Competition for Light Interception in Cotton Populations of Different Densities

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Data Collection
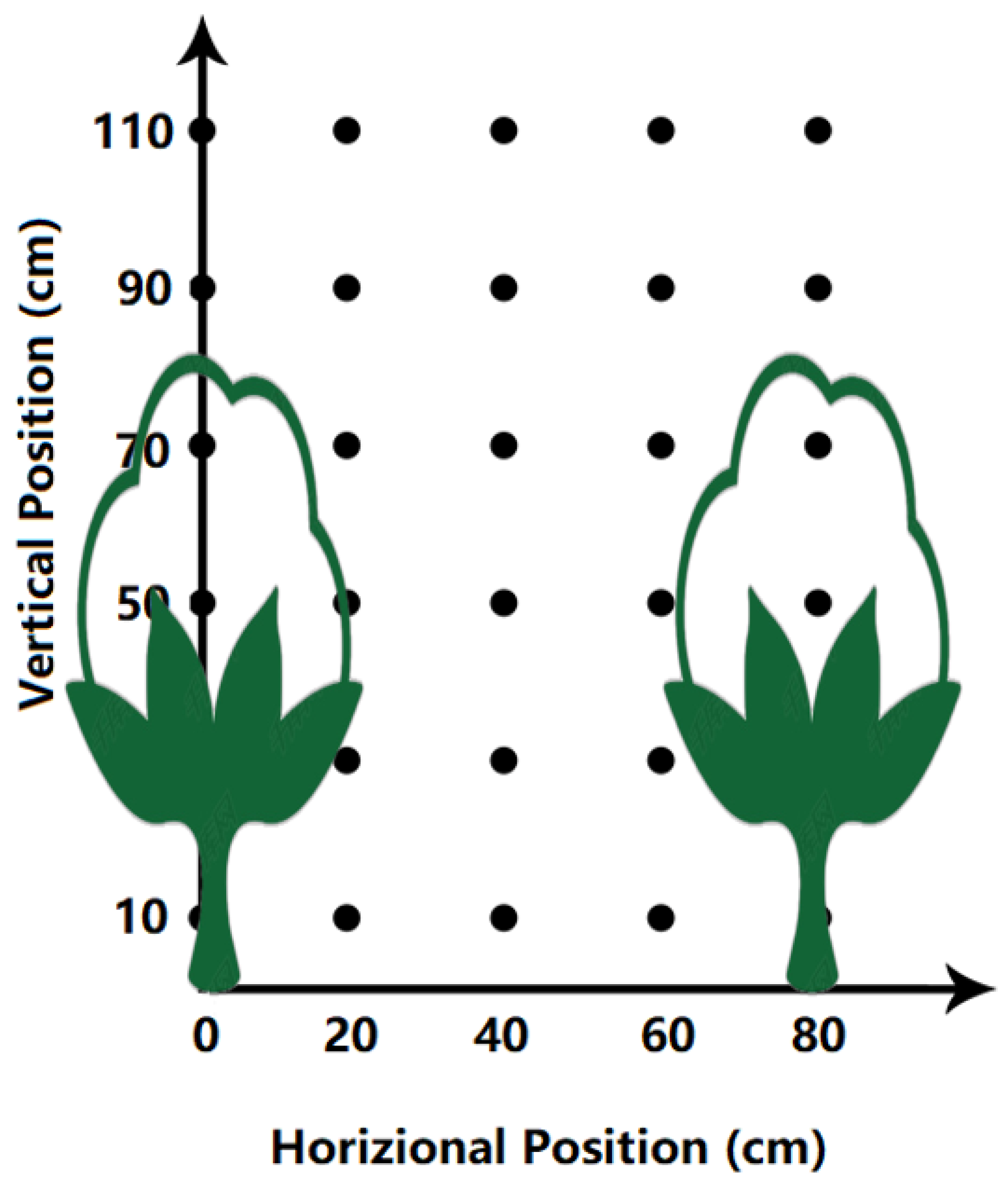
2.3.1. Light Interception
2.3.2. Leaf Area
2.3.3. Plant Height
2.3.4. Biomass
2.3.5. Yield
2.3.6. Statistical Analysis
3. Results
3.1. Light Interception
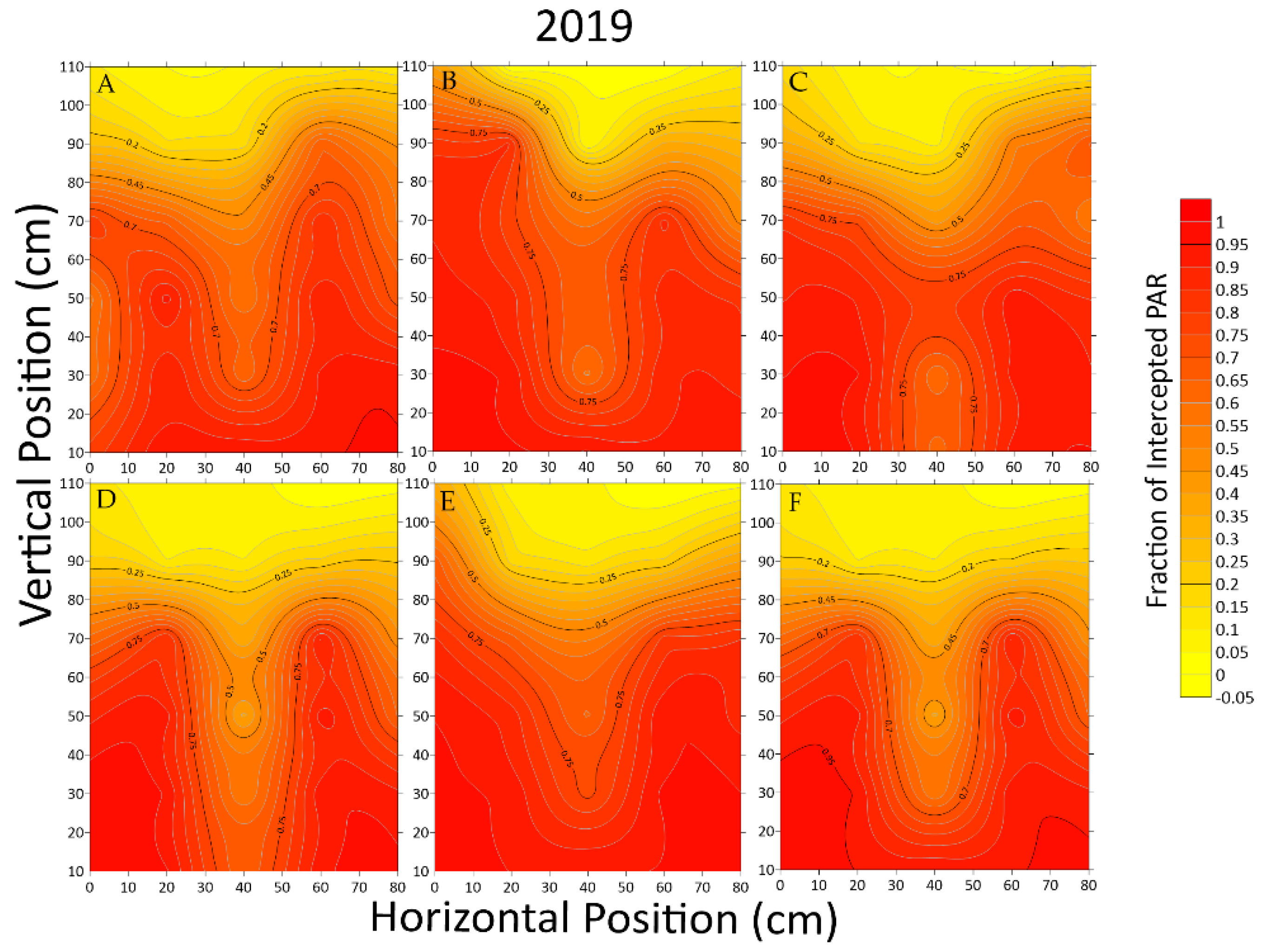
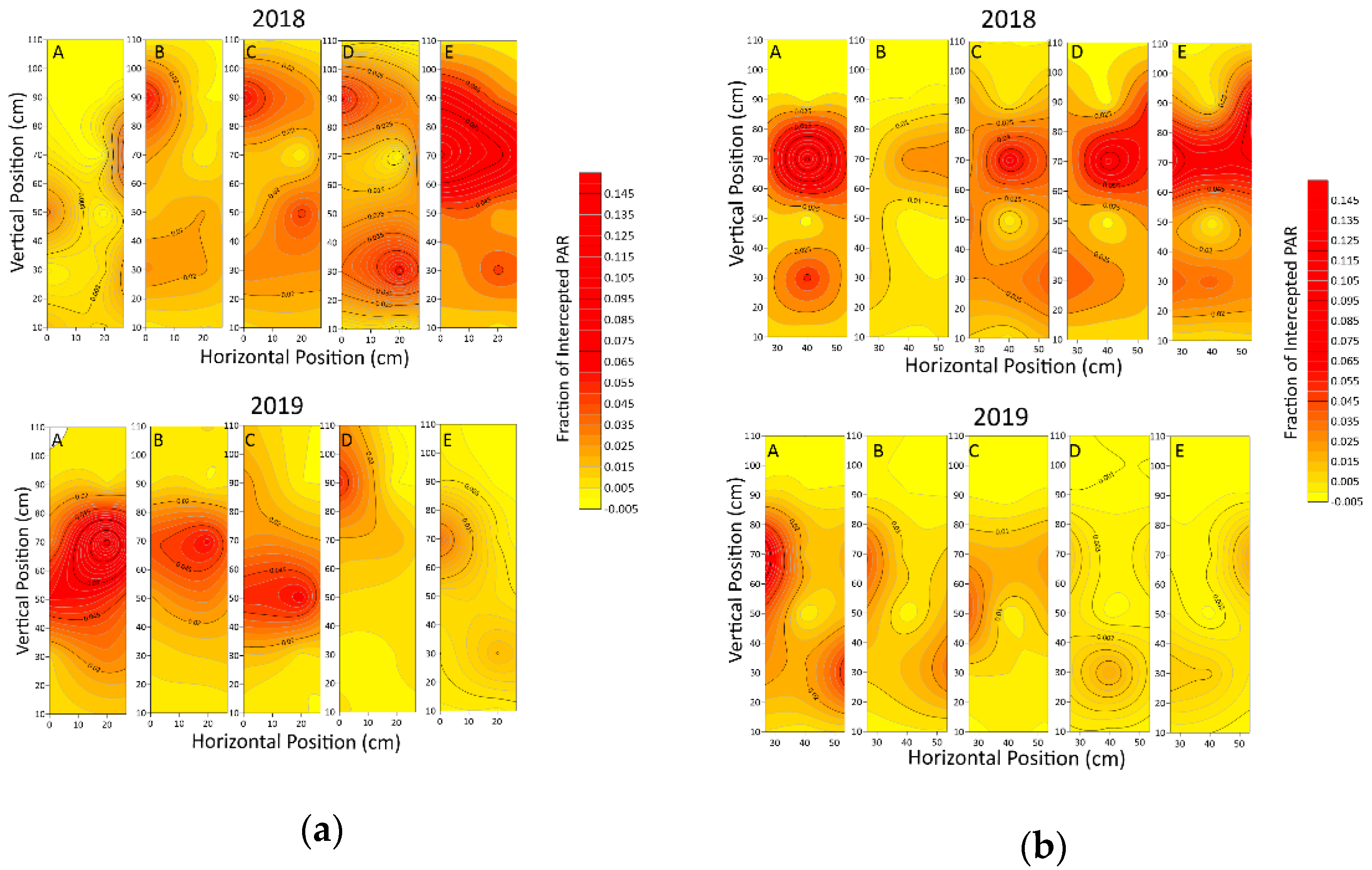
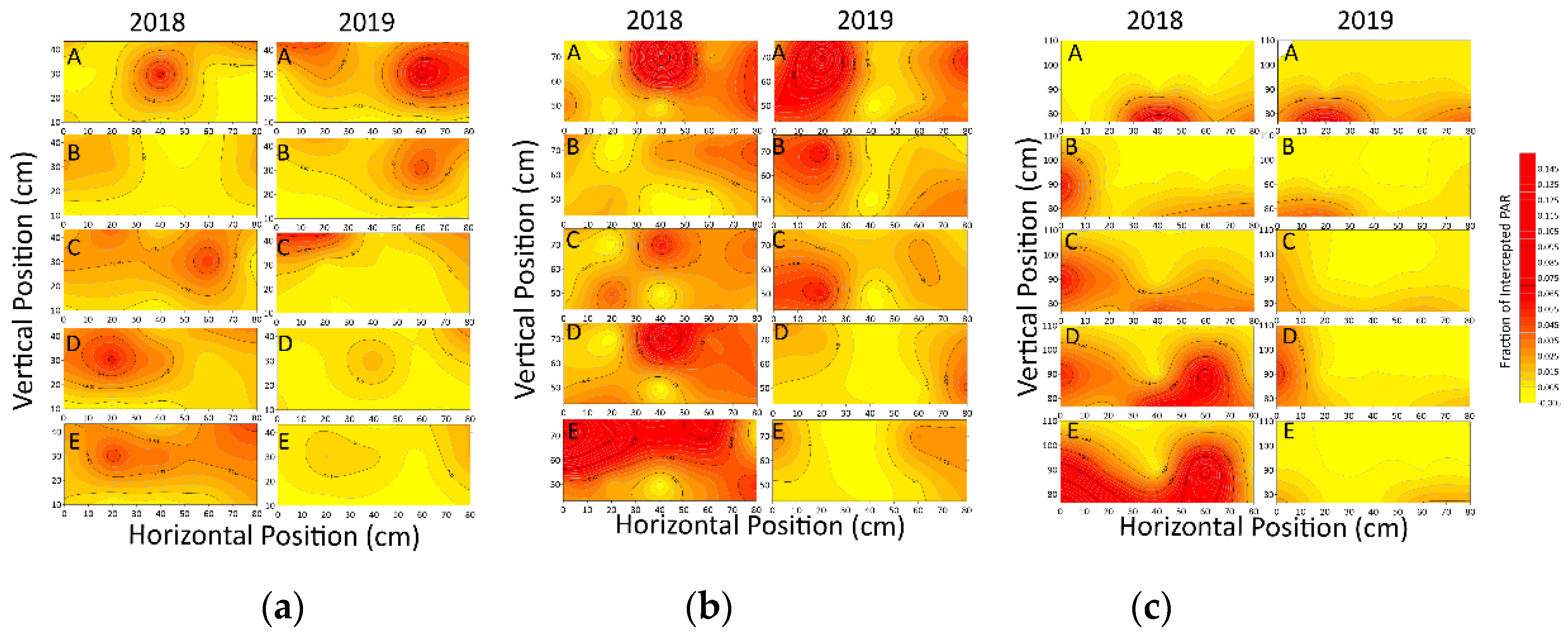
3.1.1. Light Interception at Different Points
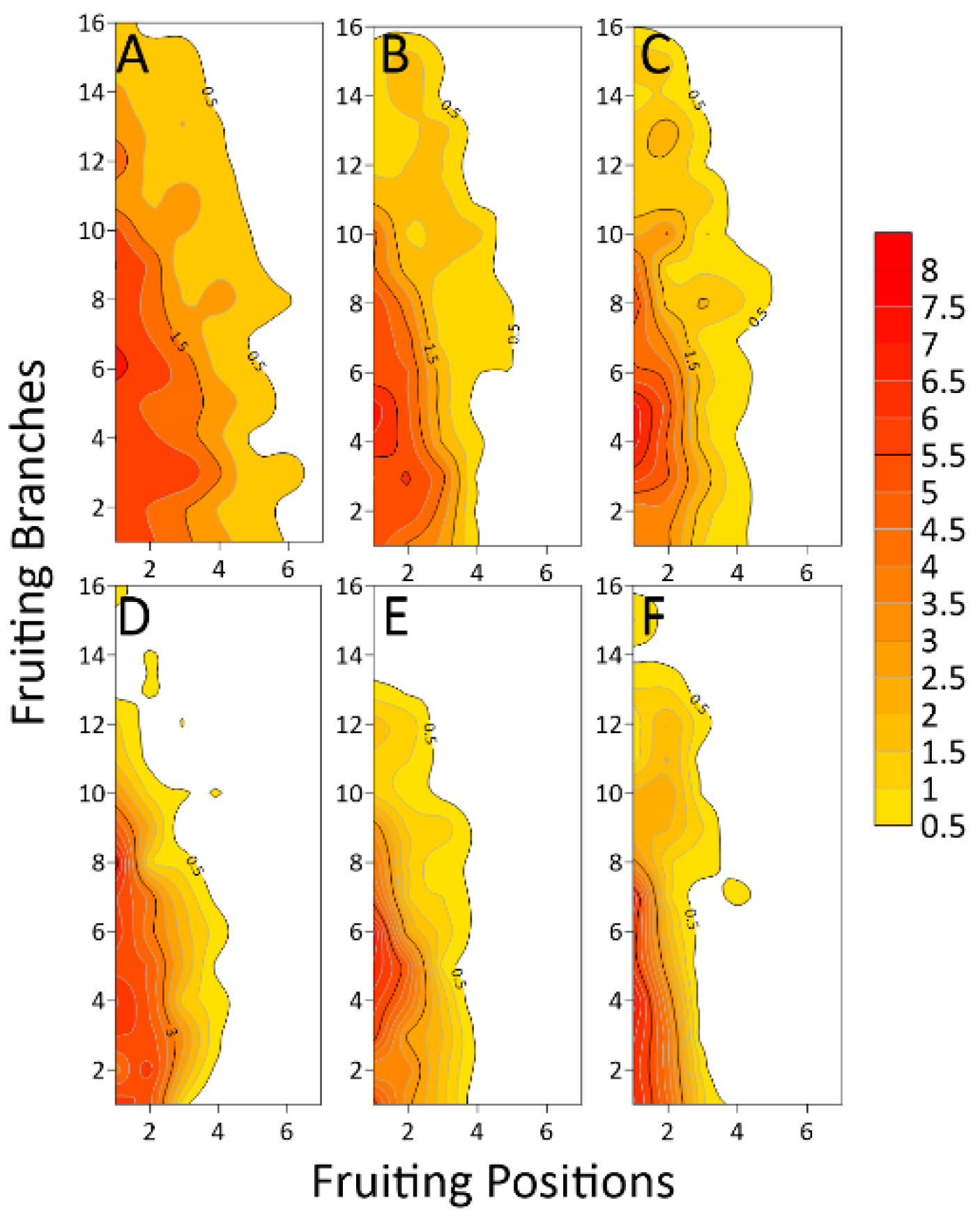
3.1.2. Variation of Light Interception in Different Positions
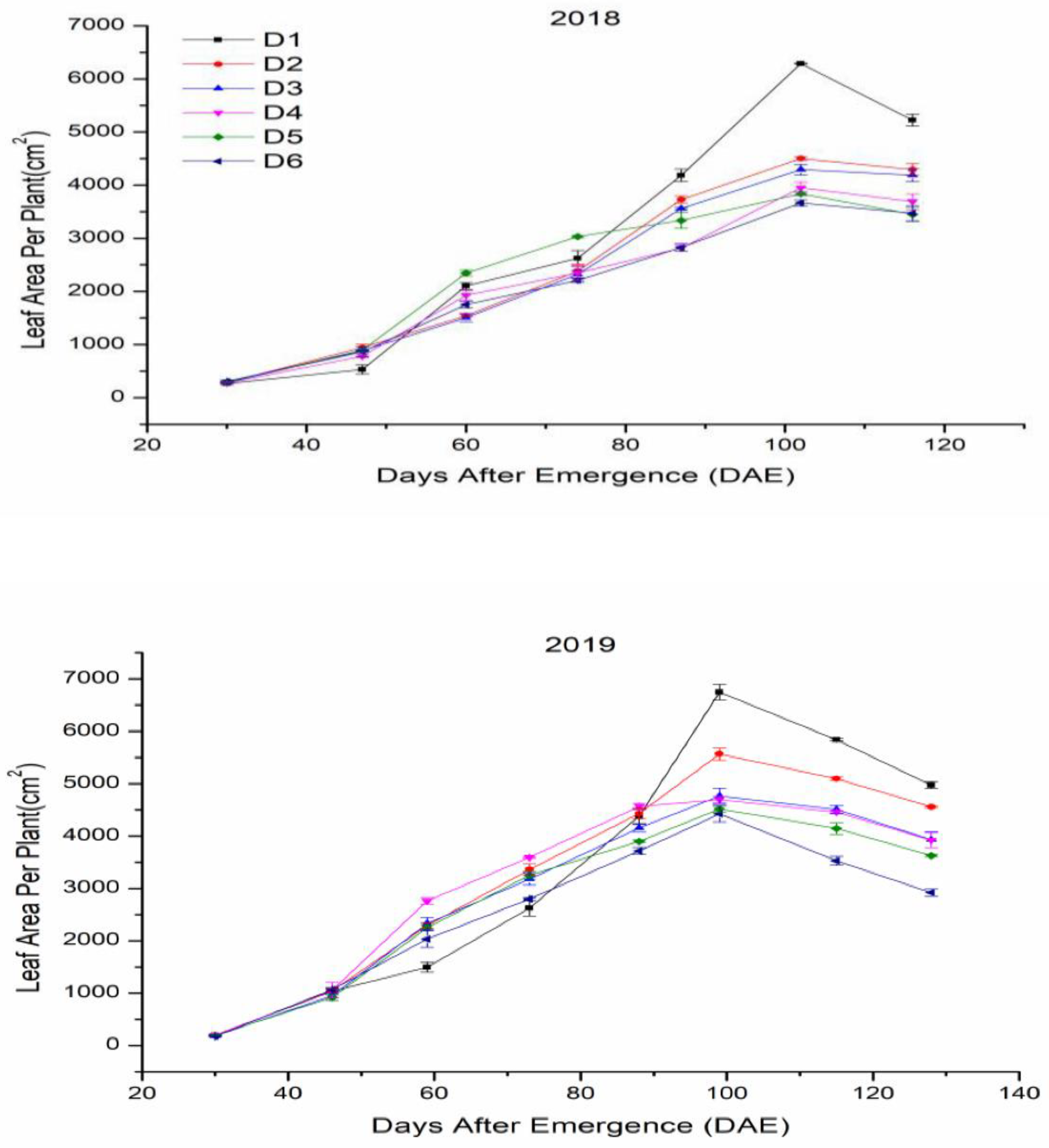
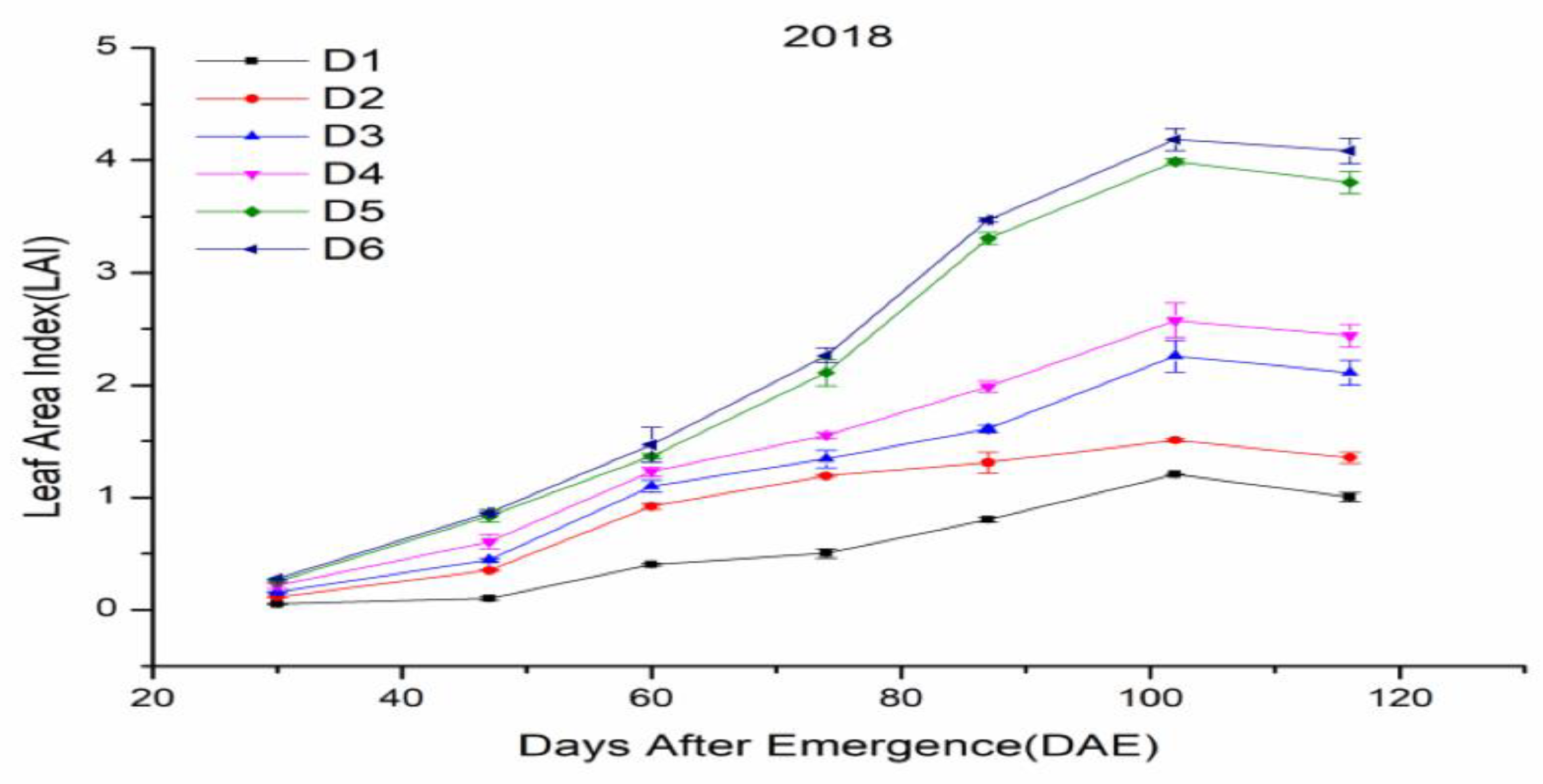
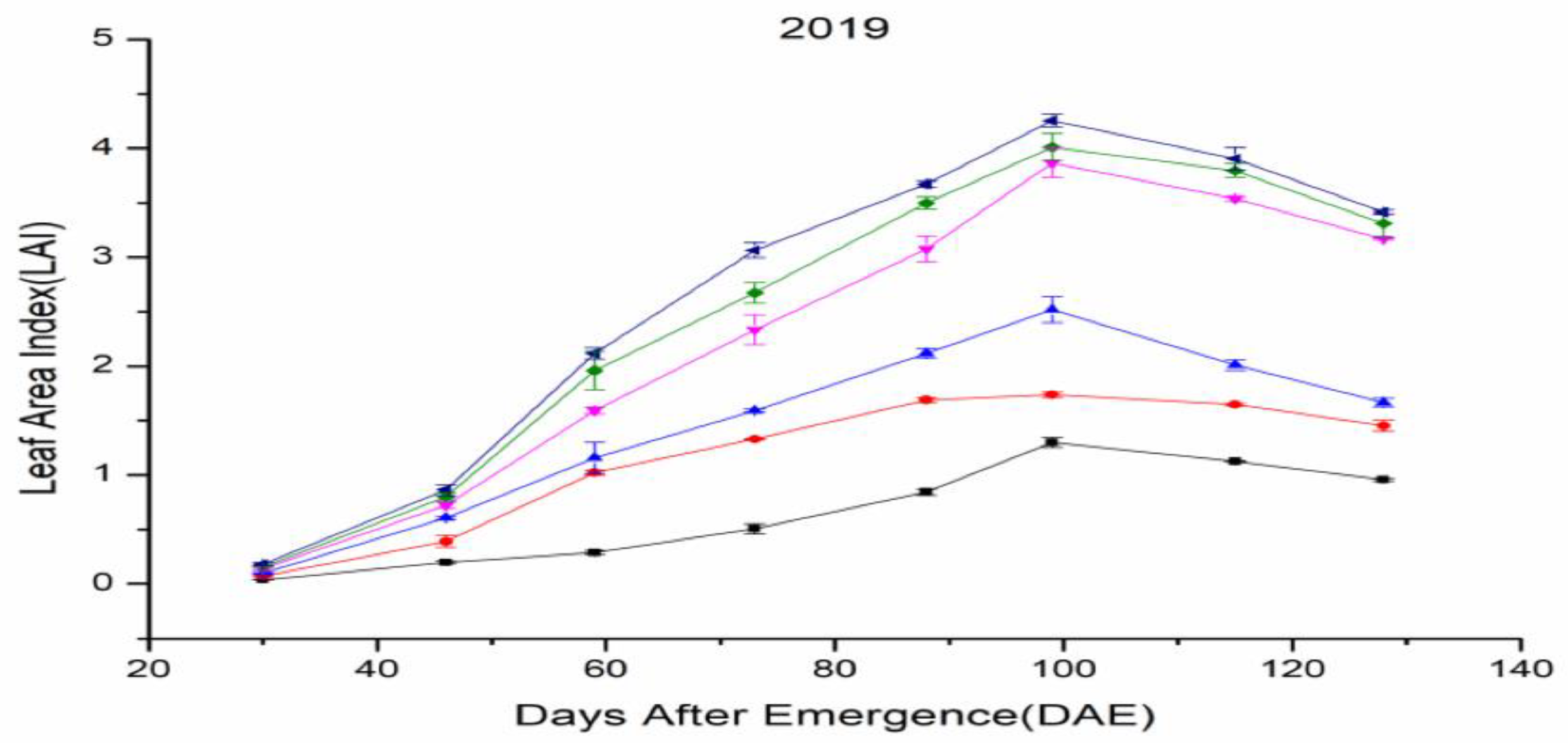
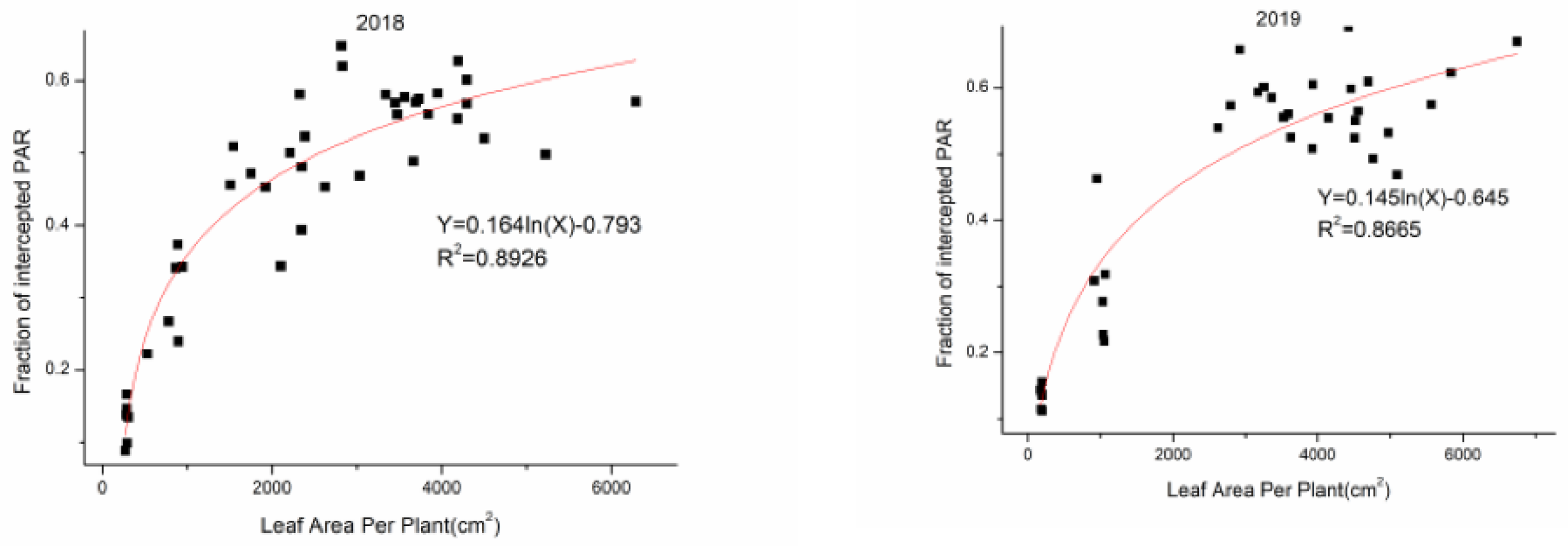
3.2. Leaf Area
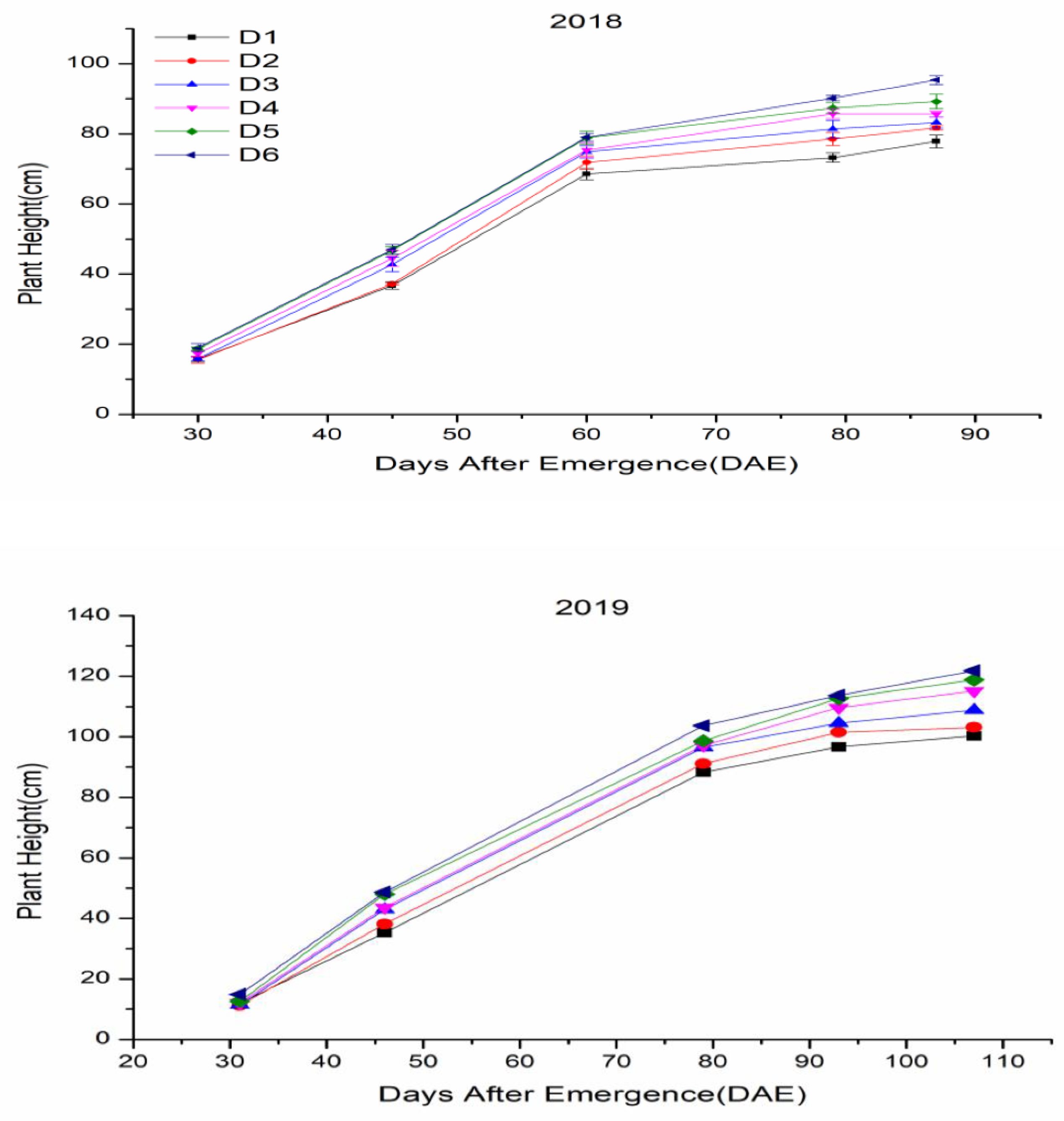
3.3. Plant Height
3.4. Biomass
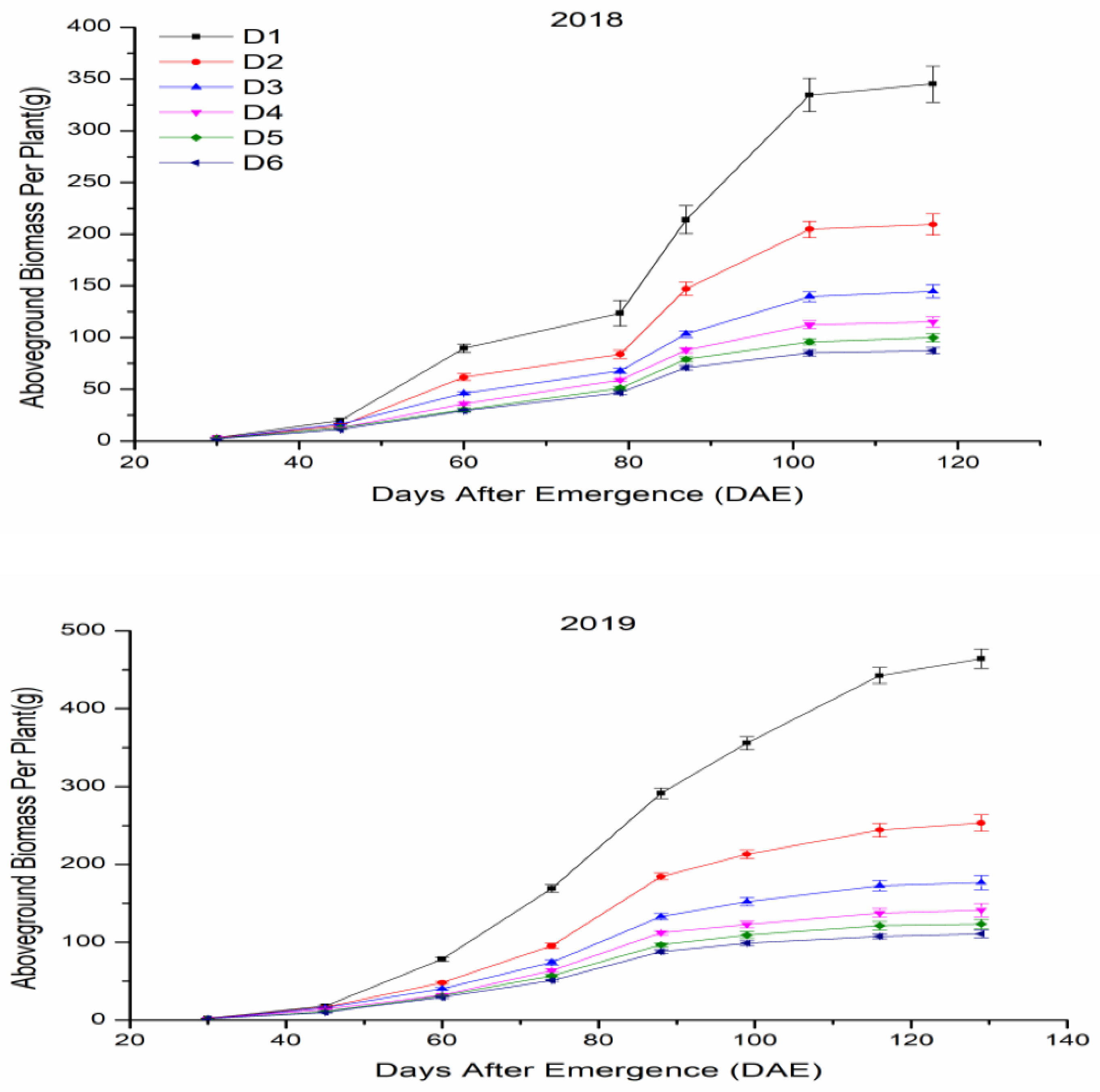
3.4.1. Aboveground Biomass of Individual Plants
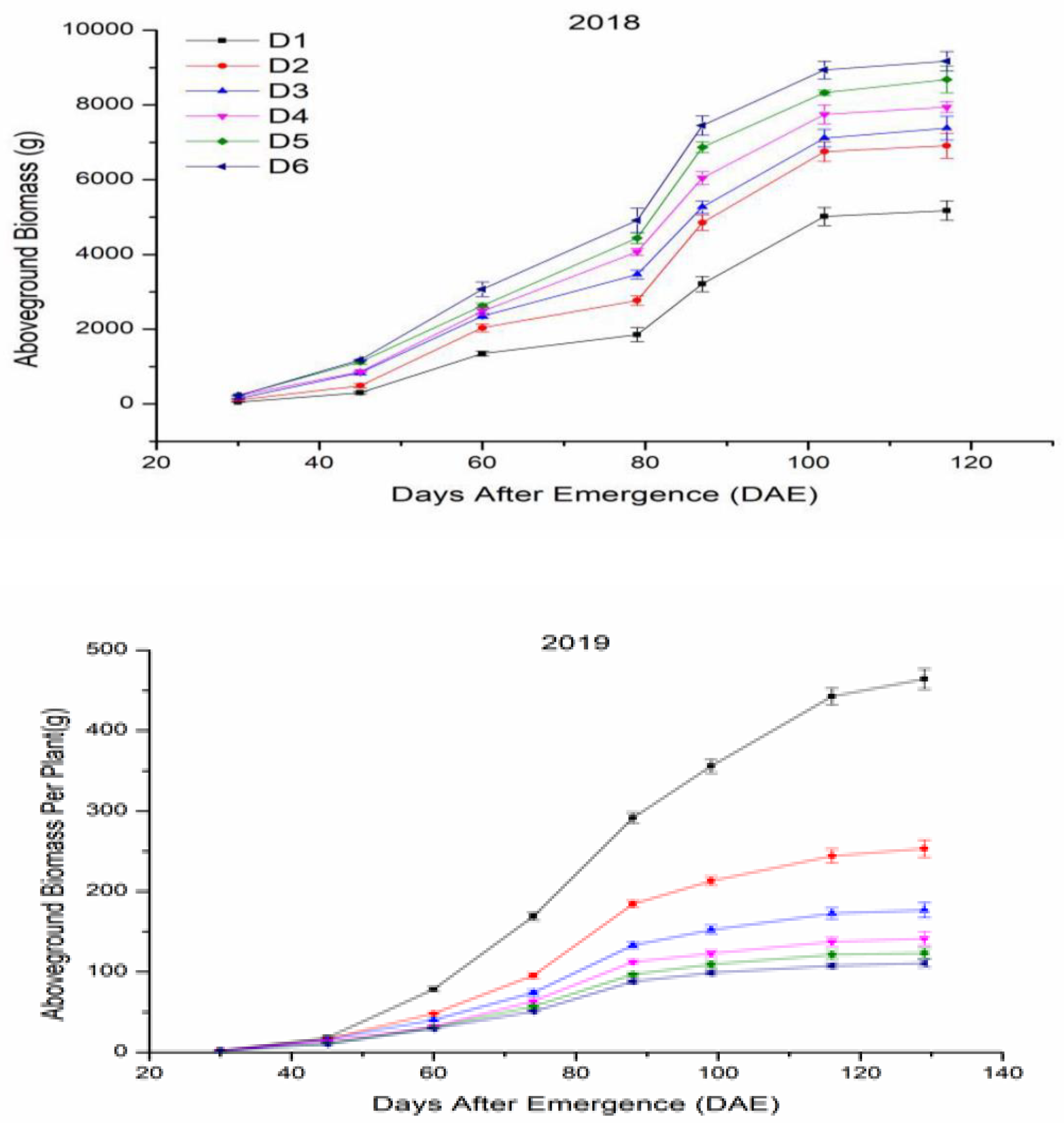
3.4.2. Aboveground Biomass Accumulation
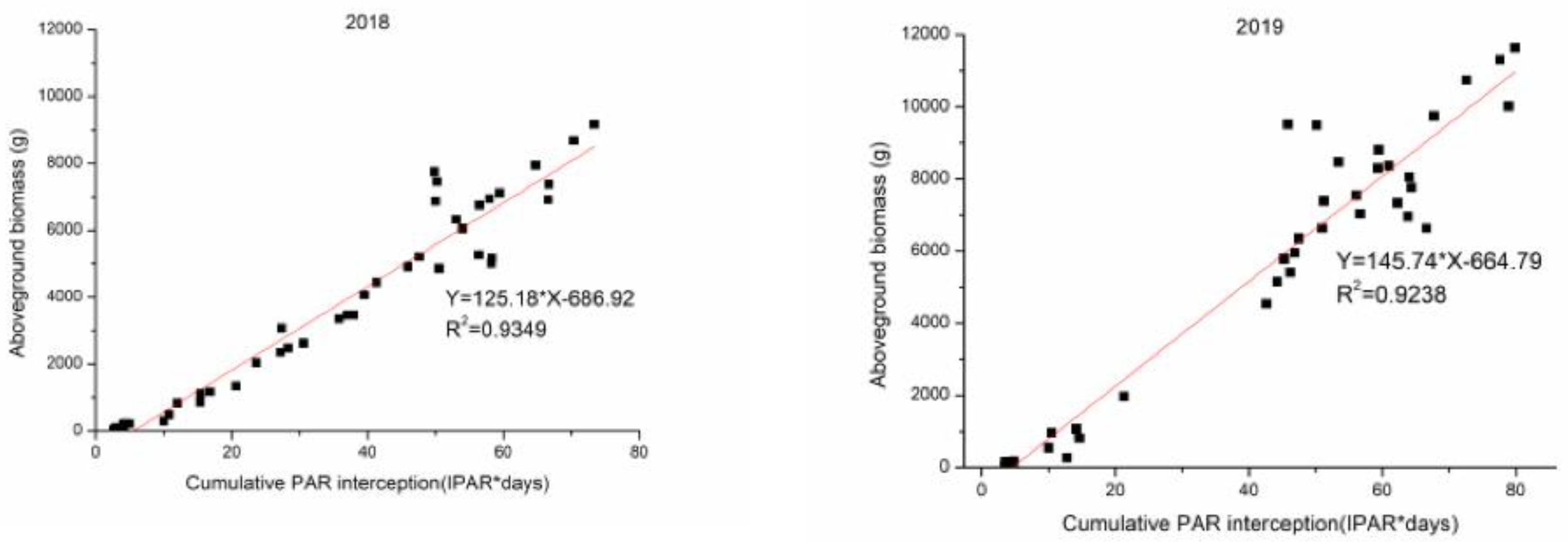
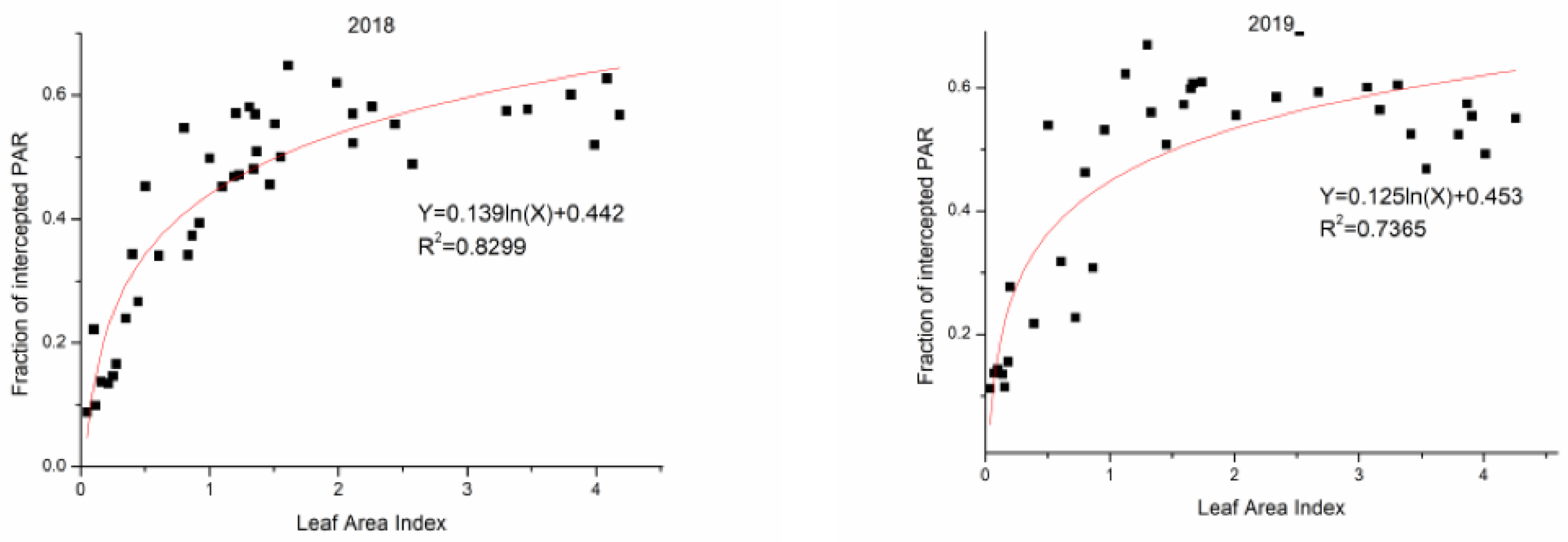
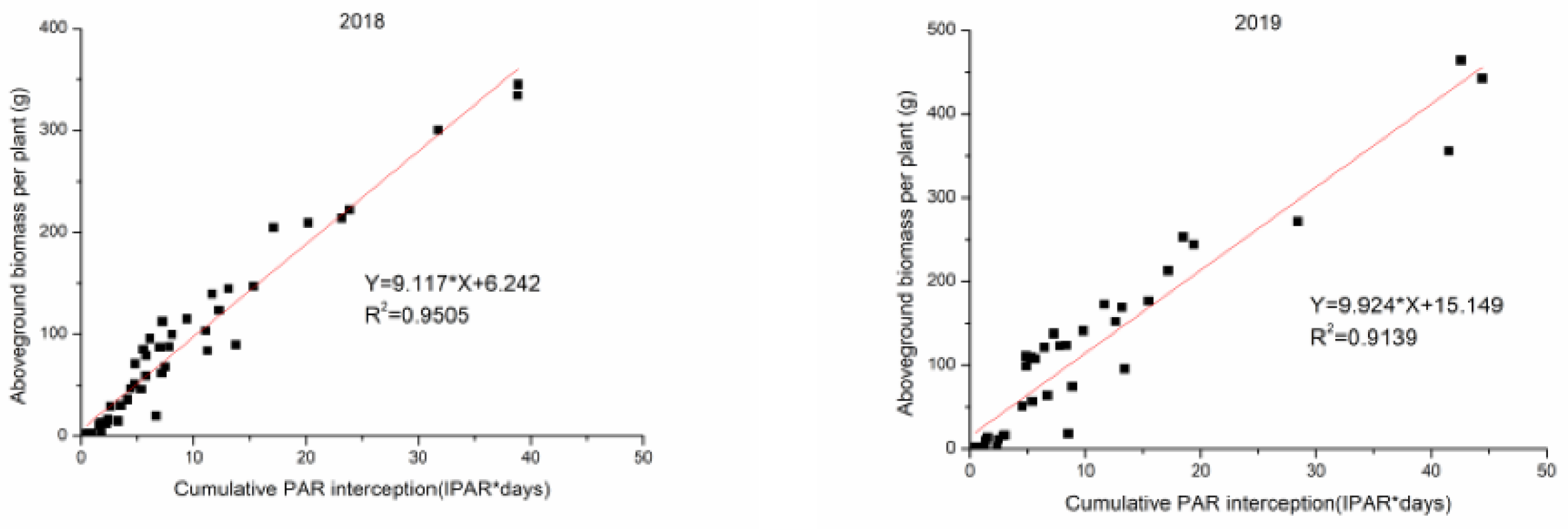
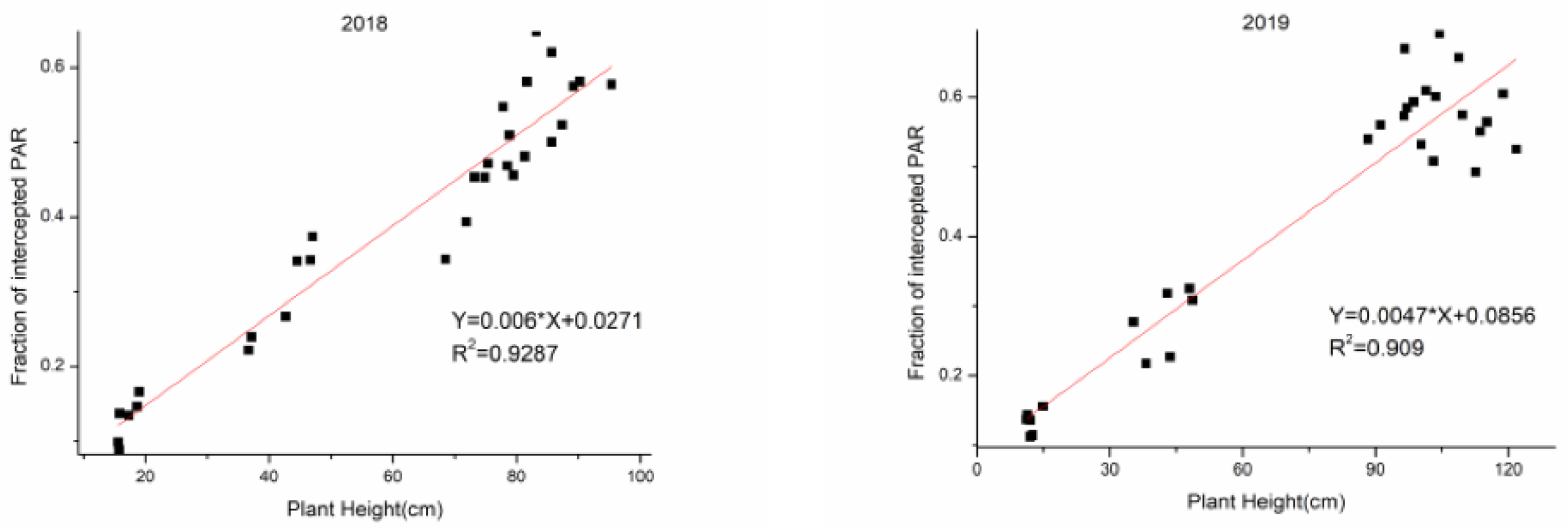
3.5. The Relationships and Fitted Models
3.6. Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, X.; Jia, X.; Niu, J. The present situation and prospects of cotton industry development in China. Sci. Agric. Sin. 2018, 51, 26–36. [Google Scholar]
- Chen, M. Research on issues related to the development of China’s cotton industry--report at the 2014 annual meeting of China cotton association. China Cotton 2014, 41, 38–43. [Google Scholar]
- Mao, S.; Li, Y. Report on China’s Cotton Production Prosperity 2017; China Agriculture Press: Beijing, China, 2018. [Google Scholar]
- Zhang, B.; Bian, X. Evaluation of compresive benefits of smart agriculture in Heilongjiang province based on AHP. Chin. J. Agric. Resour. Reg. Plan 2019, 40, 109–115. [Google Scholar]
- Mao, S. The modernization of cotton cultivation technology in China also focuses on the research related to cotton cultivation in the 12th Five-Year Plan. China Cotton 2010, 37, 2–5. [Google Scholar]
