3.1. General Characteristics of the Plant Remains in Welleholz GVAC_06
4412 remains were identified in sample GVAC_06 (
Table 4 and
Table 5) of which the majority is desiccated. The finds are subdivided into eight material categories: cereals, oil and fibre crops, weeds and wild plants, wood and charcoal, indeterminate plants, animals and insects, fungi, and other. The cereal category represents a clear majority of the finds, with an absolute quantity of 3737 (84.78%;
Table 4). The bulk of the quantified cereal remains consists of cereal straw and rye ears (
Table 5). Other finds of rye include grains, rachises and anthers (
Figure 1a,b). Cereals were further represented by grains, rachises and spikelets of common oat (
Avena sativa), six-row hulled barley (
Hordeum vulgare ssp.
vulgare, hulled) and bread wheat (
Triticum aestivum ssp.
aestivum). The group of oil and fibre crops included two finds belonging to two taxa: a fruit fragment of flax/linseed (
Linum usitatissimum,
Figure 1c) and a seed of opium poppy (
Papaver cf.
somniferum).
The category of weeds and wild plants is the second most abundant category, with 368 finds (8.35%) and includes the highest diversity of taxa (
n = 50) of all plant categories. Seeds of long-headed poppy/common poppy (
Papaver dubium/rhoeas,
Figure 1d) are represented by the highest absolute quantity and other frequent finds include red bartsia (
Odontites vernus,
Figure 1e) and white goosefoot (
Chenopodium album). Seeds and fruits form the majority of plant parts, but several taxa were also represented by flower heads, fruits, pods and leafs (
Figure 1f).
The category of indeterminate plants included a variety of remains, of which small fragments of charcoal were the most abundant. The group of animals and insects mainly consisted of diverse fragments of various insects, but also included bone fragments and caddisfly (Trichoptera) cases. The category of fungi consists of straw remains of different cereal taxa containing dark “lesions” that were likely caused by an infection with an unidentified fungus. Finally, the category of other materials included single finds of plaster, ceramics and yarn.
The rich diversity of plant taxa and plant parts in Wellerholz GVAC_06 attests to the excellent preservation conditions of the plant remains. The predominance of straw, ears and spikelets of rye indicate that it is likely that the plant material used to wrap the Wellerholz represents a harvest of rye. The high diversity of taxa of weeds and wild plants, the variety of plant parts (including flower heads, capsules and nearly complete plants) as well as the rarity of cereal grains, suggest that the plant material represents the by-products of the earliest stages of crop-processing of rye (i.e., between stages one and three as described by Stevens [
66]. Considering that the aim of crop processing is to remove impurities from the harvest, the farther one progresses through the crop-processing sequence, the less reliable the data will be with respect to reconstruction of weed ecology and cultivation practice (especially the reconstruction of sowing season; e.g., [
57]. In the case of GVAC_06, it can thus be assumed that the weeds and wild plants represent a significant part of the arable weed flora and that the data resulting from their analysis will not be severely biased.
3.2. Reconstructing Sowing Time and Cultivation Intensity
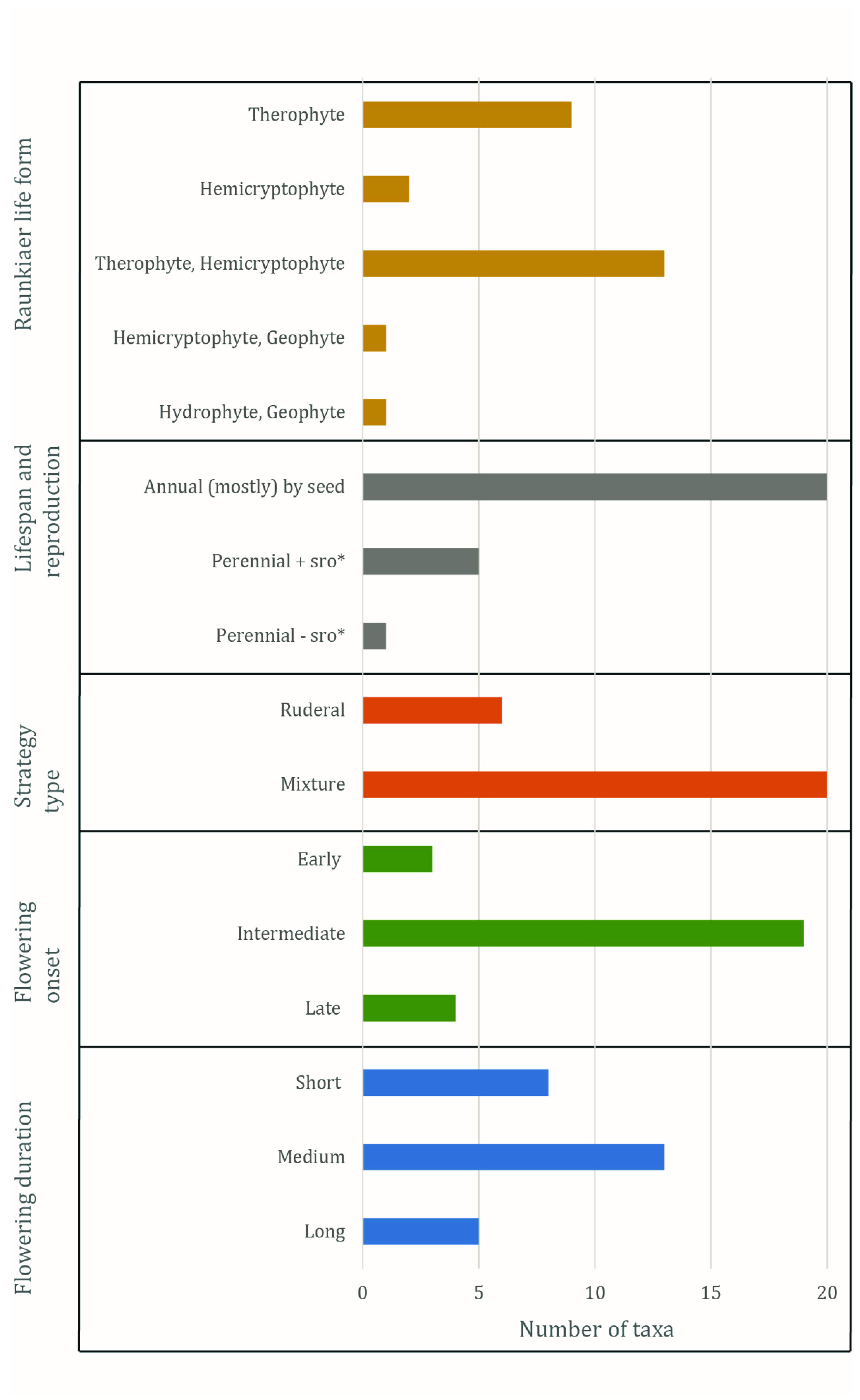
Figure 2 and
Table S1 summarise the characteristics collected on plant-sociological groups, classification, physical properties, autecology and (a part of the) functional attributes of 26 taxa of weeds and wild plants from GVAC_06. The vast majority of taxa of weeds and wild plants are part of Ellenberg’s classification [
43] of modern segetal communities (in this case, classes 3.3 and 3.4). Most of the taxa (
n = 12) are classified as Secalietea (class 3.4), a smaller portion as Chenopodietea (class 3.3;
n = 7), while a few individual taxa belong to various other groups. The bulk (
n = 21) is classified as indicator species of different levels of the plant-sociological groups. The dominant lifeform is therophyte-hemicryptophyte (
