
Ovicidal and Physiological Effects of Essential Oils Extracted from Six Medicinal Plants on the Elm Leaf Beetle, Xanthogaleruca luteola (Mull.)

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mass Rearing of Insects
2.2. Extraction of Essential Oils
2.3. Assessment of the Ovicidal Effect of Essential Oils
2.4. Preparation of Eggs Homogenates for Biochemical Assay
2.5. Protein Assay
2.6. Determination of Glucose
2.7. Determination of Triglyceride
2.8. Statistical Analysis
3. Results
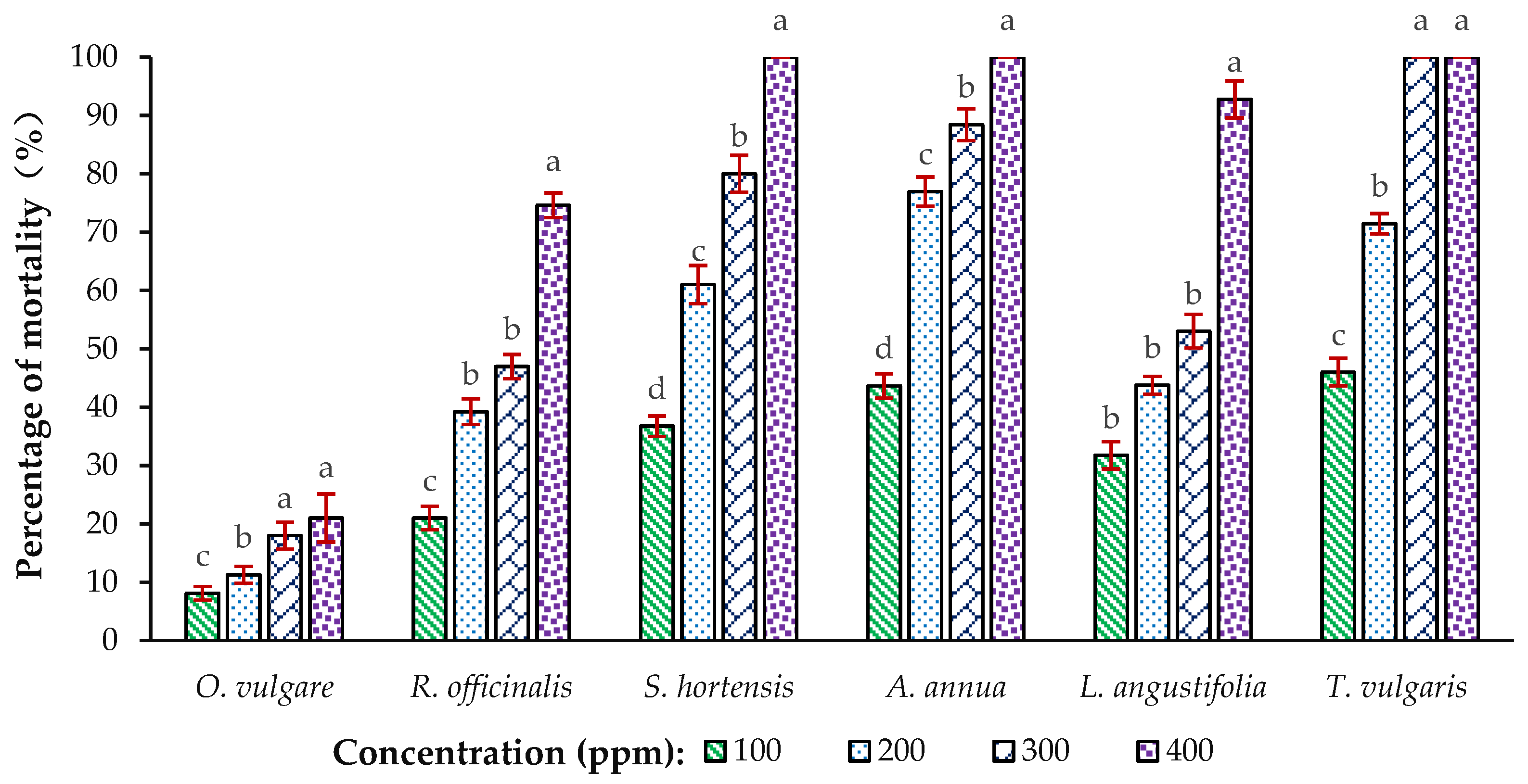
3.1. The Ovicidal Activity of EOs
3.2. Total Protein Level in X. luteola Eggs Post-Exposure to EOs
3.3. Total Glucose of X. luteola Eggs Treated
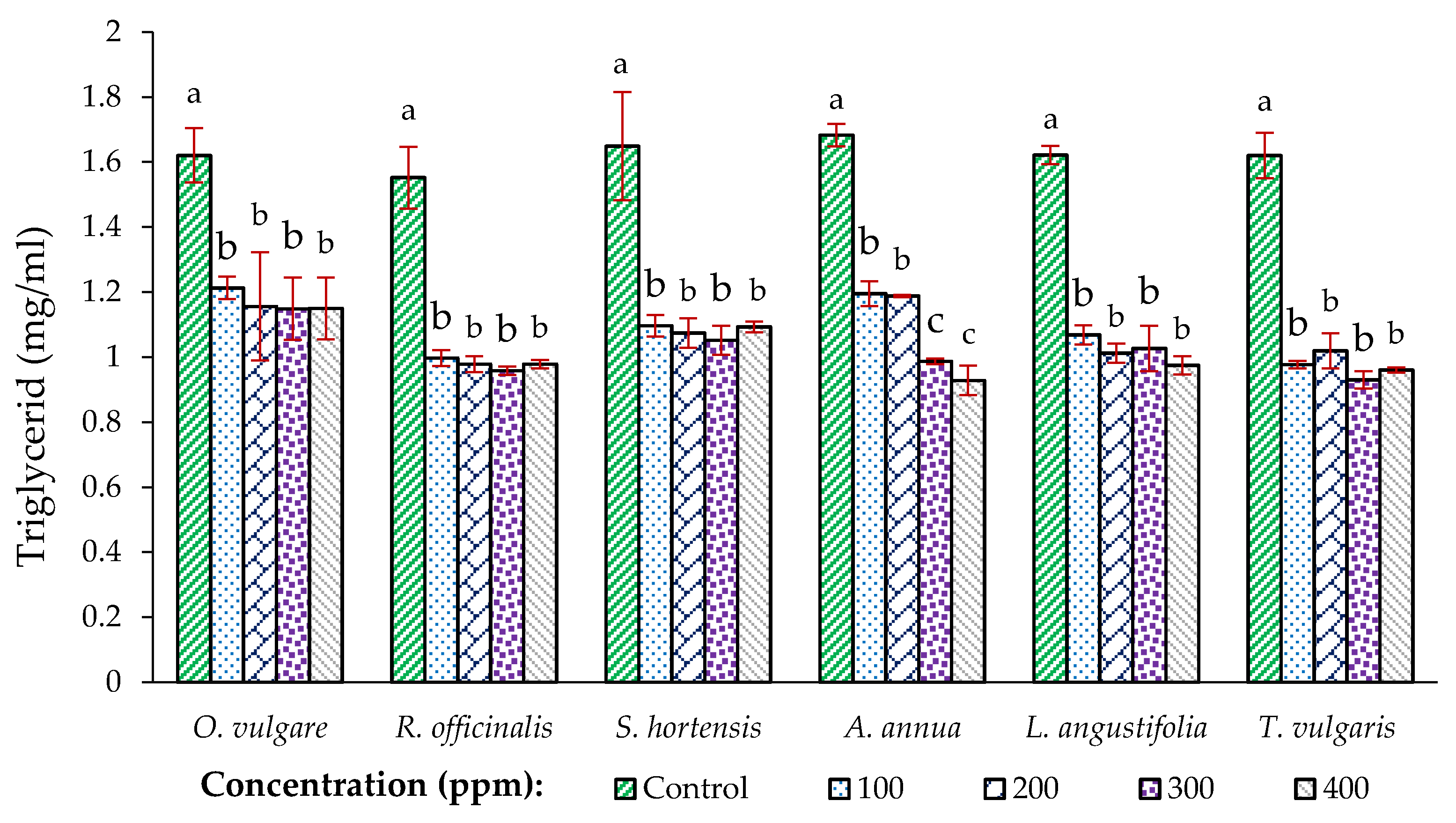
3.4. Total Triglyceride of X. luteola Eggs Treated
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chiffelle, Í.; Huerta, A.; Bobadilla, V.; Macuada, G.; Araya, J.E.; Curkovic, T.; Ceballos, R. Antifeedant and insecticidal effects of extracts from Melia azedarach fruits and Peumus boldus leaves on Xanthogaleruca luteola larvae. Chil. J. Agric. Res. 2019, 79, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo, E.; Bosch, A.M.S.; Xamaní, P.; Laborda, R. Life cycle, parasitism and damage of Xanthogaleruca luteola (Mull.) in Valencia (SE Spain): A preliminary study. Urban. For. Urban. Green 2019, 46, 126474. [Google Scholar] [CrossRef]
- Moore, G.M.; Lefoe, G. The effect of a heat wave on urban tree pests in Melbourne, Australia: Examples that may inform climate change tree management. Arboric Urban For. 2020, 46, 135–147. [Google Scholar] [CrossRef]
- Khater, H.F. Prospects of botanical biopesticides in insect pest management. Pharmacologia 2012, 3, 641–656. [Google Scholar] [CrossRef] [Green Version]
