
Parthenocarpy and Self-Incompatibility in Mandarins
by
 , , ,
, , ,
Rafael Montalt
1 ,
,
María Carmen Vives
1,
Luis Navarro
2,
Patrick Ollitrault
3,4 and
Pablo Aleza
2,* 1
Centro de Protección Vegetal y Biotecnología, Instituto Valenciano de Investigaciones Agrarias (IVIA), Carretera CV-315, km 10.7, Moncada, 46113 Valencia, Spain
2
Centro de Citricultura y Producción Vegetal, Instituto Valenciano de Investigaciones Agrarias (IVIA), Carretera CV-315, km 10.7, Moncada, 46113 Valencia, Spain
3
CIRAD, UMR AGAP, 20230 San Giuliano, France
4
UMR AGAP, Institut Agro, CIRAD, INRAE, Université Montpellier, 34060 Montpellier, France
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(10), 2023; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102023
Submission received: 13 September 2021
/
Revised: 6 October 2021
/
Accepted: 7 October 2021
/
Published: 9 October 2021
(This article belongs to the Special Issue Advances in Citrus Breeding, Genetics, Physiology and Horticultural Management)
Abstract
:Citrus reproductive biology is complex. One of its characteristic features is parthenocarpy that enables seedless fruit production. Citrus parthenocarpy and self-incompatibility knowledge is only partial and sometimes discrepant. Increasing such knowledge is relevant for better managing cultivated varieties and improving the selection of parents in breeding strategies to recover seedless varieties such as mandarins. This work develops an efficient protocol to characterize self-incompatibility and different parthenocarpy types based on emasculation, hand self-pollination, and hand cross-pollination. It analyzes fruit setting and seed production coupled with histological pollen performance observations. We analyzed the reproductive behavior of nine mandarin varieties with relevant characteristics as parents for seedless mandarin breeding. ‘Clemenules’ clementine and ‘Moncada’ mandarins were strictly self-incompatible with facultative and vegetative parthenocarpy; ‘Imperial’ mandarin and ‘Ellendale’ tangor displayed no strict self-incompatibility associated with facultative and vegetative parthenocarpy; ‘Fortune’ mandarin was self-incompatible with facultative and stimulative parthenocarpy; ‘Campeona’ and ‘Salteñita’ mandarins were self-compatible with vegetative parthenocarpy; ‘Serafines’ satsuma was associated with male sterility together with facultative and vegetative parthenocarpy; and ‘Monreal’ clementine was self-compatible and nonparthenocarpic. Our protocol can be applied for screening of mandarin germplasm and to characterize new parents. Reproductive behavior knowledge is important for optimizing seedless mandarin breeding programs based on diploidy, triploidy, or induced mutagenesis.
1. Introduction
Seedlessness is one of the most important characteristics for citrus fresh-fruit markets because consumers do not accept seedy fruit. Breeding programs that aim to obtain seedless varieties focus mainly on mandarins, which are a large and diverse group with wide genetic variability [1]. Seedlessness in citrus differs among genotypes, and is sometimes altered by environmental conditions and cross-pollination success [2].
Two main seedlessness types have been described in plants: (1) parthenocarpy in sensu stricto, which occurs when the ovary develops directly without fertilization and produce seedless fruits and (2) stenospermocarpy, in which fruits are seedless because the ovule/embryo aborts without producing mature seed [3]. In citrus, stenospermocarpy is uncommon [4] and ovule sterility or lack of its fecundation (self-incompatibility or no compatible viable pollen) coupled with parthenocarpy is necessary for the production of seedless fruits.
Ovule sterility, lack of fecundation, or no seed development at early stages often prevents seed formation and different mechanisms have been reported in citrus [2,5]. Ovule sterility can be originated in flowers whose pistils do not develop up to the functional stage [6]. For example, in nonfunctional pistils of lemon (Citrus limon (L.) Burm f.), possible blocking of further stigma and style development is associated with the presence or absence of receptive embryo sacs in the ovule [6]. Osawa [7] observed degeneration of the embryo sac in both navel sweet orange (C. sinensis (L.) Osbeck) and satsuma mandarin (C. unshiu (Mak.) Marc.) and Wong [8] reported chromosome aberrations into the embryo sac as the responsible mechanism of low seed number in hand-pollinated flowers of ‘Valencia’ sweet orange and ‘Marsh’ grapefruit (C. paradisi Macf.). In addition to ovule sterility, other mechanisms have been described in citrus originating seedless citrus fruits coupled with parthenocarpy. ‘Mukaku Kishu’ mandarin (C. kinokuni hort. ex Tanaka) produces seedless fruits because the seed development is arrested at the early stage of development [9,10]. However, for easy reading of the manuscript, hereinafter with the term female sterility we refer to the different mechanisms that prevent seeds formation in citrus fruits: (1) the failure of the pistil development up to the functional stage, (2) ovule sterility or (3) the early stage arrest of the seed development.
Male sterility prevents seed formation in the absence of cross-pollination and prevents pollen flow toward neighbor orchards. In citrus, various kinds of male sterility related to chromosome aberration occur at the diploid level and result in different degrees of pollen fertility [2]. The most marked male sterility in citrus is due to the gene–cytoplasmic interaction, as in satsumas where male sterility is associated with failed pollen grain development and scant viability [11]. Recently, two QTLs related to male sterility have been reported; MS-P1, which is a major QTL for reducing the number of pollen grains per anther and MS-F1, related to lower apparent pollen fertility [12].
Both female and male sterility can be achieved by gamma irradiation [13,14] and triploidy [2,15,16]. In fact, triploid hybrids have very little pollen and poor ovule fertility, and are generally considered sterile [17], although they can occasionally produce very few seeds. Triploid citrus plants can be recovered from sexual hybridization between two diploid parents from the union of an unreduced megagametophyte with a haploid pollen [18,19], or by hybridization between diploid and tetraploid parents [20,21].
Self-incompatibility, also called self-sterility, causes self-pollen rejection and prevents seed formation in the absence of cross-pollination. A recent report indicates that citrus gametophytic self-incompatibility is based on S-RNases, which act as S genes determinants in inhibiting pollen tube growth [22]. The most important self-incompatible horticultural citrus groups are pummelos (C. maxima (Burm.) Merr.), clementines (C. clementina Hort. ex Tan.), and several natural or artificial mandarin hybrids [22,23,24].
The term ‘Parthenokarpie’ (which literally means ‘virgin fruit’) was introduced in 1902 to refer to seedless fruit production [25]. Seeds are the consequence of fertilization and embryo development, however most citrus genotypes presents sporophytic apomixis and produce polyembryonic seeds by adventitious embryony. Even in this case, nucellar embryos depends on sexual reproduction to produce endosperm for nucellar embryo growth and development and seed set [26,27]. Apart from their reproductive function, seeds act as a source of phytohormones required for fruit development [28,29]. The ability to accumulate sufficient levels of phytohormones in developing ovaries with no need for seeds is, therefore, a condition for natural seedless fruit production, which is known as parthenocarpic ability (PA) [30]. Different degrees of PA have been reported in citrus and PA is associated with high levels of endogenous gibberellins (GAs) measured in nonfertilized developing ovaries [31,32,33].
Four parthenocarpy types have been described in citrus [28]. A distinction is made between obligatory parthenocarpy for those varieties that always produce seedless fruit, and facultative parthenocarpy in which seedless fruit is produced when cross-pollination with compatible sources of pollen is prevented. Moreover, a distinction is made between vegetative parthenocarpy (also called autonomous or autonomic parthenocarpy) to refer to seedless fruit developing without requiring any external stimulus, and stimulative parthenocarpy that requires the pollination stimulus for seedless fruit set.
In citrus, the molecular mechanisms controlling parthenocarpy are poorly understood. On the basis of the different ratio of parthenocarpic hybrids obtained for several progenies of satsuma with different second parents, Vardi et al. [25] suggested that parthenocarpy was due to the action of three dominant complementary genes and that two of these genes were in homozygosity for parthenocarpy and one in heterozygosity in satsuma. This hypothesis for three dominant genes for parthenocarpy in citrus is in agreement with the conclusion of Sykes [34] based on the parthenocarpic segregation ratio in a diploid segregating progeny recovered between ‘Imperial’ (C. reticulata Blanco) and ‘Ellendale’ (C. reticulata × C. sinensis). However, Vardi et al. [28] proposed that parthenocarpy in satsuma was controlled by at least two dominant complementary genes, with satsuma being heterozygous for these genes. These contradictory hypotheses point out that parthenocarpy is a character that needs to be studied in more detail to understand how it is determined in citrus. High-density genotyping of segregating progenies for parthenocarpy, coming from different genetic pools, should allow us to decipher the genetic determinism of parthenocarpy and to identify candidate genes. In this regard, knowledge of PA is crucial in selecting parents that could be used to obtain segregating progenies for parthenocarpy.
Current parthenocarpy information for many genotypes is inaccurate and sometimes discrepant between authors. All this indicates the complexity of the factors affecting citrus reproductive biology. This work focuses on nine different varieties selected for their importance as parents for mandarin breeding programs [15]: ‘Clemenules’ and ‘Monreal’ clementines, ‘Campeona’ (C. nobilis Lour.), ‘Imperial’, ‘Salteñita’ (C. deliciosa Ten.), ‘Fortune’ (C. clementina × C. tangerina), and ‘Moncada’ [C. clementina × (C. unshiu × C. nobilis)] mandarins, ‘Ellendale’ tangor, and ‘Serafines’ satsuma.
Clementine, which was recovered from a ‘Common’ mandarin chance seedling, is the main mandarin varietal group cultivated in the Mediterranean Basin. As a result of spontaneous budsports mutations from the original ‘Commune’ clementine, many different clementine varieties have been selected, with ‘Clemenules’ as the main variety cultivated in Spain for its exceptional fruit quality. Discrepancies are found in the literature between the stimulative parthenocarpy proposed by Vardi et al. [25] and the vegetative parthenocarpy put forward by Mesejo et al. [33]. Regarding ‘Monreal’, a self-compatible natural mutation of the ‘Commune’ clementine, previous research points out the need of seeds for fruit to set [35]. However, the possibility of seedless fruit setting by a pollination stimulus without fecundation has not yet been explored in this variety.
‘Campeona’ and ‘Ellendale’ produce fruit with a medium to high caliber and a late and very late harvesting period, respectively, whereas ‘Imperial’ and ‘Salteñita’ mandarins are characterized by their particular fruit flavor and aroma. ‘Fortune’ and ‘Moncada’ produce fruits with excellent organoleptical qualities and a very late harvesting period. Altogether, the seeds of all these varieties, except for ‘Salteñita’, are monoembryonic, which facilitates the recovery of new hybrids by sexual hybridization when used as female parents. These traits make these varieties very interesting breeding parents.
