
Association Study of Symbiotic Genes in Pea (Pisum sativum L.) Cultivars Grown in Symbiotic Conditions

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Microorganisms
2.3. Vegetation Experiment for Assesment of the Symbiotic Effectiveness of 99 Pea Cultivars
2.4. SNVs Detection and Cultivar Screening

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Identifier | Accession | Description |
|---|---|---|
| IGN1 [28] | KR047192 | Ankyrin-repeat protein with transmembrane regions |
| GICP01056763 1 | ||
| SEN1 [29] | KY888171 | Control of rhizobial differentiation |
| GDTM01047803 2 | ||
| SST1 [28] | KC008603 | Symbiotic sulphate transporter |
| GDTM01043951 2 | ||
| Sym29 | AJ495759 | CLAVATA1-like receptor |
| GICP01072631 1 | ||
| STR1 | GICP01133083 1 | ABC transporter G family member STR, required for arbuscule development |
| STR2 | GICP01179536 1 | ABC transporter G family member STR, required for arbuscule development |
| AMT2;3 | PsCam053936 3 | Ammonium transporter |
| AMT2;5 | GICP01050721 1 | Ammonium transporter |
2.5. Statistical Analysis
2.6. Analysis of AM Development
3. Results
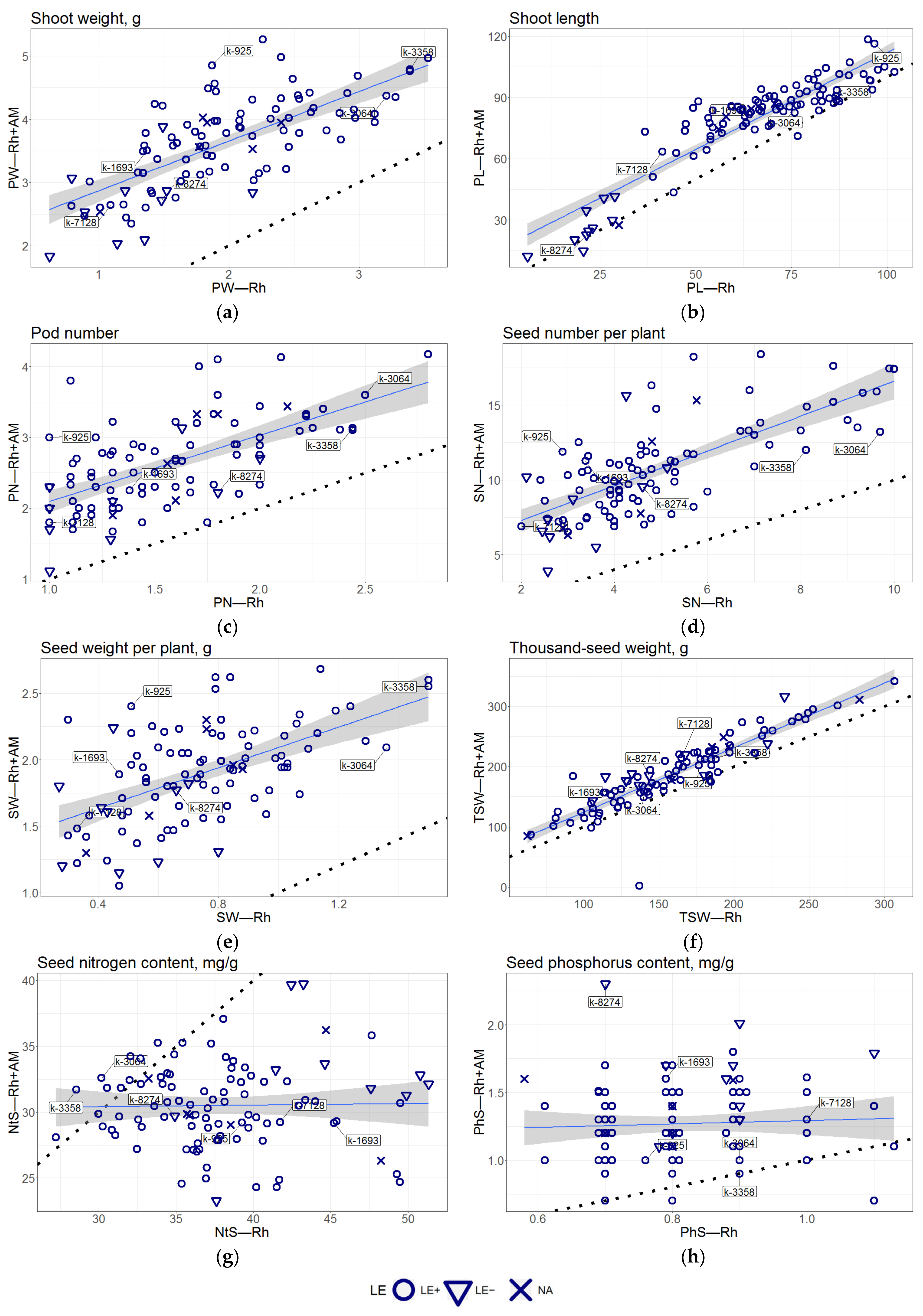
3.1. General Description of the Dataset
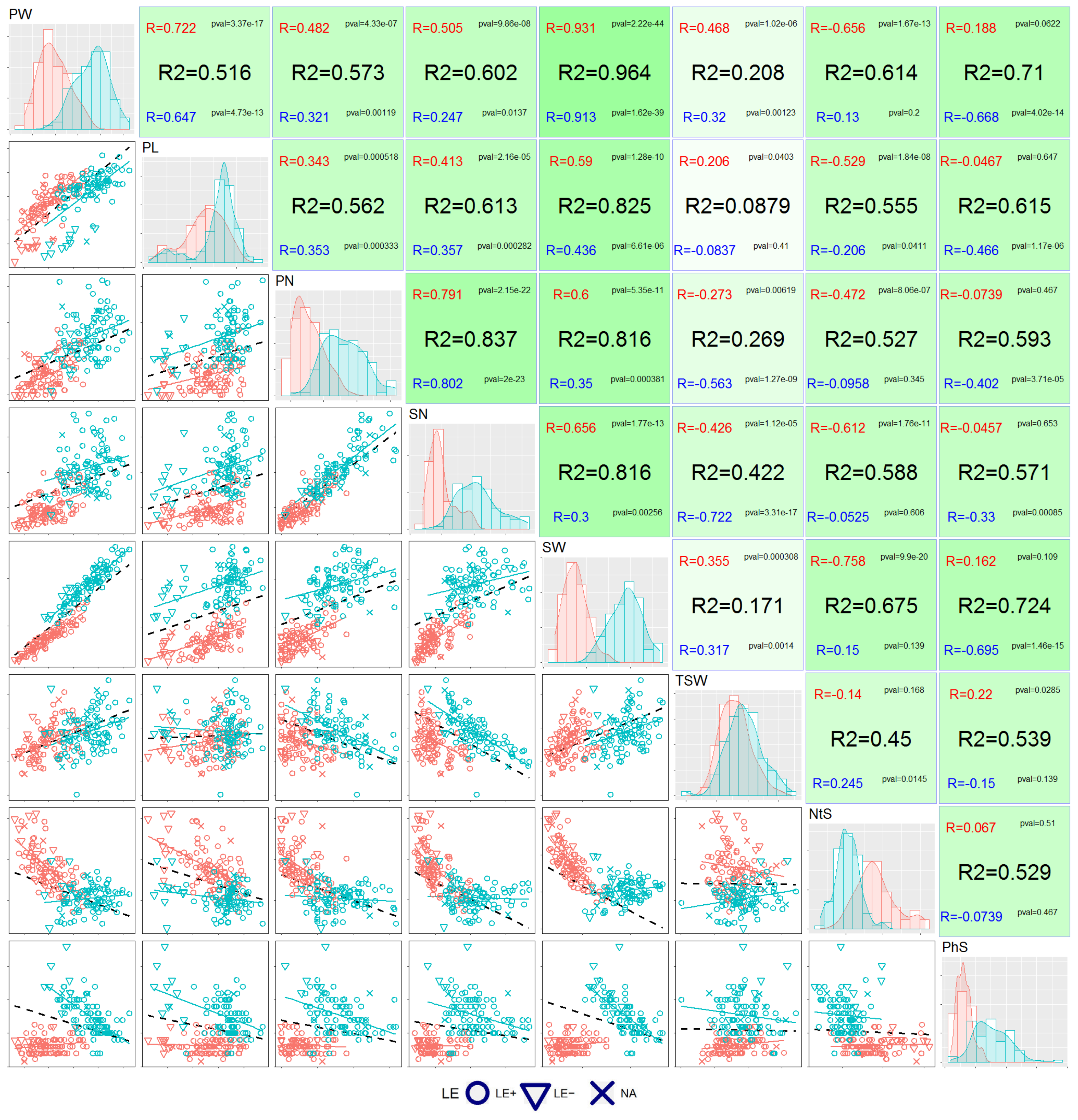
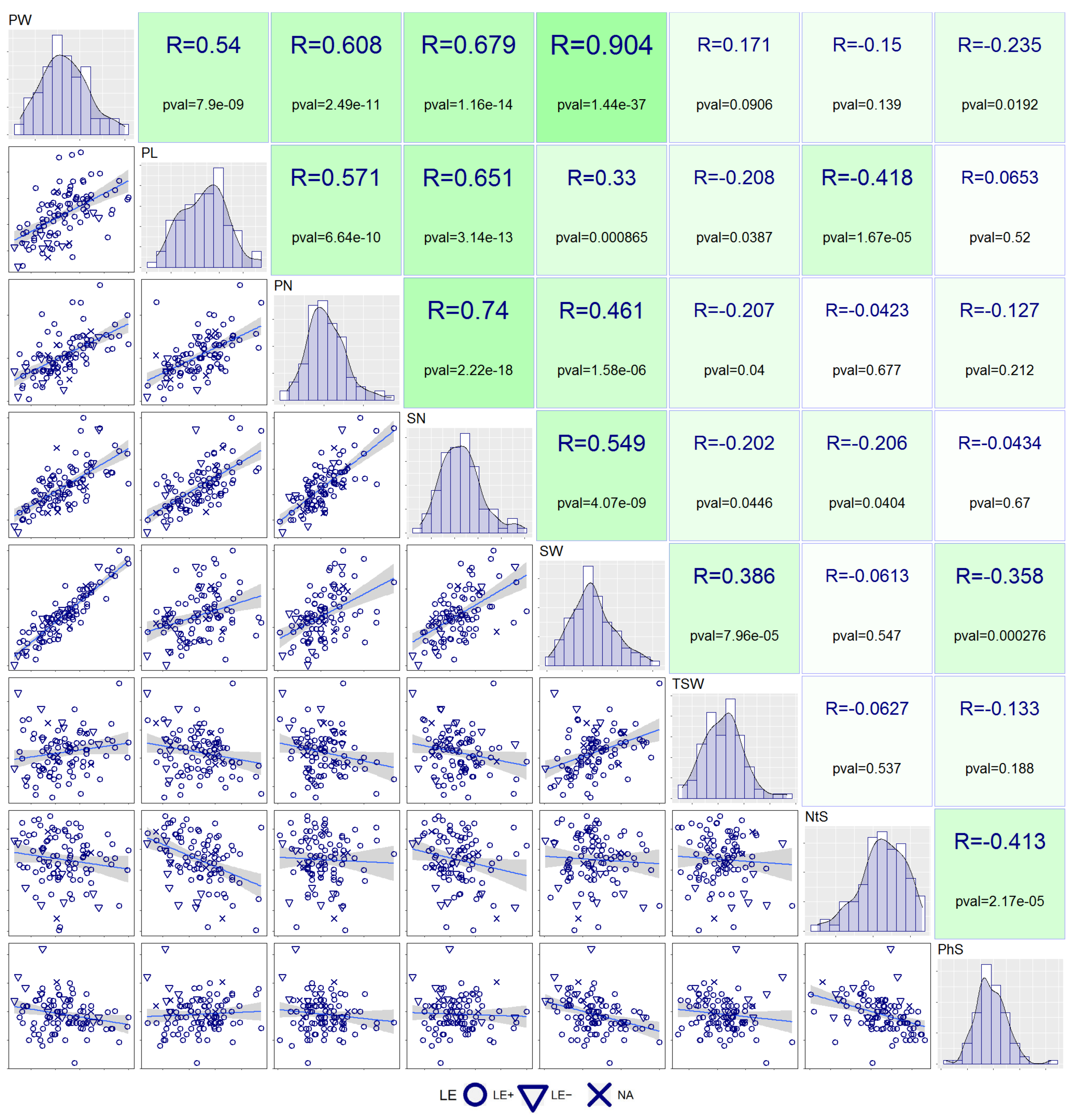
3.2. Correlation Analysis
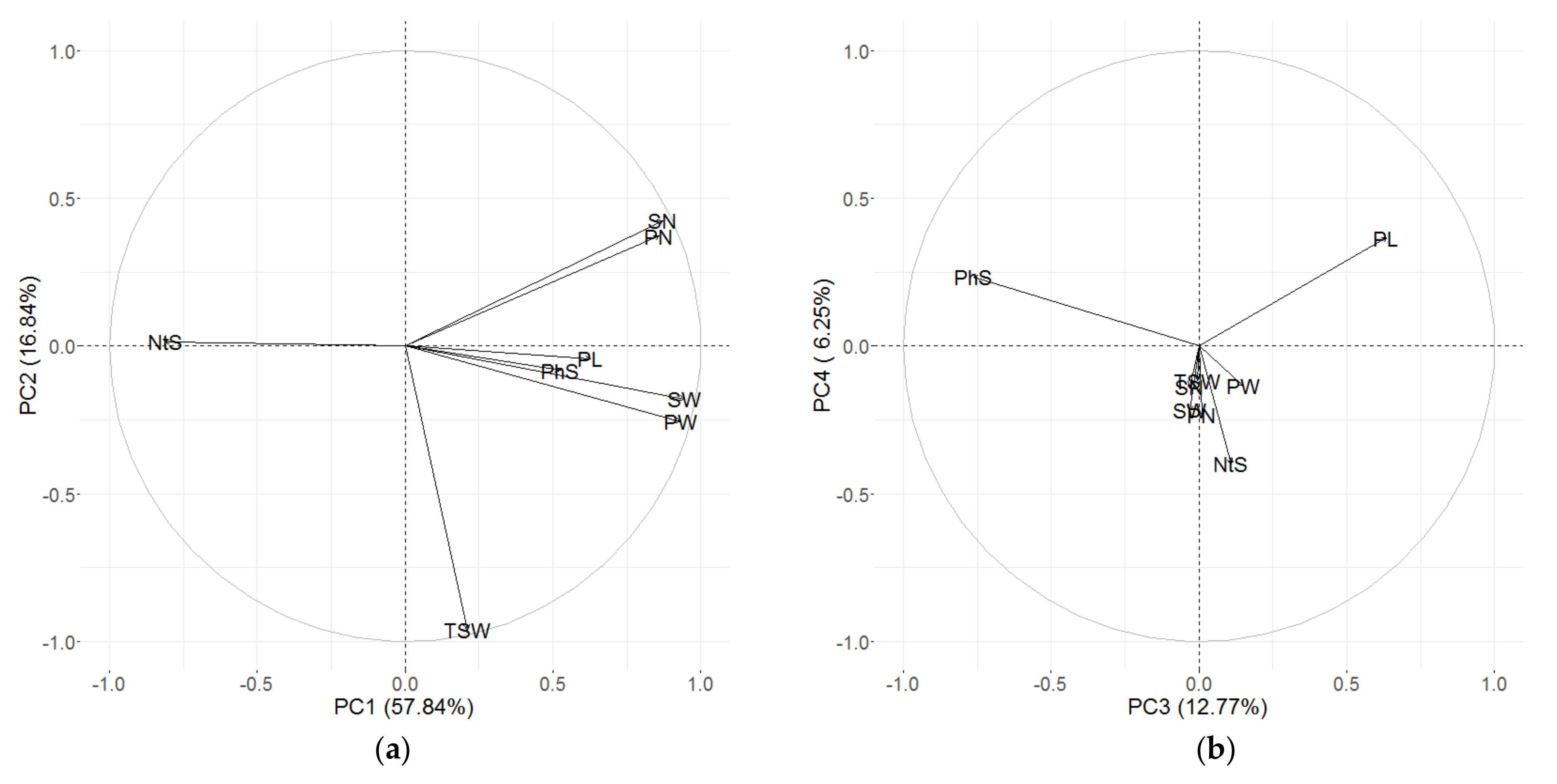
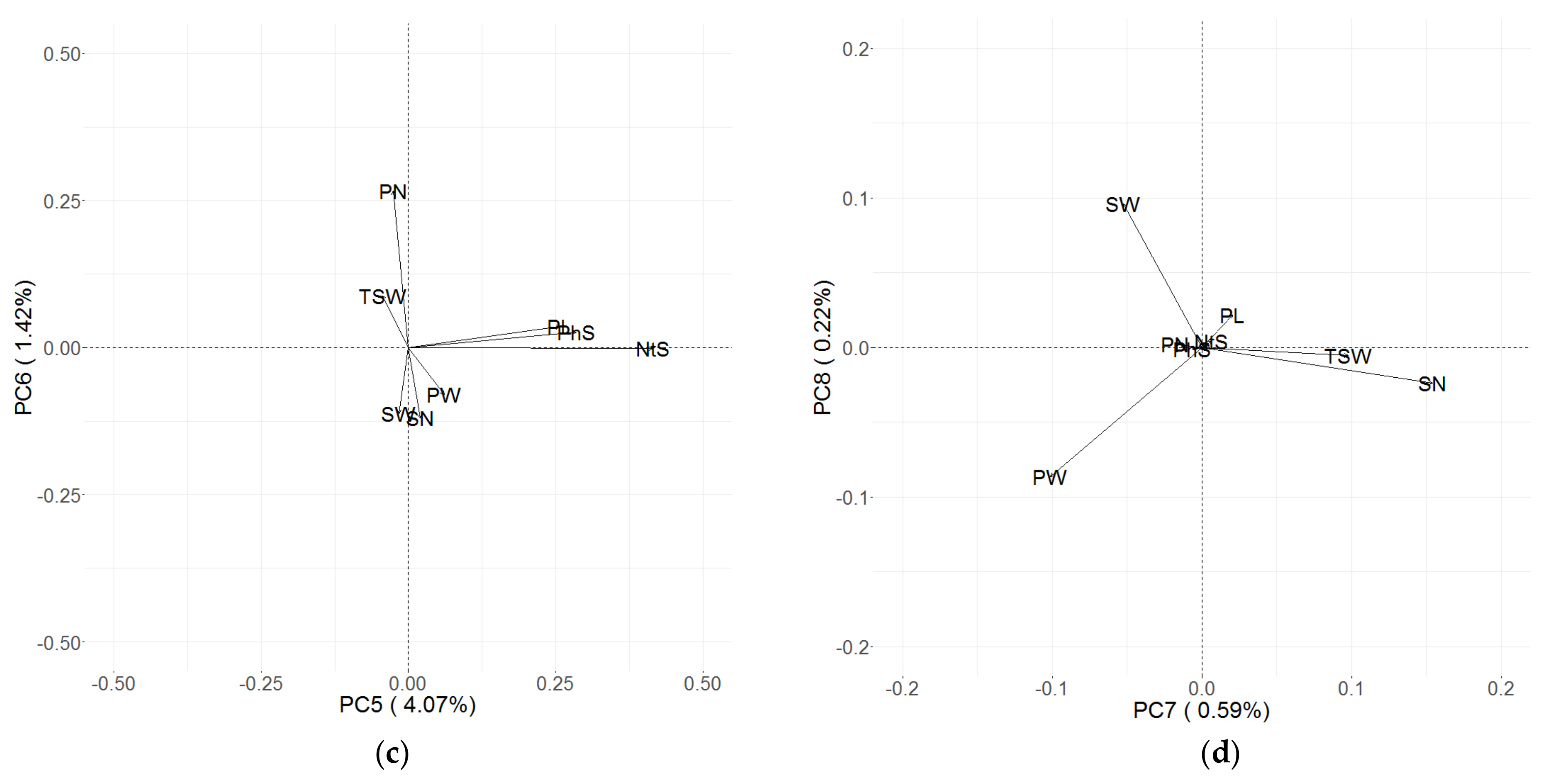
3.3. Principal Component (PC) Decomposition
3.4. Genotyping of Cultivars
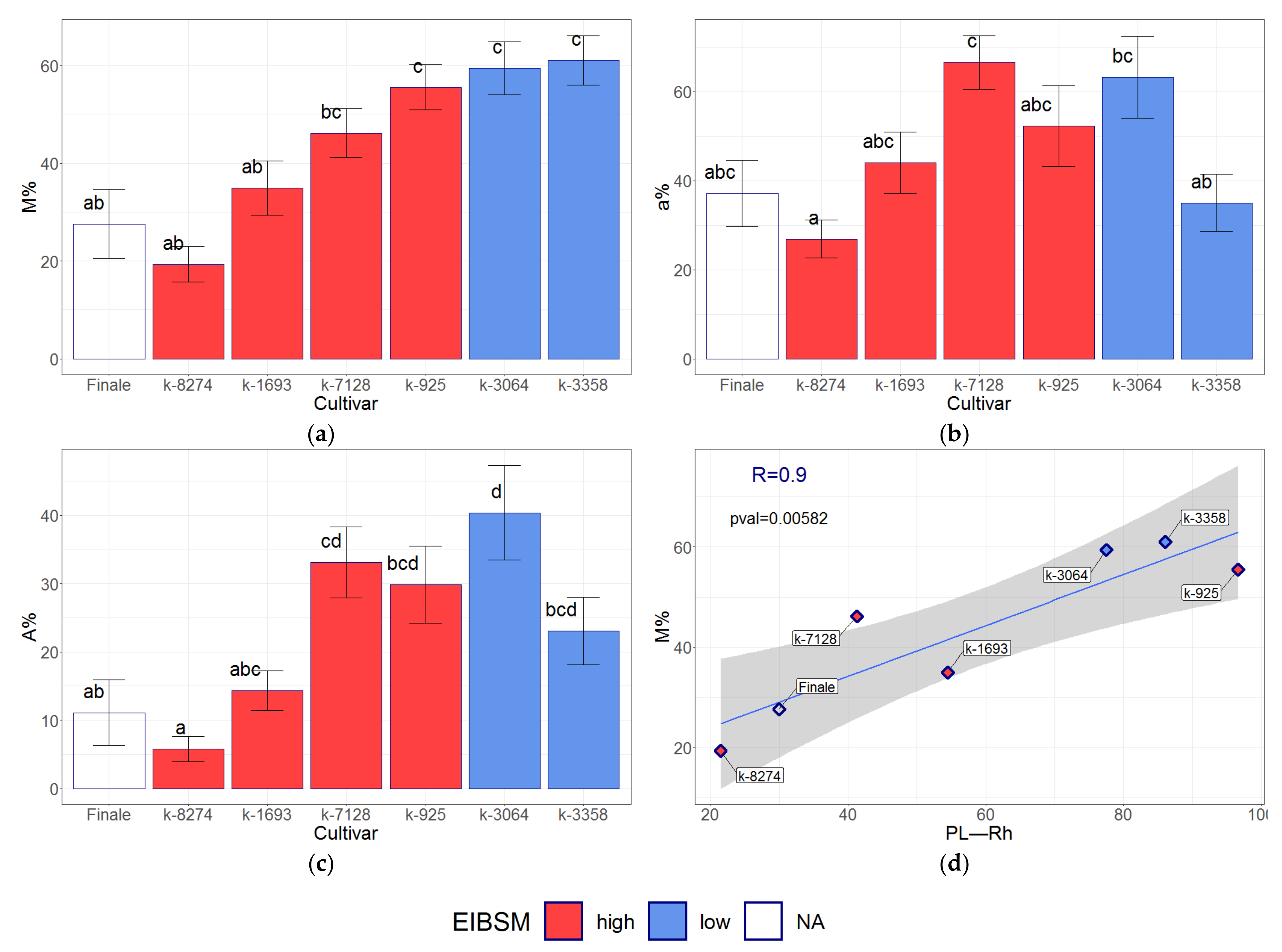
3.5. Association Analysis between the Allelic State of Sym-Genes and Traits
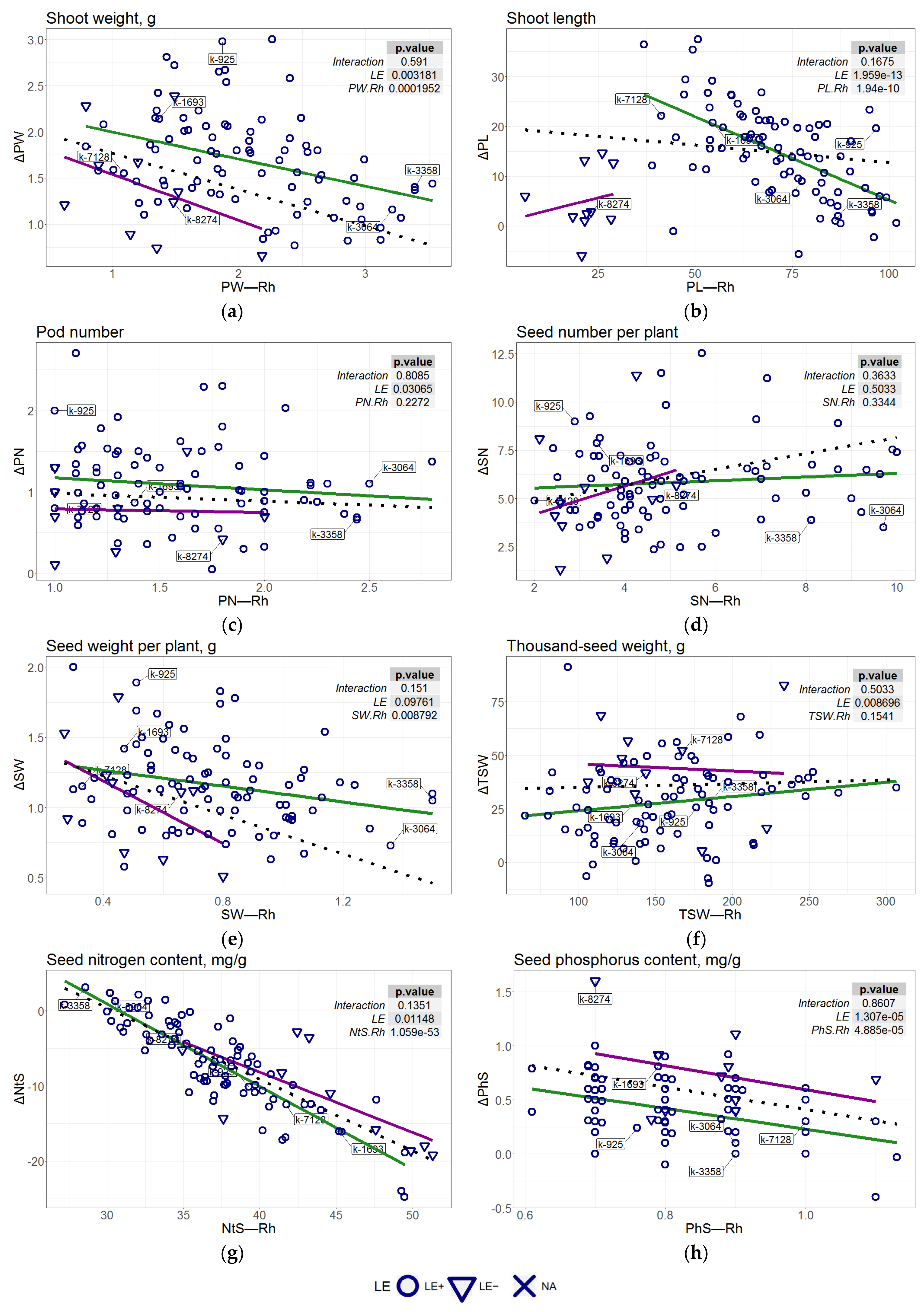
3.6. Regression Analysis of the Association of the AM-Caused Increments with SNVs
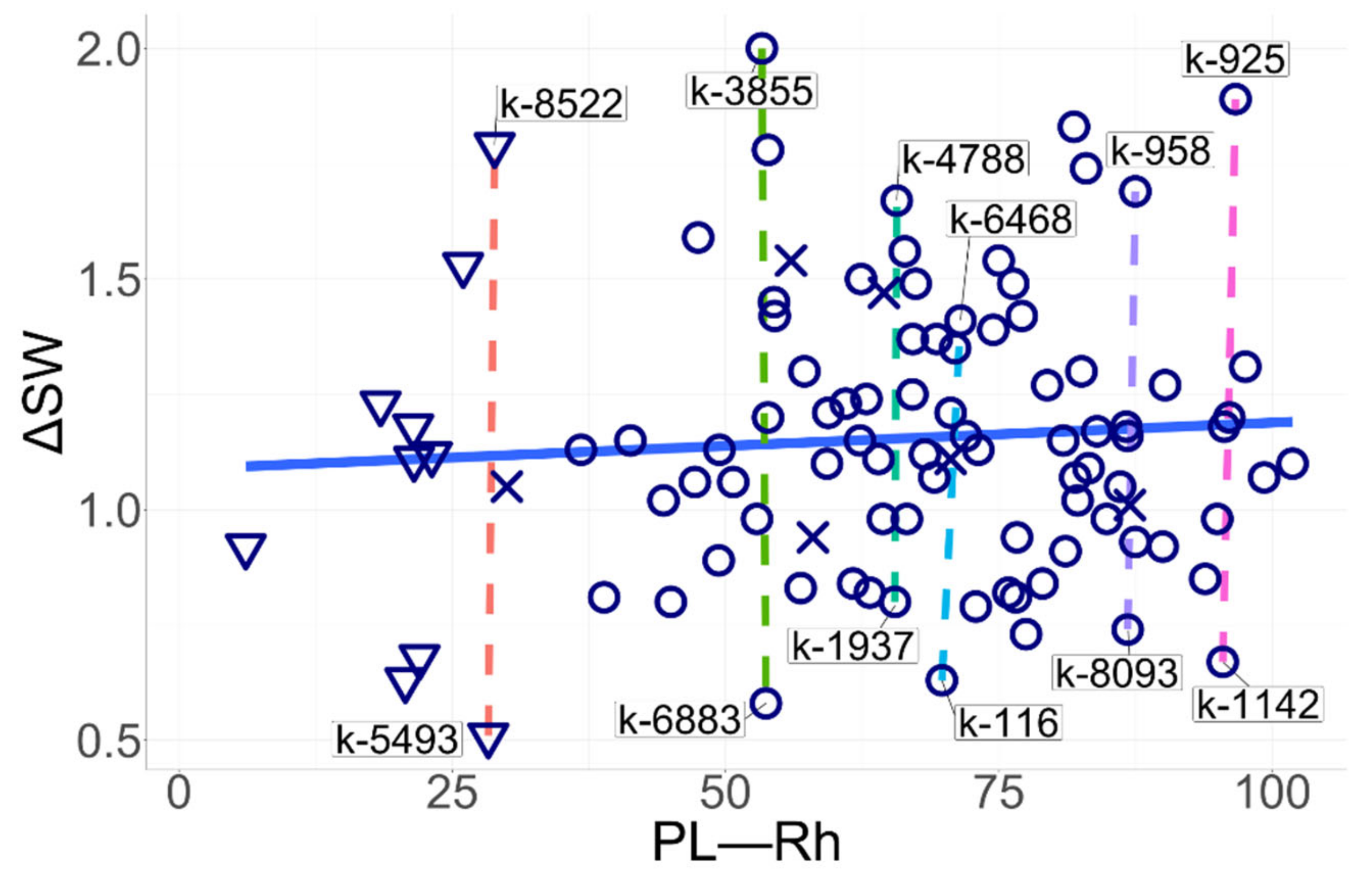
3.7. Association of the Root Mycorrhization Level and the Stem Length in Pea Cultivars
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lüscher, A.; Mueller-Harvey, I.; Soussana, J.-F.; Rees, R.M.; Peyraud, J.-L. Potential of legume-based grassland–livestock systems in Europe: A review. Grass Forage Sci. 2014, 69, 206–228. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D.; Dixon, R. Biotechnological solutions to the nitrogen problem. Curr. Opin. Biotechnol. 2014, 26, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Zhukov, V.A.; Shtark, O.Y.; Borisov, A.Y.; Tikhonovich, I.A. Breeding to Improve Symbiotic Effectiveness of Legumes. In Plant Breeding from Laboratories to Fields; InTech: Rijeka, Croatia, 2013; pp. 167–207. [Google Scholar]
- Shtark, O.Y.; Borisov, A.Y.; Zhukov, V.A.; Tikhonovich, I.A. Mutually beneficial legume symbioses with soil microbes and their potential for plant production. Symbiosis 2012, 58, 51–62. [Google Scholar] [CrossRef]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 years of genetic discoveries in legume nodulation and symbiotic nitrogen fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsyganov, V.E.; Tsyganova, A. V Symbiotic Regulatory Genes Controlling Nodule Development in Pisum sativum L. Plants 2020, 9, 1741. [Google Scholar] [CrossRef]