- Pavela, R. Essential oils from Foeniculum vulgare Miller as a safe environmental insecticide against the aphid Myzus persicae Sulzer. Environ. Sci. Pollut. Res. 2018, 25, 10904–10910. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Yao, M.; Fu, L.; Ma, Z.Q.; Zhang, X. The botanical pesticide derived from Sophora flavescens for controlling insect pests can also improve growth and development of tomato plants. Ind. Crop. Prod. 2016, 92, 13–18. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Martínez, L.C.; Rolim, G.D.S.; Coelho, R.P.; Santos, M.H.; Tavares, W.D.S.; Zanuncio, J.C.; Serrão, J.E. Insecticidal and repellent activities of Cymbopogon citratus (Poaceae) essential oil and its terpenoids (citral and geranyl acetate) against Ulomoides dermestoides. Crop. Prot. 2020, 137, 105299. [Google Scholar] [CrossRef]
- Fowsiya, J.; Madhumitha, G.A. Review of bioinsecticidal activity and mode of action of plant derived alkaloids. Res. J. Pharm Technol. 2020, 13, 963–973. [Google Scholar] [CrossRef]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia, L. genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [Green Version]
- Ebadollahi, A.; Ziaee, M.; Palla, F. Essential oils extracted from deferent species of the Lamiaceae plant family as prospective bioagents against several detrimental pests. Molecules 2020, 25, 1556. [Google Scholar] [CrossRef] [Green Version]
- Isman, M.B. Commercial development of plant essential oils and their constituents as active ingredients in bioinsecticides. Phytochem. Rev. 2020, 19, 235–241. [Google Scholar] [CrossRef]
- Domingues, P.M.; Santos, L. Essential oil of pennyroyal (Mentha pulegium): Composition and applications as alternatives to pesticides—New tendencies. Ind. Crops Prod. 2019, 139, 111534. [Google Scholar] [CrossRef]