- Dai, J.; Dong, H. Field Crops Research Intensive cotton farming technologies in China: Achievements, challenges and countermeasures. Field Crops Res. 2014, 155, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.; Li, Y.; Dong, H.; Bie, S.; Lin, Y.; Dong, H.; Tian, L.; Chen, D.; Ma, Q.; Ma, X.; et al. Institute of Cotton Research of CAAS. In Cotton Cultivation in China; Shanghai Scientific and Technical Publisher: Shanghai, China, 2013; pp. 123–124. [Google Scholar]
- Zhang, W.; Gou, L.; Li, M. Studies on The Relationship Between Canopy Apparent Photosynthesis Rate and Yield in Cotton in North Xinjiang. Cotton Sci. 1999, 11, 185–190. [Google Scholar]
- McCree, K.J. Photosynthetically Active Radiation. Physiological Plant Ecology I; Springer: Berlin/Heidelberg, Germany, 1981. [Google Scholar]
- McCree, K.J. Test of current definitions of photosynthetically active radiation against leaf photosynthesis data. Agric. Meteorol. 1972, 10, 443–453. [Google Scholar] [CrossRef]
- Gent, M.P.N. Canopy light interception, gas exchange, and biomass in reduced height isolines of winter wheat. Crop. Sci. 1995, 35, 1636–1642. [Google Scholar] [CrossRef]
- Zhang, L.Z.; Van der Werf, W.; Bastiaans, L.; Zhang, S.; Li, B.; Spiertz, J.H.J. Light interception and utilization in relay intercrops of wheat and cotton. Field Crops Res. 2008, 107, 29–42. [Google Scholar] [CrossRef]
- Trouwborst, G.; Oosterkamp, J.; Hogewoning, S.W.; Harbinson, J.; Van Ieperen, W. The responses of light interception, photosynthesis and fruit yield of cucumber to LED-lighting within the canopy. Physiol. Plant. 2010, 138, 289–300. [Google Scholar] [CrossRef]
- Zhi, X.; Han, Y.; Xing, F.; Lei, Y.; Wang, G.; Feng, L.; Yang, B.; Wang, Z.; Li, X.; Xiong, S. How do cotton light interception and carbohydrate partitioning respond to cropping systems including monoculture, intercropping with wheat, and direct-seeding after wheat? PLoS ONE 2019, 14, e0217243. [Google Scholar] [CrossRef] [PubMed]
- Louarn, G.; Lecoeur, J.; Lebon, E. A three-dimensional statistical reconstruction model of grapevine (Vitis vinifera) simulating canopy structure variability within and between cultivar/training system pairs. Ann. Bot. 2008, 101, 1167–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, L.A.; Andersen, M.N.; Jensen, C.R.; Jorgensen, U. Estimation of leaf area index, light interception and biomass accumulation of Miscanthus sinensis ‘Goliath’ from radiation measurements. Biomass Bioenergy 2002, 22, 1–14. [Google Scholar] [CrossRef]
