
Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize



Department of Agricultural and Environmental Chemistry, University of Agriculture in Krakow, al. Mickiewicza 21, 31-120 Krakow, Poland
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(12), 2582; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122582
Submission received: 30 October 2021
/
Revised: 24 November 2021
/
Accepted: 17 December 2021
/
Published: 19 December 2021
(This article belongs to the Special Issue Nutrient Recycling: Reduction in the Application of Mineral Fertilizers by Optimization of the Organic Wastes)
Abstract
:As natural plant growth stimulators, amino acids are widely used to improve crop yield and quality. There are numerous studies documenting the influence of amino acids on plants, which is not always positive. This study was conducted to determine the effect of soil-applied L-tryptophan (L-TRP) on the accumulation and utilization of nitrogen and sulfur by maize. The study was carried out under the conditions of a pot experiment. The experimental design included three treatments: soil without fertilization (control), soil with mineral fertilization (NPKS), and soil with mineral fertilization and L-tryptophan addition (NPKS + L-TRP). The application of tryptophan to the soil, supported by mineral fertilization, caused a significant increase in maize biomass. Although no significant differences in nitrogen and sulfur contents in maize biomass were found between treatments without and with the addition of L-tryptophan, significantly higher intakes of both elements were observed in the NPKS + L-TRP treatment. The application of L-tryptophan increased the biosynthesis of Chlorophyll a. Utilization of nitrogen and sulfur by maize in the NPKS + L-TRP treatment was more than 27% and 17% higher, respectively, compared to the NPKS treatment. Maintaining the recommended contents of individual nutrients in the rhizosphere is not a guarantee of optimal quantitative and qualitative intake of nutrients. Problems with maintaining optimal relationships between individual nutrients may be compounded by soil properties.
1. Introduction
Growing problems resulting from deteriorating soil fertility indicators force a search for alternative ways to reduce or eliminate them. For many years, research has been conducted to introduce new fertilizer formulas or to improve those that are already available by adding various ingredients and substances. That research has been aimed at not only increasing the effectiveness of fertilizers, but also reducing losses of fertilizer components, thus addressing both the economic aspect and environmental quality issues [1,2].
Given the increasing demands on biomass for food and feed purposes, the importance of biologically active substances, often referred to as biostimulants, is emphasized. According to Regulation (EU) 2019/1009 of the European Parliament and of the Council [3], biostimulants are certain substances, mixtures, and microorganisms that do not provide nutrients but stimulate natural plant nutritional processes. The group of biostimulants can include products containing the so-called beneficial elements (titanium, silicon), products based on algae extracts, and products containing amino acids.
Amino acids are among some of the most important organic compounds, as they are one of the building blocks of proteins, hormones, and enzymes [4]. Plant cells contain low levels of protein compared to animal cells, mainly due to the large amount of carbohydrates (cellulose and others) that make up most of the plant structure. However, the importance of proteins and amino acids cannot be overlooked here. Apart from being protein components, amino acids are also involved in many cellular reactions and therefore influence a number of physiological processes, such as plant growth and development, as well as resistance to abiotic and biotic stresses [5,6,7]. For example, glycine and glutamic acids play important roles in the formation of vegetative tissue and chlorophyll [8,9].
Amino acids are organic compounds that are synthesized by plants, but this process involves energy expenditure and takes time. The most biologically active are plant-derived L-amino acids such as L-tryptophan (L-TRP). L-TRP is known as β-3-indolylalanine. It is one of the nine essential amino acids for humans. L-tryptophan is a unique amino acid bearing an indole ring. L-TRP is applied to soil, used as a foliar spray and seed priming [9]. Numerous study results indicate that soil-applied L-TRP is taken up directly by plants or metabolized to many products, including niacin, serotonin, and auxin by the soil microbiota, and subsequently absorbed by plant roots. As a source of nitrogen and carbon, L-TRP supports microbial growth and activity in the rhizosphere. As an auxin, it is absorbed by plants and regulates various physiological and biochemical processes in plants (Ahmad et al., 2008). There are studies that have indicated a positive response after L-TRP application to increase plant yield and improve yield quality [9,10,11].