Previous information reports that ‘Fortune’ and ‘Moncada’ mandarins are self-incompatible [23], but no information about the requirement of a pollination stimulus for fruit set has yet been reported. With ‘Campeona’ and ‘Salteñita’ mandarins, the scientific literature only reports that they are seedy varieties [36], but knowledge about PA goes unnoticed. Regarding both the ‘Imperial’ mandarin and ‘Ellendale’ tangor, previous research offers discrepant results about their PA and self-incompatibility [25,34,37,38,39,40,41]
‘Serafines’ is a spontaneous mutation of the ‘Owari’ satsuma characterized by its very late harvesting period, from January to the end of February, which is an outstanding satsuma variety feature. Satsumas are characterized by their male and female sterility and parthenocarpy [2], but no specific information about ‘Serafines’ is available.
Increasing knowledge about the parthenocarpy and self-incompatibility of different mandarin genotypes to be used as parents in sexual hybridizations is crucial to improve breeding program efficiency. In this research, we assessed the self-compatibility or self-incompatibility system, the PA and the requirement of pollination stimuli for the seedless fruit production of the aforementioned mandarin varieties to classify them according to the above-described parthenocarpy types and established a relatively simple protocol to evaluate these characters in new potential parents. For this purpose, we evaluated the data obtained for fruit setting, seed production and histological observations of pollen performance obtained from emasculated, hand self-pollinated and hand cross-pollinated flowers.
2. Materials and Methods
2.1. Plant Material
Nine mandarin varieties were used to carry out the present study. These varieties were grown in plots of the Citrus Germplasm Bank of the Instituto Valenciano de Investigaciones Agrarias (IVIA), located in Moncada, Valencia (Spain) and had the following accession numbers: ‘Clemenules’ (IVIA-022), ‘Monreal’ (IVIA-459), ‘Campeona’ (IVIA-193), ‘Imperial’ (IVIA-576), ‘Salteñita’ (IVIA-361), ‘Fortune’ (IVIA-080), ‘Moncada’ (IVIA-421), ‘Ellendale’ (IVIA-194), and ‘Serafines’ (IVIA-256). Four trees of each variety were used, and all were healthy [42] and cultivated following the same agronomic practices in a Mediterranean climate.
2.2. Experimental Procedure
Terminal flowers in pre-anthesis from shoots bearing one flower and leaves were used to perform the following treatments: emasculation (E), emasculation and hand self-pollination (SP), and emasculation and hand cross-pollination (CP). To avoid any undesired pollination, flowers were bagged after treatments. For the CP treatment, ‘Fortune’ was used as apollen source for its high pollination aptitude and ‘Clemenules’ was utilized to cross-pollinate ‘Fortune’. To perform hand-pollination, pollen grains from flowers in pre-anthesis were harvested by covering all the flowering period. Anthers were removed from stamens and placed in Petri dishes on silica gel at room temperature until dehiscence. Then pollen was used for pollination. Ten days after treatment, five SP and five CP flowers per tree from each variety were fixed in FAA solution (formalin, glacial acetic acid, 70% ethanol, 1:1:18, v/v) [43] and stored at 4 °C until histological observations were made. The other flowers were left for fruit set. Fruits were collected when ripe. The data for fruit setting and number of seeds per fruit were recorded.
In order to ensure consistent results, a large number of treated flowers were needed, which involved using four trees per variety to carry out our study. Once tree effect in the obtained results was discarded, treated flowers were analyzed together. The number of flowers employed in each treatment per variety and the number of fruits obtained are shown in Supplementary Table S1. In all, 6792 flowers (320 for histological observations, plus 6472 for fruit set) were used in this work. The results obtained from the above-described treatments enabled us to assess the self-compatibility or self-incompatibility and the PA in eight of the studied varieties. For ‘Monreal’, an additional CP treatment with the pollen of the ‘Oroblanco’ (C. grandis × C. paradisi) triploid hybrid was necessary to assess PA.
2.3. Histological Observations
The pistils fixed in FAA 10 days after SP and CP treatments were used for histological observations. Pollen performance in planta, including pollen germination and pollen tube growth, was analyzed under a Leica MZ16FA epifluorescence stereomicroscope equipped with GFP1 filter (excitation filter 395–455 nm and barrier filter 480 LP). For that purpose, pistils were submerged 3 times in water for 1 h. Afterward, pistils were sliced into 14 cross-sections using a sharp blade. Stigmas were sliced into two sections, styles into eight sections, and ovaries into four sections following the methodology described by Montalt et al. [44]. Sections were stained with 0.1% aniline blue in 0.1 N K3PO4 [45]. Pollen germination was observed on the stained stigma surfaces and pollen tubes identified by its callose plugs fluorescence [46].
2.4. Statistical Analysis
In order to evaluate self-incompatibility, the average number of seeds per fruit between treatments for each variety was compared. The data of the dependent variable number of seeds per fruit were confirmed to fit the normal distribution, and outlier values based on box plots were removed prior to further analyses. Analyses of variance and LSD multiple range tests were performed for comparisons using the Statgraphics Centurion XVI statistical software package, version 16.1.03. To evaluate PA, a fruit setting comparison between treatments was analyzed in each genotype. To this end, the proportions of fruit setting obtained in each treatment were compared by pairs. Each proportion was a binomial variable with two possible outcomes: ‘successful fruit setting’ and ‘failed fruit setting’. The 95% confidence interval obtained for the differences between treatments is shown in Supplementary Table S2.
2.5. Genetic Analysis with Simple Sequence Repeat (SSR) Markers
The plants recovered from the SP treatment performed in ‘Imperial’ and ‘Ellendale’ were genotyped using 13 heterozygous SSR markers located on the reference Clementine genetic map [47]. Of them, height heterozygous markers for each variety were selected to characterize their progenies. Detailed information about all the used markers is given in Supplementary Table S3.
Genomic DNA was extracted from leaves of the samples described above using a Plant DNeasy kit from Qiagen Inc. (Valencia, CA, USA) following the manufacturer’s protocol and measured using a spectrophotometer (NanoDrop 2000C, Thermo Fisher Waltham, MA, USA). The samples were diluted with sterile water (Sigma-Aldrich, Co., Gillingham UK) at a concentration of 10 ng/mL and stored at 20 °C until use. Polymerase chain reactions (PCRs) were performed using SSR markers with a Thermocycler rep gradient S (Eppendorf©. Hamburg, Germany) according to the following protocol: reaction volume, 15 µL containing 0.5 µL of 1 U/µL of Taq DNA polymerase (Fermentas© Waltham, MA, USA), 3 µL of citrus template DNA (10 ng/µL), 1.5 µL of 2 µM welled (Sigma© Burlington, MA, USA) dye-labeled forward primer, 1.5 µL of 2 µM non-dye-labeled reverse primer, 0.2 µM of each dNTP, 1.5 µL of PCR reaction buffer 10×, and 0.45 µL of 50 mM MgCl2. The cycling program was set as follows: denaturation for 5 min at 94 °C followed by 40 cycles of 30 s at 94 °C, 30 s at 50 °C or 55 °C, 30 s at 72 °C; and a final elongation step of 8 min at 72 °C. Separation was carried out by capillary gel electrophoresis using a Genetic Analysis System 8000 (Beckman Coulter Inc. Brea, CA, USA). PCR products were initially denatured at 90 °C for 2 min, injected at 2 kV for 30 s, and separated at 6 kV for 35 min. Alleles were size-based on a DNA size standard (400 bp). The GenomeLabTM v.10.0 (Beckman Coulter Inc.) genetic analysis software was used for data collection.
3. Results and Discussion
We first analyzed whether self-incompatibility prevented self-fertilization in each variety. Next, we assessed if the studied varieties had PA and if a pollination stimulus was required for seedless fruit to set. Based on the obtained results, we discuss the self-compatibility or self-incompatibility reactions observed, the parthenocarpy type in each studied variety and their implications in breeding programs aimed to obtain seedless varieties by triploidization or induced mutagenesis.
3.1. Self-Incompatibility
In order to determine self-compatibility or self-incompatibility, we observed the presence or absence of pollen tubes reaching the ovaries in the histological sections of the flowers fixed 10 days after SP (Figure 1), as well as the production of seeded or seedless fruit from SP (Table 1).
In ‘Monreal’, ‘Campeona’ and ‘Salteñita’, the histological sections of SP flowers showed pollen tubes growing through the pistil and reaching the ovaries in all the analyzed flowers of the three varieties (Figure 1 and Supplementary Figure S1) and all the recovered fruits contained seeds (Table 1). No differences were seen in ‘Monreal’ when comparing the average seed number between SP (22.5 seeds per fruit) and CP (21 seeds per fruit) treatments. With ‘Campeona’ (4.9 seeds per fruit from SP vs. 9.9 seeds per fruit from CP) and ‘Salteñita’ (11 seeds per fruit from SP vs. 16.5 seeds per fruit from CP), the fruits obtained from the SP flowers presented lower seed numbers than those from the CP flowers (Table 1). Taking together, the results obtained for seed production and pollen tubes growth in the pistils of the self-pollinated flowers, we concluded that ‘Monreal’, ‘Campeona’ and ‘Salteñita’ are self-compatible.
Regarding ‘Clemenules’, ‘Fortune’ and ‘Moncada’, the histological observations of the self-pollinated flowers showed high pollen grain germination on stigma surfaces and pollen tube growth in the bottom half of the stigma. Nevertheless, pollen tubes stopped growing in the upper style part and no pollen tubes were identified in ovaries (Figure 1 and Supplementary Figure S1). All the recovered fruits from the SP treatment were seedless (Table 1). These results testify a self-incompatible system in ‘Clemenules’, ‘Fortune’ and ‘Moncada’.
Clementines have been studied in detail for their self-incompatibility reaction using histological approaches [23,48,49,50]. ‘Monreal’ display different features than other clementines. Distefano et al. [49] already observed that it was self-compatible, with similar results to those obtained in the present work. ‘Monreal’ was obtained from a spontaneous mutation of ‘Commune’ clementine later known as ‘Fina’ clementine which is a self-incompatible variety [49]. Liang et al. [22] identified the S-RNase gene and eleven S-locus F-box (SLF) genes on scaffold 7 of C. clementina reference genome implicated on self-incompatibility system. These findings suggest that the self-compatibility reaction in ‘Monreal’ may result from a mutation or epigenetic variation on these genes. A frameshift mutation in one S-RNase has been described to result in a loss of self-incompatibility in the genus Citrus [22]. With ‘Fortune’, our results coincide with the SI previously reported by Yamamoto et al. [23] and Distefano et al. [50].