- Heitholt, J.J.; Pettigrew, W.T.; Meredith, W.R. Light Interception and Lint Yield of Narrow-Row Cotton. Crop. Sci. 1992, 32, 728–733. [Google Scholar] [CrossRef]
- Wells, R. Soybean growth response to plant density: Relationships among canopy photosynthesis, leaf area, and light interception. Crop. Sci. 1991, 31, 755–761. [Google Scholar] [CrossRef]
- Goudriaan, J.; Monteith, J.L. A mathematical function for crop growth based on light interception and leaf area expansion. Ann. Bot. 1990, 66, 695–701. [Google Scholar] [CrossRef] [Green Version]
- Maddonni, G.A.; Otegui, M.E. Leaf area, light interception, and crop development in maize. Field Crops Res. 1996, 48, 81–87. [Google Scholar] [CrossRef]
- Du, X.; Chen, B.; Shen, T.; Zhang, Y.; Zhou, Z. Effect of cropping system on radiation use efficiency in double-cropped wheat–cotton. Field Crops Res. 2015, 170, 21–31. [Google Scholar] [CrossRef]
- Chapepa, B.; Mudada, N.; Mapuranga, R. The impact of plant density and spatial arrangement on light interception on cotton crop and seed cotton yield: An overview. J. Cotton Res. 2020, 3, 1–6. [Google Scholar] [CrossRef]
- Bhattacharya, A. Changing Climate and Resource Use Efficiency in Plants; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Zhi, X.; Han, Y.; Mao, S.; Wang, G.; Feng, L.; Yang, B.; Fan, Z.; Du, W.; Lu, J.; Li, Y. Light spatial distribution in the canopy and crop development in cotton. PLoS ONE 2014, 9, e113409. [Google Scholar] [CrossRef]
- Vos, J.; Evers, J.B.; Buck-Sorlin, G.H.; Andrieu, B.; Chelle, M.; de Visser, P.H.B. Functional-structural plant modelling: A new versatile tool in crop science. J. Exp. Bot. 2010, 61, 2101–2115. [Google Scholar] [PubMed]
- Rey, H.; Dauzat, J.; Chenu, K.; Barczi, J.F.; Dosio, G.A.A.; Lecoeur, J. Using a 3-D virtual sunflower to simulate light capture at organ, plant and plot levels: Contribution of organ interception, impact of heliotropism and analysis of genotypic differences. Ann. Bot. 2008, 101, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Evers, J.B.; Zhang, L.; Mao, L.; Zhang, S.; Zhao, X.; Liu, S.; van der Werf, W.; Li, Z. Modelling the structural response of cotton plants to mepiquat chloride and population density. Ann. Bot. 2014, 114, 877–887. