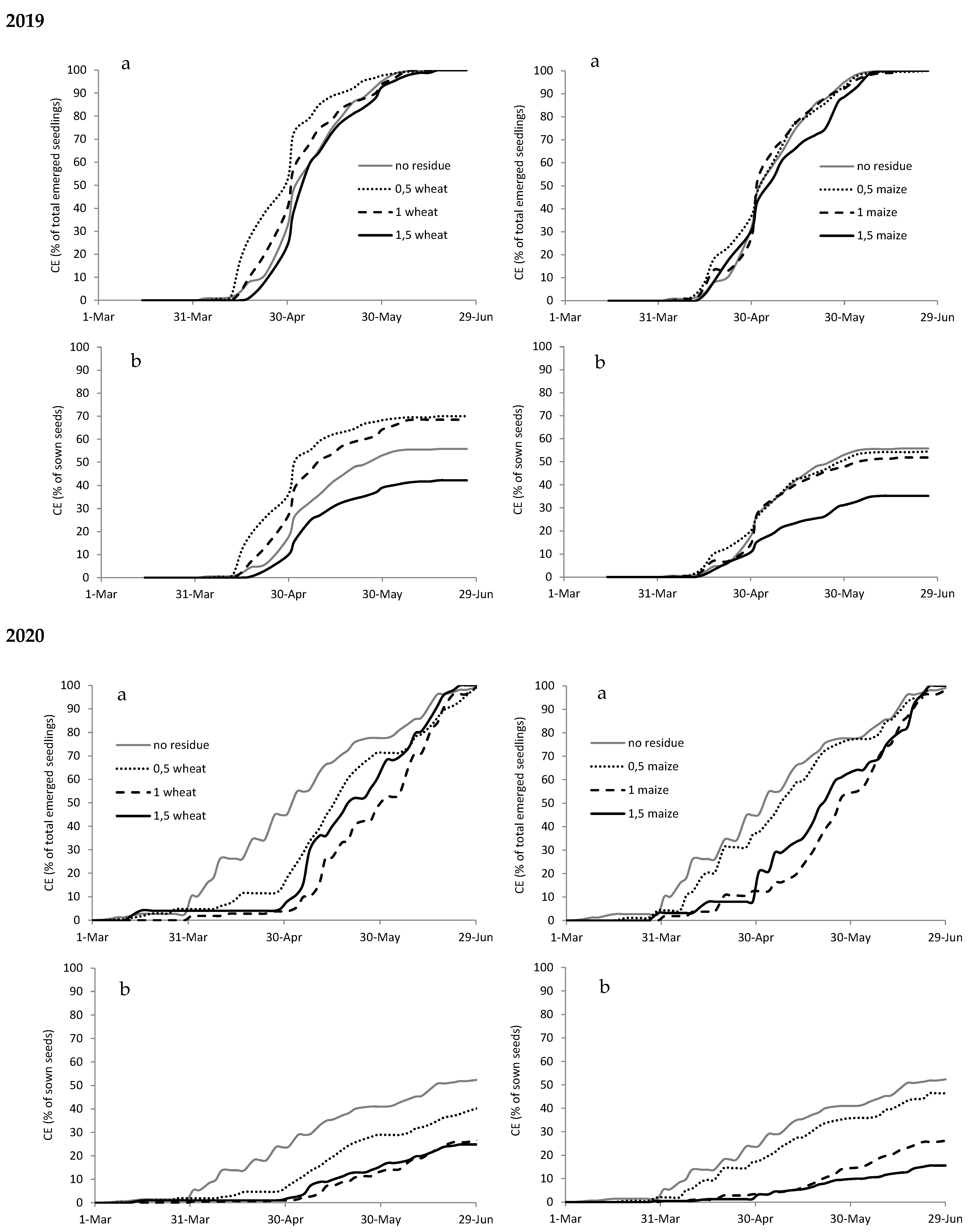
Figure A1.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Abutilon theophrasti in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.
Figure A1.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Abutilon theophrasti in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.

