
Genetic Improvement of Post-Heading Root Morphology and Physiology Facilitating Yield Increase of japonica Inbred Rice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site, Rice Cultivar, Field Design, and Crop Establishment
2.2. Sampling and Measurement
2.3. Statistical Analysis
3. Results
3.1. Grain Yield
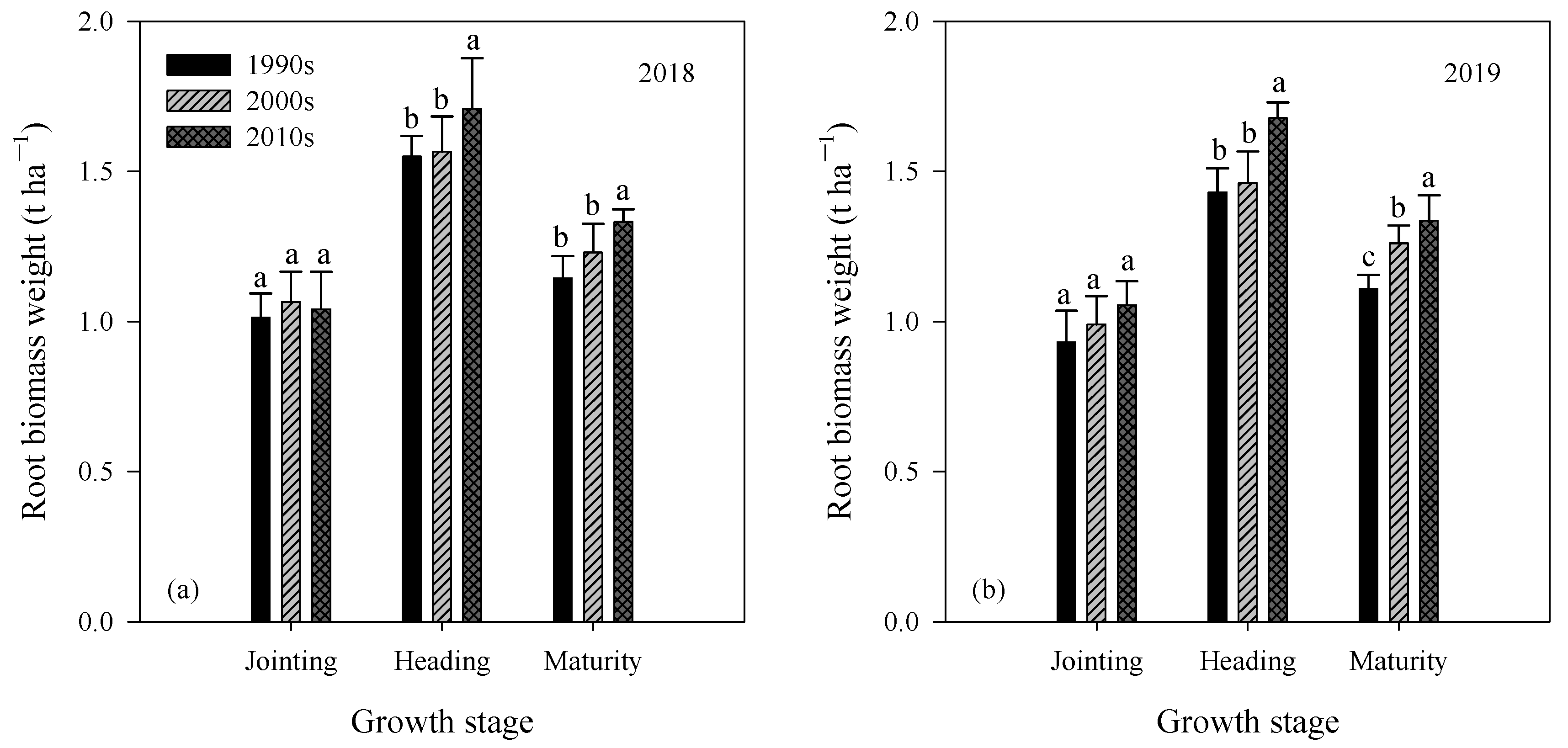
3.2. Biomass of Shoot and Root
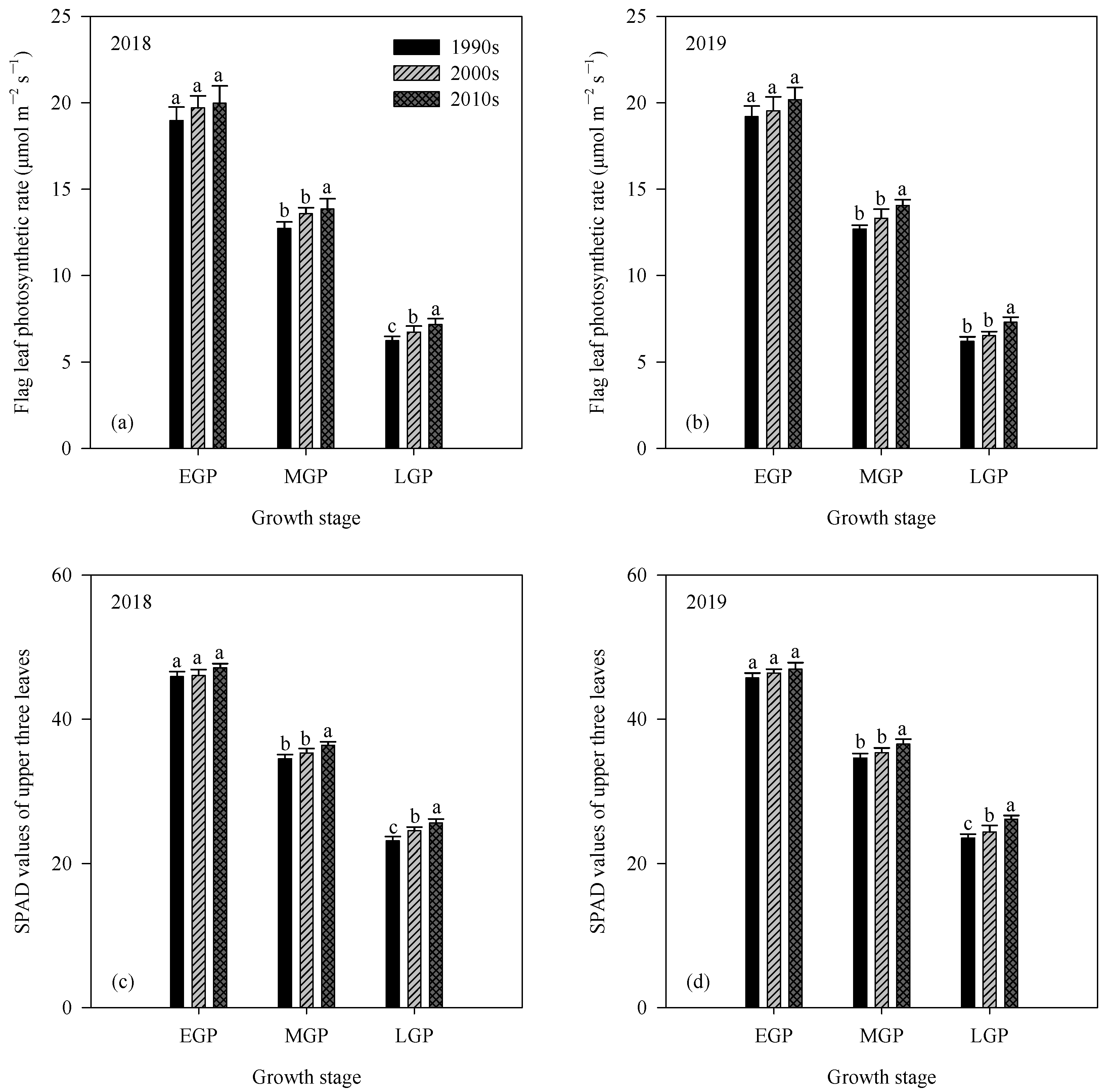
3.3. LAI, Leaf Photosynthetic Rate, and SPAD Values
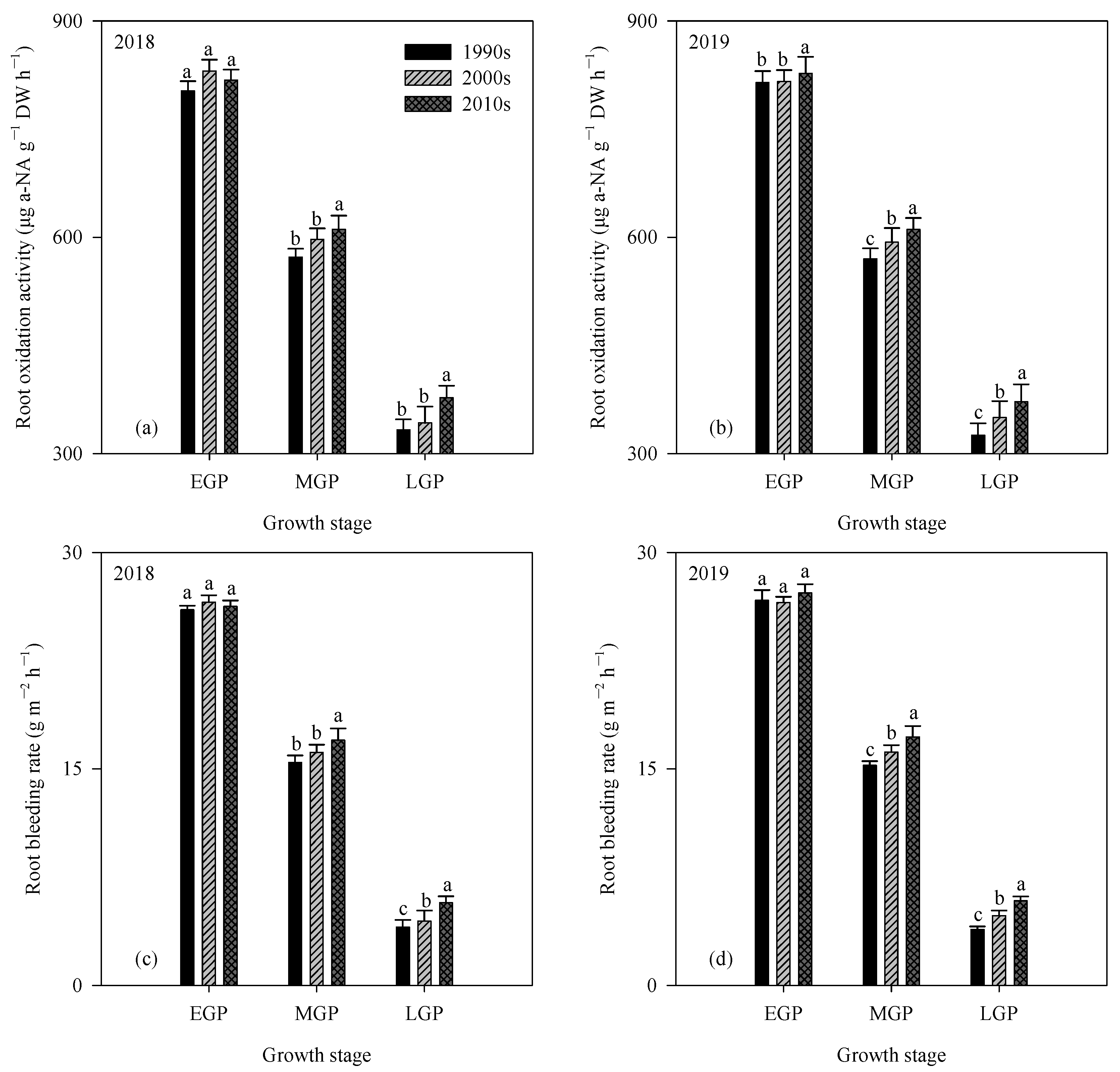
3.4. Root Length and Volume, Root Oxidation Activity, and Root Bleeding Rate
3.5. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Birla, D.S.; Malik, K.; Sainger, M.; Chaudhary, D.; Jaiwal, R.; Jaiwal, P.K. Progress and challenges in improving the nutritional quality of rice (Oryza sativa L.). Crit. Rev. Food Sci. 2017, 57, 2455–2481. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.P. Development of hybrid rice to ensure food security. Rice Sci. 2014, 21, 1–2. [Google Scholar] [CrossRef]
- Yu, Y.Q.; Huang, Y.; Zhang, W. Changes in rice yields in China since 1980 associated with cultivar improvement, climate and crop management. Field Crop Res. 2012, 136, 65–75. [Google Scholar] [CrossRef]
- Breseghello, F.; de Morais, O.P.; Pinheiro, P.V.; Silva, A.C.S.; de Castro, E.M.; Guimarães, É.P.; de Castro, A.P.; Pereira, J.A.; Lopes, A.M.; Utumi, M.M.; et al. Results of 25 years of upland rice breeding in Brazil. Crop Sci. 2011, 51, 914–923. [Google Scholar] [CrossRef]
- Wu, Z.H.; Xu, K.Z.; Zhao, Y.J.; He, X.L.; Wang, X.L.; Ling, F.L. Changes of some agronomic traits in japonica rice varieties during forty-seven years of genetic improvement in Jilin province, China. Chin. J. Rice Sci. 2007, 21, 507–512. [Google Scholar]
- Zhu, G.L.; Peng, S.B.; Huang, J.L.; Cui, K.H.; Nie, L.X.; Wang, F. Genetic improvements in rice yield and concomitant increases in radiation- and nitrogen-use efficiency in middle reaches of Yangtze River. Sci. Rep.-UK 2016, 6, 21049. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Laza, R.C.; Visperas, R.M.; Sanico, A.L.; Cassman, K.G.; Khush, G.S. Grain yield of rice cultivars and lines developed in the Philippines since 1966. Crop Sci. 2000, 40, 307–314. [Google Scholar] [CrossRef]