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.; Zhang, L.; Evers, J.B.; Henke, M.; van der Werf, W.; Liu, S.; Zhang, S.; Zhao, X.; Wang, B.; Li, Z. Identification of plant configurations maximizing radiation capture in relay strip cotton using a functional–structural plant model. Field Crops Res. 2016, 187, 1–11. [Google Scholar] [CrossRef]
- Tang, Q. Studys on Canopy Photosynthesis Characteristic of Cotton with Different Leaf-Type; Shihezi University: Shihezi, China, 2014. [Google Scholar]
- Khan, N.; Xing, F.; Feng, L.; Wang, Z.; Xin, M.; Xiong, S.; Wang, G.; Chen, H.; Du, W.; Li, Y. Comparative Yield, Fiber Quality and Dry Matter Production of Cotton Planted at Various Densities under Equidistant Row Arrangement. Agronomy 2020, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Xing, F.; Han, Y.; Feng, L.; Zhi, X.; Wang, G.; Yang, B.; Fan, Z.; Lei, Y.; Du, W.; Wang, Z.; et al. Genotypic variation in spatiotemporal distribution of canopy light interception in relation to yield formation in cotton. J. Cotton Res. 2018, 1, 13. [Google Scholar] [CrossRef]
- Monsi, M.; Saeki, T. Über den Lichtfaktor in den Pflanzengesellschaften und seine Bedeutung für die Stoffproduktion. Jpn. J. Bot. 1953, 14, 22–52. [Google Scholar]
- Zhang, W.Y.; Tang, L.; Yang, X.; Liu, L.L.; Cao, W.X.; Zhu, Y. A simulation model for predicting canopy structure and light distribution in wheat. Eur. J. Agron. 2015, 67, 1–11. [Google Scholar] [CrossRef]
- Cao, W.X. Digital Farming Technology; Science Press: Beijing, China, 2018. [Google Scholar]
- Li, Y.; Mao, S.; Wang, X.; Han, Y.; Wang, G.; Fan, Z.; Zhu, Q. Cotton Growth Condition Spatial Variance Analysis Based on Geo-statistics. Cotton Sci. 2007, 19, 214–219. [Google Scholar]
- Liu, Y.; Pan, X. The Analysis on Dominant Provinces of Cotton Production in China. Chin. Agric. Sci. Bull. 2006, 22, 360–364. [Google Scholar]
- Wang, Y.; Cui, X.; Pan, X.; Pan, S. Study on the effect of competition among cotton plants on light energy utilization and growth and development. China Cotton 1999, 08, 3–5. [Google Scholar]
- Li, F. Study on the spatial distribution of cotton bells of different planting densities in SCRC28. Cotton Sci. 2012, 34, 25–27. [Google Scholar]