Amino acids are precursors of growth substances [12]. However, they can also successfully act as nutrient carriers. Literature examples stated that amino acid molecules are electrically inert, which makes their transfer in plant cells through ectoderm to plasmalemma relatively easy. The addition of amino acids to fertilizers also significantly reduces the time of nutrient uptake, resulting in better nutrient management in the plant. Results of Garcia et al. [13] revealed that the addition of amino acids to the nutrient solution increased the content of Ca, K, Fe, Cu, Mn, and Mg in tomato leaves. Popko et al. [14] showed that the application of amino acid-based preparations increased winter wheat grain yield and improved grain technological parameters, including protein content and the Zeleny sedimentation index. Considering the effect of the applied amino acid preparations on the content of mineral components in wheat grain, an increase in the content of copper (in the range of 31–50%), sodium (35–43%), calcium (4.3–7.9%), and molybdenum (3.9–16%) was recorded. Therefore, it is confirmed scientific evidence of the beneficial effect of amino acids on the plant mineral content. On the one hand, this concerns the uptake of essential elements such as nitrogen (a building component) and sulfur (a component attributed with pro-quality properties in the formation of plant biomass), and on the other hand, this indicates a proenvironmental effect relevant to the dispersal of biogens.
As natural plant growth stimulators, amino acids are widely used to improve crop yield and quality [15]. Many studies have documented the effect of different amino acids on plants, and it was not always positive, and depended largely on the amount of the amino acid applied [9,12,16]. The study by Kravchenko et al. [17] showed that radish roots secrete 290–390 ng tryptophan per seedling per day. The inoculation of radish plants with rhizosphere pseudomonads increased the root biomass 1.4 times. The beneficial effect of inoculation of radish plants with rhizosphere pseudomonads on radish plants can be explained by the fact that the introduced rhizobacteria produce a plant growth-stimulating hormone, indole-3-acetic acid, whose precursor is tryptophan [9]. The phytohormonal action of the rhizosphere microflora was found to be effective, provided that the tryptophan concentration in the rhizosphere was sufficiently high. In this regard, a study was carried out to determine the effect of soil-applied L-tryptophan on the accumulation and utilization of nitrogen and sulfur by maize.
2. Materials and Methods
2.1. Growing Experiment
The study was carried out in 2019 on silty soil collected from the 0–0.2 m layer in southern Poland. The properties of the soil used in the experiment are presented in Table 1. The following parameters were determined in dried, 1 mm sieved initial soil samples: granulometric composition—according to PN-R-04032, pH—potentiometrically in the suspension of soil and water as well as soil and 1 mol dm−3 solution of KCl (soil: solution = 1:2.5), and electrical conductivity (EC)—conductometrically. Total contents of carbon, nitrogen, and sulfur were determined with the CNS analyzer (Vario MAX Cube, Elementar Analysensysteme). Total contents of phosphorus, potassium, calcium, magnesium, and sodium were determined after ashing the sample in a chamber furnace at 450 °C for 12 h and mineralizing its residues in a mixture of concentrated nitric and perchloric acids (3:2) (v/v). Contents of the studied elements were determined in the obtained solutions by inductively coupled plasma optical emission spectrometry (ICP-OES, Perkin Elmer Optima 7300 DV) [18].