n = 13), followed by therophyte (
n = 9), hemicryptophyte (
n = 2), hemicryptophyte-geophyte (
n = 1) and hydrophyte-geophyte (
n = 1). Annual taxa are dominant (
n = 19), followed by taxa that are able to grow both as annuals and perennials (
n = 3) and, lastly, perennials (
n = 2). The reproduction type of the taxa is for the largest part by seed (
n = 21) and a few (
n = 5) are able to reproduce by seed as well as vegetatively through subterranean reproductive organs. Most taxa (
n = 21) have a strategy type that is a mixture between ruderals, competitors and stress-tolerators, and six taxa are classified as ruderals. The most common flowering onset is intermediate (
n = 19), with nearly equal amounts of taxa having a late (
n = 4) and early (
n = 3) flowering onset. Half of the taxa have a medium flowering duration (
n = 13), while the other half have a short (
n = 8) or a long (
n = 5) duration.
The sowing time of the rye in the sample can be reconstructed from data based on the flowering onset and duration of flowering of the arable weed taxa (
Table 1 and
Table 2). Autumn sowing is typically indicated by early and intermediate onset of flowering and a short flowering duration (e.g., [
55,
67]). In the case of the arable weeds in Wellerholz GVAC_06, the former indicators are predominant, while the latter indicator is the second most common; the flowering onset is thus clearly indicative of autumn sowing, while the flowering duration leaves some room for arguing. According to studies conducted by Bogaard et al. [
67], indicators of spring sowing (i.e., late flowering onset and long flowering duration) are able to grow in autumn sown fields, while indicators for autumn sowing are usually not able to grow in spring-sown fields due to their inability to regenerate after spring ploughing. When autumn-sowing indicators occur in combination with spring-sowing indicators, it is therefore most likely that the data represent autumn sown crops. In the case of the arable weed taxa in Wellerholz GVAC_06 it can therefore be assumed that the rye in the sample was grown as a winter cereal.
The intensity of cultivation in this study is reconstructed with two sets of data and approaches: (i) descriptive values of autecological data and functional attributes, and (ii) discriminant analysis based on functional attributes. The former includes considerations of the flowering onset, the flowering duration, the Raunkiaer lifeform, the lifespan, the reproduction type and the strategy type (
Table 1 and
Table S1). The data on flowering onset and flowering duration proved to be inconclusive due to the predominance of a medium flowering onset and an intermediate flowering duration (
Figure 2;
Table S1). The data on the lifespan and reproduction type are more conclusive: the predominance of an annual lifespan and reproduction by seeds are typical indicators of cultivation practices that include a high degree of disturbance [
2,
41,
58] (p. 145, pp. 344–346, p. 188). The most abundant Raunkiaer life form is represented by therophytes-hemicryptophytes, while therophytes form the second most abundant group. The latter is characteristic for highly disturbed habitats [
59] (p. 810) and there are indications that some hemicryptophytes are as well able to grow in disturbed habitats, albeit in lower proportions [
68] (p. 38). It is unknown to the authors whether the transitional lifeform of therophytes-hemicryptophytes thrives in disturbed habitats, and therefore therophytes, the second most abundant lifeform of the arable weeds in GVAC_06, are, on their own, seen as tentative indicators for a high degree of disturbance. The strategy type of the majority of the taxa was represented by a mixture between ruderals, competitors and stress-tolerators, and six taxa were classified as ruderals. Ruderals are typical indicators of high cultivation intensity, but, similar to the Raunkiaer lifeform, due to their relatively low representation in this dataset, they are seen as complementary to the data on lifespan and reproduction type. Altogether, the descriptive values can thus be interpreted as indicative of a high cultivation intensity.
