Comparison of Drivers of Soil Microbial Communities Developed in Karst Ecosystems with Shallow and Deep Soil Depths


Abstract
:1. Introduction
2. Materials and Methods
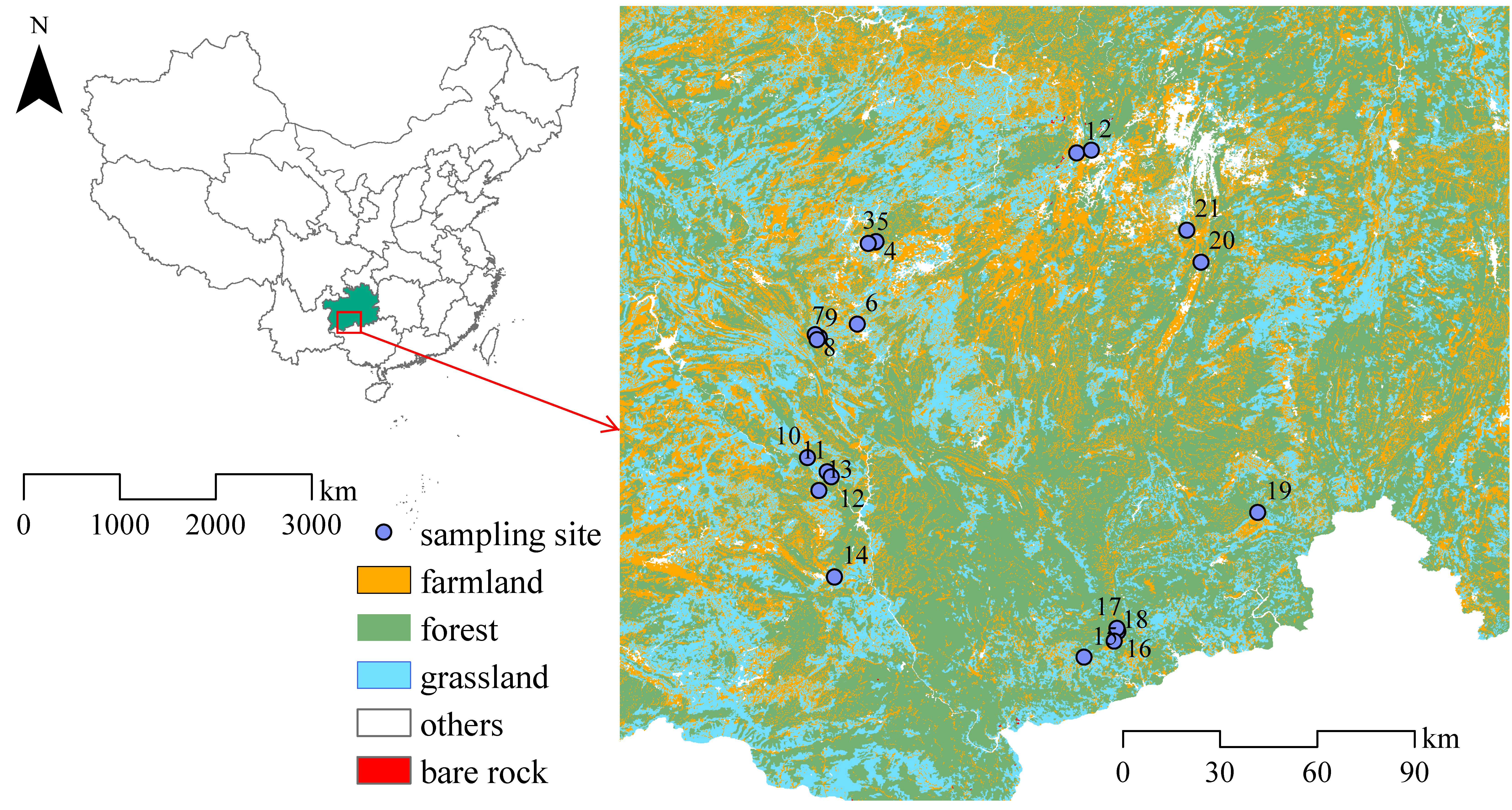
2.1. Site Description
2.2. Community Investigation and Sample Collection
2.3. Sample Analysis
2.3.1. Plant and Soil Physicochemical Properties
2.3.2. Phospholipid Fatty Acid (PLFA) Analysis
2.4. Data Analysis
2.4.1. Plant Species Diversity Calculation
- Tree Pi = (relative frequency + relative density + relative basal coverage)/3
- Shrub Pi = (relative frequency + relative density + relative coverage)/3
- Herbs Pi = (relative frequency + relative height + relative coverage)/3
2.4.2. Soil Microbial Diversity Calculation
- (1)
- Species abundance index (S)S = N
- (2)
- Shannon–Wiener species diversity index (H)where N is the number of soil microbial PLFAs in each soil sample. Microorganism Pi = relative concentration, calculated by the ratio of the content of each PLFA to the total microbial PLFA content of the sample.