In ‘Imperial’ and ‘Ellendale’, the histological observations displayed similar results in both varieties (Supplementary Figure S2). Pollen tubes were observed throughout the stigma, and the growth of most of them stopped in the upper style, although some pollen tubes reached ovaries in small percentages (15% for ‘Imperial’, 10% for ‘Ellendale’) of the self-pollinated flowers (Table 1). Unlike the other varieties studied, ‘Imperial’ and ‘Ellendale’ produced both seedless and low-seeded fruits from SP. The percentage of low-seeded fruit was 23% for ‘Imperial’ and 54% for ‘Ellendale’ and the seed number per seeded fruit was 2.3 ± 1.6 and 5.7 ± 6.2 respectively. This resulted in the values of 0.8 and 0.7 seeds per fruit shown on Table 1 for ‘Imperial’ and ‘Ellendale’ respectively. When taking into account both histological observations and poor seed production from the self-pollinated flowers, we demonstrated that some pollen tubes were able to reach ovaries, fertilize ovules and then produce seeds, which suggests no strict self-incompatibility in these two varieties under our field conditions. To rule out any uncontrolled pollination hypothesis, 12 seeds obtained from the SP treatment of each variety were cultivated under greenhouse conditions [20], and six and 11 plantlets were recovered from ‘Imperial’ mandarin and ‘Ellendale’ respectively. The progenies of each variety were analyzed with eight heterozygous markers for the female parent [19,51,52,53,54,55] and all plantlets displayed only alleles from the female parent in either heterozygosity or homozygosity (Figure 2, Supplementary Tables S3 and S4). These results indicate that the obtained plantlets originated from selfing.
Our results are in line with the low-seeded fruits obtained from self-pollinated flowers in ‘Ellendale’ reported by Vithanage [40] even though this author considered ‘Ellendale’ as a highly self-compatible variety. However, Vardi et al. [25] considered ‘Ellendale’ as self-incompatible variety. Based on the ratio 1:1 (SC:SI) of the hybrids obtained from Satsuma x ‘Ellendale’ cross, this author suggested two different self-incompatible alleles (SY and SZ) in ‘Ellendale’ which differs from the SX allele proposed for Satsuma. Recently, based on segregation distortion for male parents in high density genetic maps from reciprocal crosses between ‘Ellendale’ and ‘Fortune’, Ollitrault et al. [56] concluded that ‘Ellendale’ and ‘Fortune’ share a common self-incompatible allele and confirmed the location of the S locus at the beginning of the chromosome 7 of the Clementine reference genome. From the haplotype sequence analysis on the surrounding genomic region the same authors concluded that ‘Ellendale’ share the same two self-incompatible alleles as Clementine as already proposed by Kim et al. [57] from crosses with homozygous lines for self-incompatible alleles.
In ‘Imperial’, based on the observation of pollen tube growth and the production of seeded or seedless fruit, discrepant results have been reported regarding SI. On the one hand, Wallace and Lee [41] recorded some pollen tubes (0.2 ± 0.1) at the basis of the style and low-seeded fruits (less than one seed per fruit) form selfed flowers. However, these authors considered ‘Imperial’ to be self-incompatible since almost all pollen tubes were arrested in the stigma and upper style of self-pollinated flowers and very low number of seeds were obtained. On the other hand, Vithanage [39] and Sykes [34] concluded this variety was self-compatible since pollen tubes were observed growing into the self-pollinated styles and seeded fruits were obtained from self-pollinated flowers. In this regard, Sykes [37] suggested that the environment might affect self-pollination in ‘Imperial’ because these experiments were performed in different locations.
Taking advantage of staining techniques to observe pollen tubes growth inside pistils, different authors have demonstrated that in citrus, self-incompatibility reaction takes place from the stigma to the ovary [23,58] although in mandarin and mandarin hybrids several authors agreed that self-incompatibility reaction is originated in the first half of the style [23,48,49,50]. These studies have been carried out in different years and in different countries with different environmental conditions (Catania, Italy; Adana, Turkey; Kagoshima, Japan; Moncada, Spain) and all of them coincided with the SI classification of clementines and ‘Fortune’. Taking into account that environment may affect self-pollination in ‘Imperial’, as has been suggested by Sykes [37], and the stability of the self-incompatibility reaction in clementines and ‘Fortune’ reported in the different locations mentioned above, it seems plausible to hypothesize that the impact of environmental change on the SI reaction is dependent on the considered genotype. Beyond the stability of the SI reaction displayed in clementines and ‘Fortune’ under field conditions at different locations, previous research performed in these two varieties under controlled environmental conditions indicate that constant high and low temperatures appear to have an effect on the self-incompatibility reaction by affecting the place where pollen tubes are arrested [59]. Recently, Aloisi et al. [60] indicated that temperature contributed to a different activation of the self-incompatibility reaction in C. clementina, occurring at an optimal temperature of 25 °C and bypassed at 15 °C. The incompatible reaction resulted in enhancement of both transglutaminase enzyme activity and levels of conjugated polyamines when compared to cross-pollination [60]. However, more research is needed to understand how environmental conditions can influence the self-incompatibility reaction in citrus.
In ‘Serafines’, pale-colored anthers were obtained, which produced very scant pollen with very poor viability. Notwithstanding, we used these anthers to perform the SP treatment. We observed very few pollen tubes at the bottom of the stigma (Supplementary Figure S1), which is in line with the reduced pollen germination of satsuma reported by Vithanage [40]. However, in our observations, no pollen tubes were found in the upper style (Supplementary Figure S1). The seedless fruit set obtained from SP was, therefore, a consequence of poor pollen performance. Therefore, the self-incompatibility of ‘Serafines’ could not be assessed. Vardi et al. [25] considered satsuma as self-compatible and heterozygous with a self-compatible allele and a self-incompatible one. However, the authors did not give information on the variety used for their study.
In those varieties that produced seedless fruits from SP treatment, the seedy fruit production obtained from CP treatment indicates the potential to produce both seedless and seeded fruits, indicating that parthenocarpy itself is independent from the fertilization process and seed formation. This uncoupling of the reproductive and fruiting processes has been reported previously in ‘Fortune’ by Distefano et al. [61].
3.2. Assessing Parthenocarpic Ability (PA) and Testing the Pollination Stimulus Requirement for Fruit to Set
Under natural conditions, PA in self-incompatible genotypes is easily identified by avoiding cross-pollination. However, in self-compatible genotypes, PA can only be ascertained by emasculating and bagging (treatment E). As emasculating and bagging prevent pollination stimulus, the fruit set obtained from E (which is mandatory seedless) was assessed to check the pollination stimulus requirement for fruit to set. The fruit setting percentage from E obtained in ‘Clemenules’ (15%), ‘Campeona’ (9%), ‘Imperial’ (19%), ‘Salteñita’ (5%), ‘Moncada’ (34%), ‘Ellendale’ (5%) and ‘Serafines’ (64%) (Table 2) indicated that these varieties had PA and pollination stimulus was not required for fruit to set.
In the self-incompatible varieties ‘Clemenules’, ‘Ellendale’, and ‘Moncada’, the fruit setting comparison between treatments E and SP did not show any statistical differences (Table 2). This similar fruit setting obtained from treatments E and SP indicated not only that no pollination stimulus was required for fruit set, but also pollination stimulus did not increase fruit setting. In ‘Fortune’, no fruit was obtained from E, but the fruit setting from SP was 9% (Table 2) and all the recovered fruit was seedless (Table 1). This scenario indicated that ‘Fortune’ had PA and pollination stimulus was necessary for fruit to set.
‘Monreal’ was unable to produce fruit from emasculated flowers. Due to its self-compatibility, we used pollen from the ‘Oroblanco’ triploid hybrid to evaluate the ability of ‘Monreal’ to produce seedless fruit when seed formation was avoided and pollination stimulus was maintained. We pollinated 140 emasculated flowers and bagging. Ten of them were fixed in FAA 10 days after hand-cross pollination and were used to perform histological observations. High quantity pollen grains were observed on the stigmatic surface, which resulted in poor pollen germination and pollen tube growth. Only in two of the 10 observed pistils a few pollen tubes reached ovaries (Figure 3). The remaining 130 flowers were left for assessing fruit setting and seed content. Eight fruit were obtained and they all contained seeds with an average of three seeds per fruit. The fact that no seedless fruit was obtained by any treatment suggests lack of PA in ‘Monreal’.
Apart from identifying PA based on the ability to produce seedless fruit or not, as discussed above, a high or low degree of PA was assessed by comparing the fruit setting percentages between treatments E that produced only seedless fruit and CP that produced only seeded fruit.
In ‘Clemenules’, ‘Campeona’, ‘Imperial’, ‘Salteñita’, ‘Fortune’, ‘Moncada’, and ‘Ellendale’, the fruit setting percentages obtained from CP were higher than those obtained from E (and SP in ‘Fortune’) (Table 2), which indicates that the presence of fertilized ovules strongly influenced fruit set.
In contrast, ‘Serafines’ showed no differences in the fruit setting percentages between E (64%) and CP (65%) (Table 2), which implies greater PA in this variety. When comparing satsuma and clementine, previous research has associated high levels of endogenous GAs in developing ovaries of the satsuma with greater PA, whereas clementine produced lower GA levels and less PA [31,32]. Our results displayed ‘Serafines’ as the variety with greater PA in which seed production did not appear crucial for fruit set. Therefore, the comparison of fruit setting between E and CP offers a methodology to identify citrus genotypes with different PA levels.
In citrus, competition between flowers results in a marked drop of flowers and fruitlets [62]. Together with several factors that affect fruit setting, such as floral load, inflorescence typeb and flower position [63], conducting more work using a large quantity of flowers on different trees is necessary to assess reliable PA data. The results presented herein are supported by the large quantity of treated flowers and, thus, provide consistent PA data.
3.3. Parthenocarpic Classification
We classified the parthenocarpy of each variety according to the four types described in citrus by Vardi et al. [28]. To classify each variety as either vegetative parthenocarpy, which allows seedless fruit to set with no external stimuli, or stimulative parthenocarpy, we tested whether pollination stimulus was necessary for seedless fruit to set. Regarding the distinction made between facultative and obligatory parthenocarpy, the conditions under which seedless fruit production occurs are crucial. Facultative parthenocarpy produces seedless fruit when cross-pollination with compatible pollen is avoided, and corresponds to self-incompatible or male sterility varieties. Obligatory parthenocarpy always produces seedless fruit regardless of pollination conditions and, therefore, corresponds to varieties with female sterility. Beyond the poor pollen performance displayed by ‘Serafines’, all the studied varieties are male and female fertile ones and bear seeded fruit when cross-pollinated. Therefore, self-incompatibility for these varieties is key to produce seedless fruit under natural conditions.
