
Predicting the Emergence of Echinochloa crus-galli (L.) P. Beauv. in Maize Crop in Croatia with Hydrothermal Model


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Laboratory Experiments—Estimation of Base Temperature and Base Water Potential
2.2. Field Experiments and Laboratory Analyses
2.2.1. Monitoring of E. crus-galli Emergence in Maize
2.2.2. Soil Analysis
2.3. Statistical Analysis
Tsi < To: n = 0 if Ψsi ≤ Ψb; n = 1 if Ψsi > Ψb
Tsi > To: n = 0 if Ψsi ≤ Ψb + Kt (Tsi − To); n = 1 if Ψsi> Ψb + Kt (Tsi − To)
Model Validation
3. Results and Discussion
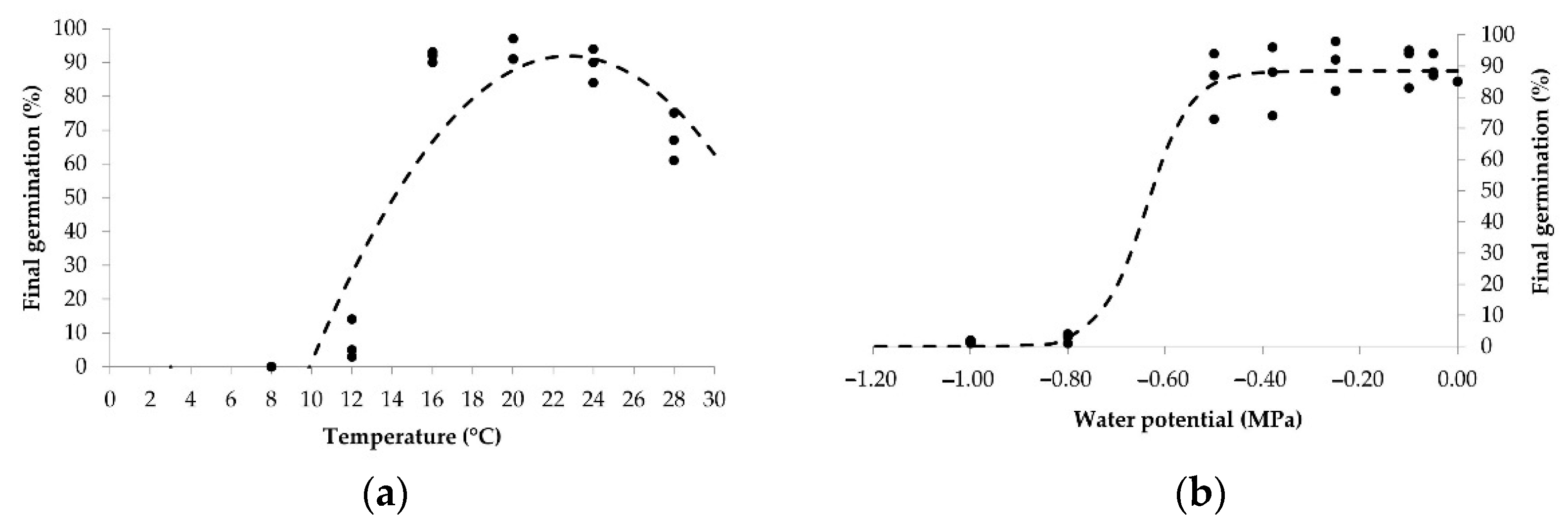
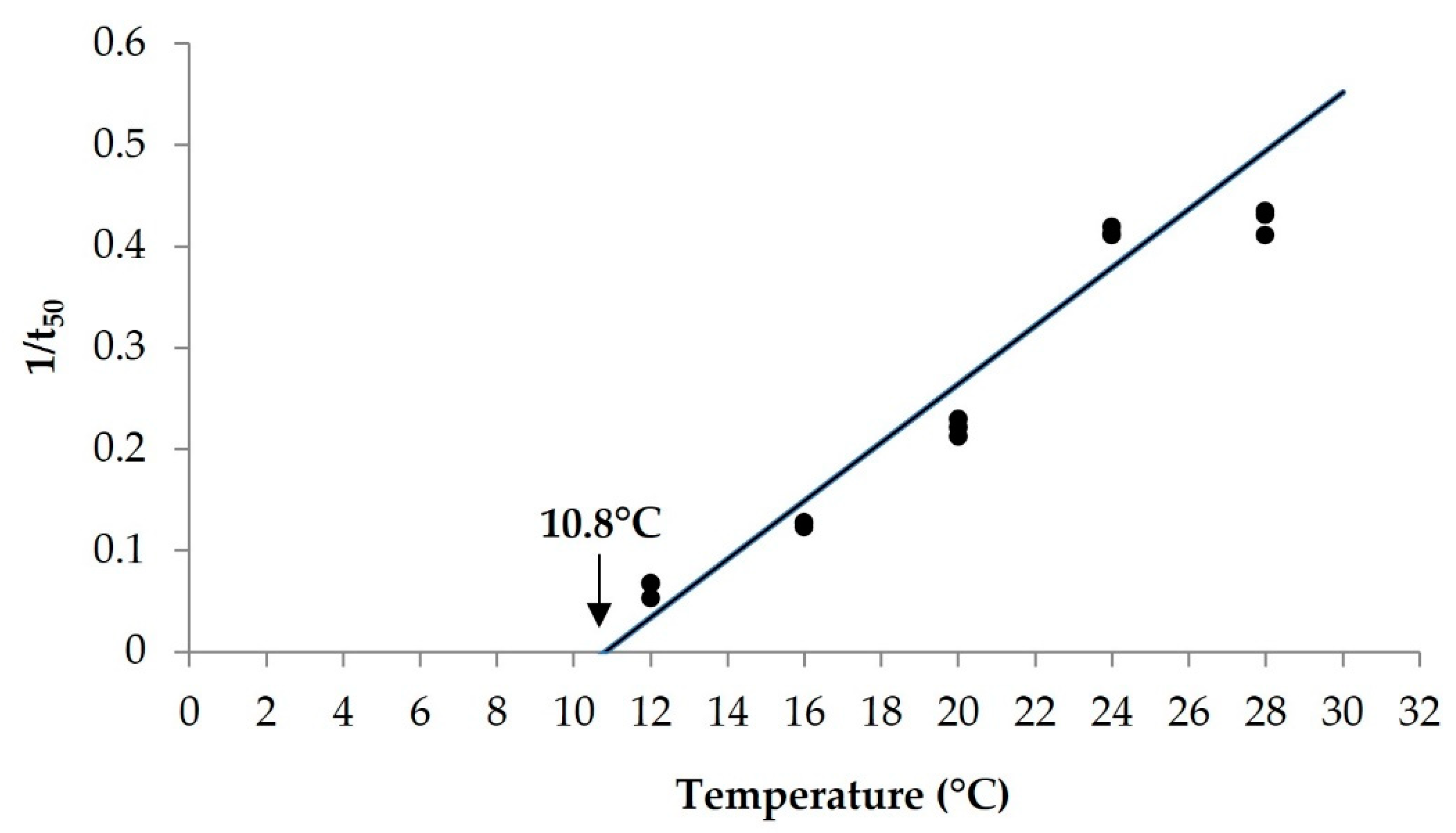
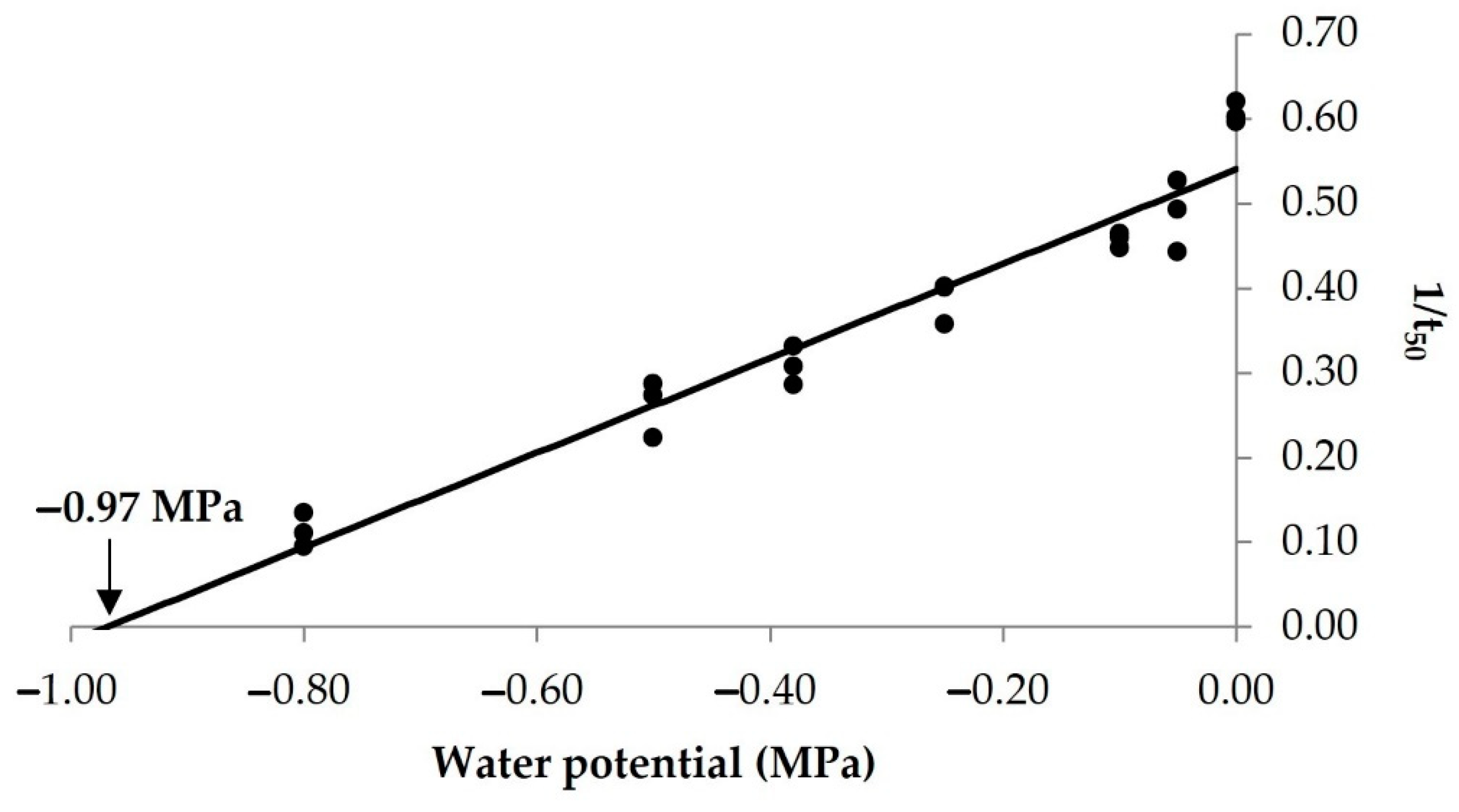
3.1. Estimation of Base Temperature and Base Water Potential
3.2. Field Experiments
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grundy, A.C. Predicting weed emergence: A review of approaches and future challenges. Weed Res. 2003, 43, 1–11. [Google Scholar] [CrossRef]
- Vleeshouwers, L.M.; Kropff, M.J. Modelling field emergence patterns in arable weeds. New Phytologist. 2000, 148, 445–457. [Google Scholar] [CrossRef]
- Merfield, C.N. False and Stale Seedbeds: The Most Effective Non-Chemical WeedManagement Tools for Cropping and Pasture Establishment; The BHU Future Farming Centre: Lincoln, New Zealand, 2013; p. 23. [Google Scholar]
- Mohler, C.L. Weed life history: Identifying vulnerabilities. In Ecological Management of Agricultural Weeds; Liebman, M., Mohler, C.L., Staver, C.P., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 40–98. [Google Scholar] [CrossRef]