- Tabien, R.E.; Samonte, S.O.P.B.; McClung, A.M. Forty-eight years of rice improvement in Texas since the release of cultivar Bluebonnet in 1944. Crop Sci. 2008, 48, 2097–2106. [Google Scholar] [CrossRef]
- Yamamoto, T.; Suzuki, T.; Suzuki, K.; Adachi, S.; Sun, J.; Yano, M.; Ookawa, T.; Hirasawa, T. Characterization of a genomic region that maintains chlorophyll and nitrogen contents during ripening in a high-yielding stay-green rice cultivar. Field Crop Res. 2017, 206, 54–64. [Google Scholar] [CrossRef]
- Zhu, G.L.; Ren, Z.; Liu, Y.Q.; Lu, F.G.; Gu, L.F.; Shi, Y.; Liu, J.W.; Zhou, G.S.; Nimir, N.E.A.; Mohapatra, P.K. Optimization of leaf properties and plant phenotype through yield-based genetic improvement of rice over a period of seventy years in the Yangtze River basin of China. Food Energy Secur. 2020, 9, e223. [Google Scholar] [CrossRef]
- Rogers, E.D.; Benfey, P.N. Regulation of plant root system architecture: Implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [Google Scholar] [CrossRef]
- Ning, P.; Li, S.; Li, X.X.; Li, C.J. New maize hybrids had larger and deeper post-silking root than old ones. Field Crop Res. 2014, 166, 66–71. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Chen, F.J.; Chen, X.C.; Long, L.Z.; Gao, K.; Yuan, L.X.; Zhang, F.S.; Mi, G.H. Genetic improvement of root growth contributes to efficient phosphorus acquisition in maize (Zea mays L.). J. Integr. Agric. 2013, 12, 1098–1111. [Google Scholar] [CrossRef]
- Cui, X.; Dong, Y.; Gi, P.; Wang, H.; Xu, K.; Zhang, Z. Relationship between root vigour, photosynthesis and biomass in soybean cultivars during 87 years of genetic improvement in the northern China. Photosynthetica 2016, 54, 81–86. [Google Scholar] [CrossRef]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Dreccer, M.F.; Watt, M. Larger root systems: Are they useful in adapting wheat to dry environments. Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Tian, Z.W.; Fan, Y.H.; Yin, M.; Wang, F.R.; Cai, J.; Jiang, D.; Dai, T.B. Genetic improvement of root growth and its relationship with grain yield of wheat cultivars in the middle-lower Yangtze River. Acta Agron. Sin. 2015, 41, 613–622. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, Z.H.; Wang, J.C.; Wang, Z.Q.; Yang, J.C. Changes in morphological and physiological traits of roots and their relationships with grain yield during the evolution of mid-season indica rice cultivars in Jiangsu Province. Acta Agron. Sin. 2011, 37, 1020–1030. [Google Scholar] [CrossRef]
- Zhang, H.C.; Zhang, J.; Gong, J.L.; Chang, Y.; Li, M.; Gao, H.; Dai, Q.G.; Huo, Z.Y.; Xu, K.; Wei, H.Y. The productive advantages and formation mechanisms of “indica rice to japonica rice”. Sci. Agric. Sin. 2013, 46, 686–704. [Google Scholar]
- Wang, C.L.; Zhang, Y.D.; Zhu, Z.; Chen, T.; Zhao, Q.Y.; Zhong, W.G.; Yang, J.; Yao, S.; Zhou, L.H.; Zhao, L.; et al. Research progress on the breeding of japonica super rice varieties in Jiangsu Province, Chin. J. Integr. Agric. 2017, 16, 992–999. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.T.; Cui, W.P.; Xu, K.; Gao, H.; Wei, H.Y.; Zhang, H.C. Effects of early- and late-sowing on starch accumulation and associated enzyme activities during grain filling stage in rice. Rice Sci. 2021, 28, 191–199. [Google Scholar]
- Zhang, W.Y.; Yu, J.X.; Xu, Y.J.; Wang, Z.Q.; Liu, L.J.; Zhang, H.; Gu, J.F.; Zhang, J.H.; Yang, J.C. Alternate wetting and drying irrigation combined with the proportion of polymer-coated urea and conventional urea rates increases grain yield, water and nitrogen use efficiencies in rice. Field Crop Res. 2021, 268, 101865. [Google Scholar] [CrossRef]
- Ando, T.; Yoshida, S.; Nishiyama, I. Nature of oxidizing power of rice roots. Plant Soil 1983, 72, 57–71. [Google Scholar] [CrossRef]
- Meng, T.Y.; Wei, H.H.; Li, X.Y.; Dai, Q.G.; Huo, Z.Y. A better root morpho-physiology after heading contributing to yield superiority of japonica/indica hybrid rice. Field Crop Res. 2018, 228, 135–146. [Google Scholar] [CrossRef]
- Anzoua, K.G.; Junichi, K.; Toshihiro, H.; Kazuto, I.; Yutaka, J. Genetic improvements for high yield and low soil nitrogen tolerance in rice (Oryza Sativa L.) under a cold environment. Field Crop Res. 2010, 116, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Saito, H.; Fukuta, Y.; Obara, M.; Tomita, A.; Ishimaru, T.; Sasaki, K.; Fujita, D.; Kobayashi, N. Two novel QTLs for the harvest index that contribute to high-yield production in rice (Oryza sativa L.). Rice 2021, 14, 18. [Google Scholar] [CrossRef]