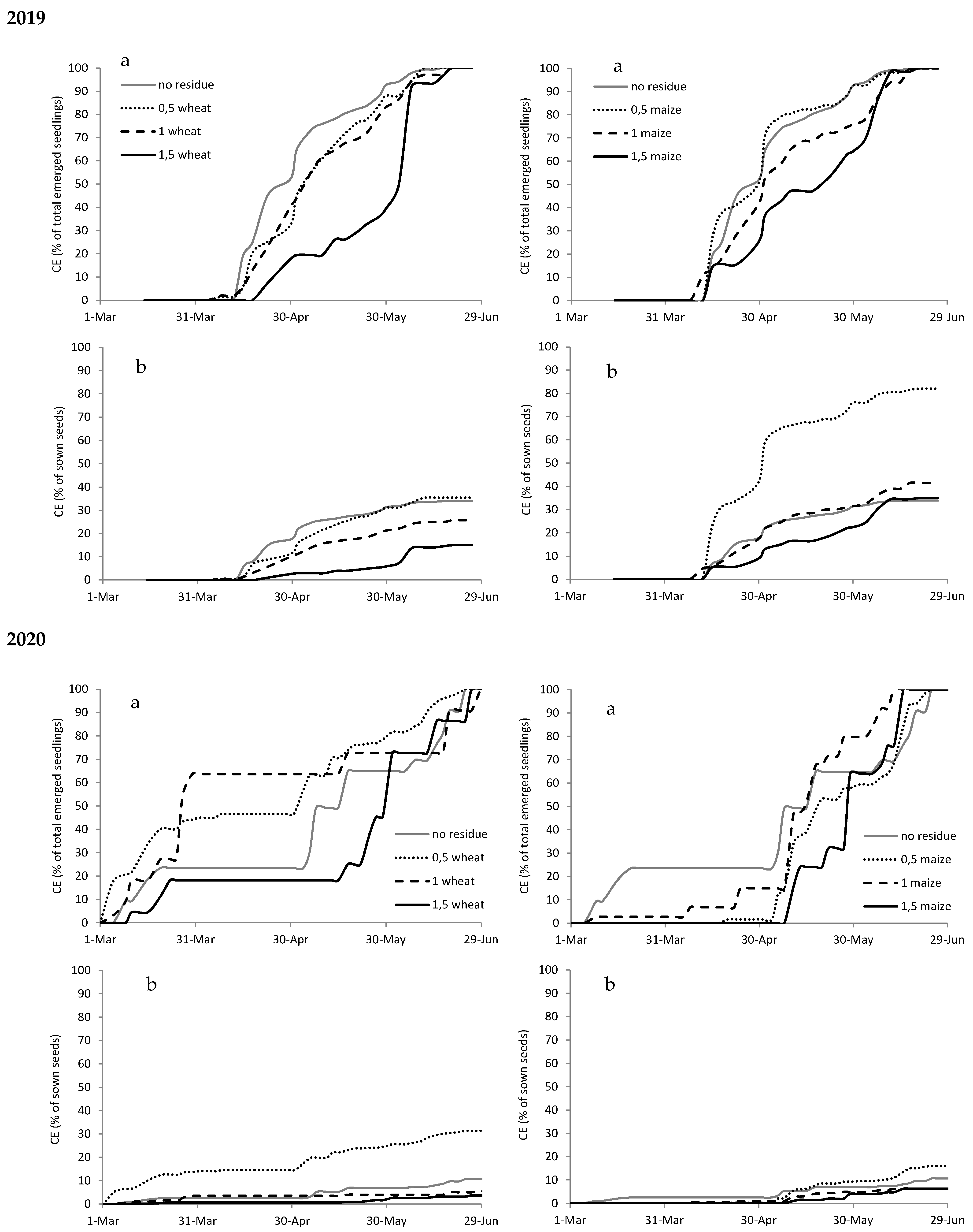
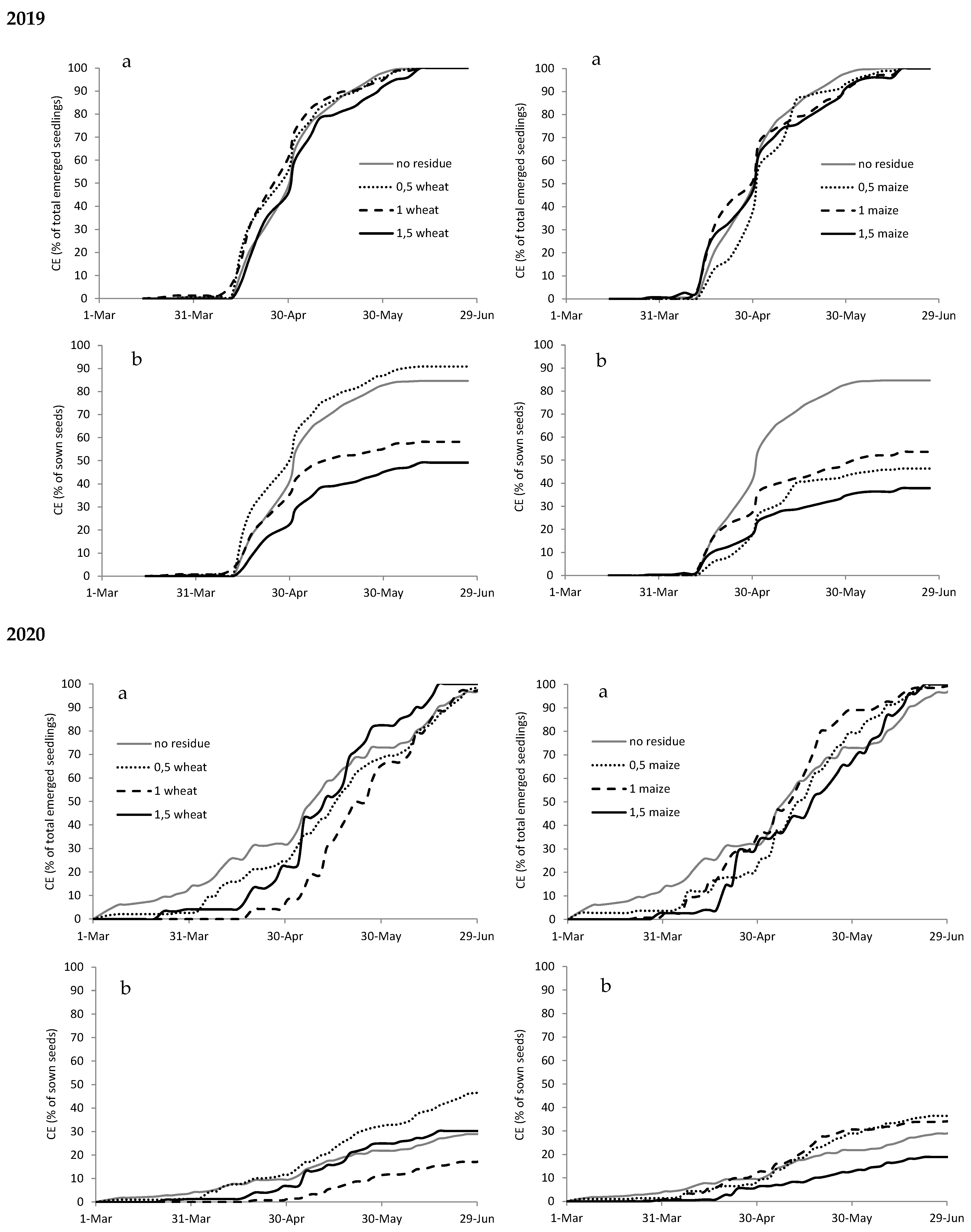
Figure A2.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Amaranthus retroflexus in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.