- Forcella, F.; Benech-Arnold, R.L.; Sanchez, R.; Ghersa, C.M. Modeling seedling emergence. Field Crop Res. 2000, 67, 123–139. [Google Scholar] [CrossRef]
- García, A.L.; Recasens, J.; Forcella, F.; Torra, J.; Royo-Esnal, A. Hydrothermal Emergence Model for Ripgut Brome (Bromus diandrus). Weed Sci. 2013, 61, 146–153. [Google Scholar] [CrossRef]
- Masin, R.; Loddo, D.; Benvenuti, S.; Zuin, M.C.; Macchia, M.; Zanin, G. Temperature and Water Potential as Parameters for Modeling Weed Emergence in Central-Northern Italy. Weed Sci. 2010, 58, 216–222. [Google Scholar] [CrossRef]
- Leguizamon, E.S.; Fernandez-Quintanilla, C.; Barroso, J.; Gonzalez-Andujar, J.L. Using thermal and hydrothermal time to model seedling emergence of Avena sterilis spp. ludoviciana in Spain. Weed Res. 2005, 45, 149–156. [Google Scholar] [CrossRef]
- Gummerson, R.J. The effect of constant temperatures and osmotic potentials on the germination of sugar beet. J. Exp. Bot. 1986, 37, 729–741. [Google Scholar] [CrossRef]
- Bradford, K.J. Water relations in seed germination6. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1995; pp. 351–396. [Google Scholar]
- Masin, R.; Loddo, D.; Benvenuti, S.; Otto, S.; Zanin, G. Modeling Weed Emergence in Italian Maize Fields. Weed Sci. 2012, 60, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Archer, D.W.; Forcella, F.; Eklund, J.J.; Gunsolus, J. WeedCast Version 2.0. 2001. Available online: http://www.morris.ars.usda.gov (accessed on 15 July 2021).