- Liu, L.L.; Wang, E.L.; Zhu, Y.; Tang, L.; Cao, W.X. Effects of warming and autonomous breeding on the phenological development and grain yield of double-rice systems in China. Agric. Ecosyst. Environ. 2013, 165, 28–38. [Google Scholar] [CrossRef]
- Jiang, Y.; Qian, H.Y.; Wang, L.; Feng, J.F.; Huang, S.; Hungate, B.A.; van Kessel, C.; Horwath, W.R.; Zhang, X.Y.; Qin, X.B.; et al. Limited potential of harvest index improvement to reduce methane emissions from rice paddies. Glob. Chang. Biol. 2019, 25, 686–698. [Google Scholar] [CrossRef]
- Marcaida, M., III; Li, T.; Angeles, O.; Evangelista, G.K.; Fontanilla, M.A.; Xu, J.L.; Gao, Y.M.; Li, Z.K.; Ali, J. Biomass accumulation and partitioning of newly developed Green Super Rice (GSR) cultivars under drought stress during the reproductive stage. Field Crop Res. 2014, 162, 30–38. [Google Scholar] [CrossRef]
- Niones, J.M.; Suralta, R.R.; Inukai, Y.; Yamauchi, A. Field evaluation on functional roles of root plastic responses on dry matter production and grain yield of rice under cycles of transient soil moisture stresses using chromosome segment substitution lines. Plant Soil 2012, 359, 107–120. [Google Scholar] [CrossRef]
- Atere, C.T.; Ge, T.; Zhu, Z.K.; Liu, S.L.; Huang, X.Z.; Shibsitova, O.; Guggenberger, G.; Wu, J.S. Assimilate allocation by rice and carbon stabilisation in soil: Effect of water management and phosphorus fertilization. Plant Soil 2019, 445, 153–167. [Google Scholar] [CrossRef]
- Madhu, M.; Hatfield, J.L. Dynamics of plant root growth under increased atmospheric carbon dioxide. Agron. J. 2013, 105, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Zang, H.D.; Xiao, M.L.; Wang, Y.D.; Ling, N.; Wu, J.S.; Ge, T.; Kuzyakov, Y. Allocation of assimilated carbon in paddies depending on rice age, chase period and N fertilization: Experiment with 13CO2 labelling and literature synthesis. Plant Soil 2019, 445, 113–123. [Google Scholar] [CrossRef]