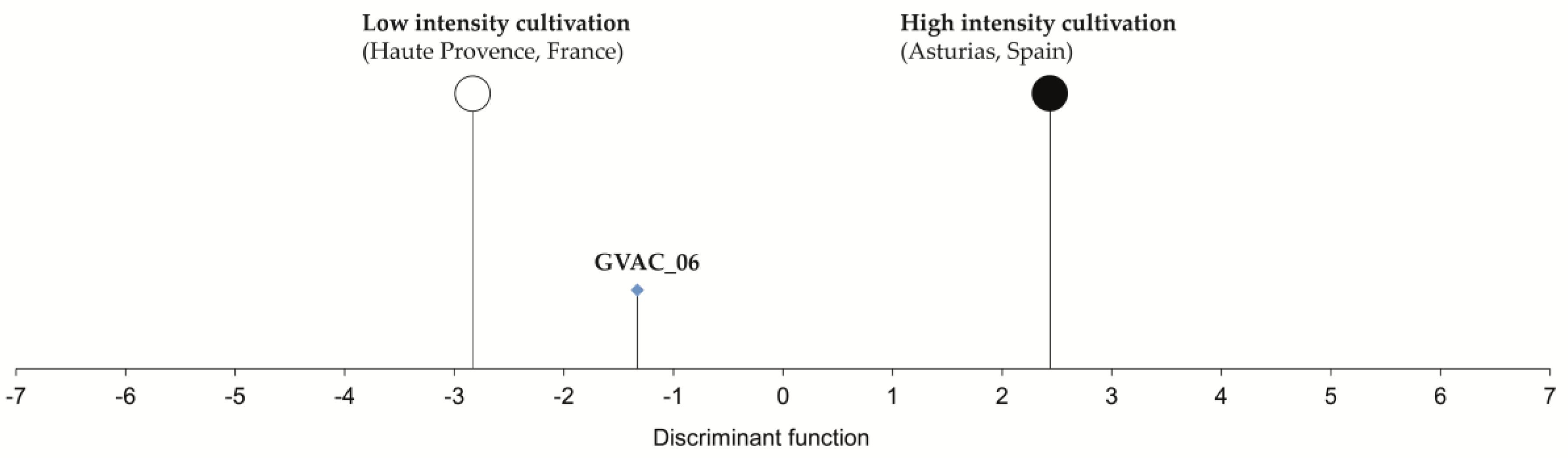
The results of the discriminant analysis (classified with a high probability of 99.7%) show that the GVAC_06 sample is grouped within the low-intensity cultivation spectrum of the Haute Provence fields, toward the high-intensity cultivation spectrum of the Asturias fields (
Figure 3). The results thus seemingly contradict those described above by suggesting cultivation practices with a low degree of disturbance. The low-intensity fields from Haute Provence were ploughed, harrowed and rarely to never manured, while the high-intensity fields from Asturias were hand weeded, ploughed, harrowed and manured regularly [
62]. The data collected on the functional attributes of the weeds in the respective regions, however, especially relate to the productivity of the habitat, and thus to the degree to which the fields were manured [
62]. It is therefore likely that the results of the discriminant analysis of Wellerholz GVAC_06 mainly reflect a lack of manuring, while not necessarily excluding a more intensive practice of soil tillage and/or weed control such as ploughing and harrowing. A likely practice in winter-rye cultivation is the use of the mouldboard plough [
15,
25] (p. 206, p. 76), which is known to have been used in the surroundings of Göttingen from the 12th century onward [
30] (p. 458). In contrast to the ard, this is a type of heavy plough has been observed to intensively disturb the soil, effectively eliminating perennial arable weeds [
2] (p. 145). Hamerow et al. [
69] (p. 597) for example describe that in Stafford (England) the arable floras of extensively cultivated fields reflect an increasing intensity from the 12th century onward as a result of the more common use of the mouldboard plough. The arable weed dataset of Wellerholz GVAC_06 indeed predominantly consists of annual taxa (
Figure 2;
Table S1), indicating intensive tillage of the soil. This observation, combined with the results of the discriminant analysis therefore suggest that the field in which the rye from Wellerholz GVAC_06 was grown was intensively tilled with the mouldboard plough, but was likely not hand weeded or manured.