- Yazdani, E.; Jalali Sendi, J.; Aliakbar, A.; Senthil-Nathan, S. Effect of Satureja hortensis L. essential oil on feeding efficiency and biochemical properties of Glyphodes pyloalis Walker (Lepidoptera: Pyralidae). Arch. Phytopathol. Plant. Protect. 2013, 46, 328–339. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R. Repellence of essential oils and selected compounds against ticks—A systematic review. Acta. Trop. 2018, 179, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Papanastasiou, S.A.; Ioannou, C.S.; Papadopoulos, N.T. Oviposition-deterrent effect of linalool–a compound of citrus essential oils–on female Mediterranean fruit flies, Ceratitis capitata (Diptera: Tephritidae). Pest. Manag. Sci. 2020, 86, 3066–3377. [Google Scholar] [CrossRef]
- Valizadeh, B.; Zibaee, A.; Jalali Sendi, J. Inhibition of digestive α-amylases from Chilo suppressalis Walker (Lepidoptera: Crambidae) by a proteinaceous extract of Citrullus colocynthis L. (Cucurbitaceae). J. Plant. Protec. Res. 2013, 53, 195–202. [Google Scholar] [CrossRef]
- Marimuthu, G.; Rajamohan, S.; Mohan, R.; Krishnamoorthy, Y. Larvicidal and ovicidal properties of leaf and seed extracts of Delonix elata (L.) Gamble (Family: Fabaceae) against malaria (Anopheles stephensi Liston) and dengue (Aedes aegypti Linn.) (Diptera: Culicidae) vector mosquitoes. Parasitol. Res. 2012, 111, 65–77. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticidal, deterrents, and repellents in modern agriculture and increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular targets for components of essential oils in the insect nervous system—A review. Molecules 2017, 23, 34. [Google Scholar] [CrossRef] [Green Version]
- Adibmoradi, G.; Jalali Sendi, J.J.; Tirgari, S.; Imani, S.; Nematolahi, S.R. Insecticidal and morpho-physiological disorders caused by Thymus vulgaris L. essential oil on the elm leaf beetle, Xanthogaleruca luteola Müller (Coleoptera: Chrysomelidae). Arch. Phytopath. Plant. Protect. 2020, 53, 765–780. [Google Scholar] [CrossRef]