‘Clemenules’ and ‘Moncada’ have self-incompatibility and produced seedless fruit from SP treatment (Table 1), which reveals facultative parthenocarpy. In both varieties, fruit setting percentage obtained when pollination stimulus was removed (E treatment) was similar to those obtained from SP treatment (Table 2) and consequently, displayed vegetative parthenocarpy. Therefore, we classified ‘Clemenules’ and ‘Moncada’ as facultative and vegetative parthenocarpic. With ‘Clemenules’, this result agrees with the pollination-independent proposed by Mesejo et al. [33] but challenges the classification of stimulative parthenocarpy, as proposed by Vardi et al. [25].
‘Fortune’, as ‘Clemenules’ and ‘Moncada’, displayed a self-incompatibility reaction and produced seedless fruits from SP treatment (Table 1). In contrast, no fruit was obtained when the pollination stimulus was removed (E treatment in Table 2). This denotes that ‘Fortune’ requires a pollination stimulus to set fruit. Therefore, apart from the facultative parthenocarpy related to self-incompatibility, we classified ‘Fortune’ as having stimulative parthenocarpy. To the best of our knowledge, we are the first to report the requirement of a pollination stimulus for fruit to set in ‘Fortune’.
‘Imperial’ and ‘Ellendale’ produced 19% and 5% of fruit setting respectively after removing the pollination stimulus (E treatment in Table 2), and all fruits were seedless. The percentage of fruit setting after SP treatment was 33% and 5% respectively (Table 2). In ‘Imperial’, 23% of fruits recovered from SP treatment contained seeds and the other 77% were seedless. Regarding ‘Ellendale’, 54% of fruits recovered from SP treatment contained seeds and the rest were seedless. The average number of seeds per fruit was 0.8 and 0.7 for ‘Imperial’ and ‘Ellendale’ respectively (Table 1). Therefore, we classified ‘Imperial’ and ‘Ellendale’ as facultative and vegetative parthenocarpic, but point out that low-seeded fruit can be produced even if cross-pollination is avoided. With ‘Imperial’, previous studies [37,38,39,41] coincided with our results and reported PA in this variety. Furthermore, Sykes [37] reported vegetative (autonomic) parthenocarpic fruit development. In contrast, ‘Ellendale’ has been studied by different authors who reached quite opposite conclusions. Vithanage [40] classified it as nonparthenocarpic, Vardi [25] as stimulative parthenocarpic, and Sykes [37] reported vegetative (autonomic) parthenocarpic fruit development. Our results obtained under environmental conditions of Valencia and from more than 800 flowers enabled us to classify ‘Ellendale’ with facultative and vegetative parthenocarpy, which rules out the hypotheses proposed by the first two authors, but agrees with Sykes [37].
For ‘Campeona’ and ‘Salteñita’ fruit setting percentages from SP treatment were respectively 42% and 58% (Table 2). Given their self-compatibility, all fruits recovered were seeded with an average of 4.9 seeds per fruit in ‘Campeona’ and 11 seeds per fruit in ‘Salteñita’ (Table 1). However, we found that they were able to produce seedless fruit when self-pollination was avoided by emasculating and bagging (E treatment), with a fruit setting percentage of 9% and 5% for ‘Campeona’ and ‘Salteñita’ respectively (Table 2), which means that they possess PA and do not need a pollination stimulus to set seedless fruit. As the term parthenocarpy is used to refer to seedless fruit production, self-compatible varieties can be classified as nonparthenocarpic. However, as ‘Campeona’ and ‘Salteñita’ possess PA, classifying these varieties as nonparthenocarpic can be confusing. The fact that in natural conditions, self-compatible genotypes produce seedy fruit even if cross pollination is avoided and seedless fruit can be recovered only from emasculation, which has to be performed by hand, explains why the scientific literature only reports them as seedy varieties [36], but information about PA goes unnoticed. We classify ‘Campeona’ and ‘Salteñita’ as self-compatible varieties with PA, which, therefore, provides relevant information on the parthenocarpy of these varieties.
For ‘Serafines’, we obtained practically the same fruit setting percentage after E, SP, and CP treatments, with values of 64, 66, and 65% respectively. Seedless fruits were obtained in all treatments except for CP, recovering an average of 5.3 seeds per fruit (Table 1). By taking into account the strictest meaning of obligate and facultative parthenocarpy, it must be considered to be facultative parthenocarpy because ‘Serafines’ is not female-sterile. As for requiring a pollination stimulus for fruit to set, ‘Serafines’ was able to produce seedless fruit from the E flowers with similar percentage values to those obtained from the CP flowers (Table 2). Considering the results herein obtained, we classified ‘Serafines’ as a facultative and vegetative parthenocarpic variety. Vegetative parthenocarpy has been reported in other satsuma varieties generally considered to be male and female sterile [2]. However, satsumas have been used in breeding programs as parents in different countries, particularly in Japan [64]. Extremely interesting new hybrid varieties have been recovered, such as ‘Kiyomi’ tangor (C. unshiu × C. sinensis) [65], ‘Queen’ mandarin (C. unshiu × unknown) [66], ‘Primosole’ mandarin (C. unshiu × C. reticulata) [67], among others. These indicate that the viability of ovules and pollen grains of satsumas is strongly influenced by not only environmental conditions but also by genotype.
Male and female gametophytes (from meiosis to zygote formation) are exposed plant structures, which potentially makes them especially susceptible to environmental conditions [68,69]. Temperature is one of the main environmental conditions that influences the success of plant sexual reproduction [70,71] and several studies report the impact of temperature on gametophytic generation and the progamic phase [44,69,72,73,74,75]. To the best of our knowledge, influence of environmental conditions in PA is unnoticed in citrus. However, previous research performed under controlled environmental conditions coincided in the influence of temperature in the self-incompatibility reaction, as we have discussed previously in this work.
The results in this work were obtained from a large number of flowers to ensure their robustness and consistency, although it was not been carried out in consecutive years, for which we cannot determine the effect of environmental conditions over data reproducibility. However, in a preliminary study carried out during 2013 and 2014 within this framework, we evaluated the seed production in SP flowers and the fruit setting from E, SP, and CP flowers [76]. A summary of the results obtained in our preliminary study is shown in Supplementary Table S5. Both the number of seeds per fruit obtained from SP treatment and fruit setting percentage comparison between E and CP treatments are in line with those obtained in the present work suggesting that SI and PA in the studied varieties are stable under our Mediterranean field conditions.
Regarding the methodology used in our study, we identified the PA by testing the ability of each genotype to produce seedless fruits. In addition, we assessed the PA degree by comparing the percentages of fruit setting between emasculated (E treatment) and cross-pollinated (CP treatment) flowers (Table 2). In order to ensure the robustness of the results presented in Table 2, a large number of treated flowers was needed (Supplementary Table S1) and we used four trees per variety. Supplementary Table S6 displays the number of treated flowers, recovered fruits, and percentage of fruit setting from E and CP treatments in each of the four trees used per variety. Data shown from the E treatment (Supplementary Table S6) suggest that PA can be identified in most of the varieties by performing E treatment in 50 flowers. Regarding the assessment of the PA degree, the comparison between E-CP treatments displayed the same differences in each replicate separately (Supplementary Table S6) and in merged data (Table 2). ‘Serafines’ displayed similar fruit set percentage from E and CP treatments in each of the four replicates (Supplementary Table S6) and also in merged data (Table 2). In the rest of varieties with PA, E treatment produced lower fruit set than CP treatment in all four replicates (Supplementary Table S6) and in merged data (Table 2). With these results, we suggest that PA can be assessed by utilizing 50 flowers in E and CP treatments, making this protocol more feasible to identify those varieties with higher PA.
Seedlessness is a major characteristic in citrus and a lot of breeding work has been accomplished to develop new seedless cultivars [2]. Diploid and triploid breeding programs and mutation breeding are different approaches followed to produce new seedless varieties. Increasing knowledge about self-incompatibility and parthenocarpy is relevant for improving the selection of parents that will be used in sexual hybridizations or mutagenesis.
Mutation breeding by gamma irradiation has been used in citrus to reduce seed production in diploid seedy hybrids [77]. Notwithstanding, the recovery of complete seedlessness or very low seedy genotypes is a very difficult issue but helpful to point out the importance of having strong parthenocarpic traits in breeding progenies [78]. Therefore, it is a key step to acquire prior knowledge of the parthenocarpic aptitude and the parthenocarpy type of the candidate diploid seedy varieties for irradiation since only those with these characteristics, and therefore capable to produce seedless fruits, will be suitable for being irradiated. Mutant lines from low or no PA varieties might require cultural practices such as girdling or GA treatments to stimulate fruit set and seedless fruit production [79].