- Leuchner, M.; Hertel, C.; Menzel, A. Spatial variability of photosyntheticallyactive radiation in European beech and Norway spruce. Agric. For. Meteorol. 2011, 151, 1226–1232. [Google Scholar] [CrossRef]
- Sarlikioti, V.; de Visser, P.H.; Marcelis, L.F. Exploring the spatial distributionof light interception and photosynthesis of canopies by means of afunctional-structural plant model. Ann. Bot. 2011, 107, 875–883. [Google Scholar] [CrossRef] [Green Version]
- Zhi, X.; Han, Y.; Wang, G.; Feng, L.; Yang, B.; Fan, Z.; Du, W.; Lei, Y.; Mao, S.; Li, Y. Changes to the PAR Spatial Distribution, Biomass, and Fiber Quality in Response to Plant Densities. Cotton Sci. 2017, 29, 365–373. [Google Scholar]
- Xue, H.; Han, Y.; Li, Y.; Wang, G.; Feng, L.; Fan, Z.; Du, W.; Yang, B.; Cao, C.; Mao, S. Spatial distribution of light interception by different plant population densities and its relationship with yield. Field Crops Res. 2015, 184, 17–27. [Google Scholar] [CrossRef]
- Han, C.; Wei, S. The study of canopy characteristics of high-yielding groups in rice and their relationship with yield. Liaoning Agric. Sci. 1996, 3, 24–27. [Google Scholar]
- Bai, Z. Study on light interception and biomass production of different cotton cultivars. PLoS ONE 2016, 11, e0156335. [Google Scholar] [CrossRef]
- Shi, W. Effects of Plant Densities in Cotton (Gossypium hirsutum L.) Dry Matter Accumulation and Distribution, Yield and Fiber Quality; Nanjing Agricultural University: Nanjing, China, 2012. [Google Scholar]
- Kiniry, J.R.; Jones, C.A.; O’toole, J.C.; Blanchet, R.; Cabelguenne, M.; Spanel, D.A. Radiation-use efficiency in biomass accumulation prior to grain-filling for five grain-crop species. Field Crops Res. 1989, 20, 51–64. [Google Scholar] [CrossRef]
- Flénet, F.; Kiniry, J.R.; Board, J.E.; Westgate, M.E.; Reicosky, D.C. Rows pacing effects on light extinction coefficients of corn, sorghum soybean and sunflower. Agron. J. 1996, 88, 185–190. [Google Scholar] [CrossRef]
- Kiniry, J.R.; Williams, J.R.; Vanderlip, R.L.; Atwood, J.D.; Reicosky, D.C.; Mulliken, J.; Cox, W.J.; Mascagni, H.J., Jr.; Hollinger, S.E.; Wiebold, W.J. Evaluation of two maize models for nine U.S. locations. Agron. J. 1997, 89, 421–426. [Google Scholar] [CrossRef]