- Loddo, D.; Bozic, D.; Calha, I.M.; Dorado, J.; Izquierdo, J.; Šćepanović, M.; Barić, K.; Carlesi, S.; Leskovsek, R.; Peterson, D.; et al. Variability in seedling emergence for European and North American populations of Abutilon theophrasti. Weed Res. 2019, 59, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Leiblein-Wild, M.C.; Kaviani, R.; Tackenberg, O. Germination and seedling frost tolerance differ between the native and invasive range in common ragweed. Oecologia 2014, 174, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Bürger, J.; Colbach, N. Germination base temperature and relative growth rate of 13 weed species—Comparing populations from two geographical origins. In Proceedings of the 28th German Conference on Weed Biology and Weed Control, Braunschweig, Germany, 27 February–1 March 2018; Volume 458, pp. 419–426. [Google Scholar] [CrossRef]
- Leblanc, M.L.; Cloutier, D.C.; Stewart, K.A.; Hamel, C. Calibration and validation of a common lambsquarters (Chenopodium album) seedling emergence model. Weed Sci. 2004, 52, 61–66. [Google Scholar] [CrossRef]
- Holm, L.; Doll, J.; Holm, E.; Pancho, J.; Herberger, J. World Weeds: Natural Histories and Distribution; John Wiley & Sons: New York, NY, USA, 1997. [Google Scholar]
- Šarić, T.; Ostojić, Z.; Stefanović, L.; Deneva Milanova, S.; Kazinczi, G.; Tyšer, L. The changes of the composition of weed flora in southeastern and central europe as affected by cropping practices. Herbologia 2011, 12, 8–12. [Google Scholar]