The growing experiment was conducted in containers with a capacity of 1400 g of soil dry mass. The experimental design included three treatments carried out in three replicates: treatment without fertilization (control), treatment with mineral fertilization (NPKS), and treatment with mineral fertilization and L-tryptophan (NPKS + L-TRP). In the experiment, nitrogen was applied in the form of chemically pure salts NH4NO3 and (NH4)2SO4, phosphorus in the form of Ca(H2PO4)2·H2O, potassium in the form of KCl, and sulfur in the form of (NH4)2SO4. L-tryptophan (BIOSYNTH® Carbosynth) was also used in the study. The doses of minerals were as follows: N 0.098 g kg−1 DM soil, P 0.075 g kg−1 DM soil, K 0.108 g kg−1 DM soil, S 0.036 g kg−1 DM soil. L-tryptophan was applied in a dose of 0.010 g kg−1 DM soil. After introducing mineral salt fertilizers and L-tryptophan, distilled water (up to 45% water capacity) was added to soils and thoroughly mixed with the material. The soil was then placed into PVC containers. After 24 h, seeds of Zea mays L. of the Karpatis FAO 249 variety were sown. The plant density per pot was four plants. Soil moisture was maintained at 60% of the maximum water holding capacity. The experiment was carried out under laboratory conditions for 60 days. After that time, the aerial parts of the plants were collected, and their roots separated from the soil block and thoroughly washed with distilled water. The collected biomass was dried to a constant weight at 65 °C, and its amount determined.
2.2. Analysis of Plant Material
Chlorophyll a and b contents were determined spectrophotometrically in fresh maize leaf samples after extraction with acetone [19]. The ash content in the dried and ground plant material was determined after ashing the sample in a chamber furnace at 550 °C for 16 h. Nitrogen content was determined with the CNS analyzer (Vario MAX Cube, Elementar Analysensysteme). Sulfur content was determined by inductively coupled plasma optical emission spectrometry (ICP-OES, Perkin Elmer Optima 7300 DV) after oxidizing the plant material in concentrated nitric acid and binding it with magnesium nitrate solution [20].The contents of other macronutrients (K, Ca, Na, Mg) in the aboveground parts of maize were determined after ashing the sample in a chamber furnace at 450 °C for 12 h and dissolving the residue in diluted (1:2) (v/v) nitric acid. The contents of K, Ca, Na, and Mg were determined in the obtained solutions by inductively coupled plasma optical emission spectrometry (ICP-OES, Perkin Elmer Optima 7300 DV) [18].
2.3. Calculations and Statistical Analysis
The uptake quantities of nitrogen (UN) and sulfur (US) were calculated based on the amount of maize biomass (aboveground parts and roots) and the elemental content in the biomass:
where
U (g pot−1) = (Y × C)/1000
Y—yield (g pot−1).
C—content element in biomass (g kg−1 DM).
Nitrogen (RN) and sulfur (RS) recovery from fertilization was calculated from Formula (2):
where
R (%) = ((U − Uo)/D) × 100%
U—nutrient uptake from the fertilized treatment (g pot−1).
Uo—nutrient uptake from the nonfertilized treatment (g pot−1).
D—nutrient dose (g pot−1).
The obtained values of individual macroelements in the aboveground parts of maize were used to calculate the following mass ratios: N:S; N:P, and the equivalent ratios: K:Ca; K:Mg; K:(Ca + Mg); (K + Na):(Ca + Mg). Equivalent ratios were calculated from Formula (3):
where
MgR = M/W
MgR—gram equivalent mass of an element.
M—molar mass of an element.
W—number of ion charges of an element.
2.4. Statistics
The experiment was carried out in three replicates. The obtained data were compiled with the use of STATISTICA 12 (StatSoft Inc., Tulsa, OK, USA). The mean values of analyzed properties were compared using Duncan’s multiple range test at p ≤ 0.05. Variations in treatments were determined by calculating the standard deviation (±S.D.).
3. Results and Discussion
Currently, changes in plant cultivation technologies are aimed at obtaining higher and better quality yields while minimizing the risk of environmental deterioration. There is a tendency in modern agriculture to reduce the number of mineral fertilizers and chemical plant protection products used. However, attempts are being made to use biostimulants (including those containing amino acids) in the cultivation technologies of many plant species [14,21,22]. From an environmental point of view, the introduction of biostimulants into plant breeding technology not only relieves stress, but also reduces the risk of nutrient dispersal by increasing their use from fertilization by plants.