3.3. Reconstructing Soil Conditions
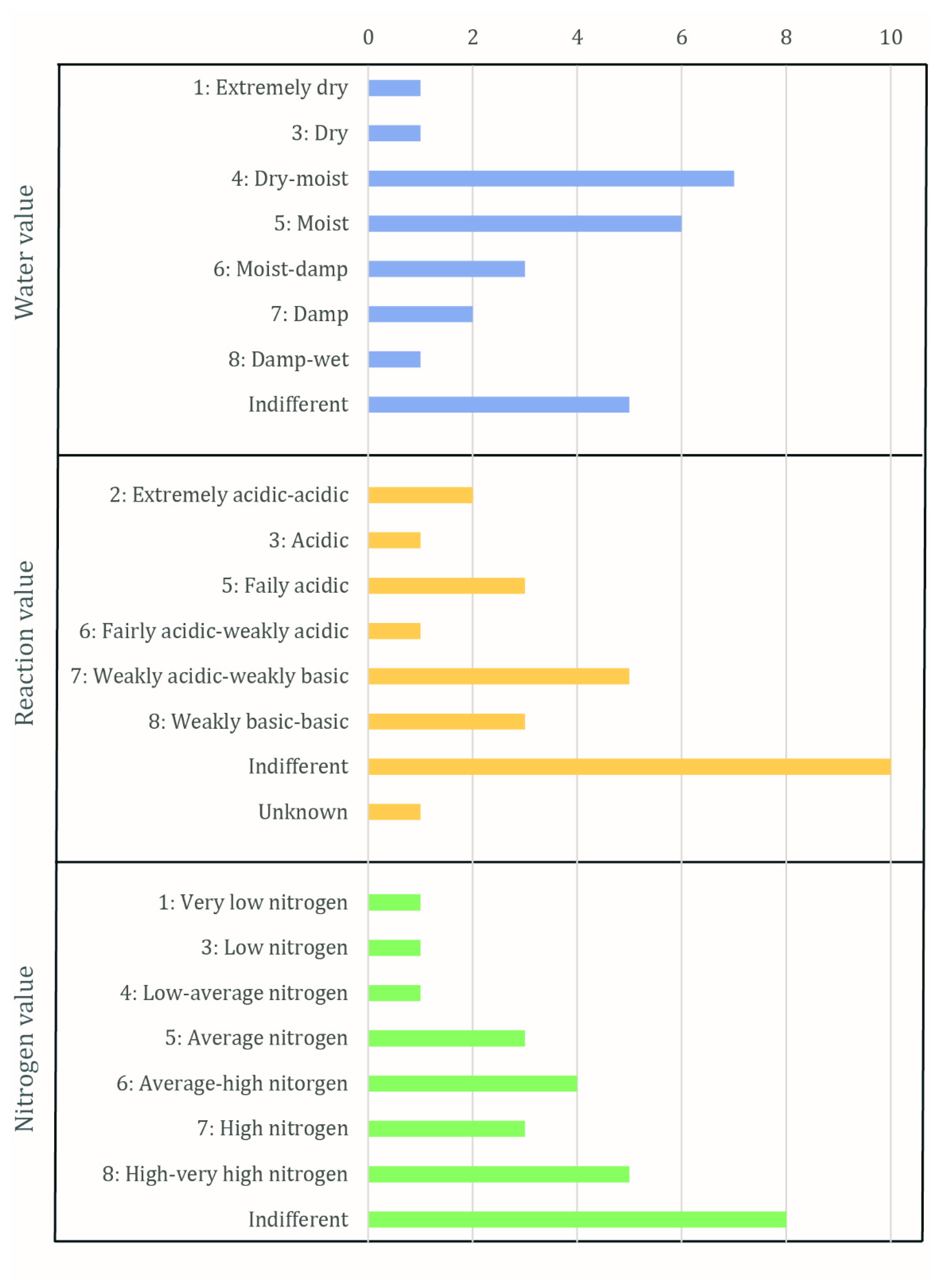
The results of Ellenberg’s indicator values for edaphic factors of the taxa of weeds and wild plants include a rather diverse range of values (
Figure 4;
Table S1). The water values show that most taxa prefer dry to moist (
n = 7) and moist (
n = 6) conditions. Five taxa are indifferent, while fewer prefer moist to damp (
n = 3) and damp (
n = 2) conditions. Extremely dry and dry conditions are represented by a single taxon each. The reaction values of the taxa indicate that 10 taxa are indifferent to soil pH. Five and three taxa, respectively, prefer to grow in weakly acidic to weakly basic and weakly basic to basic conditions. Fairly acidic to weakly acidic conditions are preferred by one taxon and fairly acidic conditions by three taxa. Acidic and extremely acidic conditions are preferred by one and two taxa, respectively. The reaction value of one taxon is unknown. The nitrogen values illustrate that eight taxa are indifferent to the nitrogen content of the soil. High to very high and high nitrogen contents are preferred by five and three taxa, respectively. Four plants prefer to grow in soil with average to high nitrogen content and three plants prefer an average nitrogen content.
On average, the values indicate moist or damp conditions (water value), more basic than acidic conditions (reaction value) and a moderate nitrogen content (nitrogen value). The water values and reaction values of the taxa correspond well to the soils and environmental conditions that prevail in the surroundings of Göttingen: the climate is temperate with a sufficient amount of rainfall to grow crops, and the typical soil in the surroundings of Göttingen is calcareous loess [
30] (pp. 457–458). The nitrogen values can be interpreted in the light of the results of the discriminant analysis described above, suggesting that manuring was not practiced on a regular basis. The diverse range of indicator values might further be interpreted as indicative of a low level of competition for minerals and water. Notably, low levels of competition are characteristic for highly disturbed habitats, especially among ruderals [
70] (p. 1171). The results of the indicator values can thus be seen to further support an interpretation of intensive soil tillage (see above).
3.4. Indications for Crop Rotation
It is interesting to note that most arable weeds in GVAC_06 are classified as Secalietea (community 3.4;
Table S1), which today are considered indicators of autumn sowing and/or extensive cultivation, while a smaller portion are classified as Chenopodietea, which today are typical of spring sowing and/or intensive cultivation [
43] (p. 627). Segetal communities are known to have altered in composition through time, developing parallel to crops over thousands of years [
39,
43] (p. 186; p. 625). The communities of arable weeds that today are indicative of specific sowing times and intensity of cultivation therefore do not necessarily express similar conditions in the past [
39] (p. 186). In the case of Wellerholz GVAC_06, however, the analysis of autecological data and functional attributes mostly correspond to the Ellenberg classification, suggesting that at the time when the rye was cultivated, the arable weed flora had already achieved the typical composition that it has today.