- Šćepanović, M.; Ostojić, Z.; Barić, K. Ograničenja mogućnosti suzbijanja korova u soji nakon nicanja. In Zbornik Sažetaka 56; seminara biljne zaštite. Cvjetković, B. (ur.); Hrvatsko društvo biljne zaštite: Opatija, Croatia, 2012; pp. 27–28. [Google Scholar]
- Bosnic, A.C.; Swanton, C.J. Influence of barnyardgrass (Echinochloa crus-galli) time of emergence and density on corn (Zea mays). Weed Sci. 1997, 45, 276–282. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The Osmotic Potential of Polyethylene Glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef] [PubMed]
- Šoštarčić, V.; Masin, R.; Loddo, D.; Brijačak, E.; Šćepanović, M. Germination parameters of selected summer weeds: Transferring of the Alertinf model to other geographical regions. Agronomy 2021, 11, 292. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports; No. 106; FAO: Rome, Italy, 2014. [Google Scholar]
- FAO. Guidelines for Soil Description, 4th ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2006; p. 97. [Google Scholar]
- Pintar, A.; Stipičević, S.; Lakić, J.; Barić, K. Phytotoxicity of mesotrione residues on sugar beet (Beta vulgaris L.) in agricultural soils differing in adsorption affinity. Sugar Tech. 2020, 22, 137–142. [Google Scholar] [CrossRef]
- van Genuchten, M.T.; Leij, F.J.; Yates, S.R. The RETC Code for Quantifying the Hydraulic Functions of Unsaturated Soils, Version 1.0; EPA Report 600/2-91/065; U.S. Salinity Laboratory, USDA, ARS: Riverside, CA, USA, 1991.