- Kobata, T.; Shinonaga, M.; Yoshida, H.; Tomisaka, K.; Akai, K. Stay-green trait assessment using the leaf incubation method to examine the maintenance of assimilation rates under high temperature conditions during the grain-filling period in rice. Plant Prod. Sci. 2015, 18, 254–266. [Google Scholar] [CrossRef]
- Shin, D.; Lee, S.; Kim, T.H.; Lee, J.H.; Park, J.; Lee, J.Y.; Cho, L.H.; Choi, J.Y.; Lee, W.; Park, D.W.; et al. Natural variations at the Stay-Green gene promoter control lifespan and yield in rice cultivars. Nat. Commun. 2020, 11, 2819. [Google Scholar] [CrossRef]
- Dass, A.; Chandra, S.; Choudhary, A.K.; Singh, G.; Sudhishri, S. Influence of field re-ponding pattern and plant spacing on rice root–shoot characteristics, yield, and water productivity of two modern cultivars under SRI management in Indian Mollisols. Paddy Water Environ. 2016, 14, 45–59. [Google Scholar] [CrossRef]
- Wei, H.H.; Meng, T.Y.; Li, X.Y.; Dai, Q.G.; Zhang, H.C.; Yin, X.Y. Sink-source relationship during rice grain filling is associated with grain nitrogen concentration. Field Crop Res. 2018, 215, 23–38. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cultivar | Releasing Year | Cross Information | Cumulative Planting Area (Mha) |
|---|---|---|---|
| Wuyujing 3 | 1992 | Zhongdan 1/79-51 × Zhongdan 1/Yangjing 1 | 6.1 |
| Zhendao 88 | 1997 | Yuezhiguang × 2507-4 | 0.5 |
| Zaofeng 9 | 1997 | Wufujing/Zhongdan 1 × Nonglin 205 | 1.0 |
| Wujing 13 | 2003 | 791 × SR 21 | 0.3 |
| Huaidao 9 | 2006 | Huai 9712 | 0.5 |
| Nanjing 44 | 2007 | Nanjing 38 | 0.5 |
| Zhendao 18 | 2013 | Zhendao 99 × Wuyunjing 7 | 0.2 |
| Nanjing 9108 | 2013 | Wuxiangjing 14 × Guandong 194 | 1.8 |
| Nanjing 2728 | 2018 | Wujing 15 × Nanjing 5055 | 0.1 |
| Year | Cultivar Type | Grain Yield (t ha−1) | Panicles per m2 | Spikelets per Panicle | Filled-Grain Percentage (%) | 1000-Grain Weight (g) |
|---|---|---|---|---|---|---|
| 2018 | 1990s | 10.1 ± 0.2 b | 319 ± 12 a | 128 ± 6 c | 89.6 ± 0.5 ab | 27.6 ± 0.3 a |
| 2000s | 10.4 ± 0.2 b | 307 ± 10 ab | 143 ± 7 b | 89.9 ± 0.6 a | 27.0 ± 0.4 a | |
| 2010s | 10.8 ± 0.2 a | 290 ± 8 b | 158 ± 6 a | 88.5 ± 0.7 b | 27.3 ± 0.2 a | |
| 2019 | 1990s | 10.0 ± 0.3 c | 322 ± 5 a | 121 ± 6 c | 90.1 ± 0.8 a | 27.5 ± 0.4 a |