Figure A2.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Amaranthus retroflexus in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.

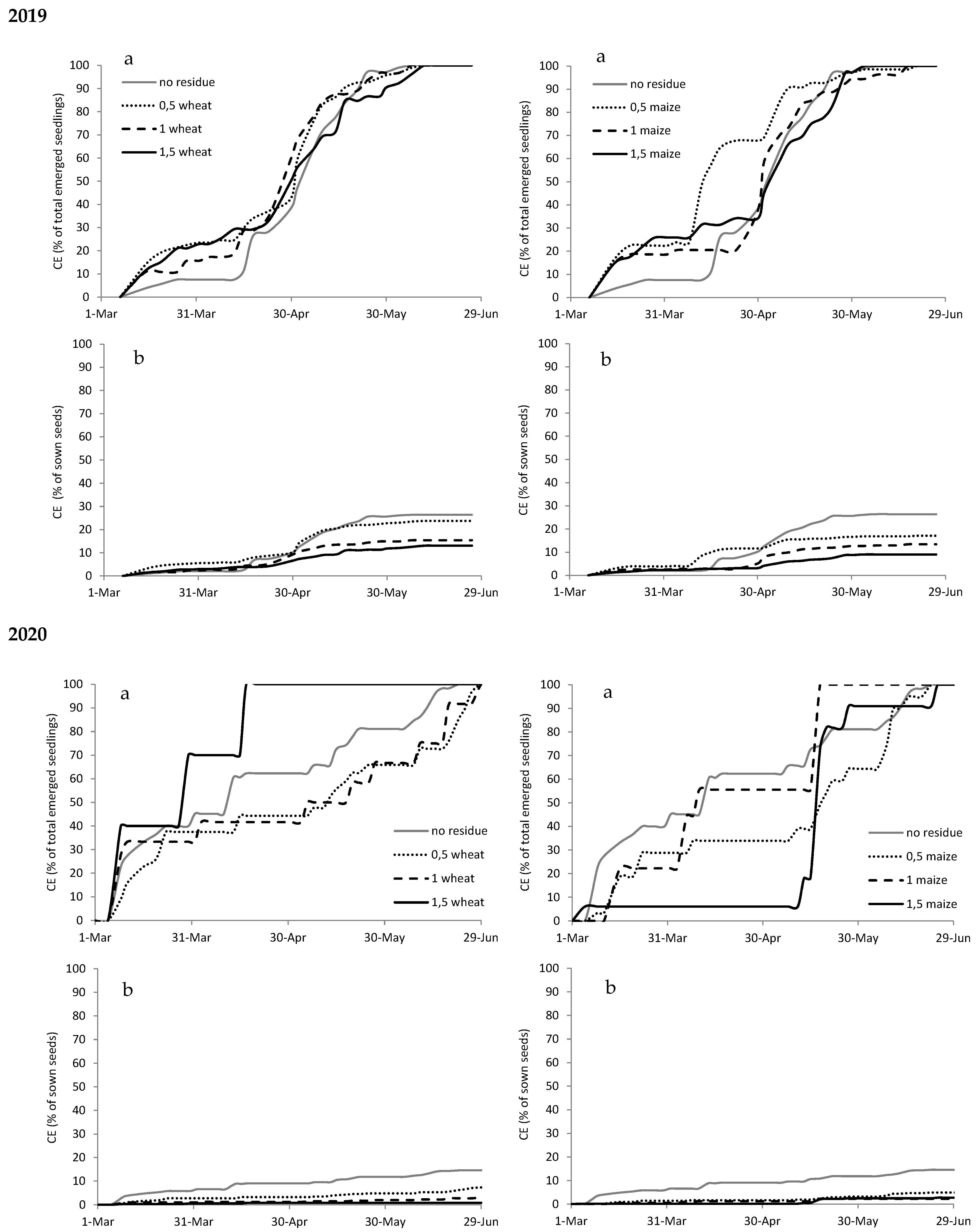
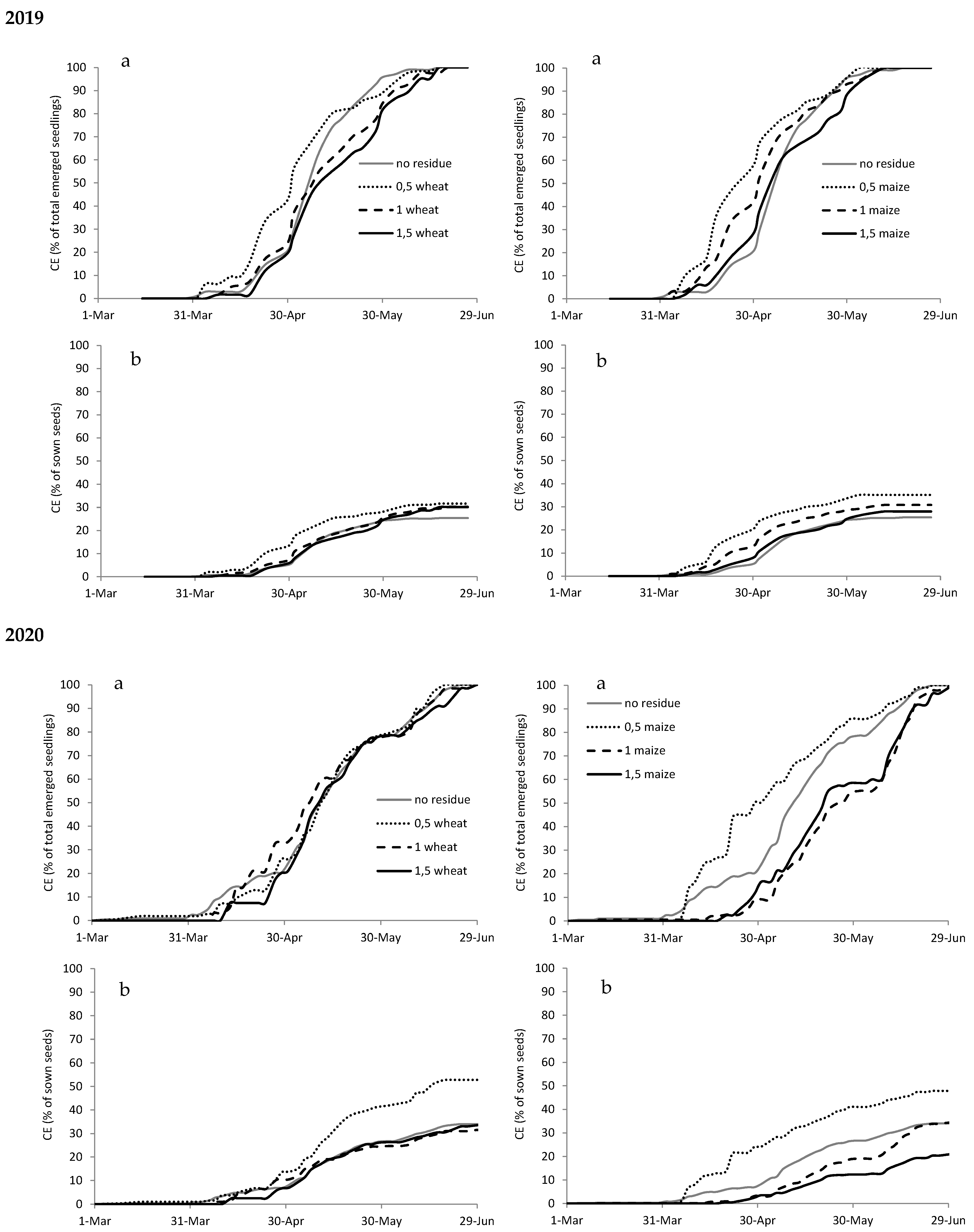
Figure A3.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Chenopodium album in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.
Figure A3.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Chenopodium album in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.