2.4.3. Selection of Minimum Data Set
2.4.4. Statistical Analysis
2.5. Other Data Source
3. Results
3.1. Physiochemical Properties of the Plant and Soil of the Two Soil Depth Community Categories
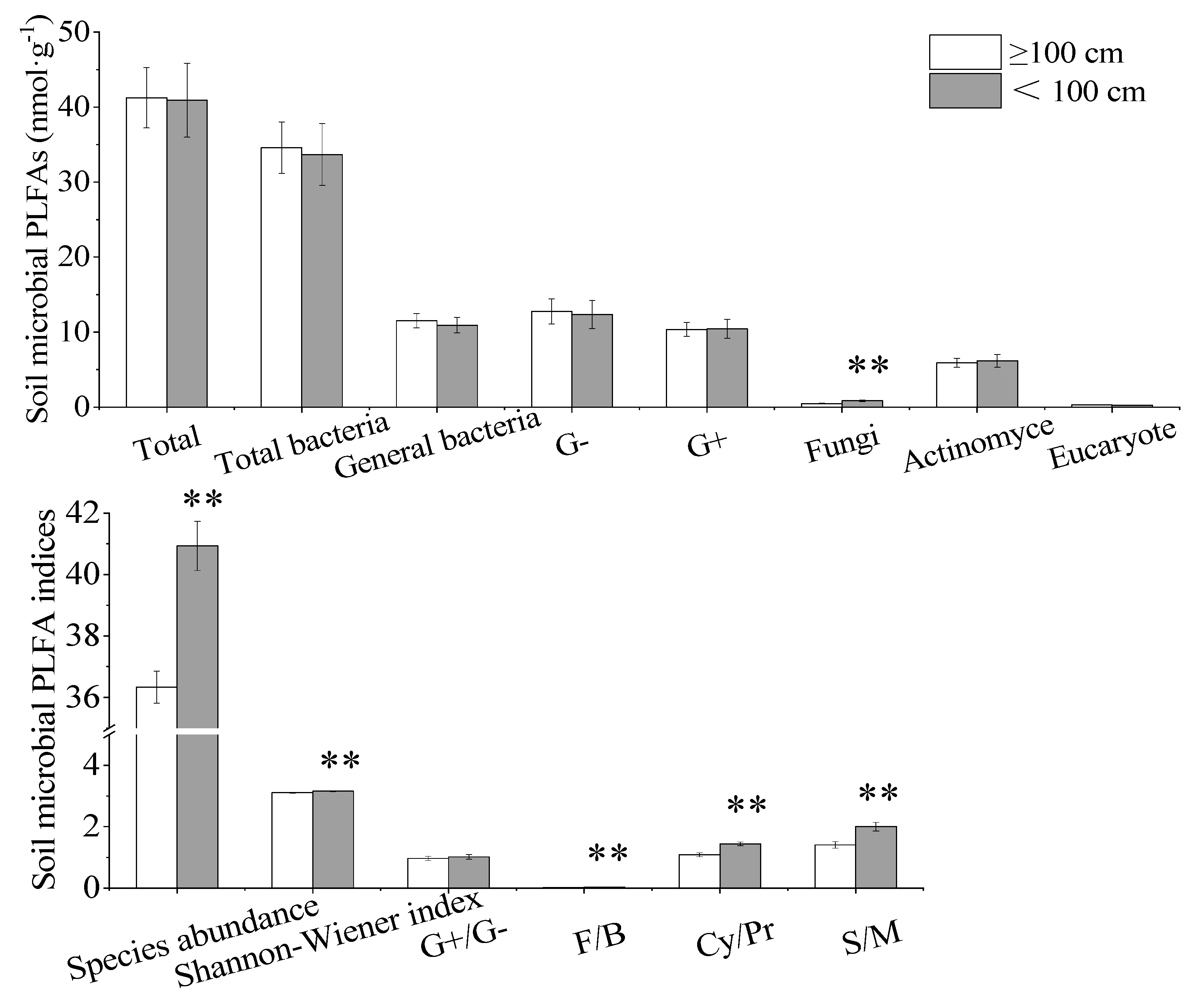
3.2. Soil PLFA Profiles of the Two Soil Depth Communitiey Categories
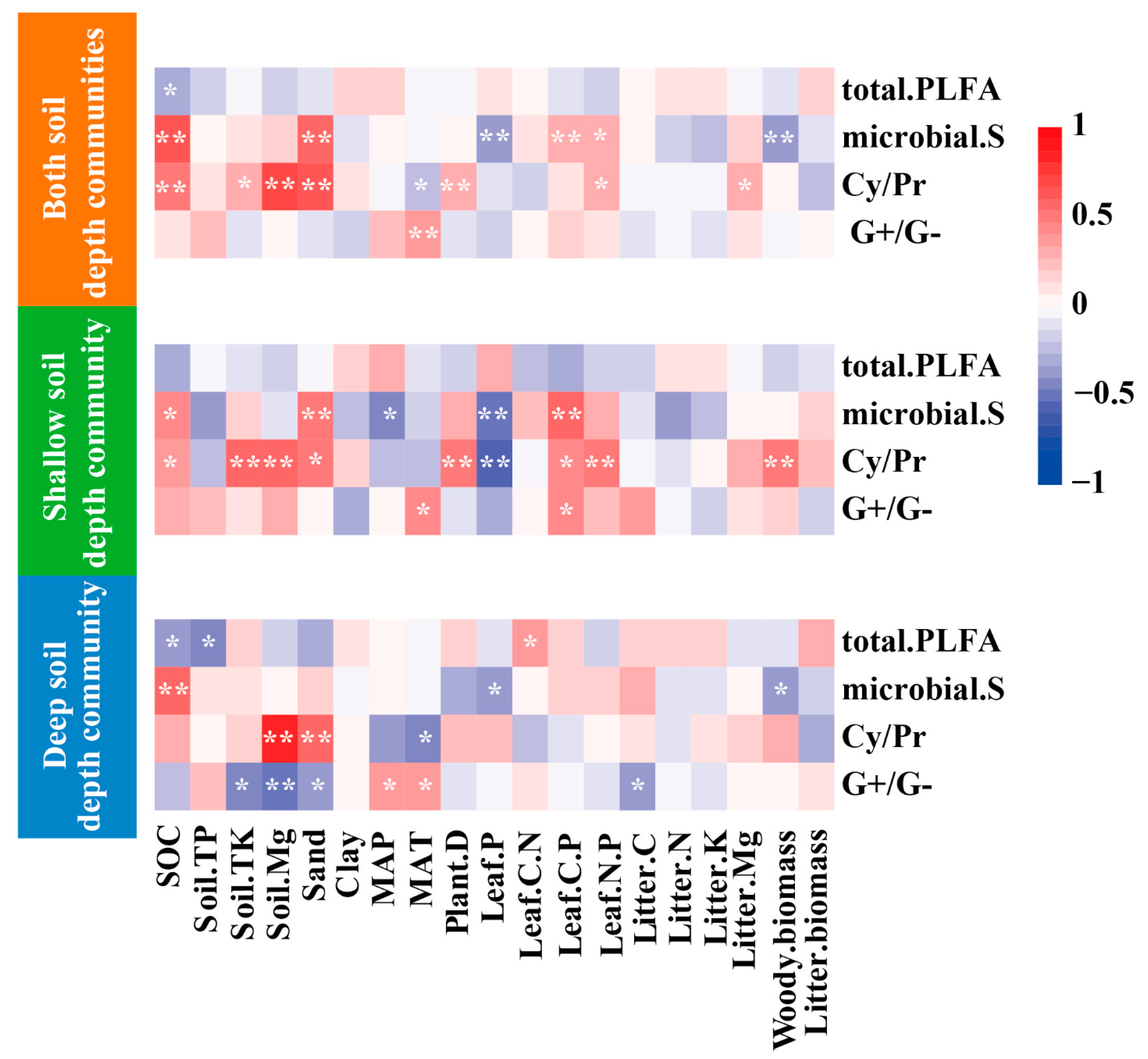
3.3. Pearson’s Correlation Analysis between Soil PLFA Profiles and Environmental Factors in the Communities with Shallow and Deep Soil Depths
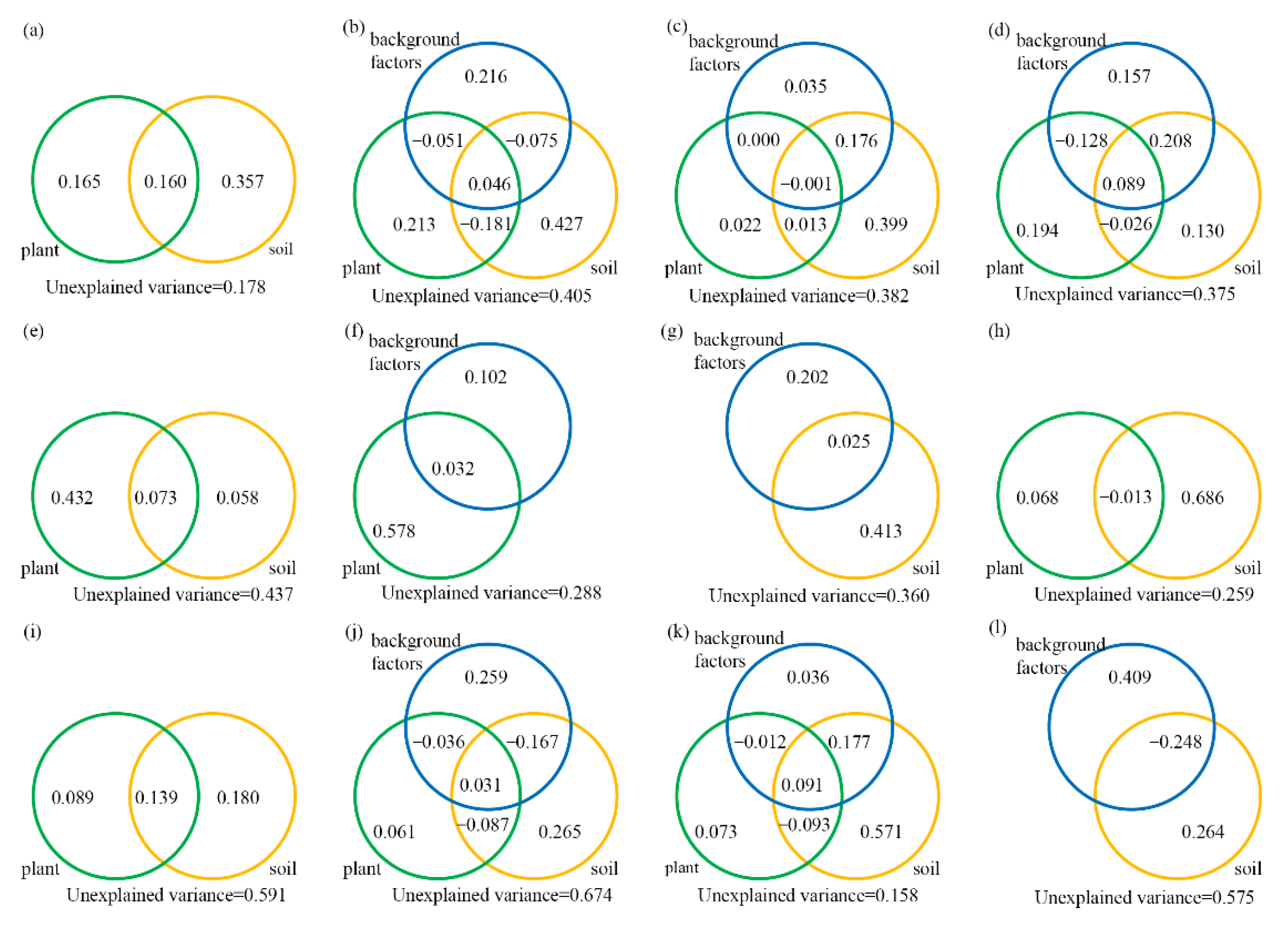
3.4. Combined Effects of Plant, Edaphic and Background Factors on Microbial Communities
4. Discussion
4.1. Driving Factors of Microbial Biomass and Diversity
4.2. Driving Factors of Microbial Community Structures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Grosso, F.; Iovieno, P.; Alfani, A.; De Nicola, F. Structure and activity of soil microbial communities in three Mediterranean forests. Appl. Soil Ecol. 2018, 130, 280–287. [Google Scholar] [CrossRef]
- Wang, X.H.; Yang, J.H.; Xie, X.F.; Chen, X.J.; Pu, L.J.; Zhang, X.W. Soil microbial succession with soil development since costal reclamation. Catena 2020, 187, 8. [Google Scholar] [CrossRef]
- Richter, A.; Schöning, I.; Kahl, T.; Bauhus, J.; Ruess, L. Regional environmental conditions shape microbial community structure stronger than local forest management intensity. For. Ecol. Manag. 2018, 409, 250–259. [Google Scholar] [CrossRef]
- Wang, J.M.; Wang, Y.; He, N.P.; Ye, Z.Q.; Chen, C.; Zang, R.G.; Feng, Y.M.; Lu, Q.; Li, J.W. Plant functional traits regulate soil bacterial diversity across temperate deserts. Sci. Total Environ. 2020, 715, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.K.; Zhang, X.Y.; Wang, X.; Fu, S.Y.; Wu, S.J.; Lu, X.Q.; Ren, C.J.; Han, X.H.; Yang, G.H. Soil bacteria and fungi respond differently to plant diversity and plant family composition during the secondary succession of abandoned farmland on the Loess Plateau, China. Plant. Soil 2020, 448, 183–200. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Caruso, T. Soil microbial community responses to climate extremes: Resistance, resilience and transitions to alternative states. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 13. [Google Scholar] [CrossRef] [Green Version]
- Deltedesco, E.; Keiblinger, K.M.; Piepho, H.P.; Antonielli, L.; Potsch, E.M.; Zechmeister-Boltenstern, S.; Gorfer, M. Soil microbial community structure and function mainly respond to indirect effects in a multifactorial climate manipulation experiment. Soil Biol. Biochem. 2020, 142, 12. [Google Scholar] [CrossRef]
- Baldrian, P.; Merhautova, V.; Cajthaml, T.; Petrankova, M.; Snajdr, J. Small-scale distribution of extracellular enzymes, fungal, and bacterial biomass in Quercus petraea forest topsoil. Biol. Fertil. Soils 2010, 46, 717–726. [Google Scholar] [CrossRef]
- Llado, S.; Lopez-Mondejar, R.; Baldrian, P. Drivers of microbial community structure in forest soils. Appl. Microbiol. Biotechnol. 2018, 102, 4331–4338. [Google Scholar] [CrossRef]
- Yang, Y.; Cheng, H.; Dou, Y.X.; An, S.S. Plant and soil traits driving soil fungal community due to tree plantation on the Loess Plateau. Sci. Total Environ. 2020, 708, 12. [Google Scholar] [CrossRef]
- Chen, X.L.; Henriksen, T.M.; Svensson, K.; Korsaeth, A. Long-term effects of agricultural production systems on structure and function of the soil microbial community. Appl. Soil Ecol. 2020, 147, 10. [Google Scholar] [CrossRef]
- Qiu, J.M.; Cao, J.H.; Lan, G.Y.; Liang, Y.M.; Wang, H.; Li, Q. The Influence of Land Use Patterns on Soil Bacterial Community Structure in the Karst Graben Basin of Yunnan Province, China. Forests 2020, 11, 15. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.S.; Scholer, A.; Nielsen, T.K.; Hansen, L.H.; Schloter, M.; Winding, A. Land use as a driver for protist community structure in soils under agricultural use across Europe. Sci. Total Environ. 2020, 717, 11. [Google Scholar] [CrossRef]
- Aho, K.A.; Weber, C.F.; Christner, B.C.; Vinatzer, B.A.; Morris, C.E.; Joyce, R.; Failor, K.C.; Werth, J.T.; Bayless-Edwards, A.L.H.; Schmale, D.G. Spatiotemporal patterns of microbial composition and diversity in precipitation. Ecol. Monogr. 2020, 90, e01394. [Google Scholar] [CrossRef] [Green Version]