- Onofri, A. Bioassay97: A new Excel VBA macro to perform statistical analyses on herbicide dose-response data. Ital. J. Agrometeorol. 2001, 3, 40–45. [Google Scholar]
- Efron, B. Bootstrap methods: Another look at the jackknife. Ann. Stat. 1979, 7, 1–26. [Google Scholar] [CrossRef]
- Masin, R.; Zuin, M.C.; Zanin, G.; Tridello, G. Weed Turf: Software for improving summer annual weed control in turf. Ital. J. Agrometeorol. 2005, 50, 46–50. [Google Scholar]
- Masin, R.; Loddo, D.; Gasparini, V.; Otto, S.; Zanin, G. Evaluation of Weed Emergence Model AlertInf for Maize in Soybean. Weed Sci. 2014, 62, 360–369. [Google Scholar] [CrossRef]
- Loddo, D.; Ghaderi-Far, F.; Rastegar, Z.; Masin, R. Base temperatures for germination of selected weed species in Iran. Plant Prot. Sci. 2018, 54, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Wiese, A.M.; Binning, L.K. Calculating the Threshold Temperature of Development for Weeds. Weed Sci. 1987, 35, 177–179. [Google Scholar] [CrossRef]
- Guillemin, J.P.; Gardarin, A.; Granger, S.; Reibel, C.; Munier-Jolain, N.; Colbach, N. Assessing potential germination period of weeds with base temperatures and base water potentials. Weed Res. 2013, 53, 76–87. [Google Scholar] [CrossRef]
- Steinmaus, S.J.; Prather, T.S.; Holt, J.S. Estimation of base temperatures for nine weed species. J. Exp. Bot. 2000, 51, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Koppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Zambrano-Navea, C.; Bastida, F.; Gonzalez-Andujar, J.L. A hydrothermal seedling emergence model for Conyza bonariensis. Weed Res. 2013, 53, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Vasileiadis, V.P.; Froud-Williams, R.J.; Loddo, D.; Eleftherohorinos, I.G. Emergence dynamics of barnyardgrass and jimsonweed from two depths when switching from conventional to reduced and no-till conditions. Span. J. Agric. 2016, 14, e1002. [Google Scholar] [CrossRef] [Green Version]
- Werle, R.; Sandell, L.D.; Buhler, D.D.; Hartzler, R.G.; Lindquist, J.L. Predicting emergence of 23 summer annual weed species. Weed Sci. 2014, 62, 267–279. [Google Scholar] [CrossRef]
- Bagavathiannan, M.V.; Norsworthy, J.K.; Smith, K.L.; Burgos, N. Seedbank Size and Emergence Pattern of Barnyardgrass (Echinochloa crusgalli) in Arkansas. Weed Sci. 2011, 59, 359–365. [Google Scholar] [CrossRef]
- Ramanarayanan, T.S.; Williams, J.R.; Dugas, W.A.; Hauck, L.M.; McFarland, A.M.S. Using APEX to Identify Alternative Practices for Animal Waste Management. Part I: Model Description and Validation; ASAE Paper No. 972209; American Society of Agricultural Engineers: St. Joseph, MI, USA, 1997. [Google Scholar]
- Egea-Cobrero, V.; Bradley, K.; Calha, I.M.; Davis, A.S.; Dorado, J.; Forcella, F.; Lindquist, J.L.; Sprague, C.L.; Gonzalez-Andujar, J.L. Validation of predictive empirical weed emergence models of Abutilon theophrasti Medik based on intercontinental data. Weed Res. 2020, 60, 297–302. [Google Scholar] [CrossRef]