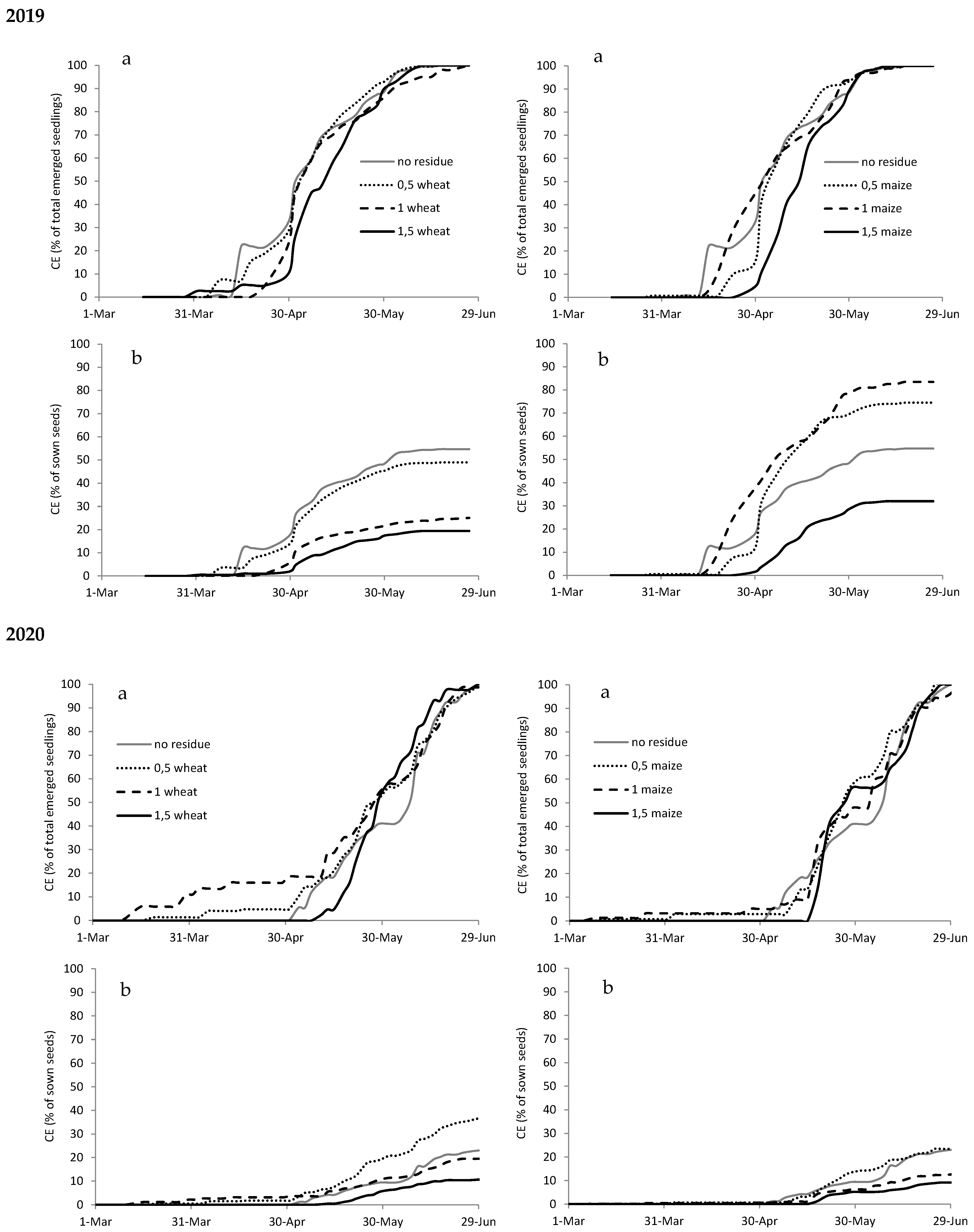
Figure A4.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Digitaria sanguinalis in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.
Figure A4.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Digitaria sanguinalis in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.

Figure A5.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Echinochloa crus-galli in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.
Figure A5.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Echinochloa crus-galli in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.

Figure A6.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Setaria pumila in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.
Figure A6.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Setaria pumila in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.

Figure A7.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Sonchus oleraceus in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.
Figure A7.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Sonchus oleraceus in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.

Figure A8.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Sorghum halepense in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.
Figure A8.
Cumulated emergence (CE) expressed as percentage of total emerged seedlings (a) and as percentage of sown seeds (b) of Sorghum halepense in 2019 and 2020 with different types (wheat on the left and maize on the right) and amounts of crop residues.