4. Conclusions
Parthenocarpic ability is a key reproductive biology component because it enables seedless fruit production when combined with other reproductive features such as male and female sterility or self-incompatibility. We developed an efficient protocol to characterize the self-incompatibility and different types of parthenocarpy. It is based on emasculation, hand self-pollination, and hand cross-pollination and the analysis of fruit setting, seed production, and histological observations of pollen performance. We applied this protocol to analyze the reproductive behavior of nine important citrus varieties used as parents for seedless mandarin breeding. Of the four parthenocarpy types previously described for citrus, we found that ‘Clemenules’ and ‘Moncada’ were strictly self-incompatible with facultative and vegetative parthenocarpy, ‘Imperial’ and ‘Ellendale’ displayed no strict self-incompatibility associated with facultative and vegetative parthenocarpy, ‘Fortune’ was self-incompatible with facultative and stimulative parthenocarpy, and ‘Campeona’ and ‘Salteñita’ were self-compatible but with vegetative PA. ‘Serafines’ associated male sterility with facultative and vegetative parthenocarpy, while ‘Monreal’ clementine was not parthenocarpic. Our protocol can be applied for screening particular parents with previously identified interesting horticultural traits (e.g., production of nonapomictic seeds, resistance to Alternaria fungus, production of red fruit color, organoleptical qualities, etc.) and candidate-selected seedy diploid varieties with the objective to remove seeds by irradiation. A good reproductive behavior knowledge is important for optimizing seedless mandarin breeding programs.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/agronomy11102023/s1, Table S1: Number of the treated flowers and recovered fruits in the performed treatments (E, SP, and CP) in each variety; Table S2: 95% confidence interval in the paired comparison of fruit setting obtained in the performed treatments (E, SP, and CP) in each variety; Table S3: SSR markers used for the genetic analysis of the recovered plants from the self-pollinated flowers of ‘Imperial’ mandarin and ‘Ellendale’ tangor; Table S4: Observed alleles with the SSR markers used for the genetic analysis of the diploid hybrids recovered from the self-pollination of ‘Imperial’ mandarin and ‘Ellendale’ tangor; Table S5. Number of seeds per fruit (nsp) from SP treatment and fruit setting percentage (fsp) from E, SP, and CP treatments obtained in the preliminary study; Table S6. Number of treated flowers, recovered fruits and fruit set percentage from treatments E and CP in each of the four replicates R1, R2, R3, and R4, and for each studied variety; Figure S1. Histological sections of pistils after 10 days of self-pollination. Self-incompatible varieties: (a–c) ‘Clemenules’ clementine and (d–f) ‘Moncada’ mandarin; Self-compatible varieties: (g–i) ‘Campeona’ mandarin and (j–l) ‘Salteñita’ mandarin; Male sterile ‘Serafines’ satsuma (m–o) with no pollen tubes growing through the middle stigma. Left side of the figure represent a pistil with red lines indicating the transversal sections observed that corresponds with middle stigma (a), (d), (g), (j), (m), upper style (b), (e), (h), (k), (n), and ovary (c), (f), (i), (l), (o). Pollen tubes are marked by an arrow; va: vascular axis; sc: stylar canal; ov: ovule. Scale bars are depicted by blue lines: (a), (d), (g), (j), (m) 0.5 mm; (b), (e), (h), (k), (n) 0.25 mm; (c), (f), (i), (l), (o) 1 mm. Figure S2. Histological sections of pistils after 10 days of self-pollination. (a–f) ‘Imperial’ mandarin: (a–c) showing self-incompatibility reaction with no pollen tubes growing into the upper style and ovary. (d–f) ‘Imperial’ mandarin displaying pollen tubes into the style and ovary indicating partial self-incompatibility. (g-l) ‘Ellendale’ tangor: (g–i) showing self-incompatibility reaction with no pollen tubes growing into the upper style and ovary. (j–l) ‘Ellendale’ tangor displaying pollen tubes into the style and ovary indicating partial self-incompatibility. Left side of the figure represent a pistil with red lines indicating the transversal sections observed that corresponds with middle stigma (a), (d), (g), (j), upper style (b), (e), (h), (k) and ovary (c), (f), (i), (l). Pollen tubes are marked by an arrow; va: vascular axis; sc: stylar canal; ov: ovule. Scale bars are depicted by blue lines: (a), (b), (d), (e), (g), (h), (j) and (k) 0.25 mm; (f) and (l) 0.5 mm; (c) and (i) 1 mm.
Author Contributions
R.M.: conceptualization, methodology, writing—original draft. M.C.V.: funding acquisition. L.N.: conceptualization, writing—review and editing. P.O.: conceptualization, writing—review and editing. P.A.: conceptualization, validation, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Project RTA2015-00069-00-00 from the Spanish Ministry of “Economía y Competividad”, and Fondo Europeo de Desarrollo Regional (FEDER).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article or as Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in: the study design; data collection, analyses or interpretation; the writing of the manuscript; or the decision to publish the results.
References
- Garcia-Lor, A.; Luro, F.; Ollitrault, P.; Navarro, L. Genetic diversity and population structure analysis of mandarin germplasm by nuclear, chloroplastic and mitochondrial markers. Tree Genet. Genomes 2015, 11. [Google Scholar] [CrossRef]
- Ollitrault, P.; Froelicher, Y.; Dambier, D.; Luro, F.; Yamamoto, M. Seedlessness and ploidy manipulation. In Citrus Genetics, Breeding and Biotechnology; Khan, I., Ed.; CABI: Wallingfor, UK, 2007; pp. 197–218. [Google Scholar]
- Picarella, M.E.; Mazzucato, A. The Occurrence of Seedlessness in Higher Plants; Insights on Roles and Mechanisms of Par-thenocarpy. Front. Plant Sci. 2019, 9, 1997. [Google Scholar] [CrossRef] [Green Version]
- Mesejo, C.; Muñoz-Fambuena, N.; Reig, C.; Martínez-Fuentes, A.; Agustí, M. Cell division interference in newly fertilized ovules induces stenospermocarpy in cross-pollinated citrus fruit. Plant Sci. 2014, 225, 86–94. [Google Scholar] [CrossRef]
- Yamamoto, M. Progress on studies for seedless breeding of citrus in Japan. Adv. Hortic. Sci. 2014, 28, 64–72. [Google Scholar]
- Wilms, H.J.; Van Went, J.L.; Cresti, M.; Ciampolini, F. Structural aspects of female sterility in Citrus limon. Acta Bot. Neerl. 1983, 32, 87–96. [Google Scholar] [CrossRef]
- Osawa, I. Cytological and experimental studies in Citrus. J. Coll. Agric. Tokyo Univ. 1912, 4, 83–116. [Google Scholar]
- Wong, C.Y. The influence of pollination on seed development in certain varieties of citrus. Soc. Hortic. Sci. 1939, 37, 161–164. [Google Scholar]
- Yamasaki, A.; Kitajima, A.; Ohara, N.; Tanaka, M.; Hasegawa, K. Characteristics of Arrested Seeds in Mukaku Kishu-type Seedless Citrus. J. Jpn. Soc. Hortic. Sci. 2009, 78, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, A.; Kitajima, A.; Ohara, N.; Tanaka, M.; Hasegawa, K. Histological Study of Expression of Seedlessness in Citrus kinokuni ‘Mukaku Kishu’ and Its Progenies. J. Am. Soc. Hortic. Sci. 2007, 132, 869–875. [Google Scholar] [CrossRef] [Green Version]
- Goto, S.; Yoshioka, T.; Ohta, S.; Kita, M.; Hamada, H.; Shimizu, T. Segregation and Heritability of Male Sterility in Populations Derived from Progeny of Satsuma Mandarin. PLoS ONE 2016, 11, e0162408. [Google Scholar] [CrossRef]
- Goto, S.; Yoshioka, T.; Ohta, S.; Kita, M.; Hamada, H.; Shimizu, T. QTL mapping of male sterility and transmission pattern in progeny of Satsuma Mandarin. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Goldenberg, L.; Yaniv, Y.; Porat, R.; Carmi, N. Effects of Gamma-Irradiation Mutagenesis for Induction of Seedlessness, on the Quality of Mandarin Fruit. Food Nutr. Sci. 2014, 5, 943–952. [Google Scholar] [CrossRef] [Green Version]
- Bermejo, A.; Pardo, J.; Cano, A. Influence of gamma irradiation on seedless citrus production: Pollen germination and fruit quality. Food Nutr. Sci. 2011, 2, 169. [Google Scholar]
- Navarro, L.; Aleza, P.; Cuenca, J.; Juárez, J.; Pina, J.A.; Ortega, C.; Navarro, A.; Ortega, V. The Mandarin Triploid Breeding Program in Spain; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2015; Volume 1065, pp. 389–396. [Google Scholar]
- Ollitrault, P.; Dambier, D.; Francois, L.; Froelicher, Y. Ploidy Manipulation for Breeding Seedless Triploid Citrus. In Plant Breed Rev; John Wiley & Sons: New York, NY, USA, 2008; Volume 30, pp. 323–352. ISBN 9780470380130. [Google Scholar]
- Otto, S.P.; Whitton, J. Polyploid incidence and evolution. Annu. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleza, P.; Juárez, J.; Cuenca, J.; Ollitrault, P.; Navarro, L. Recovery of citrus triploid hybrids by embryo rescue and flow cytometry from 2x × 2x sexual hybridisation and its application to extensive breeding programs. Plant Cell Rep. 2010, 29, 1023–1034. [Google Scholar] [CrossRef]
- Cuenca, J.; Froelicher, Y.; Aleza, P.; Juárez, J.; Navarro, L.; Ollitrault, P. Multilocus half-tetrad analysis and centromere mapping in citrus: Evidence of SDR mechanism for 2n megagametophyte production and partial chiasma interference in mandarin cv “Fortune”. Heredity (Edinburgh) 2011, 107, 462–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleza, P.; Juárez, J.; Hernández, M.; Ollitrault, P.; Navarro, L. Implementation of extensive citrus triploid breeding programs based on 4x × 2x sexual hybridisations. Tree Genet. Genomes 2012, 8, 1293–1306. [Google Scholar] [CrossRef]
- Aleza, P.; Juárez, J.; Cuenca, J.; Ollitrault, P.; Navarro, L. Extensive citrus triploid hybrid production by 2x × 4x sexual hybridizations and parent-effect on the length of the juvenile phase. Plant Cell Rep. 2012, 31, 1723–1735. [Google Scholar] [CrossRef]
- Liang, M.; Cao, Z.; Zhu, A.; Liu, Y.; Tao, M.; Yang, H.; Xu, Q.; Wang, S.; Liu, J.; Li, Y.; et al. Evolution of self-compatibility by a mutant Sm -RNase in citrus. Nat. Plants 2020, 6, 131–142. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kubo, T.; Tominaga, S. Self- and cross-incompatibility of various citrus accessions. J. Jpn. Soc. Hortic. Sci. 2006, 75, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liang, M.; Wang, N.; Xu, Q.; Deng, X.; Chai, L. Reproduction in woody perennial Citrus: An update on nucellar embryony and self-incompatibility. Plant Reprod. 2018, 31, 43–57. [Google Scholar] [CrossRef]