- Gorton, A.J.; Heath, K.D.; Pilet-Nayel, M.-L.; Baranger, A.; Stinchcombe, J.R. Mapping the genetic basis of symbiotic variation in legume-rhizobium interactions in Medicago truncatula. G3 Genes Genomes Genet. 2012, 2, 1291–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramaekers, L.; Galeano, C.H.; Garzón, N.; Vanderleyden, J.; Blair, M.W. Identifying quantitative trait loci for symbiotic nitrogen fixation capacity and related traits in common bean. Mol. Breed. 2013, 31, 163–180. [Google Scholar] [CrossRef]
- Tominaga, A.; Gondo, T.; Akashi, R.; Zheng, S.; Arima, S.; Suzuki, A. Quantitative trait locus analysis of symbiotic nitrogen fixation activity in the model legume Lotus japonicus. J. Plant Res. 2012, 125, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Smýkal, P.; Aubert, G.; Burstin, J.; Coyne, C.J.; Ellis, N.T.H.; Flavell, A.J.; Ford, R.; Hýbl, M.; Macas, J.; Neumann, P.; et al. Pea (Pisum sativum L.) in the Genomic Era. Agronomy 2012, 2, 74–115. [Google Scholar] [CrossRef]
- Bourgeois, M.; Jacquin, F.; Savois, V.; Sommerer, N.; Labas, V.; Henry, C.; Burstin, J. Dissecting the proteome of pea mature seeds reveals the phenotypic plasticity of seed protein composition. Proteomics 2009, 9, 254–271. [Google Scholar] [CrossRef]
- Deulvot, C.; Charrel, H.; Marty, A.; Jacquin, F.; Donnadieu, C.; Lejeune-Hénaut, I.; Burstin, J.; Aubert, G. Highly-multiplexed SNP genotyping for genetic mapping and germplasm diversity studies in pea. BMC Genom. 2010, 11, 468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourion, V.; Rizvi, S.M.H.; Fournier, S.; de Larambergue, H.; Galmiche, F.; Marget, P.; Duc, G.; Burstin, J. Genetic dissection of nitrogen nutrition in pea through a QTL approach of root, nodule, and shoot variability. Theor. Appl. Genet. 2010, 121, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Herridge, D.; Rose, I. Breeding for enhanced nitrogen fixation in crop legumes. Field Crop. Res. 2000, 65, 229–248. [Google Scholar] [CrossRef]
- Rengel, Z. Breeding for better symbiosis. In Food Security in Nutrient-Stressed Environments: Exploiting Plants’ Genetic Capabilities; Adu-Gyamfi, J.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 245–260. [Google Scholar] [CrossRef]
- Aliyu, O.M.; Makinde, B.O. Phenotypic analysis of seed yield and yield components in cowpea (Vigna unguiculata L., Walp). Plant Breed. Biotechnol. 2016, 4, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Chalk, P.M.; Souza, R.d.F.; Urquiaga, S.; Alves, B.J.R.; Boddey, R.M. The role of arbuscular mycorrhiza in legume symbiotic performance. Soil Biol. Biochem. 2006, 38, 2944–2951. [Google Scholar] [CrossRef]