- Connor, D.J.; Fereres, E. A dynamic model of crop growth and partitioning of biomass. Field Crops Res. 1999, 63, 139–157. [Google Scholar] [CrossRef]
- Dai, J.; Li, W.; Tang, W.; Zhang, D.; Li, Z.; Lu, H.; Eneji, A.E.; Dong, H. Manipulation of dry matter accumulation and partitioning with plant density in relation to yield stability of cotton under intensive management. Field Crops Res. 2015, 180, 207–215. [Google Scholar] [CrossRef]
- Francescangeli, N.; Sangiacomo, M.A.; Martí, H. Effects of plant density in broccoli on yield and radiation use efficiency. Sci. Hortic. 2006, 110, 135–143. [Google Scholar] [CrossRef]
- Cheng, G.; Sun, H.; Zhang, Y.; Liu, L.; Wang, M.; Li, C. Effects of canopy structure on photosynthesis and yield in cotton. J. Agric. Univ. Hebei 2015, 38, 1–7. [Google Scholar]
- Lv, X.; Wang, K.; Liu, J.; Chu, G.; Yang, Y.; Dun, J.; Yao, C.; Zhou, F. An Analysis of Characters on Wheat/Corn Compound Canopy Structure. J. Shihezi Univ. (Nat. Sci.) 2001, 5, 120–122. [Google Scholar]
- Khan, N.; Han, Y.; Xing, F.; Feng, L.; Wang, Z.; Wang, G.; Yang, B.; Fan, Z.; Lei, Y.; Xiong, S.; et al. Plant Density Influences Reproductive Growth, Lint Yield and Boll Spatial Distribution of Cotton. Agronomy 2020, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Liu, F. Experimental and Mathematical Model of Rice Canopy Structure and Dry Matter Production under Controlled Irrigation; Hehai University: Nanjing, China, 2005. [Google Scholar]
- Zhang, D. Effect of Cotton Plant Densities on Light Distribution and LUE of Jujube-Cotton Intercropped Systems; Shihezi University: Shihezi, China, 2014. [Google Scholar]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Meteorological Data | Year | April | May | June | July | August | September | October |
|---|---|---|---|---|---|---|---|---|
| Sunshine hours (h) | 2018 | 198.59 | 224.98 | 248.91 | 213.64 | 234.48 | 188.62 | 205.45 |
| 2019 | 297.67 | 256.73 | 260.16 | 186.56 | 213.74 | 139.91 | 99.93 | |
| Daily PAR (μmol·m−2·s−1) | 2018 | 348.58 | 398.81 | 469.74 | 418.51 | 388.82 | 310.04 | 234.28 |
| 2019 | 313.95 | 490.95 | 487.20 | 448.48 | 377.63 | 304.86 | 203.47 |
| Treatment | Emergence/DAE 1 | Seedling/DAE | Squaring/DAE | Flowering/DAE | Boll Opening/DAE |