- Bernhard, A.E.; Chelsky, A.; Giblin, A.E.; Roberts, B.J. Influence of local and regional drivers on spatial and temporal variation of ammonia-oxidizing communities in Gulf of Mexico salt marshes. Environ. Microbiol. Rep. 2019, 11, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Mi, J.; Chu, P.; Cheng, J.; Zhang, L.; Pan, Q.; Xie, Y.; Bai, Y. Patterns and drivers of soil microbial communities along a precipitation gradient on the Mongolian Plateau. Landsc. Ecol. 2014, 30, 1669–1682. [Google Scholar] [CrossRef]
- Huang, Y.M.; Liu, D.; An, S.S. Effects of slope aspect on soil nitrogen and microbial properties in the Chinese Loess region. Catena 2015, 125, 135–145. [Google Scholar] [CrossRef]
- Yang, Y.; Cheng, H.; Gao, H.; An, S.S. Response and driving factors of soil microbial diversity related to global nitrogen addition. Land Degrad. Dev. 2019, 15, 190–204. [Google Scholar] [CrossRef]
- Nie, Y.P.; Ding, Y.L.; Zhang, H.L.; Chen, H.S. Comparison of woody species composition between rocky outcrops and nearby matrix vegetation on degraded karst hillslopes of Southwest China. J. For. Res. 2019, 30, 911–920. [Google Scholar] [CrossRef]
- Zhang, C.H.; Qi, X.K.; Wang, K.L.; Zhang, M.Y.; Yue, Y.M. The application of geospatial techniques in monitoring karst vegetation recovery in southwest China: A review. Prog. Phys. Geogr. 2017, 41, 450–477. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; Cardenas, E.; Maas, K.R.; Leung, H.; McNeil, L.; Berch, S.; Chapman, W.; Hope, G.; Kranabetter, J.M.; Dube, S.; et al. Biogeography and organic matter removal shape long-term effects of timber harvesting on forest soil microbial communities. ISME J. 2017, 11, 2552–2568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neupane, J.; Guo, W.; Acosta-Martinez, V.; Zhang, F.; Lin, Z.; Cano, A. Assessing Spatial Pattern of Soil Microbial Community at Landscape Scale for Precision Soil Management. In Proceedings of the ASA, CSSA and SSSA International Annual Meetings, San Antonio, TX, USA, 10–13 November 2019. [Google Scholar]
- Wang, S.J.; Zhang, X.B.; Bai, X.Y. An outline of karst geomorphology zoning in the karst areas of southern China. Mt. Res. 2015, 33, 641–648. (In Chinese) [Google Scholar]
- Jiang, Z.C.; Lian, Y.Q.; Qin, X.Q. Rocky desertification in Southwest China: Impacts, causes, and restoration. Earth Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Bai, Y.E.; Liu, Q.; Gu, Z.F.; Lu, Y.R.; Sheng, Z.P. The dissolution mechanism and karst development of carbonate rocks in karst rocky desertification area of Zhenfeng-Guanling-Huajiang County, Guizhou, China. Carbonates Evaporites 2019, 34, 45–51. [Google Scholar] [CrossRef]
- Yan, Y.J.; Dai, Q.H.; Yuan, Y.F.; Peng, X.D.; Zhao, L.S.; Yang, J. Effects of rainfall intensity on runoff and sediment yields on bare slopes in a karst area, SW China. Geoderma 2018, 330, 30–40. [Google Scholar] [CrossRef]
- Zhou, H.Y.; Pan, X.Y.; Zhou, W.Z. Assessing spatial distribution of soil erosion in a karst region in southwestern China: A case study in Jinfo Mountains. In International Conference on Energy Engineering and Environmental Protection; Huang, K.L., Kim, K.W., Liu, J., Eds.; IOP Publishing: Bristol, UK, 2017; Volume 52. [Google Scholar]
- Ma, T.S.; Deng, X.W.; Chen, L.; Xiang, W.H. The soil properties and their effects on plant diversity in different degrees of rocky desertification. Sci. Total Environ. 2020, 736, 1–8. [Google Scholar] [CrossRef]
- Cheng, J.Z.; Lee, X.Q.; Tang, Y.; Pan, W.J.; Gao, W.C.; Chen, Y.; Wang, B.; Cheng, H.G. Changes in above- and below-ground nitrogen stocks and allocations following the conversion of farmland to forest in rocky desertification regions. Agric. Ecosyst. Environ. 2016, 232, 9–16. [Google Scholar] [CrossRef]
- Liu, H.Y.; Liu, F. Recovery Dynamics of Vegetation and Soil Properties in Karst Rocky Desertification Areas in Guizhou, China. In Advances in Environmental Science and Engineering; Parts, 1–6; Iranpour, R., Zhao, J., Wang, A., Yang, F.L., Li, X., Eds.; Trans Tech Publications Ltd.: Zurich, Switzerland, 2012; Volume 518–523, pp. 4532–4544. [Google Scholar]
- Sheng, M.Y.; Xiong, K.N.; Wang, L.J.; Li, X.N.; Li, R.; Tian, X.J. Response of soil physical and chemical properties to Rocky desertification succession in South China Karst. Carbonates Evaporites 2018, 33, 15–28. [Google Scholar] [CrossRef]