- Shtark, O.Y.; Danilova, T.N.; Naumkina, T.S.; Vasilchikov, A.G.; Chebotar, V.K.; Kazakov, A.E.; Zhernakov, A.I.; Nemankin, T.A.; Prilepskaya, N.A.; Borisov, A.Y. Analysis of pea (Pisum sativum L.) source material for breeding of cultivars with high symbiotic potential and choice of criteria for its evaluation. Ecol. Genet. 2006, 4, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Mamontova, T.; Afonin, A.M.; Ihling, C.; Soboleva, A.; Lukasheva, E.; Sulima, A.S.; Shtark, O.Y.; Akhtemova, G.A.; Povydysh, M.N.; Sinz, A. Profiling of seed proteome in pea (Pisum sativum L.) lines characterized with high and low responsivity to combined inoculation with nodule bacteria and arbuscular mycorrhizal fungi. Molecules 2019, 24, 1603. [Google Scholar] [CrossRef] [Green Version]
- Afonin, A.M.; Gribchenko, E.S.; Zorin, E.A.; Sulima, A.S.; Romanyuk, D.A.; Zhernakov, A.I.; Shtark, O.Y.; Akhtemova, G.A.; Zhukov, V.A. Unique transcriptome features of pea (Pisum sativum L.) lines with differing responses to beneficial soil microorganisms. Ecol. Genet. 2021, 19, 131–141. [Google Scholar] [CrossRef]
- Yakobi, L.M.; Kukalev, A.S.; Ushakov, K.V.; Tsyganov, V.E.; Naumkina, T.S.; Provorov, N.A.; Borisov, A.Y.; Tikhonovich, I.A. Polymorphism of garden pea forms by the effectiveness of symbiosis with the Endomycorrhizal fungus Glomus sp. under conditions of inoculation with rhizobia. Agric. Biol. Sel’skokhozyaistvennaya Biol. 2000, 2000, 94–102. [Google Scholar]
- Engvild, K.C. Nodulation and nitrogen fixation mutants of pea, Pisum sativum. Theor. Appl. Genet. 1987, 74, 711–713. [Google Scholar] [CrossRef]
- Muromtsev, G.S.; Marshunova, G.A.; Jacobi, L.M. USSR Inventor’s Certificate no. 1501509. Moscow USSR State Regist. Invent. 1989. [Google Scholar]
- Afonin, A.; Sulima, A.; Zhernakov, A.; Zhukov, V. Draft genome of the strain RCAM1026 Rhizobium leguminosarum bv. viciae. Genomics Data 2017, 11, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Konieczny, A.; Ausubel, F.M. A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR-based markers. Plant J. 1993, 4, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.N.; Proebsting, W.M.; Hedden, P. Mendel’s dwarfing gene: cDNAs from the Le alleles and function of the expressed proteins. Proc. Natl. Acad. Sci. USA 1997, 94, 8907–8911. [Google Scholar] [CrossRef] [Green Version]
- Sulima, A.S.; Zhukov, V.A.; Afonin, A.A.; Zhernakov, A.I.; Tikhonovich, I.A.; Lutova, L.A. Selection signatures in the first exon of paralogous receptor kinase genes from the sym2 region of the Pisum sativum L. Genome. Front. Plant Sci. 2017, 8, 1957. [Google Scholar] [CrossRef] [Green Version]
- Zhukov, V.A.; Rychagova, T.S.; Fedorina, J.V.; Pinaev, A.G.; Andronov, E.E.; Borisov, A.Y.; Tikhonovich, I.A. Features of expression of the PsSst1 and PsIgn1 genes in nodules of pea (Pisum sativum L.) symbiotic mutants. Russ. J. Genet. 2016, 52, 362–369. [Google Scholar] [CrossRef]
- Kulaeva, O.A.; Zhernakov, A.I.; Afonin, A.M.; Boikov, S.S.; Sulima, A.S.; Tikhonovich, I.A.; Zhukov, V.A. Pea Marker Database (PMD)–A new online database combining known pea (Pisum sativum L.) gene-based markers. PLoS ONE 2017, 12, e0186713. [Google Scholar] [CrossRef] [Green Version]