- Campbell, B.E.; Pereira, R.M.; Koehler, P.G. Complications with Controlling Insect Eggs: Insecticide Resistance, 1st ed.; Intech: Rijeka, Croatia, 2016; pp. 83–96. [Google Scholar]
- Amri, I.; Hamrouni, L.; Hanana, M.; Jamoussi, B.; Lebdi, K. Essential oils as biological alternatives to protect date palm (Phoenix dactylifera L.) against Ectomyelois ceratoniae Zeller (Lepidoptera: Pyralidae). Chil. J. Agric. Res. 2014, 74, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Naseri, B.; Abedi, Z.; Abdolmaleki, A.; Jafary-Jahed, M.; Borzoui, E.; Mansouri, S.M. Fumigant Toxicity and sublethal effects of Artemisia khorassanica and Artemisia sieberi on sitotroga cerealella (Lepidoptera: Gelechiidae). J. Insect Sci. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Bedini, S.; Flamini, G.; Cosci, F.; Ascrizzi, R.; Echeverria, M.C.; Gomez, E.V.; Guidi, L.; Landi, M.; Lucchi, A.; Conti, B. Toxicity and oviposition deterrence of essential oils of Clinopodium nubigenum and Lavandula angustifolia against the myiasis-inducing blowfly Lucilia sericata. PLoS ONE 2019, 14, e0212576. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Magierowicz, K.; Górska-Drabik, E.; Sempruch, C. The effect of Tanacetum vulgare essential oil and its main components on some ecological and physiological parameters of Acrobasis advenella (Zinck.) (Lepidoptera: Pyralidae). Pestic. Biochem. Physiol. 2020, 162, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Valizadeh, B.; Jalali Sendi, J. Sublethal effects of pyriproxyfen on some biological and biochemical properties of elm leaf beetle, Xanthogaleruca luteola (Col.: Chrysomelidae). J. Entomol. Soc. Iran. 2014, 33, 59–70. [Google Scholar]
- British Pharmacopoeia; HMSO: London, UK, 1988; Volume 2, pp. 137–138.