| 2000s | 10.4 ± 0.3 b | 310 ± 6 a | 141 ± 6 b | 89.1 ± 0.3 b | 27.3 ± 0.3 a | |
| 2010s | 10.9 ± 0.2 a | 279 ± 14 b | 163 ± 4 a | 88.7 ± 0.4 b | 26.8 ± 0.4 b | |
| Analysis of variance (ANOVA) | ||||||
| Year | ns | ns | ns | ns | ns | |
| Cultivar type | ** | ** | ** | ** | ns | |
| Year × Cultivar type | ns | ns | ns | ns | ns | |
| Year | Cultivar Type | Shoot Biomass Weight (t ha−1) | Harvest Index | Shoot Biomass Accumulation (t ha−1) | |||
|---|---|---|---|---|---|---|---|
| Jointing | Heading | Maturity | Jointing-Heading | Heading-Maturity | |||
| 2018 | 1990s | 5.4 ± 0.3 a | 10.4 ± 0.3 b | 18.2 ± 0.3 b | 0.475 ± 0.005 b | 5.0 ± 0.6 a | 7.8 ± 0.4 b |
| 2000s | 5.5 ± 0.4 a | 10.5 ± 0.5 ab | 18.5 ± 0.4 b | 0.483 ± 0.006 a | 5.0 ± 0.5 a | 8.0 ± 0.2 b | |
| 2010s | 5.5 ± 0.2 a | 10.7 ± 0.6 a | 19.2 ± 0.3 a | 0.486 ± 0.005 a | 5.2 ± 0.6 a | 8.5 ± 0.3 a | |
| 2019 | 1990s | 5.3 ± 0.3 a | 10.4 ± 0.4 b | 18.4 ± 0.3 b | 0.470 ± 0.006 b | 5.1 ± 0.2 b | 8.0 ± 0.2 b |
| 2000s | 5.4 ± 0.2 a | 10.6 ± 0.3 b | 18.6 ± 0.4 b | 0.479 ± 0.003 a | 5.2 ± 0.5 a | 8.0 ± 0.3 b | |
| 2010s | 5.6 ± 0.5 a | 11.0 ± 0.3 a | 19.5 ± 0.3 a | 0.482 ± 0.003 a | 5.4 ± 0.5 a | 8.5 ± 0.4 a | |
| Analysis of variance (ANOVA) | |||||||
| Year | ns | ns | ns | ns | ns | ns | |
| Cultivar type | ns | * | ** | ** | ns | ** | |
| Year × Cultivar type | ns | ns | ns | ns | ns | ns | |
| Year | Cultivar Type | Root Biomass Weight at Jointing (kg ha−1) | Root Biomass Weight at Heading (kg ha−1) | Root Biomass Weight at Maturity (kg ha−1) | |||
|---|---|---|---|---|---|---|---|
| 0–15 cm | 15–30 cm | 0–15 cm | 15–30 cm | 0–15 cm | 15–30 cm | ||
| 2018 | 1990s | 888 ± 68 a | 124 ± 15 a | 1386 ± 64 a | 163 ± 11 b | 1023 ± 69 b | 120 ± 9 b |
| 2000s | 924 ± 78 a | 141 ± 22 a | 1394 ± 102 a | 172 ± 22 b | 1098 ± 79 ab | 132 ± 16 b | |
| 2010s | 903 ± 112 a | 138 ± 11 a | 1428 ± 147 a | 279 ± 30 a | 1157 ± 47 a | 175 ± 15 a | |
| 2019 | 1990s | 804 ± 79 a | 127 ± 26 a | 1273 ± 75 b | 157 ± 28 b | 983 ± 37 b | 126 ± 15 b |
| 2000s | 867 ± 83 a | 123 ± 13 a | 1297 ± 76 b | 164 ± 32 b | 1112 ± 26 a | 157 ± 24 b | |
| 2010s | 917 ± 58 a | 139 ± 22 a | 1456 ± 55 a | 221 ± 42 a | 1156 ± 100 a | 180 ± 31 a | |