The study showed a significant increase in maize biomass in the treatment with soil-applied tryptophan (NPKS + L-TRP) compared to the control and the treatment with only mineral fertilizer (NPKS) (Figure 1).
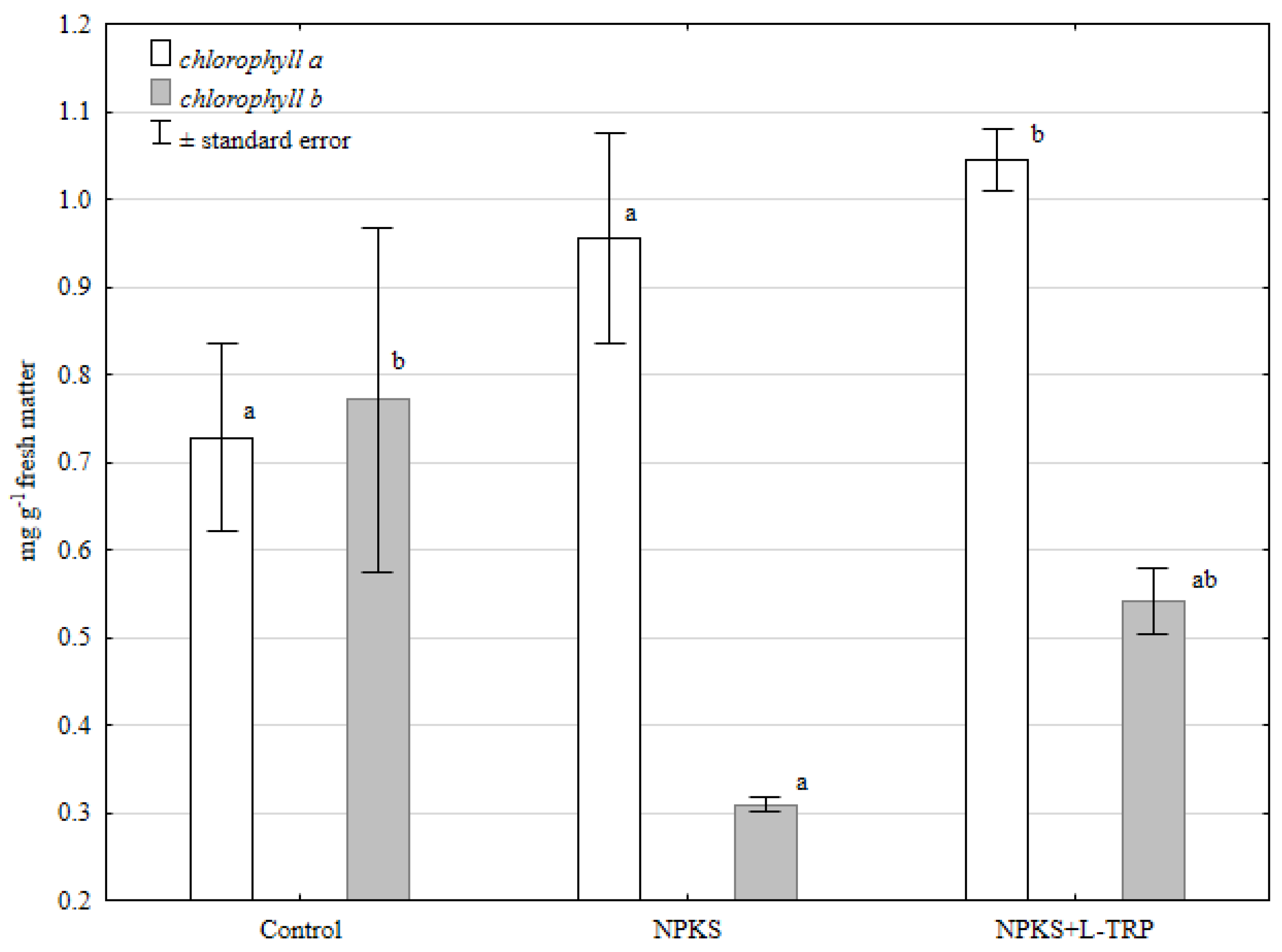
L-tryptophan is a precursor of the growth hormone, 3-indoleacetic acid, and stimulates plant growth at very low concentrations. The study of Ahmad et al. [23] revealed that the application of L-tryptophan-enriched compost to fertilize maize significantly increased the cob and grain yield compared to the mineral-fertilized treatment. Mohammandipour and Souri [24] used another amino acid, glycine, in the medium, and also showed an increase in the amount of Coriandrum sativum L. biomass. According to these authors, moderate glycine levels in the medium can improve the growth and biological value of Coriandrum sativum L. However, the authors emphasized that higher glycine concentrations in the medium (e.g., 40 mg L−1) can lower some plant growth parameters [25]. In the present study, the increase in maize biomass was probably due to the stimulatory effect of L-tryptophan and the role of this amino acid in