- Afonin, A.M.; Leppyanen, I.V.; Kulaeva, O.A.; Shtark, O.Y.; Tikhonovich, I.A.; Dolgikh, E.A.; Zhukov, V.A. A high coverage reference transcriptome assembly of pea (Pisum sativum L.) mycorrhizal roots. Vavilov J. Genet. Breed. 2020, 24, 331–339. [Google Scholar] [CrossRef]
- Zhukov, V.A.; Zhernakov, A.I.; Kulaeva, O.A.; Ershov, N.I.; Borisov, A.Y.; Tikhonovich, I.A. De Novo Assembly of the Pea (Pisum sativum L.) Nodule Transcriptome. Int. J. Genom. 2015, 2015, 695947. [Google Scholar] [CrossRef] [Green Version]
- Alves-Carvalho, S.; Aubert, G.; Carrère, S.; Cruaud, C.; Brochot, A.L.; Jacquin, F.; Klein, A.; Martin, C.; Boucherot, K.; Kreplak, J.; et al. Full-length de novo assembly of RNA-seq data in pea (Pisum sativum L.) provides a gene expression atlas and gives insights into root nodulation in this species. Plant J. 2015, 84, 1–19. [Google Scholar] [CrossRef]
- Shtark, O.Y.; Sulima, A.S.; Zhernakov, A.I.; Kliukova, M.S.; Fedorina, J.V.; Pinaev, A.G.; Kryukov, A.A.; Akhtemova, G.A.; Tikhonovich, I.A.; Zhukov, V.A. Arbuscular mycorrhiza development in pea (Pisum sativum L.) mutants impaired in five early nodulation genes including putative orthologs of NSP1 and NSP2. Symbiosis 2016, 68, 129–144. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; Arnon, D.I., Ed.; California Agricultural Experiment Station, The College of Agriculture, University of California: Berkeley, CA, USA, 1950. [Google Scholar]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piché, Y. Ink and Vinegar, a Simple Staining Technique for Arbuscular-Mycorrhizal Fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Estimation of vesicular arbuscular mycorrhizal infection levels. Research for methods having a functional significance. In Proceedings of the Physiological and Genetical Aspects of Mycorrhizae=Aspects Physiologiques et Genetiques des Mycorhizes: Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, 1–5 July 1985; Institut National de le Recherche Agronomique: Paris, France, 1986. [Google Scholar]
- Shtark, O.Y.; Puzanskiy, R.K.; Avdeeva, G.S.; Yurkov, A.P.; Smolikova, G.N.; Yemelyanov, V.V.; Kliukova, M.S.; Shavarda, A.L.; Kirpichnikova, A.A.; Zhernakov, A.I.; et al. Metabolic alterations in pea leaves during arbuscular mycorrhiza development. PeerJ 2019, 2019, e7495. [Google Scholar] [CrossRef] [Green Version]
- Shtark, O.; Puzanskiy, R.; Avdeeva, G.; Yemelyanov, V.; Shavarda, A.; Romanyuk, D.; Kliukova, M.; Kirpichnikova, A.; Tikhonovich, I.; Zhukov, V. Metabolic Alterations in Pisum sativum Roots during Plant Growth and Arbuscular Mycorrhiza Development. Plants 2021, 10, 1033. [Google Scholar] [CrossRef]
- Kaeppler, S.M.; Parke, J.L.; Mueller, S.M.; Senior, L.; Stuber, C.; Tracy, W.F. Variation among maize inbred lines and detection of quantitative trait loci for growth at low phosphorus and responsiveness to arbuscular mycorrhizal fungi. Crop Sci. 2000, 40, 358–364. [Google Scholar] [CrossRef]