References
- Lingenfelter, D.D.; Hartwig, N.L. Introduction to Weeds and Herbicides; Professor of Weed Science, Pennsylvania State University, College of Agricultural Sciences, University Park: State College, PA, USA, 2013; pp. 1–38. [Google Scholar]
- Kohli, R.; Singh, H.P.; Batish, D.R. Allelopathic potential in rice germplasm against ducksalad, redstem and barnyard grass. J. Crop. Prot. 2008, 4, 287–301. [Google Scholar]
- Nichols, V.; Verhulst, N.; Cox, R.; Govaerts, B. Weed dynamics and conservation agriculture principles: A review. Field Crop. Res. 2015, 183, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Chen, E.; Chen, Z.; Cheng, J.; Dou, C.; Du, J.; Dickinson, R.E.; Fan, W.; Fang, H.; Fu, Q.; et al. Land Cover and Land use Changes. In Advanced Remote Sensing; Liang, S., Li, X., Wang, J., Eds.; Elsevier Inc.: San Diego, CA, USA, 2012; pp. 703–772. ISBN 9780123859549. [Google Scholar]
- Rusinamhodzi, L. Crop rotations and residue management in conservation agriculture. In Conservation Agriculture; Farooq, M., Siddique, K.H.M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–665. ISBN 9783319116204. [Google Scholar]
- Bewley, J.D. Seed Germination and Dormancy. Plant Cell Online 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Zimdahl, L.R. Fundamentals of Weed Science, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2007; ISBN 9780080549859. [Google Scholar]
- Savabi, M.R.; Stott, D.E. Plant residue impact on rainfall interception. Trans. Am. Soc. Agric. Eng. 1994, 37, 1093–1098. [Google Scholar] [CrossRef]
- Ruan, H.; Ahuja, L.R.; Green, T.R.; Benjamin, J.G. Residue Cover and Surface-Sealing Effects on Infiltration. Soil Sci. Soc. Am. J. 2001, 65, 853–861. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, J.R.; Mohler, C.L. Light Transmittance, Soil Temperature, and Soil Moisture under Residue of Hairy Vetch and Rye. Agron. J. 1993, 85, 673–680. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Singh, R.G.; Mahajan, G. Ecology and management of weeds under conservation agriculture: A review. Crop. Prot. 2012, 38, 57–65. [Google Scholar] [CrossRef]
- Arriaga, F.J.; Guzman, J.; Lowery, B. Conventional Agricultural Production Systems and Soil Functions; Elsevier Inc.: Amsterdam, The Netherlands, 2017; ISBN 9780128054017. [Google Scholar]
- FAO Conservation Agriculture. Available online: http://www.fao.org/conservation-agriculture/en/ (accessed on 9 November 2020).
- Kassam, A.; Friedrich, T.; Derpsch, R.; Kienzle, J. Overview of the Worldwide Spread of Conservation Agriculture. FACTS Rep. 2015, 8, 1–11. [Google Scholar]
- Eslami, S.V. Weed management in conservation agriculture systems. In Recent Advances in Weed Management; Springer Science+Business Media: New York, NY, USA, 2014; ISBN 9781493910199. [Google Scholar]
- Teasdale, J.R.; Mohler, C.L. The quantitative relationship between weed emergence and the physical properties of mulches. Weed Sci. 2000, 48, 385–392. [Google Scholar] [CrossRef]
- Kruidhof, H.M.; Bastiaans, L.; Kropff, M.J. Cover crop residue management for optimizing weed control. Plant Soil 2009, 318, 169–184. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Abugho, S.B. Effect of Crop Residue on Seedling Emergence and Growth of Selected Weed Species in a Sprinkler-Irrigated Zero-Till Dry-Seeded Rice System. Weed Sci. 2013, 61, 403–409. [Google Scholar] [CrossRef]
- Eurostat Agricultural Production—Crops | Statistics Explained. Agric. For. Fish. Stat. 2019, 1–19. [CrossRef]
- Acharya, C.L.; Hati, K.M.; Bandyopadhyay, K.K. Mulches. In Encyclopedia of Soils in the Environment; Elsevier: Amsterdam, The Netherlands, 2005; Volume 4, pp. 521–532. ISBN 9780080547954. [Google Scholar] [CrossRef]
- Liebman, M.; Gallandt, E.R. Many Little Hammers: Ecological Management of Crop-Weed Interactions. Ecol. Agric. 1997, 291–343. [Google Scholar] [CrossRef]
- Ranaivoson, L.; Naudin, K.; Ripoche, A.; Affholder, F.; Rabeharisoa, L.; Corbeels, M. Agro-ecological functions of crop residues under conservation agriculture. A review. Agron. Sustain. Dev. 2017, 37. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.T.; Du, G.Z.; Chen, J.K. Seed size in relation to phylogeny, growth form and longevity in a subalpine meadow on the east of the Tibetan Plateau. Folia Geobot. 2004, 39, 129–142. [Google Scholar] [CrossRef]
- Norden, N.; Daws, M.I.; Antoine, C.; Gonzalez, M.A.; Garwood, N.C.; Chave, J. The relationship between seed mass and mean time to germination for 1037 tree species across five tropical forests. Funct. Ecol. 2009, 23, 203–210. [Google Scholar] [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Tanveer, A.; Tasneem, M.; Khaliq, A.; Javaid, M.M.; Chaudhry, M.N. Influence of seed size and ecological factors on the germination and emergence of field bindweed (Convolvulus arvensis). Planta Daninha 2013, 31, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Emrah, C.; Fahrettin, T. Seed Size Effects on Germination, Survival and Seedling Growth of Castanea sativa Mill. J. Biol. Sci. 2007, 7, 438–441. [Google Scholar]
- Leishman, M.R.; Wright, I.J.; Moles, A.T.; Westoby, M. The evolutionary ecology of seed size. In Seeds Ecol. Regen. Plant Communities; Fenner, M., Ed.; CABI: Wallingford, UK, 2000; pp. 31–57. ISBN 9780851999470. [Google Scholar]