plant metabolism. Exogenous application of amino acids can increase photosynthesis rate and Chlorophyll biosynthesis, which was also demonstrated in the present study, especially for Chlorophyll a (Figure 2). In our study, the content of Chlorophyll b in maize leaves was lower in the fertilized treatments compared to the control.
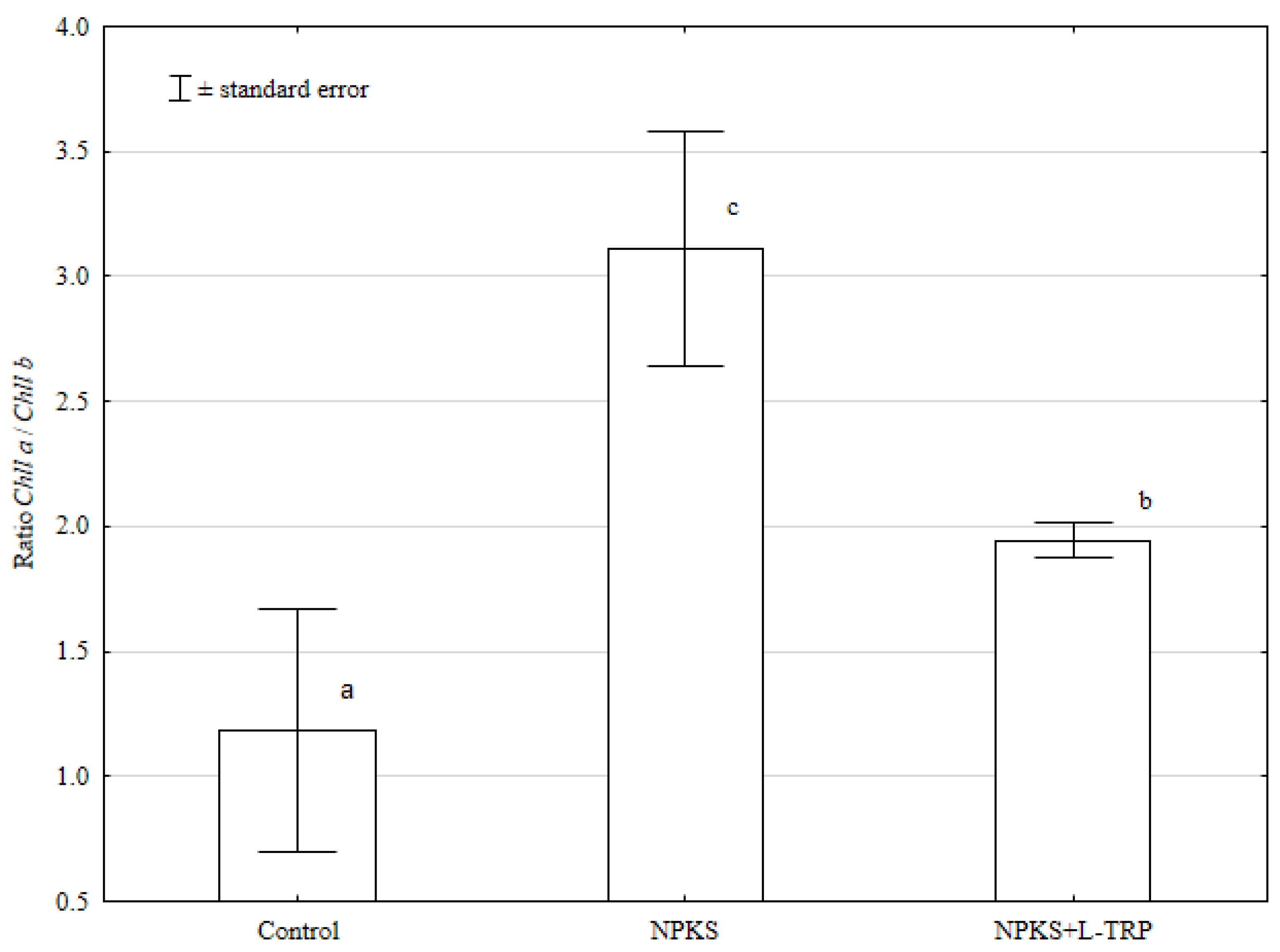
The value of the Chll a/Chll b ratio in the L-TRP-supplemented treatment was lower compared to the NPKS treatment (Figure 3).
Arshad et al. (1995) argued that the beneficial effect of L-tryptophan on plant growth can most probably be attributed to the conversion of this amino acid into auxins by the rhizosphere microflora before uptake by the plant. However, the cited authors did not rule out other mechanisms of tryptophan action, such as direct uptake of this amino acid by plant roots and subsequent auxin production in plant tissues or alteration of the rhizosphere microflora balance affecting plant growth [11]. Mustafa et al. [9] stated that L-tryptophan contains about 14% of nitrogen that is released in the rhizosphere or in the plant during metabolism, which may play a significant role in increasing crop productivity. Furthermore, nitrogen, which is a product of L-TRP metabolism, is absorbed much faster by plant cells than nitrogen from fertilizer.
Studies conducted by other authors showed that foliar or soil application of amino acids has a positive effect on nutrient uptake by plants [13,24]. In our study, the nitrogen and sulfur contents in maize aboveground parts did not differ significantly between the NPKS and the NPKS + L-TRP treatments (Table 2). Only in the case of maize roots was the sulfur content found to be significantly higher in the treatment with soil-applied tryptophan.
The calculated amounts of nitrogen and sulfur uptake by maize biomass were significantly different in individual treatments (Table 3). The significantly highest uptake amounts for both elements were found in the NPKS + L-TRP treatment. The component uptake amounts were derived from the biomass amount and the component content in the biomass. Although no significant differences in N and S content in maize biomass were found, a notable increase in the biomass amount in NPKS + L-TRP made the uptake of both elements substantially higher.