| Analysis of variance (ANOVA) | |||||||
| Year | ns | ns | ns | ns | ns | ns | |
| Cultivar type | ns | ns | * | ** | ** | ** | |
| Year × Cultivar type | ns | ns | ns | ns | ns | ns | |
| Year | Cultivar Type | Leaf Area Index (LAI, m2 m−2) | ||
|---|---|---|---|---|
| EGP | MGP | LGP | ||
| 2018 | 1990s | 5.8 ± 0.3 c | 4.1 ± 0.3 c | 2.3 ± 0.2 b |
| 2000s | 7.2 ± 0.3 b | 5.0 ± 0.2 b | 2.6 ± 0.2 ab | |
| 2010s | 7.8 ± 0.2 a | 5.4 ± 0.1 a | 2.9 ± 0.2 a | |
| 2019 | 1990s | 6.1 ± 0.2 c | 4.3 ± 0.1 c | 2.4 ± 0.2 b |
| 2000s | 7.1 ± 0.4 b | 5.0 ± 0.2 b | 2.6 ± 0.2 b | |
| 2010s | 7.7 ± 0.2 a | 5.5 ± 0.1 a | 3.0 ± 0.2 a | |
| Analysis of variance (ANOVA) | ||||
| Year | ns | ns | ns | |
| Cultivar type | ** | ** | ** | |
| Year × Cultivar type | ns | ns | ns | |
| Year | Cultivar Type | EGP | MGP | LGP | |||
|---|---|---|---|---|---|---|---|
| Root Length (km m−2) | Root Volume (m3 ha−1) | Root Length (km m−2) | Root Volume (m3 ha−1) | Root Length (km m−2) | Root Volume (m3 ha−1) | ||
| 2018 | 1990s | 14.5 ± 0.8 b | 21.6 ± 1.2 b | 9.8 ± 0.5 b | 18.9 ± 0.6 b | 7.9 ± 0.7 b | 14.9 ± 1.0 b |
| 2000s | 15.7 ± 0.5 ab | 25.2 ± 0.6 a | 10.8 ± 0.6 ab | 19.2 ± 1.1 b | 8.4 ± 0.7 b | 15.8 ± 0.8 b | |
| 2010s | 16.9 ± 0.5 a | 26.4 ± 0.8 a | 11.5 ± 0.9 a | 20.3 ± 0.9 a | 9.9 ± 0.6 a | 17.3 ± 0.8 a | |
| 2019 | 1990s | 14.8 ± 0.6 b | 22.4 ± 1.0 b | 10.0 ± 0.4 b | 18.6 ± 0.5 b | 8.2 ± 0.7 b | 13.8 ± 0.8 c |
| 2000s | 15.8 ± 0.7 ab | 25.1 ± 0.4 a | 10.8 ± 0.5 ab | 18.6 ± 0.6 b | 8.9 ± 0.5 b | 15.6 ± 0.8 b | |
| 2010s | 16.7 ± 0.5 a | 26.5 ± 1.2 a | 11.6 ± 0.6 a | 20.7 ± 1.0 a | 9.7 ± 0.7 a | 17.2 ± 0.6 a | |
| Analysis of variance (ANOVA) | |||||||
| Year | ns | ns | ns | ns | ns | ns | |
| Cultivar type | ** | ** | ** | ** | ** | ** | |
| Year × Cultivar type | ns | ns | ns | ns | ns | ns | |
| Item | Shoot Biomass Weight | Grain Yield | |||
|---|---|---|---|---|---|
| Jointing | Heading | Maturity | |||
| Root biomass weight | Jointing | 0.49 * | 0.25 | 0.28 | 0.28 |
| Heading | 0.17 | 0.48 * | 0.52 * | 0.60 ** | |
| Maturity | 0.42 | 0.52 * | 0.83 ** | 0.77 ** | |
| Item | EGP | MGP | LGP | Shoot Biomass Accumulation from Heading to Maturity | Grain Yield | ||||
|---|---|---|---|---|---|---|---|---|---|