- Ribeiro, W.L.C.; Camurça-Vasconcelos, A.L.F.; Macedo, I.T.F.; dos Santos, J.M.L.; de Araújo-Filho, J.V.; Ribeiro, J.D.C.; Pereira, V.D.A.; Viana, D.D.A.; de Paula, H.C.B.; Bevilaqua, C.M.L. In vitro effects of Eucalyptus staigeriana nanoemulsion on Haemonchus contortus and toxicity in rodents. Veter. Parasitol. 2015, 212, 444–447. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Siegert, K.J. Carbohydrate metabolism in Manduca sexta during late larval development. J. Insect Physiol. 1987, 33, 421–427. [Google Scholar] [CrossRef]
- Fossati, P.; Prencipe, L. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Khosravi, R.; Jalali Sendi, J.J. Toxicity, development and physiological effect of Thymus vulgaris and Lavandula angustifolia essential oils on Xanthogaleruca luteola (Coleoptera: Chrysomelidae). J. King Saud Univ. Sci. 2013, 25, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Adibmoradi, G.; Sendi, J.J.; Tirgari, S.; Imani, S.; Nematolahi, S.R. Effect of 1,8-cineol on the biology and physiology of elm leaf beetle, Xanthogaleruca luteola (Col.: Chrysomelidae). J. Plant. Protect. Res. 2018, 58, 420–430. [Google Scholar] [CrossRef]
- Shekari, M.; Jalali Sendi, J.; Etebari, K.; Zibaee, A.; Shadparvar, A. Effects of Artemisia annua L. (Asteracea) on nutritional physiology and enzyme activities of elm leaf beetle, Xanthogaleruca luteola Mull. (Coleoptera: Chrysomelidae). Pestic. Biochem. Physiol. 2008, 9, 66–74. [Google Scholar] [CrossRef]
- Krzyżowski, M.; Baran, B.; Łozowski, B.; Francikowski, J. The effect of Rosmarinus officinalis essential oil fumigation on biochemical, behavioral, and physiological parameters of Callosobruchus maculatus. Insects 2020, 11, 344. [Google Scholar] [CrossRef]
- Al-Murmidhi, M.M.A.F.; Al-Hasnawi, M.R.A. The toxicity of phenolic compounds to some plants in the cumulative loss of the adult stages of domestic flies. Musca domestica (Diptera: Muscidae). J. Phys. 2019, 1294, 062009. [Google Scholar] [CrossRef] [Green Version]
- Modarres-Najafabadi, S.S. Control of Tetranychus urticae Koch by thyme, lavender and eucalyptus essential oils. J. Med. Plants By-Prod. 2012, 1, 43–47. [Google Scholar] [CrossRef]
- Gong, X.; Ren, Y. Larvicidal and ovicidal activity of carvacrol, p-cymene, and γ-terpinene from Origanum vulgare essential oil against the cotton bollworm, Helicoverpa armigera (Hübner). Environ. Sci. Pollut. Res. 2020, 27, 18708–18716. [Google Scholar] [CrossRef]
- Cotchakaew, N.; Soonwera, M. Toxicity of several botanical essential oils and their combinations against females of Aedes albopictus (Skuse) and Anopheles minimus (Theobald): Oviposition deterrent, ovicidal and adulticidal efficacies. Asian Pac. J. Trop. Biomed. 2019, 9, 1–29. [Google Scholar] [CrossRef]
- Sarma, R.; Adhikari, K.; Mahanta, S. Twenty essential oils as ovicidal agent against Aedes aegypti (Diptera: Culicidae). Natl. Acad. Sci. Lett. 2020, 43, 497–500. [Google Scholar] [CrossRef]
- Gustafson, J.E.; Liew, Y.C.; Chew, S.; Markham, J.L.; Bell, H.C.; Wyllie, S.G. Effects of tea tree oil on Escherichia coli. Lett. Appl. Microbiol. 1998, 26, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Oussalah, M.; Caillet, S.; Salmieri, S.; Saucier, L.; Lacroix, M. Antimicrobial effects of alginate-based films containing essential oils on Listeria monocytogenes and Salmonella typhimurium present in bologna and ham. J. Food Prot. 2007, 70, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino Acids: Biochemistry and Nutrition, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021; pp. 510–545. [Google Scholar]