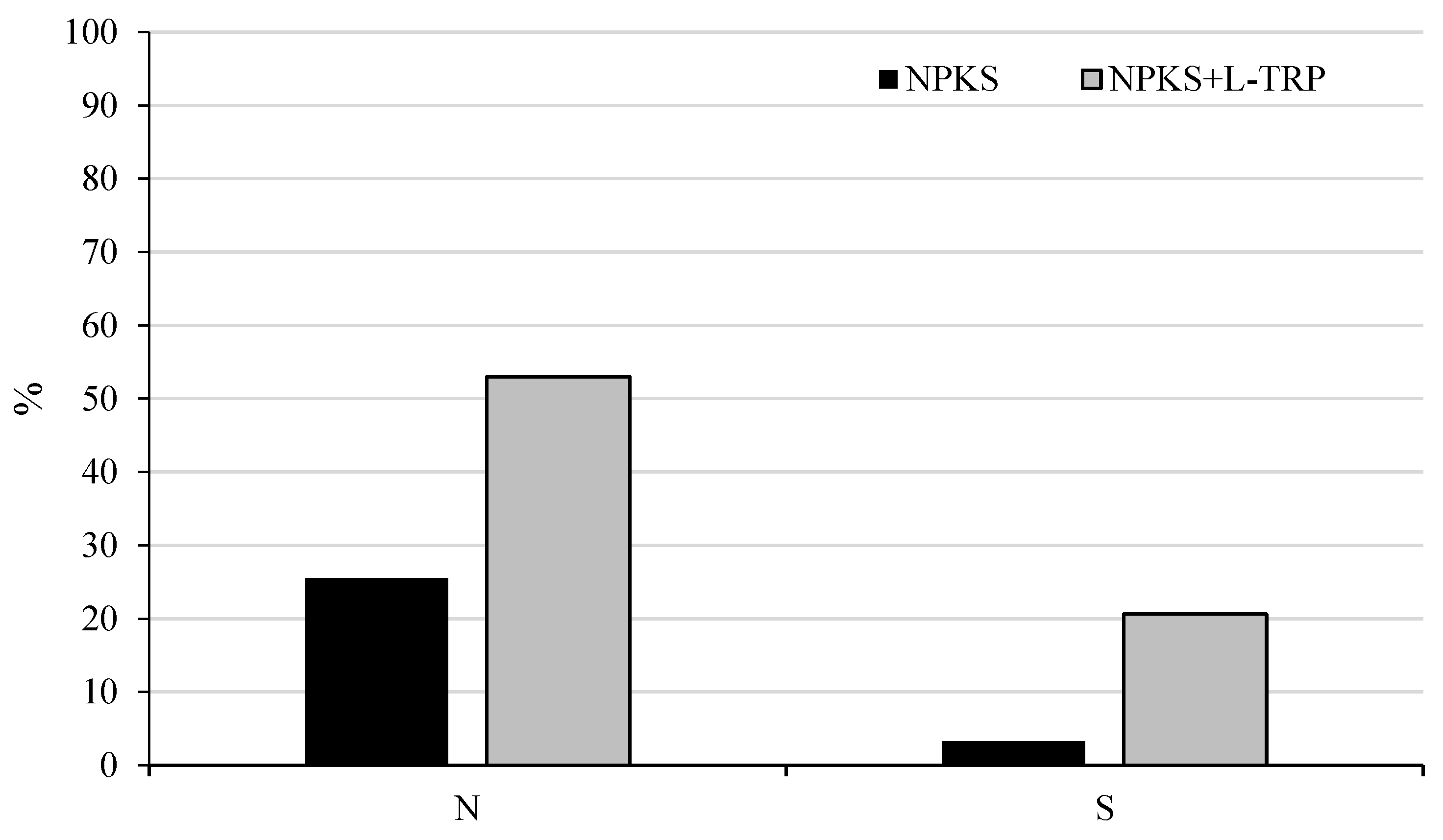
The significantly higher N and S uptakes in the tryptophan-supplemented treatment were reflected in the level of utilization of both elements from fertilization (Figure 4).
Despite the relatively short period of maize vegetation, nitrogen utilization in the NPKS + L-TRP treatment was over 27% higher than in the NPKS treatment. Sulfur utilization by maize plants in the treatment with the addition of tryptophan (NPKS + L-TRP) was over 17% higher than in the NPKS treatment. The results showed that the application of amino acids combined with mineral fertilization can, by increasing plant yield, reduce nutrient dispersion in the environment due to the better use of nutrients from fertilizers [26].
Individual cations cross semipermeable membranes at different rates. Not only the contents, but also relationships between individual elements are significant in terms of yielding and quality of the crop obtained. Maintaining the recommended contents of individual nutrients in the rhizosphere is not a guarantee of optimal quantitative and qualitative intake of nutrients. The values of the studied ratios in the aboveground parts of maize were derived from the chemical composition (Table 4).