- Vardi, A.; Neumann, H.; Frydman-Shani, A.; Yaniv, Y.; Spiegel-Roy, P. Tentative model on the inheritance of juvenility, self-incompatibility and parthenocarpy. Acta Hortic. 2000, 535, 199–206. [Google Scholar] [CrossRef]
- Esen, A.; Soost, R.K. Adventive Embroyogenesis in Citrus and its Relation to Pollination and Fertilization. Am. J. Bot. 1977, 64, 607–614. [Google Scholar] [CrossRef]
- Koltunow, A.M. Apomixis: Embryo Sacs and Embryos Formed without Meiosis or Fertilization in Ovules. Plant Cell 1993, 5, 1425–1437. [Google Scholar] [CrossRef] [Green Version]
- Vardi, A.; Levin, I.; Carmi, N.; Sciences, P.; Box, P.O. Induction of Seedlessness in Citrus: From Classical Techniques to Emerging Biotechnological Approaches. J. Amer. Soc. Hort. Sci. 2008, 133, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Ozga, J.A.; Reinecke, D.M. Hormonal Interactions in Fruit Development. J. Plant Growth Regul. 2003, 22, 73–81. [Google Scholar] [CrossRef]
- Talon, M.; Zacarias, L.; Primo-Millo, E. Hormonal changes associated with fruit set and development in mandarins differing in their parthenocarpic ability. Physiol. Plant. 1990, 79, 400–406. [Google Scholar] [CrossRef]
- Talon, M.; Zacarias, L.; Primo-Millo, E. Gibberellins and parthenocarpic ability in developing ovaries of seedless mandarins. Plant Physiol. 1992, 99, 1575–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesejo, C.; Yuste, R.; Reig, C.; Martínez-Fuentes, A.; Iglesias, D.J.; Muñoz-Fambuena, N.; Bermejo, A.; Germanà, M.A.; Primo-Millo, E.; Agustí, M. Gibberellin reactivates and maintains ovary-wall cell division causing fruit set in parthenocarpic Citrus species. Plant Sci. 2016, 247, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Mesejo, C.; Yuste, R.; Martínez-Fuentes, A.; Reig, C.; Iglesias, D.J.; Primo-Millo, E.; Agustí, M. Self-pollination and parthenocarpic ability in developing ovaries of self-incompatible Clementine mandarins (Citrus clementina). Physiol. Plant. 2013, 148, 87–96. [Google Scholar] [CrossRef]
- Sykes, S.R. Segregation in an “Imperial” mandarin X “Ellendale” tangor family for characteristics that contribute to the seedless phenotype. J. Hortic. Sci. Biotechnol. 2008, 83, 719–724. [Google Scholar] [CrossRef]
- Garcia-Papi, M.A.; Garcia-Martinez, J.L. Endogenous plant growth substances content in young fruits of seeded and seedless Clementine mandarin as related to fruit set and development. Sci. Hortic. (Amsterdam) 1984, 22, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, R.W. Horticultural Varieties of Citrus. In The Citrus Industry. Volume I. History, World Distribution, Botany, and Varieties; Reuther, W., Webber, H., Batchelor, L., Eds.; University of California: Berkeley, CA, USA, 1967; pp. 431–611. [Google Scholar]
- Sykes, S.R. The effect on Citrus fruit of excluding pollinating insects at flowering and implications for breeding new seedless cultivars. J. Hortic. Sci. Biotechnol. 2008, 83, 713–718. [Google Scholar] [CrossRef]
- Sykes, S.R.; Possingham, J. V The effect of excluding insect pollinators on seediness of Imperial mandarin fruits. Aust. J. Exp. Agric. 1992, 32, 409–411. [Google Scholar] [CrossRef]
- Vithanage, V. Incompatibility relationships among some mandarin cultivars. Plant Cell Incompat. Newsl. 1986, 18, 41–45. [Google Scholar]
- Vithanage, V. Effect of different pollen parents on seediness and quality of ‘Ellendale’ tangor. Sci. Hortic. (Amsterdam) 1991, 48, 253–260. [Google Scholar] [CrossRef]
- Wallace, H.M.; Lee, L.S. Pollen source, fruit set and xenia in mandarins. J. Hortic. Sci. Biotechnol. 1999, 74, 82–86. [Google Scholar] [CrossRef]
- Navarro, L.; Pina, J.A.; Juárez, J.; Ballester-Olmos, J.F.; Arregui, J.M.; Ortega, C.; Navarro, A.; Duran-Vila, N.; Guerri, J.; Moreno, P.; et al. The Citrus Variety Improvement Program in Spain in the Period 1975–2001; Duran-Vila, N., Milne, R., da Graça, J., Eds.; International Organization Citrus Virologists (IOCV): Riverside, CA, USA, 2002; pp. 306–316. [Google Scholar]
- Johansen, D. Plant Microtechniques; McGraw-Hill: New York, NY, USA, 1940. [Google Scholar]
- Montalt, R.; Cuenca, J.; Vives, M.C.; Navarro, L.; Ollitrault, P.; Aleza, P. Influence of temperature on the progamic phase in Citrus. Environ. Exp. Bot. 2019, 166, 103806. [Google Scholar] [CrossRef]
- Linskens, F.H.; Esser, K. Über eine spezifische anfarbung der pollenschlauche im griffel und die zahl der kallospefropfen nach slbstdung und femddung. Naturwissenschaften 1957, 44, 16. [Google Scholar] [CrossRef]
- Adhikari, P.B.; Liu, X.; Kasahara, R.D. Mechanics of Pollen Tube Elongation: A Perspective. Front. Plant Sci. 2020, 11, 1612. [Google Scholar] [CrossRef]
- Ollitrault, P.; Terol, J.; Chen, C.; Federici, C.T.; Lotfy, S.; Hippolyte, I.; Ollitrault, F.; Bérard, A.; Chauveau, A.; Cuenca, J.; et al. A reference genetic map of C. clementina hort. ex Tan.; citrus evolution inferences from comparative mapping. BMC Genomics 2012, 13, 593. [Google Scholar] [CrossRef] [PubMed]
- Eti, S.; Stosser, R. Pollen Tube Growth and Development of Ovules in Relation to Fruit Set in Mandarines, cv.’Clementine’ (Citrus reticulata Blanco); International Society for Horticultural Science (ISHS): Leuven, Belgium, 1992; pp. 621–625. [Google Scholar]
- Distefano, G.; Caruso, M.; la Malfa, S.; Gentile, A.; Tribulato, E. Histological and molecular analysis of pollen-pistil interaction in clementine. Plant Cell Rep. 2009, 28, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Distefano, G.; Las Casas, G.; La Malfa, S.; Gentile, A.; Tribulato, E.; Herrero, M. Pollen tube behavior in different Mandarin hybrids. J. Am. Soc. Hortic. Sci. 2009, 134, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Froelicher, Y.; Dambier, D.; Bassene, J.B.; Constantino, G.; Lotfy, S.; Didout, C.; Beaumont, V.; Brottier, P.; Risterucci, A.M.; Luro, F.; et al. Characterization of microsatellite markers in mandarin orange (Citrus reticulata Blanco). Mol. Ecol. Resour. 2008, 8, 119–122. [Google Scholar] [CrossRef]
- García-Lor, A.; Luro, F.; Navarro, L.; Ollitrault, P. Comparative use of InDel and SSR markers in deciphering the interspecific structure of cultivated citrus genetic diversity: A perspective for genetic association studies. Mol. Genet. Genom. 2012, 287, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Kamiri, M.; Stift, M.; Srairi, I.; Costantino, G.; El Moussadik, A.; Hmyene, A.; Bakry, F.; Ollitrault, P.; Froelicher, Y. Evidence for non-disomic inheritance in a Citrus interspecific tetraploid somatic hybrid between C. reticulata and C. limon using SSR markers and cytogenetic analysis. Plant Cell Rep. 2011, 30, 1415–1425. [Google Scholar] [CrossRef]
- Kijas, J.M.H.; Thomas, M.R.; Fowler, J.C.S.; Roose, M.L. Integration of trinucleotide microsatellites into a linkage map of Citrus. Theor. Appl. Genet. 1997, 94, 701–706. [Google Scholar] [CrossRef]
- Aleza, P.; Froelicher, Y.; Schwarz, S.; Agustí, M.; Hernández, M.; Juárez, J.; Luro, F.; Morillon, R.; Navarro, L.; Ollitrault, P. Tetraploidization events by chromosome doubling of nucellar cells are frequent in apomictic citrus and are dependent on genotype and environment. Ann. Bot. 2011, 108, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Ollitrault, P.; Ahmed, D.; Costantino, G.; Evrard, J.C.; Cardi, C.; Mournet, P.; Perdereau, A.; Froelicher, Y. Segregation distortion for male parents in high density genetic maps from reciprocal crosses between two self-incompatible cultivars confirms a gametophytic system for self-incompatibility in citrus. Agriculture 2021, 11, 379. [Google Scholar] [CrossRef]
- Kim, J.-H.; Handayani, E.; Wakana, A.; Sato, M.; Miyamoto, M.; Miyazaki, R.; Zhou, X.; Sakai, K.; Mizunoe, Y.; Shigyo, M.; et al. Distribution and evolution of Citrus accessions with S3 and/or S11 alleles for self-incompatibility with an emphasis on sweet orange [Citrus sinensis (L.) Osbeck; SfS3 or SfS3sm]. Genet. Resour. Crop Evol. 2020, 67, 2101–2117. [Google Scholar] [CrossRef]
- Ngo, B.X.; Wakana, A.; Park, S.M.; Nada, Y.; Fukudome, I. Pollen tube behaviors in self-incompatible and self-compatible Citrus cultivars. J. Fac. Agric. Kyushu Univ. 2001, 45, 443–457. [Google Scholar] [CrossRef]
- Distefano, G.; Hedhly, A.; Las Casas, G.; La Malfa, S.; Herrero, M.; Gentile, A. Male-female interaction and temperature variation affect pollen performance in Citrus. Sci. Hortic. (Amsterdam) 2012, 140, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Aloisi, I.; Distefano, G.; Antognoni, F.; Potente, G.; Parrotta, L.; Faleri, C.; Gentile, A.; Bennici, S.; Mareri, L.; Cai, G.; et al. Temperature-Dependent Compatible and Incompatible Pollen-Style Interactions in Citrus clementina Hort. ex Tan. Show Different Transglutaminase Features and Polyamine Pattern. Front. Plant Sci. 2020, 11, 1018. [Google Scholar] [CrossRef] [PubMed]
- Distefano, G.; Gentile, A.; Herrero, M. Pollen-pistil interactions and early fruiting in parthenocarpic citrus. Ann. Bot. 2011, 108, 499–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agustí, M.; García-Marí, F.; Guardiola, J.L. The influence of flowering intensity on the shedding of reproductive structures in sweet orange. Sci. Hortic. (Amsterdam) 1982, 17, 343–352. [Google Scholar] [CrossRef]
- Garcia-Papi, M.A.; Garcia-Martinez, J.L. Fruit set and development in seeded and seedless Clementine mandarin. Sci. Hortic. (Amsterdam) 1984, 22, 113–119. [Google Scholar] [CrossRef]
- Omura, M.; Shimada, T. Citrus breeding, genetics and genomics in Japan. Breed. Sci. 2016, 66, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Nishiura, M.; Shichijo, T.; Ueno, I.; Iwamasa, M.; Kihara, T.; Yamada, Y. Kiyomi tangor: A new variety of Citrus. Bull. Fruit Tree Res. Stn. B 1983, 10, 1–9. [Google Scholar]
- de Teresa, E. Mandarin treee named “Mandarin Queen”. US PP22,062 P3, pp. 1–22. Available online: https://patents.google.com/patent/USPP22062P3/en (accessed on 9 August 2011).