- Etebari, K.; Matindoost, L. A study on the effects of larval age on biochemical macromolecules abundance of haemolymph in silkworm Bombyx mori L. (Lepidoptera: Bombycidae). J. Entomol. Soc. Iran. 2004, 24, 1–16. [Google Scholar]
- Oftadeh, M.; Jalali Sendi, J.; Ebadollahi, A.; Setzer, W.N.; Krutmuang, P. Mulberry protection through flowering-stage essential oil of Artemisia annua against the lesser mulberry pyralid, Glyphodes pyloalis Walker. Foods 2021, 10, 210. [Google Scholar] [CrossRef]
- Valková, V.; Ďúranová, H.; Galovičová, L.; Vukovic, N.L.; Vukic, M.; Kačániová, M. In Vitro antimicrobial activity of lavender, mint, and rosemary essential oils and the effect of their vapours on growth of Penicillium spp. in a bread model system. Molecules 2021, 26, 3859. [Google Scholar] [CrossRef]
- Goyal, S.; Tewari, G.; Pandey, H.K.; Kumari, A. Exploration of productivity, chemical composition, and antioxidant potential of Origanum vulgare L. grown at different geographical locations of Western Himalaya. India J. Chem. 2021, 12. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Taghinezhad, E.; Setzer, W.N.; Chen, G. Susceptibility of Tribolium castaneum (Coleoptera: Tenebrionidae) to the fumigation of two essential Satureja oils: Optimization and modeling. Processes 2021, 9, 1243. [Google Scholar] [CrossRef]
- Ed-Dra, A.; Nalbone, L.; Filali, F.R.; Trabelsi, N.; El Majdoub, Y.O.; Bouchrif, B.; Giarratana, F.; Giuffrida, A. Comprehensive evaluation on the use of Thymus vulgaris essential oil as natural additive against different serotypes of Salmonella enterica. Sustainability 2021, 13, 4594. [Google Scholar] [CrossRef]
- Pavela, R. Insecticidal properties of phenols on Culex quinquefasciatus Say and Musca domestica L. Parasitol. Res. 2011, 109, 1547–1553. [Google Scholar] [CrossRef]
- Aissaoui, F.; Hedjal-Chebheb, M.; Soltani, A.; Haouel-Hamdi, S.; Talhi, O.; Chérif Ziani, B.E.; Mediouni-Ben Jemâa, J. Variations of chemical composition of two Algerian essential oils collected for different seasons and assessment of their insecticidal toxicity against three moth pests. Chem. Rev. 2007, 107, 2411–2502. [Google Scholar] [CrossRef]
- Liu, T.-T.; Chao, L.K.-P.; Hong, K.-S.; Huang, Y.-J.; Yang, T.-S. Composition and insecticidal activity of essential oil of Bacopa caroliniana and interactive effects of individual compounds on the activity. Insects 2020, 11, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patiño-Bayona, W.; Galeano, L.N.; Cortes, J.B.; Ávila, W.D.; Daza, E.H.; Suárez, L.; Prieto-Rodríguez, J.; Patiño-Ladino, O. Effects of Essential Oils from 24 Plant Species on Sitophilus zeamais Motsch (Coleoptera, Curculionidae). Insects 2021, 12, 532. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R. Acute, synergistic and antagonistic effects of some aromatic compounds on the Spodoptera littoralis Boisd. (Lep., Noctuidae) larvae. Ind. Crop. Prod. 2014, 60, 247–258. [Google Scholar] [CrossRef]
- Pavela, R. Acute toxicity and synergistic and antagonistic effects of the aromatic compounds of some essential oils against Culex quinquefasciatus Say larvae. Parasitol. Res. 2015, 114, 3835–3853. [Google Scholar] [CrossRef] [PubMed]
- Campos, E.V.; Proença, P.L.; Oliveira, J.L.; Mansi Bakshi, P.C.; Fraceto, L.F.A. Use of botanical insecticides for sustainable agriculture: Future perspectives. Ecol. Indic. 2019, 105, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Carayon, J.-L.; Téné, N.; Bonnafé, E.; Alayrangues, J.; Hotier, L.; Armengaud, C.; Treilhou, M. Thymol as an alternative to pesticides: Persistence and effects of Apilife Var on the phototactic behaviour of the honeybee Apis mellifera. Environ. Sci. Pollut. Res. Int. 2014, 21, 4934–4939. [Google Scholar] [CrossRef]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial properties of plant essential oils against human pathogens and their mode of action: An updated review. Evid. Based Complement. Alternat. Med. 2016, 2016, 3012462. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Scientific Name | Family | GPS/Location | Part | Essential Oil Yield (% (w/w)) |