- Vazquez, E.; Benito, M.; Espejo, R.; Teutscherova, N. Response of soil properties and microbial indicators to land use change in an acid soil under Mediterranean conditions. Catena 2020, 189, e104486. [Google Scholar] [CrossRef]
- Li, J.; Li, M.; Dong, L.; Wang, K.; Liu, Y.; Hai, X.; Deng, L. Plant productivity and microbial composition drive soil carbon and nitrogen sequestrations following cropland abandonment. Sci. Total Environ. 2020, 744, 1–10. [Google Scholar] [CrossRef]
- Liu, Y.P.; Lu, M.X.; Zhang, X.W.; Sun, Q.B.; Liu, R.L.; Lian, B. Shift of the microbial communities from exposed sandstone rocks to forest soils during pedogenesis. Int. Biodeterior. Biodegrad. 2019, 140, 21–28. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, P.C.; Tian, H.M.; Xiao, Q.Q.; Jiang, H.K. Pyrosequencing-based assessment of soil microbial community structure and analysis of soil properties with vegetable planted at different years under greenhouse conditions. Soil Tillage Res. 2019, 187, 1–10. [Google Scholar] [CrossRef]
- Hu, N.; Li, H.; Tang, Z.; Li, Z.; Tian, J.; Lou, Y.; Hu, X. Community diversity, structure and carbon footprint of nematode food web following reforestation on degraded Karst soil. Sci. Rep. 2016, 6, 28138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, N.; Sun, N.X.; Du, H.M.; Umair, M.; Kang, H.Z.; Liu, X.X.; Romantschuk, M.; Liu, C.J. Karst rocky desertification does not erode ectomycorrhizal fungal species richness but alters microbial community structure. Plant. Soil 2019, 445, 383–396. [Google Scholar] [CrossRef]
- Song, M.; Peng, W.X.; Zeng, F.P.; Du, H.; Peng, Q.; Xu, Q.G.; Chen, L.; Zhang, F. Spatial Patterns and Drivers of Microbial Taxa in a Karst Broadleaf Forest. Front. Microbiol. 2018, 9, 13. [Google Scholar] [CrossRef]
- Yan, Y.J.; Dai, Q.H.; Hu, G.; Jiao, Q.; Mei, L.; Fu, W.B. Effects of vegetation type on the microbial characteristics of the fissure soil-plant systems in karst rocky desertification regions of SW China. Sci. Total Environ. 2020, 712, 11. [Google Scholar] [CrossRef]
- Zhao, C.; Long, J.; Liao, H.K.; Zheng, C.L.; Li, J.; Liu, L.F.; Zhang, M.J. Dynamics of soil microbial communities following vegetation succession in a karst mountain ecosystem, Southwest China. Sci. Rep. 2019, 9, 10. [Google Scholar] [CrossRef]
- Quideau, S.A.; McIntosh, A.C.S.; Norris, C.E.; Lloret, E.; Swallow, M.J.B.; Hannam, K. Extraction and Analysis of Microbial Phospholipid Fatty Acids in Soils. J. Vis. Exp. 2016, 9, e54360. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Wu, L.; Wang, J.; Ma, J.; Zheng, N.; Hill, P.W.; Chadwick, D.R.; Jones, D.L. Fertilizer regime changes the competitive uptake of organic nitrogen by wheat and soil microorganisms: An in-situ uptake test using C-13, N-15 labelling, and C-13-PLFA analysis. Soil Biol. Biochem. 2018, 125, 319–327. [Google Scholar] [CrossRef]
- Pasayat, M. PLFA Profiling of Microbial Community Structure Influencing Ecosystem Restoration in Chronosequence Iron Mine Overburden Spoil. Int. J. Innov. Eng. Technol. 2017, 8, 81–96. [Google Scholar]
- Veum, K.S.; Lorenz, T.; Kremer, R.J. Phospholipid Fatty Acid Profiles of Soils under Variable Handling and Storage Conditions. Agron. J. 2019, 111, 1090–1096. [Google Scholar]
- Zhang, J.D. Quantitative Ecology; Science Press: Beijing, China, 2004; pp. 20–23. (In Chinese) [Google Scholar]
- Guan, H.; Fan, J. Effects of vegetation restoration on soil quality in fragile karst ecosystems of southwest China. PeerJ 2020, 8, e9456. [Google Scholar] [PubMed]
- Ye, C.; Cheng, X.L.; Zhang, Q.F. Recovery approach affects soil quality in the water level fluctuation zone of the Three Gorges Reservoir, China: Implications for revegetation. Environ. Sci. Pollut. Res. 2014, 21, 2018–2031. [Google Scholar]
- Askari, M.S.; Holden, N.M. Indices for quantitative evaluation of soil quality under grassland management. Geoderma 2014, 230, 131–142. [Google Scholar] [CrossRef]
- Andrews, S.S.; Karlen, D.L.; Cambardella, C.A. The soil management assessment framework: A quantitative soil quality evaluation method. Soil Sci. Soc. Am. J. 2004, 68, 1945–1962. [Google Scholar]