| Leaf Photosynthetic Rate | SPAD Values | Leaf Photosynthetic Rate | SPAD Values | Leaf Photosynthetic Rate | SPAD Values | ||||
| EGP | Root length | 0.52 * | 0.68 ** | 0.70 ** | 0.79 ** | 0.66 ** | 0.80 ** | 0.59 ** | 0.84 ** |
| Root volume | 0.63 ** | 0.54 * | 0.83 ** | 0.71 ** | 0.74 ** | 0.77 ** | 0.52 * | 0.84 ** | |
| Root oxidation activity | 0.25 | −0.11 | 0.43 | 0.14 | 0.40 | 0.29 | 0.65 ** | 0.48 * | |
| Root bleeding rate | 0.25 | 0.17 | 0.29 | 0.32 | 0.33 | 0.41 | 0.48 * | 0.29 | |
| MGP | Root length | 0.44 | 0.53 * | 0.67 ** | 0.68 ** | 0.74 ** | 0.73 ** | 0.68 ** | 0.77 ** |
| Root volume | 0.28 | 0.36 | 0.49 * | 0.51 * | 0.63 ** | 0.57 * | 0.72 ** | 0.74 ** | |
| Root oxidation activity | 0.60 ** | 0.38 | 0.83 ** | 0.58 * | 0.75 ** | 0.65 ** | 0.67 ** | 0.83 ** | |
| Root bleeding rate | 0.36 | 0.41 | 0.69 ** | 0.62 ** | 0.82 ** | 0.69 ** | 0.72 ** | 0.84 ** | |
| LGP | Root length | 0.35 | 0.56 * | 0.60 ** | 0.73 ** | 0.72 ** | 0.79 ** | 0.70 ** | 0.76 ** |
| Root volume | 0.51 * | 0.66 | 0.74 ** | 0.76 ** | 0.76 ** | 0.75 ** | 0.66 ** | 0.92 ** | |
| Root oxidation activity | 0.41 | 0.34 | 0.65 ** | 0.54 * | 0.65 ** | 0.62 ** | 0.74 ** | 0.84 ** | |
| Root bleeding rate | 0.52 * | 0.59 ** | 0.75 ** | 0.75 ** | 0.78 ** | 0.80 ** | 0.69 ** | 0.90 ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, T.; Chen, X.; Zhang, X.; Ge, J.; Zhou, G.; Dai, Q.; Wei, H. Genetic Improvement of Post-Heading Root Morphology and Physiology Facilitating Yield Increase of japonica Inbred Rice. Agronomy 2021, 11, 2457. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122457
Meng T, Chen X, Zhang X, Ge J, Zhou G, Dai Q, Wei H. Genetic Improvement of Post-Heading Root Morphology and Physiology Facilitating Yield Increase of japonica Inbred Rice. Agronomy. 2021; 11(12):2457. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122457
Chicago/Turabian StyleMeng, Tianyao, Xi Chen, Xubin Zhang, Jialin Ge, Guisheng Zhou, Qigen Dai, and Huanhe Wei. 2021. "Genetic Improvement of Post-Heading Root Morphology and Physiology Facilitating Yield Increase of japonica Inbred Rice" Agronomy 11, no. 12: 2457. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122457