- Tribulato, E.; La Rosa, G. Primosole e Simeto: Due nuovi ibridi di mandarino. Italus Hortus 1993, 1, 21–25. [Google Scholar]
- Chasan, R.; Walbot, V. Mechanisms of Plant Reproduction: Questions and Approaches. Plant Cell 1993, 5, 1139–1146. [Google Scholar] [CrossRef]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. Global warming and sexual plant reproduction. Trends Plant Sci. 2009, 14, 30–36. [Google Scholar] [CrossRef]
- Iizumi, T.; Furuya, J.; Shen, Z.; Kim, W.; Okada, M.; Fujimori, S.; Hasegawa, T.; Nishimori, M. Responses of crop yield growth to global temperature and socioeconomic changes. Sci. Rep. 2017, 7, 7800. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [Green Version]
- Koti, S.; Reddy, K.R.; Reddy, V.R.; Kakani, V.G.; Zhao, D. Interactive effects of carbon dioxide, temperature, and ultraviolet-B radiation on soybean (Glycine max L.) flower and pollen morphology, pollen production, germination, and tube lengths. J. Exp. Bot. 2005, 56, 725–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Distefano, G.; Gentile, A.; Hedhly, A.; La Malfa, S. Temperatures during flower bud development affect pollen germination, self-incompatibility reaction and early fruit development of clementine (Citrus clementina Hort. ex Tan.). Plant Biol. 2018. [Google Scholar] [CrossRef]
- Lora, J.; Herrero, M.; Hormaza, J.I. The coexistence of bicellular and tricellular pollen in Annona cherimola (Annonaceae): Implications for pollen evolution. Am. J. Bot. 2009, 96, 802–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lora, J.; Herrero, M.; Hormaza, J.I. Stigmatic receptivity in a dichogamous early-divergent angiosperm species, Annona cherimola (Annonaceae): Influence of temperature and humidity. Am. J. Bot. 2011, 98, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Montalt, R.; Ollitrault, P.; Navarro, L.; Aleza, P. Evaluation of parthenocarpy ability in different citrus genotypes. In Proceedings of the 13 th International Citrus Congress, IAPAR, Foz do Iguaçu, Brazil; p. 100. Available online: https://www.semanticscholar.org/paper/Evaluation-of-parthenocarpy-ability-in-different-Montalt-Ollitrault/b706e659355dd0e57951258326d65abd6233c5d1 (accessed on 6 October 2021).
- Roose, M.L.; Williams, T.E. Mutation Breeding. In Citrus Genetics, Breeding and Biotechnology; Khan, I., Ed.; CABI: Wallingfor, UK, 2007; pp. 345–352. [Google Scholar]
- Caruso, M.; Smith, M.; Froelicher, Y.; Russo, G.; Gmitter, F. Traditional breeding. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F.G., Eds.; Woodhead Publishing: Duxford, UK, 2020; pp. 129–148. ISBN 9780128121634. [Google Scholar]
- Garmendia, A.; Beltrán, R.; Zornoza, C.; García-Breijo, F.J.; Reig, J.; Merle, H. Gibberellic acid in Citrus spp. flowering and fruiting: A systematic review. PLoS ONE 2019, 14, e0223147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
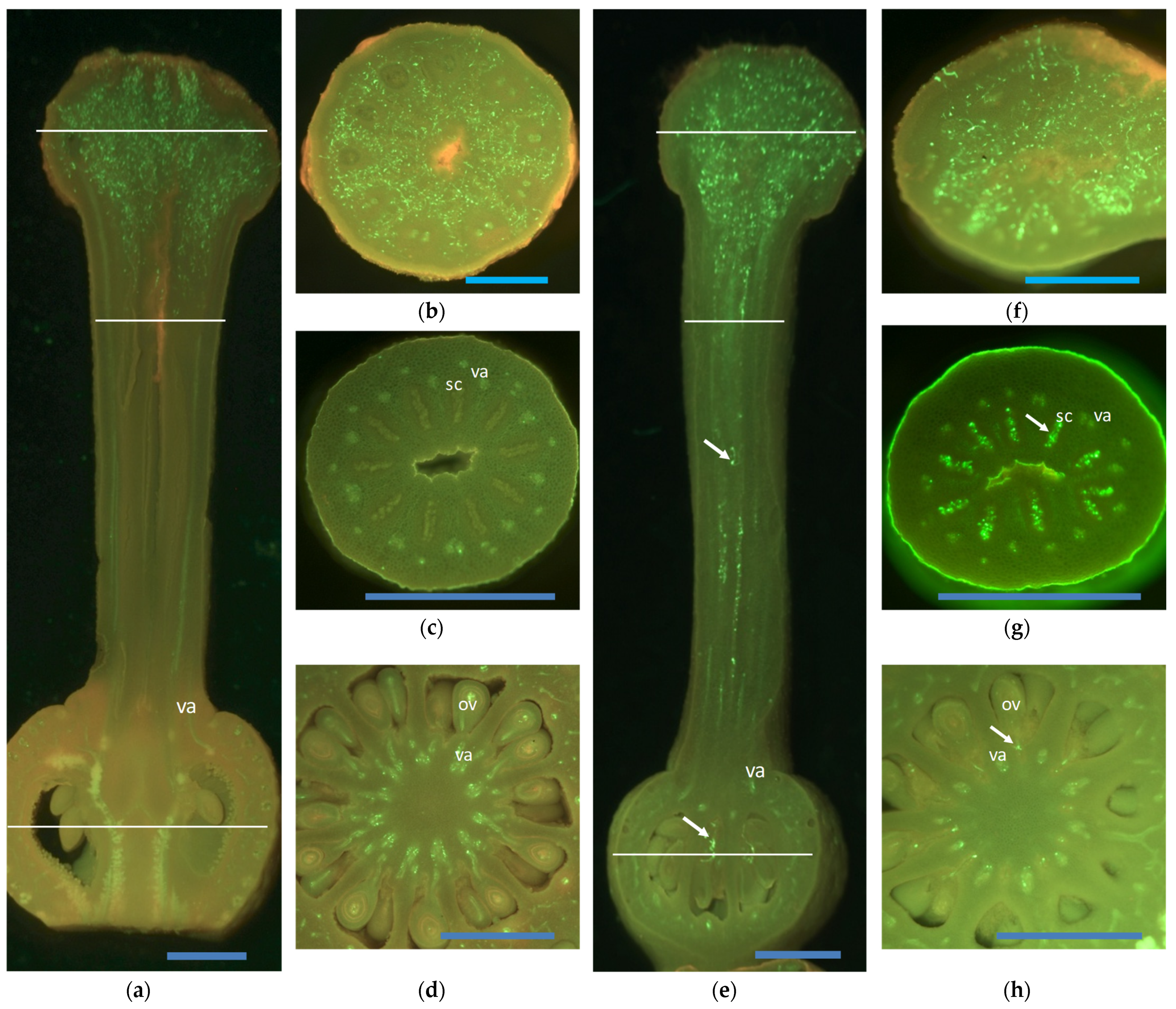
Figure 1.
Histological sections of self-pollinated pistils. (a–d) Self-incompatible pistil 10 days after self-pollination. (a) Longitudinal section of the entire pistil in which high quantity pollen tubes were observed thorough the stigma whose growth further stopped in the upper style. (e–h) Self-compatible pistil 10 days after self-pollination. (e) Longitudinal section of the entire pistil in which high quantity pollen tubes were observed throughout the stigma, along the entire style and reached ovaries. The white lines on the longitudinal pistil sections indicate the transversal section that corresponds to the middle stigma (b,f), upper style (c,g), and ovary (d,h). Pollen tubes are marked by an arrow; va: vascular axis; sc: stylar canal; ov: ovule. Scale bars are depicted by blue lines: (a,e) 1 mm; (b–g) 0.5 mm. Pictures of ‘Fortune’ mandarin and ‘Monreal’ clementine are shown as examples of self-incompatible and self-compatible varieties, respectively.
Figure 1.
Histological sections of self-pollinated pistils. (a–d) Self-incompatible pistil 10 days after self-pollination. (a) Longitudinal section of the entire pistil in which high quantity pollen tubes were observed thorough the stigma whose growth further stopped in the upper style. (e–h) Self-compatible pistil 10 days after self-pollination. (e) Longitudinal section of the entire pistil in which high quantity pollen tubes were observed throughout the stigma, along the entire style and reached ovaries. The white lines on the longitudinal pistil sections indicate the transversal section that corresponds to the middle stigma (b,f), upper style (c,g), and ovary (d,h). Pollen tubes are marked by an arrow; va: vascular axis; sc: stylar canal; ov: ovule. Scale bars are depicted by blue lines: (a,e) 1 mm; (b–g) 0.5 mm. Pictures of ‘Fortune’ mandarin and ‘Monreal’ clementine are shown as examples of self-incompatible and self-compatible varieties, respectively.