- Davis, E.O.; Evans, I.J.; Johnston, A.W.B. Identification of nodX, a gene that allows Rhizobium leguminosarum biovar viciae strain TOM to nodulate Afghanistan peas. Mol. Gen. Genet. MGG 1988, 212, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Sulima, A.S.; Zhukov, V.A.; Kulaeva, O.A.; Vasileva, E.N.; Borisov, A.Y.; Tikhonovich, I.A. New sources of Sym2A allele in the pea (Pisum sativum L.) carry the unique variant of candidate LysM-RLK gene LykX. PeerJ 2019, 7, e8070. [Google Scholar] [CrossRef] [Green Version]
- Solovev, Y.V.; Igolkina, A.A.; Kuliaev, P.O.; Sulima, A.S.; Zhukov, V.A.; Porozov, Y.B.; Pidko, E.A.; Andronov, E.E. Towards understanding Afghanistan pea symbiotic phenotype through the molecular modeling of the interaction between LykX-Sym10 receptor heterodimer and Nod factors. Front. Plant Sci. 2021, 12, 824. [Google Scholar] [CrossRef]
- Zhukov, V.; Radutoiu, S.; Madsen, L.H.; Rychagova, T.; Ovchinnikova, E.; Borisov, A.; Tikhonovich, I.; Stougaard, J. The Pea Sym37 Receptor Kinase Gene Controls Infection-Thread Initiation and Nodule Development. Mol. Plant-Microbe Interact. 2008, 21, 1600–1608. [Google Scholar] [CrossRef] [Green Version]
- Kirienko, A.N.; Porozov, Y.B.; Malkov, N.V.; Akhtemova, G.A.; Le Signor, C.; Thompson, R.; Saffray, C.; Dalmais, M.; Bendahmane, A.; Tikhonovich, I.A. Role of a receptor-like kinase K1 in pea Rhizobium symbiosis development. Planta 2018, 248, 1101–1120. [Google Scholar] [CrossRef]
- Kirienko, A.N.; Vishnevskaya, N.A.; Kitaeva, A.B.; Shtark, O.Y.; Kozyulina, P.Y.; Thompson, R.; Dalmais, M.; Bendahmane, A.; Tikhonovich, I.A.; Dolgikh, E.A. Structural variations in LysM domains of LysM-RLK psK1 may result in a different effect on Pea–Rhizobial symbiosis development. Int. J. Mol. Sci. 2019, 20, 1624. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Knox, M.R.; Edwards, A.; Hogg, B.; Ellis, T.H.N.; Wei, G.; Downie, J.A. Natural variation in host-specific nodulation of pea is associated with a haplotype of the SYM37 LysM-type receptor-like kinase. Mol. Plant-Microbe Interact. 2011, 24, 1396–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansa, J.; Smith, F.A.; Smith, S.E. Are there benefits of simultaneous root colonization by different arbuscular mycorrhizal fungi? New Phytol. 2008, 177, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Maillet, F.; Poinsot, V.; André, O.; Puech-Pagès, V.; Haouy, A.; Gueunier, M.; Cromer, L.; Giraudet, D.; Formey, D.; Niebel, A.; et al. Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature 2011, 469, 58–63. [Google Scholar] [CrossRef]
- Streng, A.; op den Camp, R.; Bisseling, T.; Geurts, R. Evolutionary origin of rhizobium Nod factor signaling. Plant Signal. Behav. 2011, 6, 1510–1514. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Chen, R. Nitrogen and Phosphorus Signaling and Transport during Legume–Rhizobium Symbiosis. Front. Plant Sci. 2021, 12, 683601. [Google Scholar] [CrossRef] [PubMed]
- Irzykowska, L.; Wolko, B. Interval mapping of QTLs controlling yield-related traits and seed protein content in Pisum sativum. J. Appl. Genet. 2004, 45, 297–306. [Google Scholar] [PubMed]