- Bastida, F.; Moreno, J.L.; Hernandez, T.; Garcia, C. Microbiological degradation index of soils in a semiarid climate. Soil Biol. Biochem. 2006, 38, 3463–3473. [Google Scholar] [CrossRef]
- Pang, D.B.; Cao, J.H.; Dan, X.Q.; Guan, Y.H.; Peng, X.W.; Cui, M.; Wu, X.Q.; Zhou, J.X. Recovery approach affects soil quality in fragile karst ecosystems of southwest China: Implications for vegetation restoration. Ecol. Eng. 2018, 123, 151–160. [Google Scholar] [CrossRef]
- Wang, Y.; Dungait, J.A.J.; Xing, K.; Green, S.M.; Hartley, I.; Tu, C.; Quine, T.A.; Tian, J.; Kuzyakov, Y. Persistence of soil microbial function at the rock-soil interface in degraded karst topsoils. Land Degrad. Dev. 2019, 31, 251–265. [Google Scholar]
- Jatoi, M.T.; Lan, G.Y.; Wu, Z.X.; Sun, R.; Yang, C.; Tan, Z.H. Comparison of Soil Microbial Composition and Diversity Between Mixed and Monoculture Rubber Plantations in Hainan Province, China. Trop. Conserv. Sci. 2019, 12, 1–9. [Google Scholar]
- Ma, L.; Guo, C.; Lü, X.; Yuan, S.; Wang, R. Soil moisture and land use are major determinants of soil microbial community composition and biomass at a regional scale in northeastern China. Biogeosciences 2015, 12, 2585–2596. [Google Scholar]
- Waldrop, M.P.; Holloway, J.M.; Smith, D.B.; Goldhaber, M.B.; Drenovsky, R.E.; Scow, K.M.; Dick, R.; Howard, D.; Wylie, B.; Grace, J.B. The interacting roles of climate, soils, and plant production on soil microbial communities at a continental scale. Ecology 2017, 98, 1957–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhalnina, K.; Dias, R.; de Quadros, P.D.; Davis-Richardson, A.; Camargo, F.A.O.; Clark, I.M.; McGrath, S.P.; Hirsch, P.R.; Triplett, E.W. Soil pH Determines Microbial Diversity and Composition in the Park Grass Experiment. Microb. Ecol. 2015, 69, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Holden, S.R.; Berhe, A.A.; Treseder, K.K. Decreases in soil moisture and organic matter quality suppress microbial decomposition following a boreal forest fire. Soil Biol. Biochem. 2015, 87, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Waring, B.G.; Hawkes, C.V. Short-Term Precipitation Exclusion Alters Microbial Responses to Soil Moisture in a Wet Tropical Forest. Microb. Ecol. 2015, 69, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, H.; Tuo, D.F.; Wang, C.; Fu, B.J.; Lv, Y.H.; Liu, G.H. Land use change and stand age regulate soil respiration by influencing soil substrate supply and microbial community. Geoderma 2020, 359, 9. [Google Scholar] [CrossRef]
- Ren, C.; Zhao, F.; Shi, Z.; Chen, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Differential responses of soil microbial biomass and carbon-degrading enzyme activities to altered precipitation. Soil Biol. Biochem. 2017, 115, 1–10. [Google Scholar] [CrossRef]
- Evans, S.E.; Wallenstein, M.D. Climate change alters ecological strategies of soil bacteria. Ecol. Lett. 2014, 17, 155–164. [Google Scholar] [CrossRef]
- Yu, S.F.; She, G.H.; Ye, S.M.; Zhou, X.G.; Yao, X.Y.; Li, Y.F. Characteristics of soil microbial biomass and community composition in Pinus yunnanensis var. Tenuifolia secondary forests. J. Sustain. For. 2018, 37, 753–770. [Google Scholar] [CrossRef]
- Peng, X.D.; Dai, Q.H.; Ding, G.J.; Li, C.L. Role of underground leakage in soil, water and nutrient loss from a rock-mantled slope in the karst rocky desertification area. J. Hydrol. 2019, 578, 10. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, A.; Yang, W.; Xu, Z.; Wu, F.; Tan, B.; Liu, Y.; Chen, L. Soil microbial abundance and community structure vary with altitude and season in the coniferous forests, China. J. Soils Sediments 2016, 17, 2318–2328. [Google Scholar] [CrossRef]
- Preusser, S.; Poll, C.; Marhan, S.; Angst, G.; Mueller, C.W.; Bachmann, J.; Kandeler, E. Fungi and bacteria respond differently to changing environmental conditions within a soil profile. Soil Biol. Biochem. 2019, 137, 12. [Google Scholar] [CrossRef]