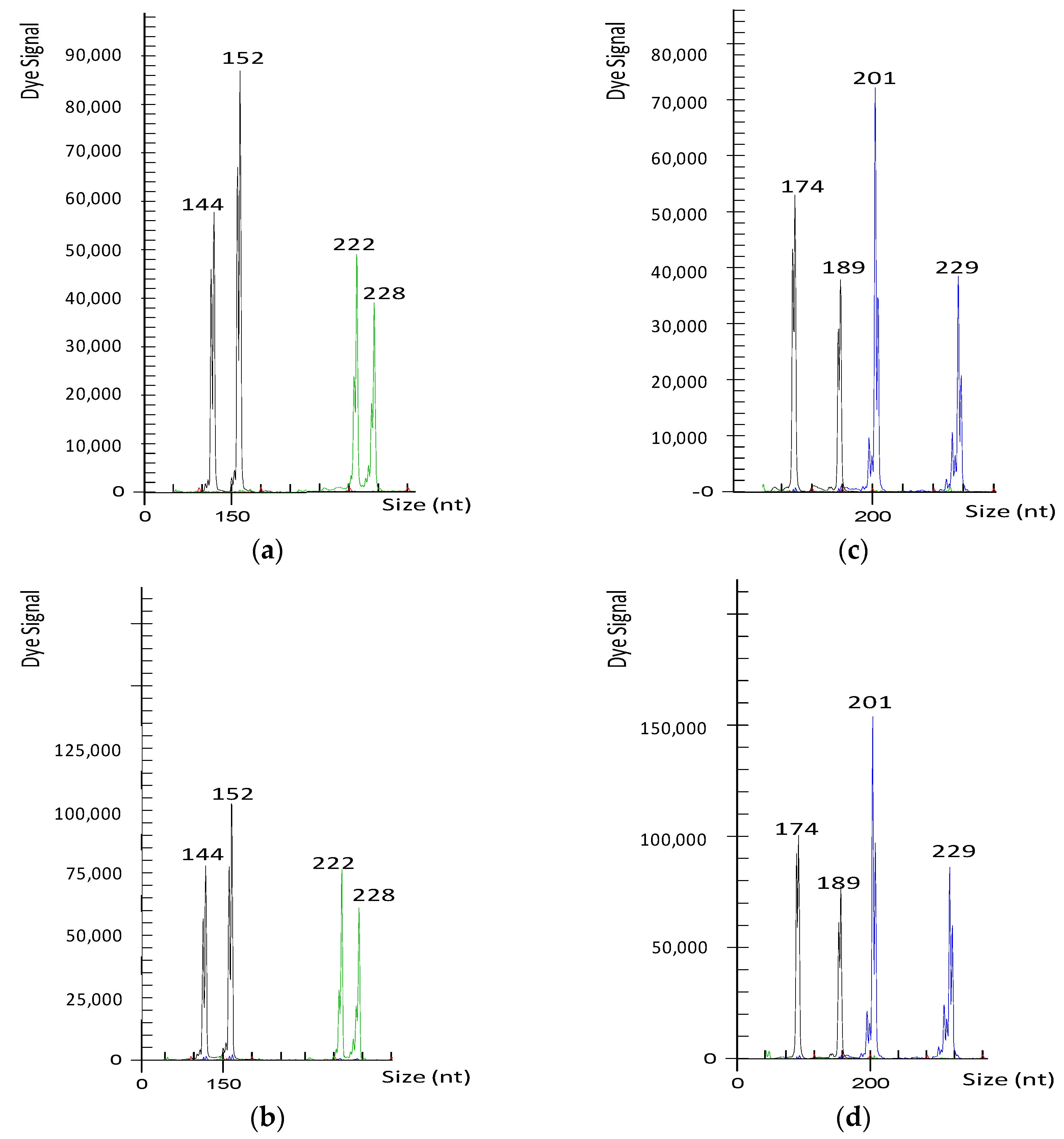
Figure 2.
Electropherograms obtained using the mCrCIR05A05 (black) and mCrCIR03G05 (green) SSR markers for ‘Imperial’ mandarin (a), a diploid hybrid recovered from the self-pollination of ‘Imperial’ (b), the MEST15 (black) and Ci02D04 (blue) loci for ‘Ellendale’ tangor (c), and a diploid hybrid recovered from the self-pollination of ‘Ellendale’ tangor (d). Numbers indicate the size of alleles in nucleotides (nt) for each variety.
Figure 2.
Electropherograms obtained using the mCrCIR05A05 (black) and mCrCIR03G05 (green) SSR markers for ‘Imperial’ mandarin (a), a diploid hybrid recovered from the self-pollination of ‘Imperial’ (b), the MEST15 (black) and Ci02D04 (blue) loci for ‘Ellendale’ tangor (c), and a diploid hybrid recovered from the self-pollination of ‘Ellendale’ tangor (d). Numbers indicate the size of alleles in nucleotides (nt) for each variety.

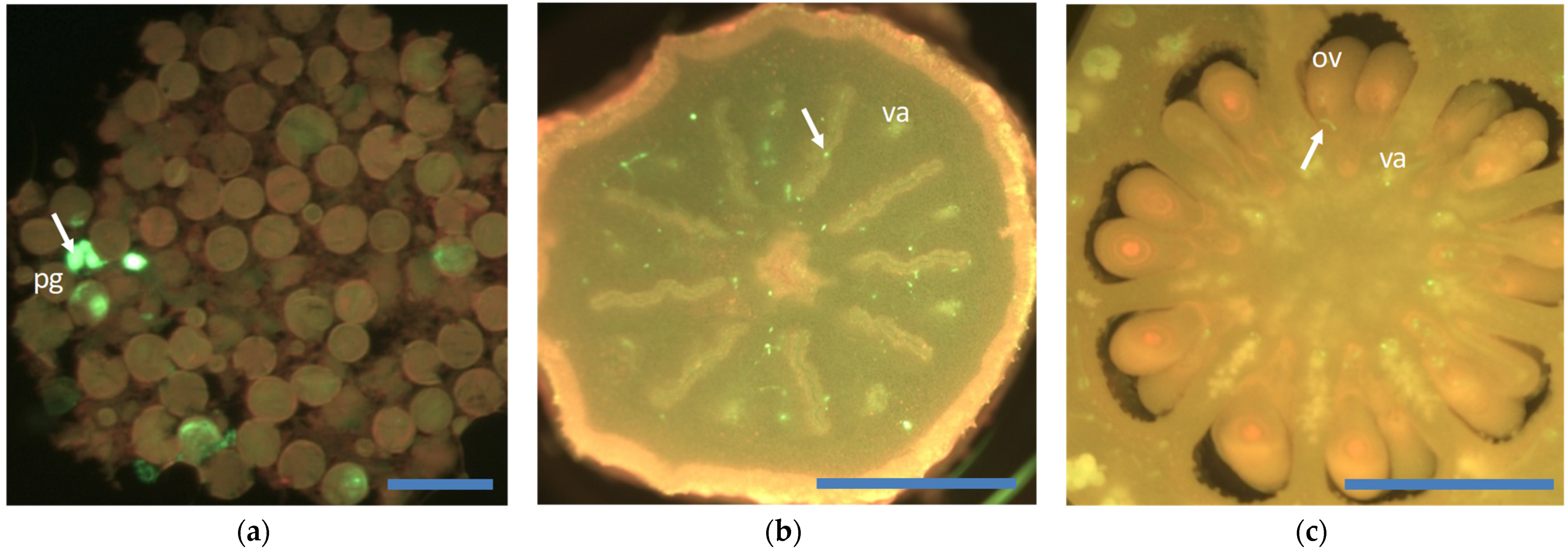
Figure 3.
Histological observations in the ‘Monreal’ clementine x ‘Oroblanco’ triploid hybrid 10 days after pollination. (a) Large quantity of pollen grains observed on the stigma surface displaying poor pollen germination; (b) very low quantity pollen tubes growing throughout the bottom half of the stigma; (c) one pollen tube reached ovules. Pollen tubes marked by an arrow; pg: pollen grain; va: vascular axis; ov: ovule. Scale bars are denoted by blue lines: (a) 100 µm; (b) and (c) 1 mm.
Figure 3.
Histological observations in the ‘Monreal’ clementine x ‘Oroblanco’ triploid hybrid 10 days after pollination. (a) Large quantity of pollen grains observed on the stigma surface displaying poor pollen germination; (b) very low quantity pollen tubes growing throughout the bottom half of the stigma; (c) one pollen tube reached ovules. Pollen tubes marked by an arrow; pg: pollen grain; va: vascular axis; ov: ovule. Scale bars are denoted by blue lines: (a) 100 µm; (b) and (c) 1 mm.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Self-compatibility (SC) or self-incompatibility (SI) classification based on pollen tube growth and number of seeds per fruit obtained from hand self-pollination (SP) and hand cross-pollination (CP) treatments in each variety.
Table 1.
Self-compatibility (SC) or self-incompatibility (SI) classification based on pollen tube growth and number of seeds per fruit obtained from hand self-pollination (SP) and hand cross-pollination (CP) treatments in each variety.
| Pollen Tubes Growth | Number of Seeds per Fruit | ||||
|---|---|---|---|---|---|
| SC/SI | SP | CP | SP | CP | |
| ‘Clemenules’ | SI | 0 | 100 | 0 | 24.3 ± 2.6 |
| ‘Monreal’ | SC | 100 | 100 | 22.5 ± 3.6 (a) | 21.0 ± 4.4 (a) |
| ‘Campeona’ | SC | 100 | 100 | 4.9 ± 1.8 (a) | 9.9 ± 2.9 (b) |
| ‘Imperial’ | SI | 15 | 100 | 0.8 ± 1.4 (a) | 9.3 ± 2.5 (b) |
| ‘Salteñita’ | SC | 100 | 100 | 11.0 ± 4.3 (a) | 16.5 ± 4.1 (b) |
| ‘Fortune’ | SI | 0 | 100 | 0 | 20.4 ± 5.6 |
| ‘Moncada’ | SI | 0 | 100 | 0 | 11.6 ± 5.2 |
| ‘Ellendale’ | SI | 10 | 100 | 0.7 ± 1.0 (a) | 33.1 ± 5.8 (b) |
| ‘Serafines’ | NA | 0 | 100 | 0 | 5.3 ± 2.1 |
Pollen tubes growth is expressed as the percentage of pistils in which pollen tubes were observed reaching the ovaries. Number of seeds per fruit are given as the mean ± SD (n = 16 to 40 depending on the number of fruits obtained in the treatment). For each variety, significant differences (p = 0.05 Fisher LSD) between treatments are indicated by different letters in brackets.
Table 2.
Fruit setting percentage obtained in each treatment.
| E | SP | CP | |
|---|---|---|---|
| ‘Clemenules’ | 15 (a) | 16 (a) | 84 (b) |
| ‘Monreal’ | 0 | 68 (a) | 74 (a) |
| ‘Campeona’ | 9 (a) | 42 (b) | 56 (c) |
| ‘Imperial’ | 19 (a) | 33 (b) | 67 (c) |
| ‘Salteñita’ | 5 (a) | 58 (b) | 70 (c) |
| ‘Fortune’ | 0 | 9 (a) | 63 (b) |
| ‘Moncada’ | 34 (a) | 33 (a) | 72 (b) |
| ‘Ellendale’ | 5 (a) | 5 (a) | 60 (b) |
| ‘Serafines’ | 64 (a) | 66 (a) | 65 (a) |
E: Emasculation; SP: hand self-pollination; CP: hand cross-pollination. For each variety, significant differences (95% confidence interval for fruit setting proportions) between treatments are indicated by different letters in brackets.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Montalt, R.; Vives, M.C.; Navarro, L.; Ollitrault, P.; Aleza, P. Parthenocarpy and Self-Incompatibility in Mandarins. Agronomy 2021, 11, 2023. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102023
AMA Style
Montalt R, Vives MC, Navarro L, Ollitrault P, Aleza P. Parthenocarpy and Self-Incompatibility in Mandarins. Agronomy. 2021; 11(10):2023. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102023
Chicago/Turabian StyleMontalt, Rafael, María Carmen Vives, Luis Navarro, Patrick Ollitrault, and Pablo Aleza. 2021. "Parthenocarpy and Self-Incompatibility in Mandarins" Agronomy 11, no. 10: 2023. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102023
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.