Despite the existing division into communities of Secalietea and Chenopodietea, many of the taxa are not strictly confined to the respective communities and therefore variations do exist in their composition [
43] (p. 628). The occurrence of portions of Chenopodietea in extensive and/or autumn-sown fields could, for instance, be an indication of crop rotation with summer, and/or intensively cultivated, crops [
31,
43] (p. 60; p. 627). In Wellerholz GVAC_06, a small portion of the arable weed assemblage is indeed classified as Chenopodietea, which can be considered an indication of crop rotation rather than rye monoculture. A further indication of crop rotation is the presence of a small portion of cultivated crops other than rye, which include bread wheat, hulled barley, common oat, flax and opium poppy (
Table 4). According to Martin and Sauerborn [
29] (p. 135), crops cultivated in previous rotation are able to emerge as part of the arable flora in the following rotations, although contradicting ethnographic observations by Jones and Halstead [
71] (p. 112) suggest that minor admixtures of other crops are more likely the result of contamination on the threshing floor rather than contamination from previous rotations. In this case, the crop taxa observed as admixtures in Wellerholz GVAC_06 fit into the typical model of the three-field system in the surroundings of Göttingen during the Middle Ages and Early Modern period, whereby rye, wheat and possibly barley were grown as winter crops, and oat, flax and poppy were grown as summer crops [
30] (pp. 457–458). It is therefore likely that, combined with the occurrence of different classes of arable weeds, the admixtures of crops in the rye harvest should be seen as evidence for the practice of the three-field system.
3.5. Implications for Harvesting Height
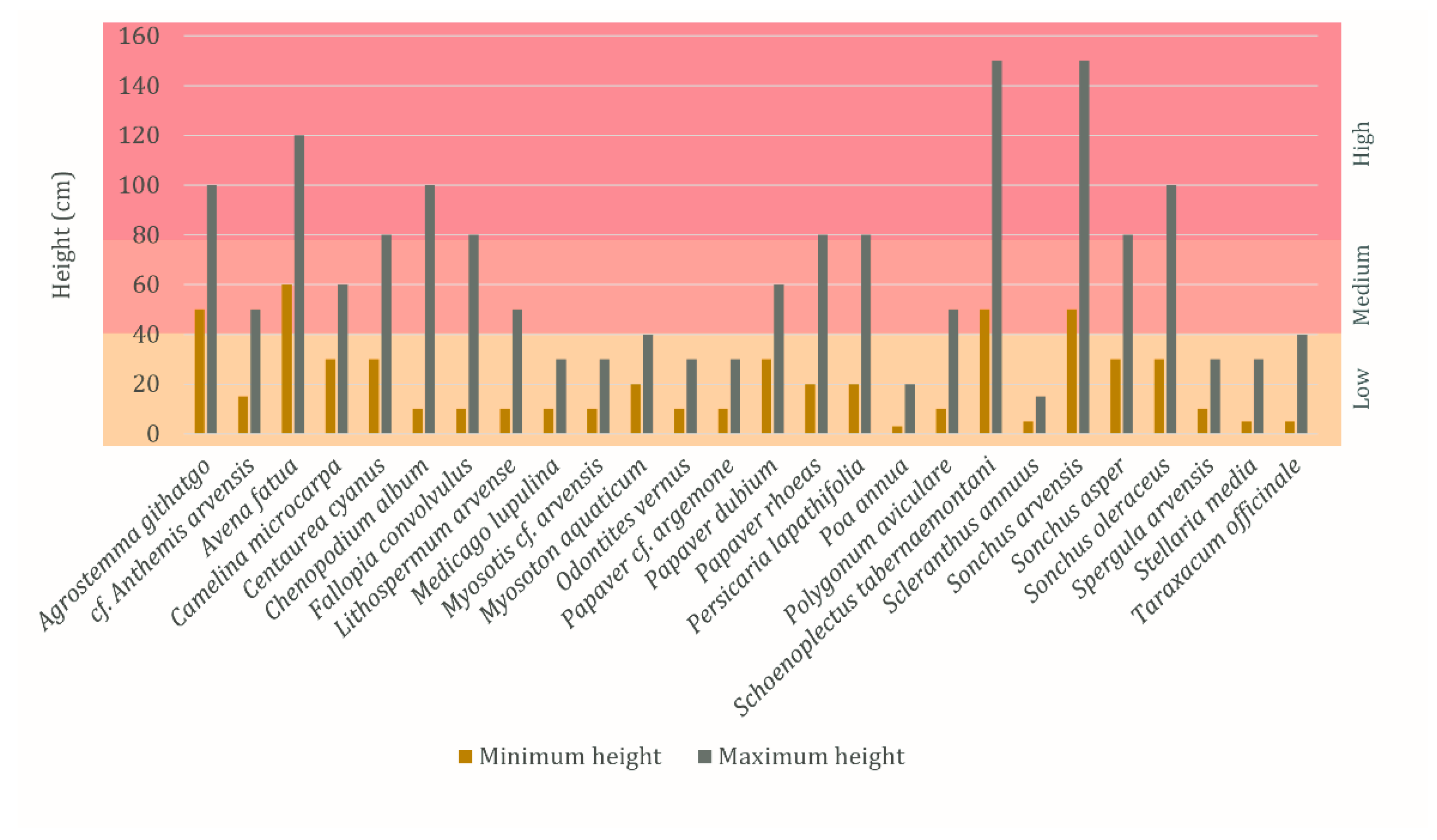
The minimum and maximum growing heights of the weeds and wild plants are shown in
Figure 5 and
Table S1. Low minimum growing heights (0–40 cm) are the most common and low and medium (41–80 cm) maximum growing heights are equally represented. Four taxa, respectively, have a medium minimum growing height and a high maximum growing height.
A total of 260 individual straw remains were measured that could be identified as rye straw due to the presence of rye ears. The measurements were subdivided into seven height categories 0–20 cm, 21–40 cm, 41–60 cm, 61–80 cm, 81–100 cm, 101–120 cm and 121–140 cm (
Figure 6). The majority of the straw was fragmented into short segments and thus fall into category A. The following categories are represented by less counts, with the tallest category (category G) containing a single measurement.