|---|---|---|---|---|---|
| Garden thyme | Thymus vulgaris | Lamiaceae | 37.159592, 49.001185 | Leaves | 1.27 ± 0.02 c |
| Lavender | Lavandula angustifolia | Lamiaceae | 38.419401, 48.860797 | Leaves | 1.61 ± 0.06 b |
| Sweet wormwood | Artemisia annua | Asteraceae | 37.198539, 49.653243 | Leaves | 2.90 ± 0.03 a |
| Summer Savoury | Satureja hortensis | Lamiaceae | 35.322589, 51.625637 | Leaves | 2.11 ± 0.09 ab |
| Rosemary | Rosmarinus officinalis | Lamiaceae | 37.193775, 49.641361 | Leaves | 1.10 ± 0.00 c |
| Oregano | Origanum vulgare | Lamiaceae | 36.949912, 50.603010 | Leaves | 1.72 ± 0.01 b |
| Essential Oil | Slope ± SE | X2 (df = 3) | LC50 (95% CI) a (ppm) | Relative Potency b |
|---|---|---|---|---|
| Artemisia annua | 1.74 ± 0.26 | 4.35 | 152.8 (64.24–291.00) | 8.94 |
| Lavandula angustifolia | 1.66 ± 0.28 | 4.62 | 287.5 (99.81–566.01) | 4.75 |
| Origanum vulgare | 2.04 ± 0.34 | 2.55 | 1366.2 (904.03–1094.6) | 1.00 |
| Rosmarinus officinalis | 1.70 ±0.27 | 4.15 | 315.9 (117.2–578.5) | 4.32 |
| Satureja hortensis | 1.31 ± 0.22 | 4.20 | 180.6 (53.09–409.29) | 7.56 |
| Thymus vulgaris | 1.79 ± 0.293 | 4.31 | 122.8 (56.2–235.0) | 11.12 |
| Essential Oil | Main Components (%) | References |
|---|---|---|
| A. annua | Camphor OM (13.1%), artemisia ketone OM (11.8%), β-selinene SH (10.7%), pinocarvone OM (7.4%), 1,8-cineole OM (6.8%), α-pinene MH (5.9%), caryophyllene oxide OS (5.4%), and β-caryophyllene SH (3.1%). | [47] |
| L. angustifolia | Linalool acetate OM (35.0%), linalool OM (32.7%), 1,8-cineole OM (8.1%), camphor OM (6.4%), borneol OM (2.4%), and lavandulyl acetate OM (2.0%). | [48] |
| R. officinalis | 1,8-Cineole OM (40.4%), camphor OM (11.9%), α-pinene MH (8.7%), β-pinene MH (6.9%), caryophyllene SH (5.3%), borneol OM (3.9%), and camphene MH (3.5%). | [48] |
| O. vulgare | Thymol OM (38.8%), γ-terpinene MH (13.6%), linalool MH (13.0%), p-cymene MH (9.5%), caryophyllene SH (4.5%), and β-myrcene MH (3.4%). | [49] |
| S. hortensis | Estragole PP (68.0%), eugenol PP (5.6%), (E)-p-methoxycinnamaldehyde PP (4.4%), allo-spathulenol OS (3.5%), α-pinene MH (1.3%), and β-pinene MH (1.2%). | [50] |
| T. vulgaris | Thymol OM (38.7%), p-cymene MH (15.7%), carvacrol OM (14.9%), γ-terpinene MH (13.3%), β-caryophyllene SH (2.7%), and 3-carene MH (2.2%). | [51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valizadeh, B.; Jalali Sendi, J.; Oftadeh, M.; Ebadollahi, A.; Krutmuang, P. Ovicidal and Physiological Effects of Essential Oils Extracted from Six Medicinal Plants on the Elm Leaf Beetle, Xanthogaleruca luteola (Mull.). Agronomy 2021, 11, 2015. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102015
Valizadeh B, Jalali Sendi J, Oftadeh M, Ebadollahi A, Krutmuang P. Ovicidal and Physiological Effects of Essential Oils Extracted from Six Medicinal Plants on the Elm Leaf Beetle, Xanthogaleruca luteola (Mull.). Agronomy. 2021; 11(10):2015. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102015
Chicago/Turabian StyleValizadeh, Bita, Jalal Jalali Sendi, Marziyeh Oftadeh, Asgar Ebadollahi, and Patcharin Krutmuang. 2021. "Ovicidal and Physiological Effects of Essential Oils Extracted from Six Medicinal Plants on the Elm Leaf Beetle, Xanthogaleruca luteola (Mull.)" Agronomy 11, no. 10: 2015. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102015