- Milberg, P.; Andersson, L.; Thompson, K. Large-seeded spices are less dependent on light for germination than small-seeded ones. Seed Sci. Res. 2000, 10, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Testani, E.; Ciaccia, C.; Campanelli, G.; Leteo, F.; Salvati, L.; Canali, S. Mulch-Based No-Tillage Effects on Weed Community and Management in an Organic Vegetable System. Agronomy 2019, 9, 594. [Google Scholar] [CrossRef] [Green Version]
- Trichard, A.; Alignier, A.; Chauvel, B.; Petit, S. Identification of weed community traits response to conservation agriculture. Agric. Ecosyst. Environ. 2013, 179, 179–186. [Google Scholar] [CrossRef]
- Shaban, M. Effect of water and temperature on seed germination and emergence as a seed hydrothermal time model. Int. J. Adv. Biol. Biomed. Res. 2013, 1, 1686–1691. [Google Scholar]
- Calado, J.M.G.; Basch, G.; de Carvalho, M. Weed emergence as influenced by soil moisture and air temperature. J. Pest. Sci. 2009, 82, 81–88. [Google Scholar] [CrossRef]
- Bir, M.S.H.; Eom, M.Y.; Uddin, M.R.; Park, T.S.; Kang, H.W.; Kim, D.S.; Park, K.W. Weed Population Dynamics under Climatic Change. Weed Turfgrass Sci. 2014, 3, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Nussbaum, E.S.; Wiese, A.F.; Crutchfield, D.E.; Chenault, E.W. The Effects of Temperature and Rainfall on Emergence and Growth of Eight Weeds; Nussbaum, E.S., Wiese, A.F., Crutchfield, D.E., Chenault, E.W., Lavake, D., Eds.; Weed Science Society of America and Allen Press S: Cambridge, UK, 2008; Volume 33, pp. 165–170. [Google Scholar]
- Dekker, J. The foxtail (Setaria) species-group. Weed Sci. 2003, 51, 641–656. [Google Scholar] [CrossRef]
- Gâdea, Ş.; Bogdan, I.; Vâtcă, S. Seed germination study for Setaria glauca L. Species. Agric. Sci. Pract. 2014, 4, 91–92. [Google Scholar]
- Gallart, M.; Mas, M.T.; Antoni, V.M.C. Demography of Digitaria sanguinalis: Effect of the emergence time on survival, reproduction, and biomass. Weed Biol. Manag. 2010, 10, 132–140. [Google Scholar] [CrossRef]
- Leguizamòn, E.S. Seed survival and patterns of seedling emergence in Sorghum halepense (L.) Pers. Weed Res. 1986, 26, 397–404. [Google Scholar] [CrossRef]
- Peerzada, A.M.; Ali, H.H.; Hanif, Z.; Bajwa, A.A.; Kebaso, L.; Frimpong, D.; Iqbal, N.; Namubiru, H.; Hashim, S.; Rasool, G.; et al. Eco-biology, impact, and management of Sorghum halepense (L.) Pers. Biol. Invasions 2017, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Ma, Y.L.; Feng, G.J.; Li, H.H. Abiotic factors affecting seed germination and early seedling emergence of large crabgrass (Digitaria sanguinalis). Planta Daninha 2018, 36, 1–10. [Google Scholar] [CrossRef]
- Loddo, D.; Sousa, E.; Masin, R.; Calha, I.; Zanin, G.; Fernández-Quintanilla, C.; Dorado, J. Estimation and Comparison of Base Temperatures for Germination of European Populations of Velvetleaf (Abutilon theophrasti) and Jimsonweed (Datura stramonium). Weed Sci. 2013, 61, 443–451. [Google Scholar] [CrossRef]
- Sadeghloo, A.; Asghari, J.; Ghaderi-Far, F. Seed germination and seedling emergence of velvetleaf (Abutilon theophrasti) and Barnyardgrass (Echinochloa crus-galli). Planta Daninha 2013, 31, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, M.; Painuli, D.K. Modeling rice seedling emergence and growth under tillage and residue management in a rice-wheat system on a Vertisol in Central India. Soil Tillage Res. 2004, 76, 167–174. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Bruce, S.E.; Kirkegaard, J.A. Longer coleoptiles improve emergence through crop residues to increase seedling number and biomass in wheat (Triticum aestivum L.). Plant Soil 2005, 272, 87–100. [Google Scholar] [CrossRef]
- Pagnani, G.; Galieni, A.; D’Egidio, S.; Visioli, G.; Stagnari, F.; Pisante, M. Effect of soil tillage and crop sequence on grain yield and quality of durum wheat in Mediterranean areas. Agronomy 2019, 9, 488. [Google Scholar] [CrossRef] [Green Version]
- Radicetti, E.; Mancinelli, R.; Moscetti, R.; Campiglia, E. Management of winter cover crop residues under different tillage conditions affects nitrogen utilization efficiency and yield of eggplant (Solanum melanogena L.) in Mediterranean environment. Soil Tillage Res. 2016, 155, 329–338. [Google Scholar] [CrossRef]
- Swanepoel, P.A.; le Roux, P.J.G.; Agenbag, G.A.; Strauss, J.A.; Maclaren, C. Seed-drill opener type and crop residue load affect canola establishment, but only residue load affects yield. Agron. J. 2019, 111, 1658–1665. [Google Scholar] [CrossRef] [Green Version]
- Fracchiolla, M.; Stellacci, A.M.; Cazzato, E.; Tedone, L.; Ali, S.A.; De Mastro, G. Effects of conservative tillage and nitrogen management on weed seed bank after a seven-year durum wheat—Faba Bean rotation. Plants 2018, 7, 82. [Google Scholar] [CrossRef] [Green Version]
- Kremer, R.J.; Li, J. Developing weed-suppressive soils through improved soil quality management. Soil Tillage Res. 2003, 72, 193–202. [Google Scholar] [CrossRef]
- Nikolić, N.; Squartini, A.; Concheri, G.; Stevanato, P.; Zanin, G.; Masin, R. Weed Seed Decay in No-Till Field and Planted Riparian Buffer Zone. Plants 2020, 9, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Experimental site; (a) 5 cm of soil removed; (b) redistribution of soil after sterilisation; (c) sowing and covering with residues; (d) final appearance.