- Yan, N.; Marschner, P.; Cao, W.; Zuo, C.; Qin, W. Influence of salinity and water content on soil microorganisms. Int. Soil Water Conserv. Res. 2015, 3, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Preusser, S.; Marhan, S.; Poll, C.; Kandeler, E. Microbial community response to changes in substrate availability and habitat conditions in a reciprocal subsoil transfer experiment. Soil Biol. Biochem. 2017, 105, 138–152. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Vo, D.; Graham, K.J.; Scow, K.M. Soil water content and organic carbon availability are major determinants of soil microbial community composition. Microb. Ecol. 2004, 48, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.Z.; Lu, S.Y.; Liu, S.; Guo, H.; Wang, T.; Zhou, J.X.; Peng, X.W. Changes in Plant Rhizosphere Microbial Communities under Different Vegetation Restoration Patterns in Karst and Non-karst Ecosystems. Sci. Rep. 2019, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.S.; Hu, B.; Qi, K.B.; Chen, W.J.; Pang, X.Y.; Bao, W.K.; Tian, G.L. Effects of phosphorus addition on soil microbial biomass and community composition in a subalpine spruce plantation. Eur. J. Soil Biol. 2016, 72, 35–41. [Google Scholar] [CrossRef]
- Bonner, M.T.L.; Herbohn, J.; Gregorio, N.; Pasa, A.; Avela, M.S.; Solano, C.; Moreno, M.O.M.; Almendras-Ferraren, A.; Wills, J.; Shoo, L.P.; et al. Soil organic carbon recovery in tropical tree plantations may depend on restoration of soil microbial composition and function. Geoderma 2019, 353, 70–80. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Ding, J.-Z.; Peng, Y.-F.; Li, F.; Yang, G.-B.; Liu, L.; Qin, S.-Q.; Fang, K.; Yang, Y.-H. Patterns and drivers of soil microbial communities in Tibetan alpine and global terrestrial ecosystems. J. Biogeogr. 2016, 43, 2027–2039. [Google Scholar] [CrossRef]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Du, Y.; Niu, W.; Zhang, Q.; Cui, B.; Gu, X.; Guo, L.; Liang, B. Effects of Nitrogen on Soil Microbial Abundance, Enzyme Activity, and Nitrogen Use Efficiency in Greenhouse Celery under Aerated Irrigation. Soil Sci. Soc. Am. J. 2018, 82, 606–613. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.X.; Zhu, Y.F.; Song, M.; Du, H.; Song, T.Q.; Zeng, F.P.; Zhang, F.; Wang, K.L.; Luo, Y.Q.; Lan, X.; et al. The spatial distribution and drivers of soil microbial richness and diversity in a karst broadleaf forest. For. Ecol. Manag. 2019, 449, 8. [Google Scholar] [CrossRef]
- Xing, Y.W.; Li, C.Y.; Liu, J.; Wang, Y.; Jing, L.J.; Wang, C.R.; Xue, Y.L.; Dang, T.H. Effects of long-term fertilization on soil microbial abundance in farmland of the Loess Plateau, China. J. Appl. Ecol. 2019, 30, 1351–1358. [Google Scholar]
- Bickel, S.; Or, D. Climate and soil properties affect diversity and abundance of soil microbial life across scales--from grains to biomes. Egu Gen. Assem. Conf. Abstr. 2018, 20, 8486. [Google Scholar]
- Orwin, K.H.; Dickie, I.A.; Holdaway, R.; Wood, J.R. A comparison of the ability of PLFA and 16S rRNA gene metabarcoding to resolve soil community change and predict ecosystem functions. Soil Biol. Biochem. 2018, 117, 27–35. [Google Scholar] [CrossRef]
- Blagodatskaya, E.V.; Blagodatsky, S.A.; Anderson, T.H.; Kuzyakov, Y. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies. Appl. Soil Ecol. 2007, 37, 95–105. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Steenwerth, K.L.; Jackson, L.E.; Scow, K.M. Land use and climatic factors structure regional patterns in soil microbial communities. Glob. Ecol. Biogeogr. 2010, 19, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- De Vries, F.T.; Shade, A. Controls on soil microbial community stability under climate change. Front. Microbiol 2013, 4, 265. [Google Scholar] [CrossRef] [Green Version]
- Ananyeva, N.D.; Castaldi, S.; Stolnikova, E.V.; Kudeyarov, V.N.; Valentini, R. Fungi-to-bacteria ratio in soils of European Russia. Arch. Agron. Soil Sci. 2015, 61, 427–446. [Google Scholar] [CrossRef]
- Jing, Y.L.; Wang, Y.; Liu, S.R.; Zhang, X.D.; Wang, Q.K.; Liu, K.; Yin, Y.; Deng, J.F. Interactive effects of soil warming, throughfall reduction, and root exclusion on soil microbial community and residues in warm-temperate oak forests. Appl. Soil Ecol. 2019, 142, 52–58. [Google Scholar] [CrossRef]
- Li, Q.; Bai, H.H.; Liang, W.J.; Xia, J.Y.; Wan, S.Q.; van der Putten, W.H. Nitrogen Addition and Warming Independently Influence the Belowground Micro-Food Web in a Temperate Steppe. PLoS ONE 2013, 8, e60441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Q.L.; Pan, K.W.; Zhang, L.; Wang, Y.J.; Li, W.; He, X.J.; Luo, H.Y. Warming and nitrogen deposition are interactive in shaping surface soil microbial communities near the alpine timberline zone on the eastern Qinghai-Tibet Plateau, southwestern China. Appl. Soil Ecol. 2016, 101, 72–83. [Google Scholar] [CrossRef]