- Sachs, J.L.; Quides, K.W.; Wendlandt, C.E. Legumes versus rhizobia: A model for ongoing conflict in symbiosis. New Phytol. 2018, 219, 1199–1206. [Google Scholar] [CrossRef] [Green Version]
- Bourion, V.; Laguerre, G.; Depret, G.; Voisin, A.-S.; Salon, C.; Duc, G. Genetic Variability in Nodulation and Root Growth Affects Nitrogen Fixation and Accumulation in Pea. Ann. Bot. 2007, 100, 589–598. [Google Scholar] [CrossRef]
- Novák, K.; Biedermannová, E.; Vondrys, J. Symbiotic and Growth Performance of Supernodulating Forage Pea Lines. Crop Sci. 2009, 49, 1227–1234. [Google Scholar] [CrossRef]
- Wang, C.; Reid, J.B.; Foo, E. The Art of Self-Control–Autoregulation of Plant–Microbe Symbioses. Front. Plant Sci. 2018, 9, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlo, M.; Boschiero, C.; Landerslev, K.G.; Blanco, G.S.; Wen, J.; Mysore, K.S.; Dai, X.; Zhao, P.X.; de Bang, T.C. The CLE53–SUNN genetic pathway negatively regulates arbuscular mycorrhiza root colonization in Medicago truncatula. J. Exp. Bot. 2020, 71, 4972–4984. [Google Scholar] [CrossRef]
- Mortier, V.; Den Herder, G.; Whitford, R.; Van de Velde, W.; Rombauts, S.; D’haeseleer, K.; Holsters, M.; Goormachtig, S. CLE peptides control Medicago truncatula nodulation locally and systemically. Plant Physiol. 2010, 153, 222–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, L.M.; Flokova, K.; Schnabel, E.; Sun, X.; Fei, Z.; Frugoli, J.; Bouwmeester, H.J.; Harrison, M.J. A CLE–SUNN module regulates strigolactone content and fungal colonization in arbuscular mycorrhiza. Nat. Plants 2019, 5, 933–939. [Google Scholar] [CrossRef]
- Zhukov, V.A.; Akhtemova, G.A.; Zhernakov, A.I.; Sulima, A.S.; Shtark, O.Y.; Tikhonovich, I.A. Evaluation of the symbiotic effectiveness of Pea (Pisum Sativum L.) Genotypes in pot experiment. Sel’skokhozyaistvennaya Biol. 2017, 52, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Nishida, Y.; Hiraoka, R.; Kawano, S.; Suganuma, N.; Sato, S.; Watanabe, S.; Anai, T.; Arima, S.; Tominaga, A.; Suzuki, A. SEN1 gene from Lotus japonicus MG20 improves nitrogen fixation and plant growth. Soil Sci. Plant Nutr. 2020, 66, 864–869. [Google Scholar] [CrossRef]








| Measured Trait | Mean Value, Rh | Mean Value, Rh + AM | Mean Increase (abs) 1 | Mean Increase (rel), % |
|---|---|---|---|---|
| Shoot weight, g | 1.96, SD = 0.668 | 3.62, SD = 0.725 | 1.67 ± 0.1 | 98.90% |
| Shoot length, cm | 65.5, SD = 21.3 | 79.3, SD = 22.3 | 13.8 ± 1.9 | 24.00% |
| Pod number | 1.56, SD = 0.43 | 2.61, SD = 0.622 | 1.05 ± 0.097 | 73.10% |
| Seed number per plant | 4.87, SD = 1.97 | 10.6, SD = 3.13 | 5.75 ± 0.43 | 133.40% |
| Seed weight per plant, g | 0.74, SD = 0.267 | 1.89, SD = 0.364 | 1.15 ± 0.061 | 183.30% |
| Thousand-seed weight, g | 160, SD = 49.0 | 190, SD = 55.0 | 30.2 ± 3.7 | 20.30% |
| Seed nitrogen content, mg/g | 38, SD = 5.25 | 30.5, SD = 3.17 | −7.54 ± 1.2 | −18.40% |
| Seed phosphorus content, mg/g | 0.816, SD = 0.116 | 1.27, SD = 0.283 | 0.453 ± 0.06 | 58.30% |