The minimum and maximum growing heights of the weeds and wild plants are various, ranging from short-growing plants (e.g., Medicago lupulina and Poa annua) to tall-growing plants (e.g., Schoenoplectus tabernaemontani and Sonchus arvensis). The maximum height of rye straw attested in Wellerholz GVAC_06 reached a height of 140 cm. The diversity in growing heights of weeds and wild plants alongside the presence of long straw segments of rye, suggests that the rye had been harvested by cutting close to the ground.
3.6. Sequence Analysis
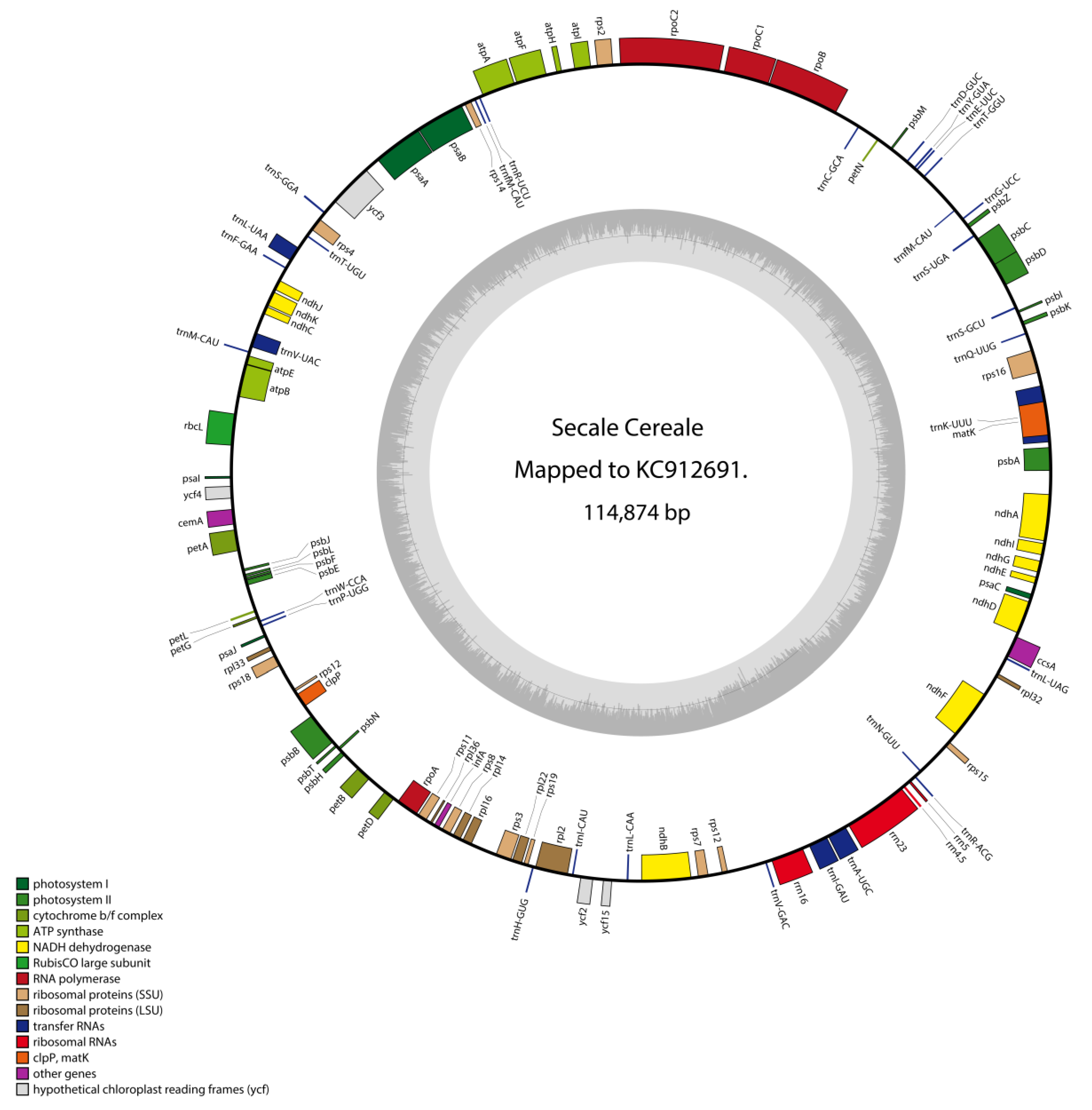
We were able to reconstruct and annotate the complete chloroplast sequence of the rye sample obtained from Wellerholz GVAC_06 by mapping to a modern reference (
Figure 7). The sequencing of the GVAC_06 sample resulted in 45,076,991 high quality reads. The chloroplast sequence consists of 114,874 base pairs and 57 open reading frames (ORFs) larger than 400 basepairs, including all known genetic domains, with a mean coverage of 29.3× (
Table S2). Compared to the reference sequence, we found 235 variable sites.