- Glassman, S.I.; Weihe, C.; Li, J.; Albright, M.B.N.; Looby, C.I.; Martiny, A.C.; Treseder, K.K.; Allison, S.D.; Martiny, J.B.H. Decomposition responses to climate depend on microbial community composition. Proc. Natl. Acad. Sci. USA 2018, 115, 11994–11999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peay, K.G.; von Sperber, C.; Cardarelli, E.; Toju, H.; Francis, C.A.; Chadwick, O.A.; Vitousek, P.M. Convergence and contrast in the community structure of Bacteria, Fungi and Archaea along a tropical elevation-climate gradient. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [Green Version]
- Carnovale, D.; Bissett, A.; Thrall, P.H.; Baker, G. Plant genus (Acacia and Eucalyptus) alters soil microbial community structure and relative abundance within revegetated shelterbelts. Appl. Soil Ecol. 2019, 133, 1–11. [Google Scholar] [CrossRef]
- Grosso, F.; Baath, E.; De Nicola, F. Bacterial and fungal growth on different plant litter in Mediterranean soils: Effects of C/N ratio and soil pH. Appl. Soil Ecol. 2016, 108, 1–7. [Google Scholar] [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Jiang, L.; Luo, Y. Trends in soil microbial communities during secondary succession. Soil Biol. Biochem. 2017, 115, 92–99. [Google Scholar] [CrossRef]
- Philippot, L.; Spor, A.; Henault, C.; Bru, D.; Bizouard, F.; Jones, C.M.; Sarr, A.; Maron, P.A. Loss in microbial diversity affects nitrogen cycling in soil. ISME J. 2013, 7, 1609–1619. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C: N: P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Bolscher, T.; Agren, G.I.; Herrmann, A.M. Land-use alters the temperature response of microbial carbon-use efficiency in soils—A consumption-based approach. Soil Biol. Biochem. 2020, 140, 107639. [Google Scholar] [CrossRef]
- Kieft, T.L.; Wilch, E.; Oconnor, K.; Ringelberg, D.B.; White, D.C. Survival and phospholipid fatty acid profiles of surface and subsurface bacteria in natural sediment microcosms. Appl. Environ. Microbiol. 1997, 63, 1531–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, V.J.; Miller, R.M.; Jastrow, J.D.; Matamala, R.; Zak, D.R. Changes in soil microbial community structure in a tallgrass prairie chronosequence. Soil Sci. Soc. Am. J. 2005, 69, 1412–1421. [Google Scholar] [CrossRef] [Green Version]
- Bossio, D.A.; Scow, K.M. Impacts of carbon and flooding on soil microbial communities: Phospholipid fatty acid profiles and substrate utilization patterns. Microb. Ecol. 1998, 35, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, B.; Zhao, W.; Wang, L.; Xie, D.; Huo, W.; Wu, Y.; Zhang, J. Comparative effects of sulfuric and nitric acid rain on litter decomposition and soil microbial community in subtropical plantation of Yangtze River Delta region. Sci. Total Environ. 2017, 601, 669–678. [Google Scholar] [CrossRef]
- Romaniuk, R.; Costantini, A.; Giuffre, L.; Nannipieri, P. Catabolic response and phospholipid fatty acid profiles as microbial tools to assess soil functioning. Soil Use Manag. 2016, 32, 603–612. [Google Scholar] [CrossRef]
- Sun, D.; Bi, Q.; Li, K.; Dai, P.; Yu, Y.; Zhou, W.; Lv, T.; Liu, X.; Zhu, J.; Zhang, Q.; et al. Significance of temperature and water availability for soil phosphorus transformation and microbial community composition as affected by fertilizer sources. Biol. Fertil. Soils 2018, 54, 229–241. [Google Scholar] [CrossRef]
- Wu, L.; Wu, H.; Chen, J.; Wang, J.; Lin, W. Microbial community structure and its temporal changes in Rehmannia glutinosa rhizospheric soils monocultured for different years. Eur. J. Soil Biol. 2016, 72, 1–5. [Google Scholar] [CrossRef]
- Carrasco, L.; Gattinger, A.; Fliessbach, A.; Roldan, A.; Schloter, M.; Caravaca, F. Estimation by PLFA of Microbial Community Structure Associated with the Rhizosphere of Lygeum spartum and Piptatherum miliaceum Growing in Semiarid Mine Tailings. Microb. Ecol. 2010, 60, 265–271. [Google Scholar] [CrossRef]