| Measured Trait | Paired t-Test 1 t-Statistic, p-Value | F-Test 2 F-Statistic, p-Value | Pearson Correlation Coefficient (R) | Coefficient of Determination, Adjusted (R2adj) |
|---|---|---|---|---|
| PW | 31.64, p-value = 3.07 × 10−53 | 1.18, p-value = 0.425 | 0.720, p-value = 4.55 × 10−17 | 0.513 |
| PL | 14.71, p-value = 1.53 × 10−26 | 1.09, p-value = 0.659 | 0.910, p-value = 8.37 × 10−39 | 0.826 |
| PN | 21.49, p-value = 7.21 × 10−39 | 2.09, p-value = 3.29 × 10−4 | 0.625, p-value = 4.93 × 10−12 | 0.384 |
| SN | 26.47, p-value = 1.95 × 10−46 | 2.53, p-value = 6.53 × 10−6 | 0.731, p-value = 8.73 × 10−18 | 0.53 |
| SW | 37.32, p-value = 9.76 × 10−60 | 1.86, p-value = 2.45 × 10−3 | 0.560, p-value = 1.62 × 10−9 | 0.307 |
| TSW | 16.03, p-value = 3.89 × 10−29 | 1.26, p--value = 0.253 | 0.941, p-value = 1.40 × 10−47 | 0.885 |
| NtS | −12.34, p-value = 1.17 × 10−21 | 0.37, p-value = 1.13 × 10−6 | 0.019, p-value = 0.85 | −0.01 |
| PhS | 15.05, p-value = 3.23 × 10−27 | 5.96, p-value < 2.2 × 10−16 | 0.052, p-value = 0.611 | −0.008 |
| Gene Identifier | Protein Substitutions |
|---|---|
| IGN1 | D108E |
| SEN1 | P22A |
| SST1 | S545Y |
| Sym29 | F16S |
| P43H | |
| A53G | |
| I171F | |
| S618P | |
| T623I | |
| STR1 | E109K |
| P397S | |
| A401P | |
| H475Y | |
| STR2 | T676I |
| AMT2;3 | A21P * |
| N215I | |
| Y385N | |
| AMT2;5 | — |
| SNV | Trait | p-Value | FDR | SNV | Trait | p-Value | FDR |
|---|---|---|---|---|---|---|---|
| Le.A229T | PL | 4.00 × 10−38 | 3.20 × 10−37 | SEN1.P22A | PL | 1.24 × 10−3 | 9.92 × 10−3 |
| Le.A229T | PW | 9.54 × 10−9 | 3.81 × 10−8 | LykX.L42K | TSW | 1.53 × 10−3 | 8.65 × 10−3 |
| Le.A229T | NtS | 7.82 × 10−7 | 2.08 × 10−6 | K1.E90D | TSW | 1.86 × 10−3 | 0.0149 |
| LykX.L13V | SN | 4.43 × 10−6 | 3.55 × 10−5 | LykX.L42K | SN | 2.16 × 10−3 | 8.60 × 10−3 |
| Le.A229T | PhS | 8.36 × 10−6 | 1.67 × 10−5 | Sym37.L56V | SN | 2.21 × 10−3 | 8.80 × 10−3 |
| LykX.L9F | TSW | 1.20 × 10−5 | 9.57 × 10−5 | IGN1.D108E | PN | 2.36 × 10−3 | 9.20 × 10−3 |
| LykX.A142I | TSW | 2.35 × 10−5 | 1.88 × 10−4 | IGN1.D108E | PL | 3.47 × 10−3 | 9.20 × 10−3 |
| Le.A229T | SW | 3.37 × 10−5 | 5.39 × 10−5 | LykX.A142I | SN | 3.58 × 10−3 | 0.0143 |
| LykX.L9F | SN | 4.96 × 10−5 | 1.98 × 10−4 | LykX.S134G | SW | 5.82 × 10−3 | 0.0466 |
| LykX.L13V | PN | 1.30 × 10−4 | 5.19 × 10−4 | AMT2;3.N215I | SW | 6.46 × 10−3 | 0.0204 |
| LykX.V86I | SN | 1.97 × 10−4 | 1.02 × 10−3 | AMT2;3.N215I | PL | 7.63 × 10−3 | 0.0204 |
| LykX.F184S | SN | 1.97 × 10−4 | 1.02 × 10−3 | IGN1.D108E | TSW | 7.68 × 10−3 | 0.0154 |
| IGN1.D108E | SN | 2.09 × 10−4 | 1.67 × 10−3 | Sym37.L56V | PN | 8.71 × 10−3 | 0.0232 |
| Sym37.S45L | TSW | 2.30 × 10−4 | 1.84 × 10−3 | K1.E90D | SN | 0.0103 | 0.0411 |
| LykX.V86I | TSW | 2.54 × 10−4 | 1.02 × 10−3 | AMT2;3.N215I | TSW | 0.0109 | 0.0218 |
| LykX.F184S | TSW | 2.54 × 10−4 | 1.02 × 10−3 | LykX.L9F | PN | 0.0119 | 0.0316 |
| Le.A229T | SN | 4.29 × 10−4 | 5.72 × 10−3 | LykX.L13V | PL | 0.0136 | 0.0273 |
| Sym37.L56V | TSW | 5.40 × 10−4 | 4.32 × 10−3 | Sym37.S45L | PN | 0.0168 | 0.0480 |