Figure 1.
Experimental site; (a) 5 cm of soil removed; (b) redistribution of soil after sterilisation; (c) sowing and covering with residues; (d) final appearance.

Figure 2.
Influence of crop residues (both maize and wheat) on weed species emergence expressed as % on control (no residues). Values are means of all weed species. Different letters indicate significant difference (p-value < 0.05), and the bars indicate the standard error.
Figure 2.
Influence of crop residues (both maize and wheat) on weed species emergence expressed as % on control (no residues). Values are means of all weed species. Different letters indicate significant difference (p-value < 0.05), and the bars indicate the standard error.

Figure 3.
Differences in emergence between weed species in the presence of crop residues. The emergence is expressed as a percentage relative to control (no residues). Emergence values are means of the two types of residues (wheat and maize) and three levels (0.5, 1, and 1.5) of residues quantities. Different letters indicate significant difference (p-value < 0.05), and the bars indicate the standard error.
Figure 3.
Differences in emergence between weed species in the presence of crop residues. The emergence is expressed as a percentage relative to control (no residues). Emergence values are means of the two types of residues (wheat and maize) and three levels (0.5, 1, and 1.5) of residues quantities. Different letters indicate significant difference (p-value < 0.05), and the bars indicate the standard error.

Figure 4.
Daily soil temperature and precipitations in 2019 (a) and 2020 (b).


Figure 5.
Differences in emergence between weed species in different years in the presence of crop residues. The emergence is expressed as a percentage of control (no residues). Emergence values are means of the two types of residues (wheat and maize) and three levels (0.5, 1, and 1.5) of residues quantities. Different letters indicate significant difference (p-value < 0.05), and the bars indicate the standard error.
Figure 5.
Differences in emergence between weed species in different years in the presence of crop residues. The emergence is expressed as a percentage of control (no residues). Emergence values are means of the two types of residues (wheat and maize) and three levels (0.5, 1, and 1.5) of residues quantities. Different letters indicate significant difference (p-value < 0.05), and the bars indicate the standard error.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Quantities of maize and wheat residues used for the trials.
| Quantity | Residue (kg/m2) | |
|---|---|---|
| Maize | Wheat | |
| 0.5 | 0.308 | 0.176 |
| 1 | 0.616 | 0.352 |
| 1.5 | 0.924 | 0.528 |
Table 2.
Full factorial ANOVA on total emergence monitored in the two years.
| Factors | Df | F-Value | p-Value |
|---|---|---|---|
| Years (Y) | 1 | 1.209 | ns |
| Species (S) | 42 | 1.455 | 0.048 |
| Crop residues (Cr) | 24 | 0.605 | ns |
| Residue quantities (Rq) | 32 | 2.078 | 0.001 |
| Interactions | |||
| Y × S | 7 | 3.155 | 0.004 |
| Y × Cr | 1 | 0.203 | ns |
| Y × Rq | 2 | 2.050 | ns |
| S × Rq | 28 | 0.704 | ns |
| S × Cr | 21 | 0.375 | ns |
| Cr × Rq | 16 | 0.558 | ns |
| Y × S × Cr | 7 | 0.174 | ns |
| Y × S × Rq | 14 | 0.664 | ns |
| Y × Cr × Rq | 2 | 1.253 | ns |
| S × Cr × Rq | 14 | 0.395 | ns |
| Y × S × Cr × Rq | 14 | 0.294 | ns |
ns = not significant, Df = Degrees of freedom.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nikolić, N.; Loddo, D.; Masin, R. Effect of Crop Residues on Weed Emergence. Agronomy 2021, 11, 163. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010163
AMA Style
Nikolić N, Loddo D, Masin R. Effect of Crop Residues on Weed Emergence. Agronomy. 2021; 11(1):163. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010163
Chicago/Turabian StyleNikolić, Nebojša, Donato Loddo, and Roberta Masin. 2021. "Effect of Crop Residues on Weed Emergence" Agronomy 11, no. 1: 163. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010163
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.