|---|---|---|---|---|---|
| Year 2018 | |||||
| D1 | April 28/0 | May 17/19 | May 29/31 | June 24/57 | August 6/100 |
| D2 | April 28/0 | May 17/19 | May 30/32 | June 25/58 | August 8/102 |
| D3 | April 28/0 | May 17/19 | May 30/32 | June 25/58 | August 8/102 |
| D4 | April 28/0 | May 17/19 | May 30/32 | June 25/58 | August 10/104 |
| D5 | April 28/0 | May 17/19 | May 30/32 | June 25/58 | August 10/104 |
| D6 | April 28/0 | May 17/19 | May 31/33 | June 27/60 | August 13/107 |
| Year 2019 | |||||
| D1 | April 28/0 | May 20/22 | May 28/30 | June 24/57 | August 14/108 |
| D2 | April 28/0 | May 20/22 | May 28/30 | June 24/57 | August 15/109 |
| D3 | April 28/0 | May 20/22 | May 30/32 | June 24/57 | August 15/109 |
| D4 | April 28/0 | May 20/22 | May 30/32 | June 24/57 | August 16/110 |
| D5 | April 28/0 | May 20/22 | May 31/33 | June 26/59 | August 17/111 |
| D6 | April 28/0 | May 20/22 | May 31/33 | June 26/59 | August 19/112 |
| Treatment | A (gm m−2 d−1) | Fast Accumulation Period | Fastest Accumulation Point | ||||
|---|---|---|---|---|---|---|---|
| t1 (DAE) | t2 (DAE) | Δt (d) | VA (gm m−2 d−1) | VM (gm m−2 d−1) | at DAE | ||
| Year 2018 | |||||||
| D1 | 609.105 | 64.0 | 108.0 | 44.1 | 8.0 | 9.1 | 86.0 |
| D2 | 784.542 | 58.4 | 100.3 | 41.9 | 10.8 | 12.3 | 79.4 |
| D3 | 936.003 | 54.9 | 104.6 | 49.6 | 10.9 | 12.4 | 79.7 |
| D4 | 1025.805 | 54.5 | 101.7 | 47.2 | 12.6 | 14.3 | 78.1 |
| D5 | 1130.505 | 51.3 | 100.3 | 49.0 | 13.3 | 15.2 | 75.8 |
| D6 | 1149.106 | 48.9 | 99.4 | 50.5 | 13.1 | 15.0 | 74.2 |
| Average | 55.3 | 102.4 | 47.0 | 11.5 | 13.1 | 78.9 | |
| Year 2019 | |||||||
| D1 | 716.274 | 64.7 | 100.5 | 35.8 | 11.1 | 13.2 | 82.6 |
| D2 | 843.626 | 63.0 | 94.1 | 31.1 | 15.6 | 17.8 | 78.6 |
| D3 | 918.050 | 60.2 | 93.2 | 33.0 | 16.0 | 18.3 | 76.7 |
| D4 | 982.470 | 59.3 | 90.7 | 31.4 | 18.8 | 20.6 | 75.0 |
| D5 | 1091.457 | 58.6 | 91.0 | 32.4 | 19.3 | 22.2 | 74.8 |
| D6 | 1177.606 | 58.2 | 90.4 | 32.2 | 21.0 | 24.1 | 74.3 |
| Average | 60.7 | 93.3 | 32.6 | 17.0 | 19.4 | 77.0 | |
| Treatment | Regression Equation | R2 |
|---|---|---|
| Year 2018 | ||
| D1 | Y = 6091.05095/(1 + 170.72e−0.05978t) | 0.985 *** |
| D2 | Y = 7845.41809/(1 + 147.37e−0.0629t) | 0.993 *** |
| D3 | Y = 9360.03681/(1 + 68.70e−0.05305t) | 0.992 *** |