| Le.A229T | PN | 6.08 × 10−4 | 6.95 × 10−4 | Sym37.S45L | SN | 0.0180 | 0.0480 |
| LykX.L13V | TSW | 7.74 × 10−4 | 2.06 × 10−3 | LykX.L13V | SW | 0.0296 | 0.0461 |
| AMT2;3.N215I | PW | 1.17 × 10−3 | 9.36 × 10−3 | LykX.L13V | NtS | 0.0346 | 0.0461 |
| SNV | PC | p-Value | FDR | SNV | PC | p-Value | FDR |
|---|---|---|---|---|---|---|---|
| Le.A229T | PC3 | 1.23 × 10−25 | 4.93 × 10−25 | LykX.L42K | PC2 | 2.40 × 10−3 | 9.61 × 10−3 |
| Le.A229T | PC1 | 2.04 × 10−13 | 4.08 × 10−13 | IGN1.D108E | PC2 | 3.66 × 10−3 | 0.0130 |
| Le.A229T | PC4 | 1.66 × 10−8 | 2.22 × 10−8 | K1.E90D | PC2 | 4.98 × 10−3 | 0.0199 |
| LykX.L9F | PC2 | 1.13 × 10−5 | 4.51 × 10−5 | AMT2;3.N215I | PC1 | 5.13 × 10−3 | 0.0205 |
| Sym37.S45L | PC2 | 9.71 × 10−5 | 3.88 × 10−4 | IGN1.D108E | PC1 | 6.51 × 10−3 | 0.0130 |
| LykX.A142I | PC2 | 2.02 × 10−4 | 8.06 × 10−4 | STR2.T676I | PC2 | 7.70 × 10−3 | 0.0308 |
| LykX.L13V | PC2 | 2.21 × 10−4 | 8.85 × 10−4 | LykX.S134G | PC1 | 0.0112 | 0.0450 |
| Sym37.L56V | PC2 | 2.48 × 10−4 | 9.90 × 10−4 | SEN1.P22A | PC4 | 0.0145 | 0.0484 |
| LykX.F184S | PC2 | 2.85 × 10−4 | 1.14 × 10−3 | K1.E90D | PC4 | 0.0219 | 0.0437 |
| LykX.V86I | PC2 | 2.85 × 10−4 | 1.14 × 10−3 | SEN1.P22A | PC3 | 0.0242 | 0.0484 |
| LykX.L13V | PC1 | 4.43 × 10−4 | 8.87 × 10−4 | LykX.L13V | PC3 | 0.0353 | 0.0471 |
| Trait | The Formula Used in the Model |
|---|---|
| PW | dPW~PW.Rh + LE |
| PL | dPL~PL.Rh + LE |
| PN | dPN~LE |
| SN | dSN~1 |
| SW | dSW~SW.Rh |
| TSW | dTSW~LE |
| NtS | dNtS~NtS.Rh + LE |
| PhS | dPhS~PhS.Rh + LE |
| SNV | Trait | p-Value | FDR |
|---|---|---|---|
| AMT2;3.215NI | dTSW | 0.0050 | 0.0402 |
| K1.31LI | dTSW | 0.0047 | 0.0379 |
| LykX.16VF | dTSW | 0.0038 | 0.0301 |
| LykX.184FS | dTSW | 0.0056 | 0.0447 |
| SNV | PC | p-Value | FDR |
|---|---|---|---|
| AMT2;3.215NI | dPC2 | 0.0061 | 0.0243 |
| LykX.16VF | dPC2 | 0.0093 | 0.0371 |
| SST1.545SY | dPC1 | 0.0215 | 0.0430 |
| SST1.545SY | dPC2 | 0.0164 | 0.0430 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhukov, V.A.; Zhernakov, A.I.; Sulima, A.S.; Kulaeva, O.A.; Kliukova, M.S.; Afonin, A.M.; Shtark, O.Y.; Tikhonovich, I.A. Association Study of Symbiotic Genes in Pea (Pisum sativum L.) Cultivars Grown in Symbiotic Conditions. Agronomy 2021, 11, 2368. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112368
Zhukov VA, Zhernakov AI, Sulima AS, Kulaeva OA, Kliukova MS, Afonin AM, Shtark OY, Tikhonovich IA. Association Study of Symbiotic Genes in Pea (Pisum sativum L.) Cultivars Grown in Symbiotic Conditions. Agronomy. 2021; 11(11):2368. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112368
Chicago/Turabian StyleZhukov, Vladimir A., Aleksandr I. Zhernakov, Anton S. Sulima, Olga A. Kulaeva, Marina S. Kliukova, Alexey M. Afonin, Oksana Y. Shtark, and Igor A. Tikhonovich. 2021. "Association Study of Symbiotic Genes in Pea (Pisum sativum L.) Cultivars Grown in Symbiotic Conditions" Agronomy 11, no. 11: 2368. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112368