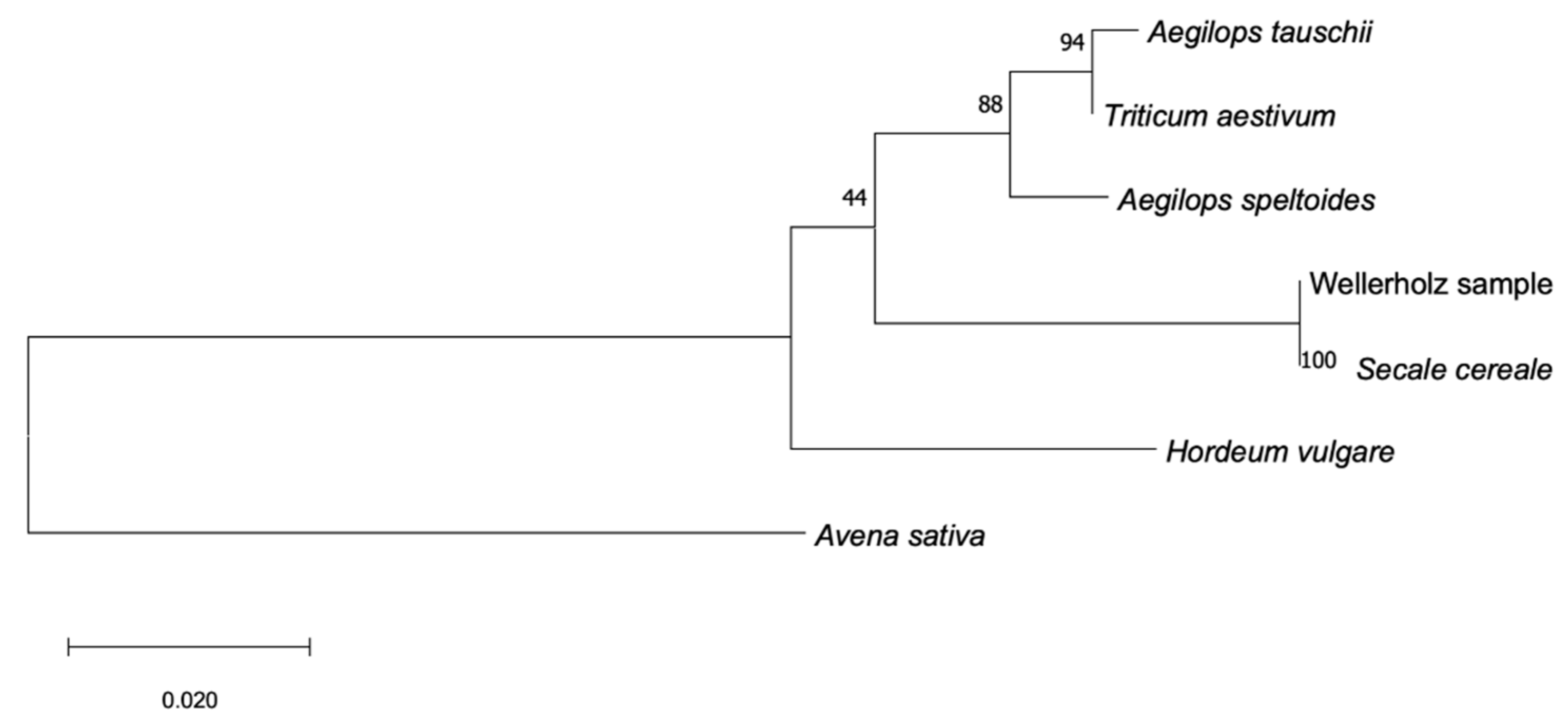
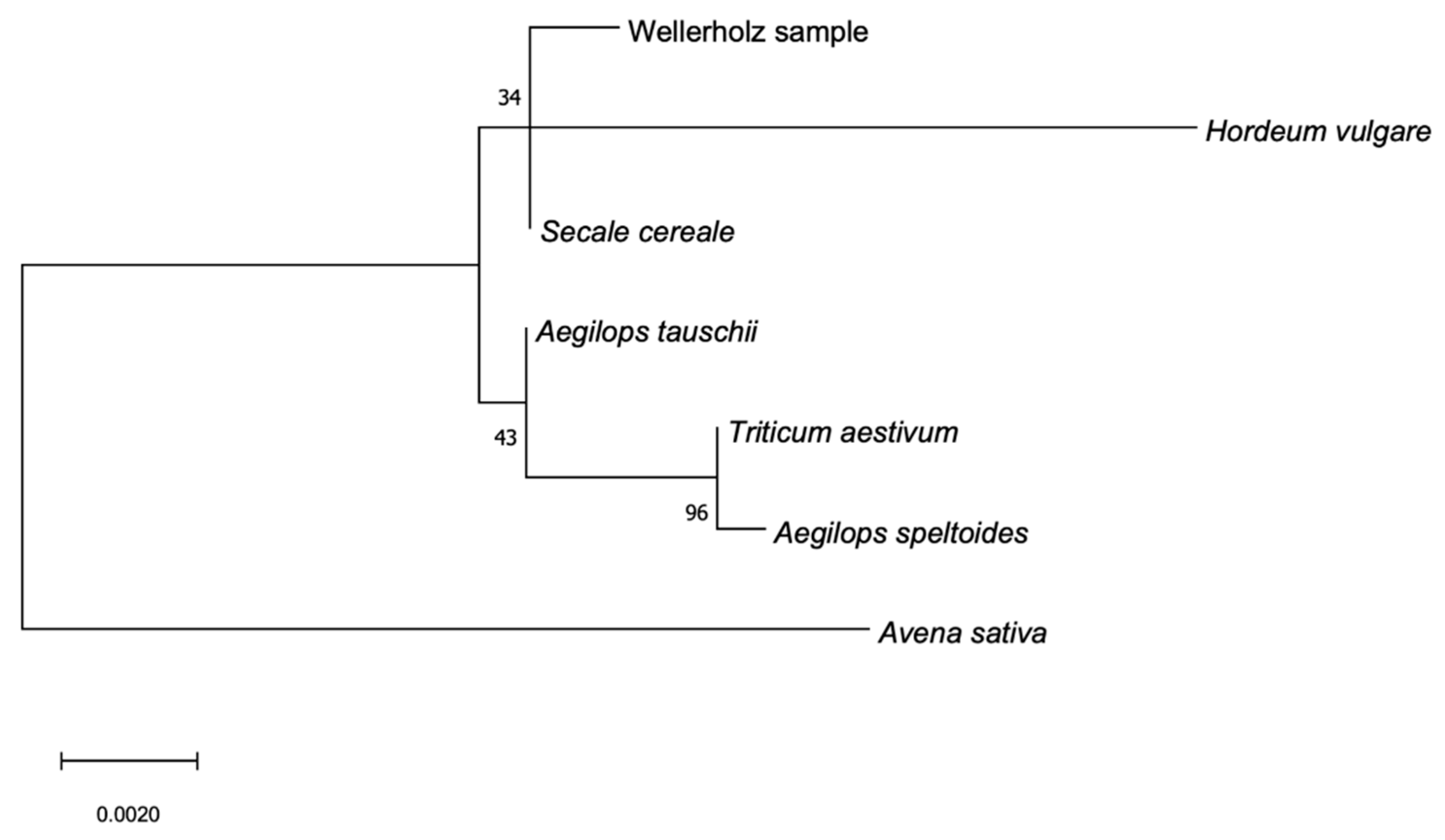
Phylogenetic analysis of the nuclear Internal Transcribed Spaces (ITS) locus (
Figure 8) as well as the chloroplast matK region (
Figure 9) show the GVAC_06 Wellerholz-sample forming a clade with the
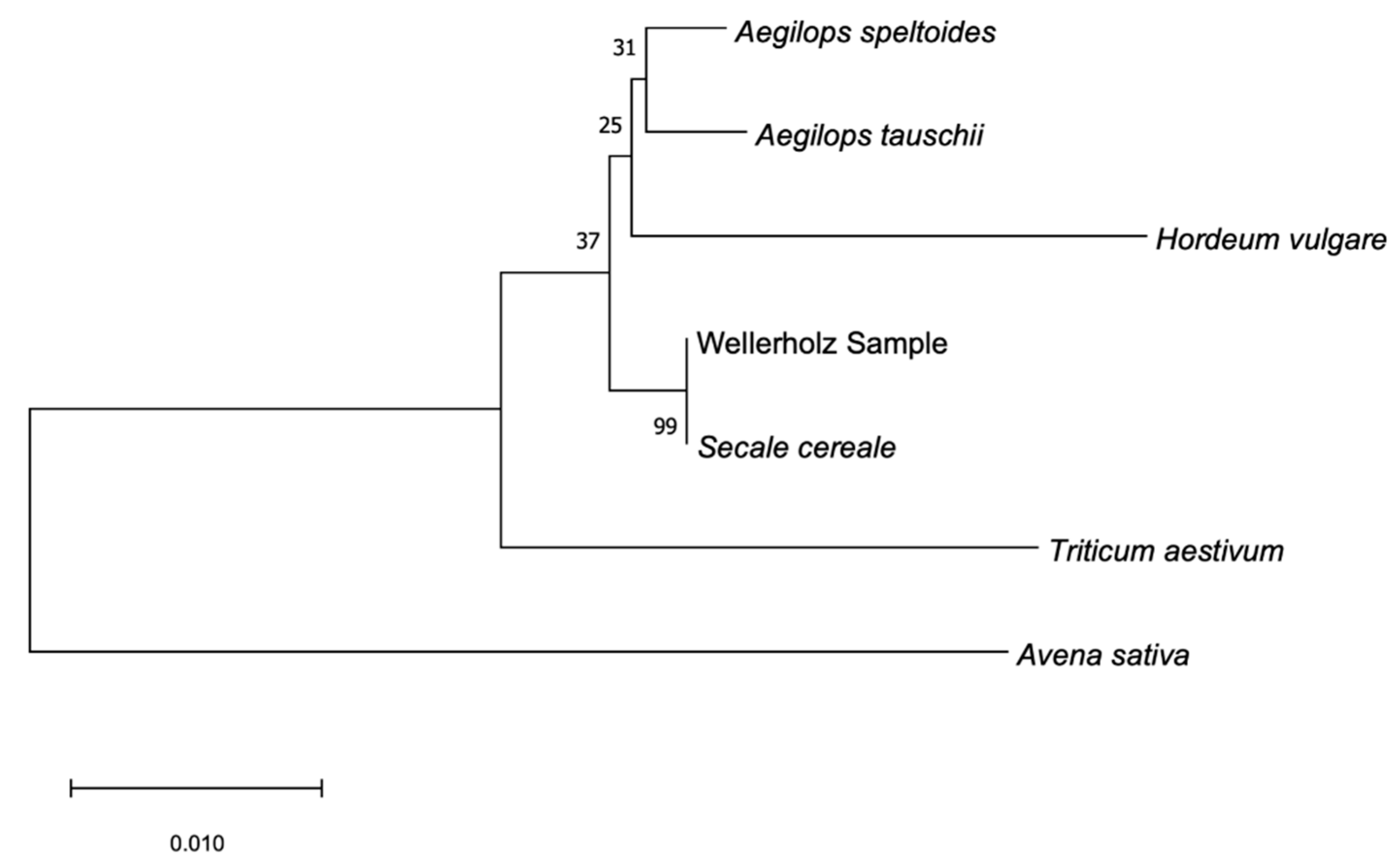
Secale cereale reference, indicating close relationship between the two, while the analysis of the rbCL-region (
Figure 10) suggests more ambiguities between the Wellerholz sample and the
Hordeum vulgare and
Secale cereale references. Higher bootstrapping values for ITS and matK phylogenetic trees establish higher confidence in their accuracy in presenting the most likely phylogenetic relationship. This suggests that matK and ITS markers are more suitable for phylogenetic analysis of the Triticeae tribe than the rbCL region. The ITS region has a mean coverage of 130.8×, out of the total 807 covered sites, 105 were variable and 34 parsimony informative.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}