| D4 | Y = 10,258.05161/(1 + 78.27e−0.05585t) | 0.991 *** |
| D5 | Y = 11,305.05257/(1 + 59.01e−0.05377t) | 0.989 *** |
| D6 | Y = 11,491.06419/(1 + 47.72e−0.05211t) | 0.989 *** |
| Year 2019 | ||
| D1 | Y = 7162.73984/(1 + 433.56e−0.07351t) | 0.998 *** |
| D2 | Y = 8436.2559/(1 + 772.58e−0.08463t) | 0.997 *** |
| D3 | Y = 9180.49668/(1 + 455.31e−0.07978t) | 0.997 *** |
| D4 | Y = 9824.69778/(1 + 544.36e−0.08399t) | 0.996 *** |
| D5 | Y = 10,914.569/(1 + 439.83e−0.0814t) | 0.997 *** |
| D6 | Y = 11,776.0588/(1 + 436.68e−0.08186t) | 0.996 *** |
| Treatment | Seed Cotton Yield (kg ha−1) | Lint Yield (kg ha−1) | Lint Percentage (%) |
|---|---|---|---|
| Year 2018 | |||
| D1 | 2730.9 ± 96.0d | 1185.5 ± 32.0c | 43.6 ± 0.44a |
| D2 | 3080.5 ± 66.0cd | 1249.8 ± 27.4bc | 41.3 ± 0.58a |
| D3 | 3369.3 ± 84.5bc | 1291.0 ± 23.2bc | 38.4 ± 0.79a |
| D4 | 3679.5 ± 94.9b | 1369.4 ± 33.1b | 37.3 ± 0.28a |
| D5 | 4232.2 ± 116.1a | 1545.3 ± 28.2a | 36.5 ± 0.37a |
| D6 | 3682.2 ± 67.5b | 1316.8 ± 26.8bc | 35.8 ± 0.62a |
| Year 2019 | |||
| D1 | 3299.1 ± 128.2d | 1306.4 ± 41.8d | 39.3 ± 1.07a |
| D2 | 3767.0 ± 68.4c | 1451.9 ± 23.3cd | 38.6 ± 1.15a |
| D3 | 4054.6 ± 20.2b | 1552.7 ± 10.4bc | 39.0 ± 1.10a |
| D4 | 4095.8 ± 72.8ab | 1649.4 ± 28.2ab | 39.1 ± 1.24a |
| D5 | 4351.5 ± 159.0a | 1789.5 ± 32.5a | 39.1 ± 1.31a |
| D6 | 4172.3 ± 105.4ab | 1712.8 ± 17.3ab | 38.7 ± 1.14a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Zhao, X.; Han, Y.; Xing, F.; Feng, L.; Wang, Z.; Wang, G.; Yang, B.; Lei, Y.; Xiong, S.; et al. Competition for Light Interception in Cotton Populations of Different Densities. Agronomy 2021, 11, 176. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010176
Chen H, Zhao X, Han Y, Xing F, Feng L, Wang Z, Wang G, Yang B, Lei Y, Xiong S, et al. Competition for Light Interception in Cotton Populations of Different Densities. Agronomy. 2021; 11(1):176. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010176
Chicago/Turabian StyleChen, Huanxuan, Xinxin Zhao, Yingchun Han, Fangfang Xing, Lu Feng, Zhanbiao Wang, Guoping Wang, Beifang Yang, Yaping Lei, Shiwu Xiong, and et al. 2021. "Competition for Light Interception in Cotton Populations of Different Densities" Agronomy 11, no. 1: 176. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11010176