- Romaniuk, R.; Giuffre, L.; Costantini, A.; Nannipieri, P. Assessment of soil microbial diversity measurements as indicators of soil functioning in organic and conventional horticulture systems. Ecol. Indic. 2011, 11, 1345–1353. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomarkers | PLFAs in This Study |
|---|---|
| General bacterial | 11:0; 12:0; 13:0; 14:0; 15:0; 16:0; 17:0; 18:0; 20:0; 16:0 DMA; 17:0 DMA; 18:0 DMA; 18:1 ω7c DMA; 16:1ω7c DMA; 16:1ω9c DMA; 16:2 DMA; 18:2 DMA; cy19:0 9, 10 DMA |
| Gram–negative bacteria | 12:1ω8c; 13:1ω3c; 15:1ω7c; 15:1ω6c; 15:0 DMA; 16:0 N alcohol; 2OH 16:0; 16:1ω9c; 16:1ω7c; 16:1ω5c; 17:1ω8c; 17:1ω4c; 17:0 cyclo ω7c; 18:1ω7c; 18:1ω6c; 18:1ω5c; 19:0 cyclo ω7c; 19:1ω8c; 20:1ω8c; 20:1ω9c; 21:1ω3c; 21:1ω5c; 21:1ω8c; 21:1ω9c; 21:1ω6c; 22:1ω3c; 22:1ω8c; 24:1ω7c |
| Gram–positive bacteria | a11:0; a13:0; a14:0; a15:0; a16:0; a17:0; a19:0; i13:0; i14:0; i15:0; i16:0; i17:0; i18:0; i19:0; i20:0; i22:0; 15:1 iso ω9c; 15:1 iso ω6c |
| Actinomycetes | 10Me 16:0; 10Me 17:0; 10Me18:0; 10Me 17:1ω7c; 10Me18:1ω7c |
| Fungi | 18:2ω6 |
| Eucaryote | 15:4ω3c; 18:3ω6c; 19:3ω6c; 19:3ω3c; 19:4ω6c; 20:3ω6c; 20:4ω6c; 20:5ω3c; 21:3ω6c; 22:2ω6c; 22:5ω6c; 22:6ω3c; 22:5ω3c; 23:1ω4c; 24:3ω3c |
| Soil Indices | Soil Depth | Significance | Plant Indices | Soil Depth | Significance | ||
|---|---|---|---|---|---|---|---|
| ≥100 cm | <100 cm | ≥100 cm | <100 cm | ||||
| SOC (g/kg) | 20.78 ± 1.56 | 38.49 ± 2.81 | ** | S | 9.43 ± 0.67 | 7.69 ± 0.59 | |
| TN (g/kg) | 2.34 ± 0.11 | 3.83 ± 0.22 | ** | H | 1.97 ± 0.14 | 1.86 ± 0.16 | |
| TP (g/kg) | 0.68 ± 0.06 | 1.10 ± 0.11 | ** | D | 1.09 ± 0.08 | 1.12 ± 0.10 | |
| C:N | 8.67 ± 0.26 | 9.77 ± 0.26 | J | 1.21 ± 0.10 | 1.23 ± 0.12 | ||
| C:P | 33.65 ± 2.41 | 42.70 ± 4.92 | ** | Woody biomass (kg/m2) | 23.89 ± 6.78 | 3.31 ± 0.73 | ** |
| N:P | 3.88 ± 0.22 | 4.16 ± 0.40 | ** | Herb biomass (kg/m2) | 0.13 ± 0.02 | 0.14 ± 0.03 | |
| Ts (°C) | 23.60 ± 0.49 | 24.81 ± 0.55 | Litter (kg/m2) | 0.32 ± 0.06 | 0.20 ± 0.04 | ||
| pH | 6.07 ± 0.18 | 7.45 ± 0.12 | ** | Leaf C (g/kg) | 465.93 ± 5.77 | 440.01 ± 7.61 | |
| SWC (%) | 30.68 ± 1.18 | 34.66 ± 1.94 | * | Leaf N (g/kg) | 15.97 ± 1.40 | 17.67 ± 1.43 | |
| BD (g/cm3) | 1.27 ± 0.02 | 1.16 ± 0.03 | Leaf P (g/kg) | 1.76 ± 0.13 | 1.70 ± 0.14 | ||
| NH4+ (mg/kg) | 12.90 ± 0.77 | 22.02 ± 2.63 | ** | Leaf C:N | 34.87 ± 2.19 | 32.43 ± 3.40 | ** |
| NO3− (mg/kg) | 4.17 ± 1.13 | 5.95 ± 1.06 | Leaf C:P | 335.47 ± 33.60 | 337.32 ± 36.83 | ||
| AP (mg/kg) | 6.01 ± 1.09 | 12.61 ± 2.87 | ** | Leaf N:P | 9.95 ± 0.70 | 11.59 ± 1.16 | |
| AK (mg/kg) | 122.64 ± 10.90 | 203.92 ± 9.38 | Litter C (g/kg) | 422.58 ± 9.56 | 414.17 ± 6.70 | * | |
| TK (g/kg) | 16.67 ± 1.16 | 16.25 ± 1.81 | Litter N (g/kg) | 10.68 ± 0.81 | 11.76 ± 0.89 | ||
| Ca (g/kg) | 5.44 ± 1.12 | 26.13 ± 4.42 | ** | Litter P (g/kg) | 0.91 ± 0.08 | 1.17 ± 0.20 | |
| Mg (g/kg) | 6.78 ± 0.68 | 10.90 ± 1.28 | Litter C:N | 44.67 ± 2.79 | 39.98 ± 2.47 | ||
| Sand (%) | 39.29 ± 1.44 | 53.25 ± 1.39 | Litter C:P | 582.08 ± 58.27 | 596.51 ± 86.68 | ||
| Silt (%) | 59.33 ± 1.38 | 45.33 ± 1.37 | Litter N:P | 12.54 ± 0.53 | 13.92 ± 1.30 | ** | |
| Clay (%) | 1.38 ± 0.17 | 1.45 ± 0.15 | Litter K (g/kg) | 3.55 ± 0.32 | 3.15 ± 0.26 | * | |
| Litter Ca (g/kg) | 15.89 ± 1.13 | 21.97 ± 1.91 | ** | ||||
| Litter Mg (g/kg) | 2.78 ± 0.23 | 4.35 ± 0.77 | ** | ||||
| Leaf K (g/kg) | 10.66 ± 0.84 | 13.91 ± 1.40 | |||||
| Leaf Ca (g/kg) | 12.02 ± 0.89 | 21.46 ± 1.98 | ** | ||||
| Leaf Mg (g/kg) | 3.12 ± 0.17 | 3.96 ± 0.35 | ** | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, H.; Fan, J.; Zhang, H.; Harris, W. Comparison of Drivers of Soil Microbial Communities Developed in Karst Ecosystems with Shallow and Deep Soil Depths . Agronomy 2021, 11, 173. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010173
Guan H, Fan J, Zhang H, Harris W. Comparison of Drivers of Soil Microbial Communities Developed in Karst Ecosystems with Shallow and Deep Soil Depths . Agronomy. 2021; 11(1):173. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010173
Chicago/Turabian StyleGuan, Huiling, Jiangwen Fan, Haiyan Zhang, and Warwick Harris. 2021. "Comparison of Drivers of Soil Microbial Communities Developed in Karst Ecosystems with Shallow and Deep Soil Depths " Agronomy 11, no. 1: 173. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010173