- Myers, M.W.; Curran, W.S.; VanGessel, M.J.; Calvin, D.D.; Mortensen, D.A.; Majek, B.A. Predicting weed emergence for eight annual species in the northeastern United States. Weed Sci. 2004, 52, 913–919. [Google Scholar] [CrossRef]
- Oriade, C.; Forcella, F. Maximizing efficacy and economics of mechanical weed control in row crops through forecasts of weed emergence. J. Crop Prod. 1999, 2, 189–205. [Google Scholar] [CrossRef]
- Otto, S.; Masin, R.; Casari, G.; Zanin, G. Weed–Corn Competition Parameters in Late-Winter Sowing in Northern Italy. Weed Sci. 2009, 57, 194–201. [Google Scholar] [CrossRef]
- Loddo, D.; Scarabel, L.; Sattin, M.; Pederzoli, A.; Morsiani, C.; Canestrale, R.; Tommasini, M.G. Combination of herbicide band application and inter-row cultivation provides sustainable weed control in maize. Agronomy 2020, 10, 20. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Period | Average Air Temperature (°C) | Average Soil Temperature (°C) | Precipitation (mm) |
|---|---|---|---|
| 2019 | |||
| 8–31 May | 13.0 | 15.2 | 54.8 |
| 1–30 June | 22.6 | 24.9 | 85.6 |
| 1–5 July | 22.0 | 22.6 | 12.4 |
| 2020 | |||
| 5–31 May | 15.3 | 17.8 | 58.6 |
| 1–29 June | 19.3 | 22.1 | 85.6 |
| Soil Physicochemical Properties a | Soil Water Retention Properties (Vol %) b | ||||||
|---|---|---|---|---|---|---|---|
| Texture | Organic matter % | CaCO3 % | pHKCl | Bulk density g cm−3 | Field capacity | Plant wilting point | Plant- available water |
| Silt loam [25] | 2.7 [25] | 7.0 | 7.28 [25] | 1.16 | 44.0 | 20.6 | 23.4 |
| °C | t10 | t50 | t90 | MPa | t10 | t50 | t90 |
|---|---|---|---|---|---|---|---|
| 28 | 1.6 a | 2.4 a | 3.4 a | 0.00 | 1.3 a | 1.6 a | 2.1 a |
| 24 | 3.2 b | 3.8 ab | 4.6 ab | −0.05 | 1.6 b | 2.2 ab | 3.0 a |
| 20 | 3.1 b | 4.6 b | 6.7 b | −0.10 | 1.8 b | 2.6 ab | 3.6 a |
| 16 | 5.2 e | 8.1 c | 12.6 c | −0.25 | 2.3 c | 3.3 bc | 4.7 a |
| 12 | 13.9 d | 16.2 d | 18.7 d | −0.38 | 2.6 e | 3.6 bc | 4.3 a |
| −0.50 | 2.9 d | 3.9 c | 5.8 a | ||||
| −0.80 | 1.6 b | 9.0 d | 50.0 b |
| Population | Tb (°C) | ±95 CI | r2 | Ψb (MPa) | ±95 CI | r2 |
|---|---|---|---|---|---|---|
| Croatia | 10.8 | 0.27 | 0.94 | −0.97 | 0.06 | 0.94 |
| Italy [7] | 11.7 | 0.28 | 0.89 | −0.97 | 0.04 | 0.95 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šoštarčić, V.; Masin, R.; Loddo, D.; Svečnjak, Z.; Rubinić, V.; Šćepanović, M. Predicting the Emergence of Echinochloa crus-galli (L.) P. Beauv. in Maize Crop in Croatia with Hydrothermal Model. Agronomy 2021, 11, 2072. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102072
Šoštarčić V, Masin R, Loddo D, Svečnjak Z, Rubinić V, Šćepanović M. Predicting the Emergence of Echinochloa crus-galli (L.) P. Beauv. in Maize Crop in Croatia with Hydrothermal Model. Agronomy. 2021; 11(10):2072. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102072
Chicago/Turabian StyleŠoštarčić, Valentina, Roberta Masin, Donato Loddo, Zlatko Svečnjak, Vedran Rubinić, and Maja Šćepanović. 2021. "Predicting the Emergence of Echinochloa crus-galli (L.) P. Beauv. in Maize Crop in Croatia with Hydrothermal Model" Agronomy 11, no. 10: 2072. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11102072