The recorded value of the N:S mass ratio in the NPKS + L-TRP treatment did not differ from the value recorded in the aboveground parts of maize harvested in the NPKS treatment, but it should be noted that it was closer to the values considered optimal for most plant species [27]. The introduction of L-tryptophan into the soil against the NPKS fertilization significantly increased the N:P mass ratio. The values of other ratios, i.e., K:Ca, K:Mg, K:(Ca + Mg), and (K + Na):(Ca + Mg) in NPKS + L-TRP, were generally significantly lower than in NPKS. The lower values of ionic ratios in the NPKS + L-TRP treatment were indirectly a consequence of the tryptophan effect, indirectly because the addition of tryptophan significantly increased the amount of biomass and, consequently, diluted the mineral composition of the maize aboveground parts. This was also confirmed by the notably lower ash content in the aboveground parts of plants from the treatment amended with both tryptophan and NPKS fertilization (Table 2).
The use of too much calcium and magnesium in fertilization may cause a potassium deficiency in plants. No fertilization with Ca and Mg was used in the study, and the content of these elements, similarly to potassium, was a consequence of the content of their available forms in the soil. It should be noted that an excess of available magnesium may cause interference in the intake of other nutrients, e.g., K, which, in consequence, creates a competition for carriers in cell membranes. Problems with maintaining optimal relationships between individual nutrients may be compounded by soil properties. Of course, competition between monovalent and divalent ions is not the only case of antagonism between nutrients. The relationships between macronutrients and microelements are important from the plant nutrition point of view. For example, a too-high concentration of potassium may prevent the absorption of some micronutrients, such as zinc. Another issue that has to be addressed is the similarity of, e.g., Na+ and K+ ion rays, because carriers in the cell membrane can transport Na+ ions instead of K+ ions [28,29]. It seems that this is the main reason for the imbalance between Na+ and K+ ions in the plant.
4. Conclusions
The present study proved that the addition of L-TRP to the soil, supported by mineral fertilization, effectively increased plant yielding, chlorophyll biosynthesis, and utilization of nutrients. Although no significant differences in nitrogen and sulfur content in maize biomass were found between treatments without and with the addition of L-tryptophan, significantly higher intakes of both elements were observed in the NPKS + L-TRP treatment. Utilization of nitrogen and sulfur by maize in the NPKS + L-TRP treatment was more than 27% and 17% higher, respectively, compared to the NPKS treatment. Maintaining the recommended contents of individual nutrients in the rhizosphere is not a guarantee of optimal quantitative and qualitative intake of nutrients. Problems with maintaining optimal relationships between individual nutrients may be compounded by soil properties.
Author Contributions
Conceptualization, K.G. and M.M.-H.; data curation, K.G. and M.M.-H.; formal analysis, K.G.; methodology, K.G. and M.M.-H.; project administration, M.M.-H.; software, K.G.; supervision, M.M.-H.; visualization, K.G.; writing—original draft, K.G. and M.M.-H.; writing—review and editing, K.G. and M.M.-H. All authors have read and agreed to the published version of the manuscript.
Funding
The study was financed by the Ministry of Science and Higher Education of the Republic of Poland.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available from corresponding author.
Conflicts of Interest
The authors declare there are no conflict of interest.
References
- Regulation of the Minister of Agriculture and Rural Development of 9 March 2015, Item 344, on the Issue of Norms in the Scope of Good Agricultural Culture in Accordance with Environmental Protection. Available online: http://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20150000344/O/D20150344.pdf (accessed on 16 December 2021). (In Polish)
- Pecio, A. Innovative products used in plant production. Stud. Rep. IUNG 2020, 63, 163–178. (In Polish) [Google Scholar]
- Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 Laying down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repaling Regulation (EC) No 2003/2003. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32019R1009 (accessed on 16 December 2021).
- Wu, G. Amino acids: Metabolizm, functions, and nutrition. Amino Acids. 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Häusler, R.E.; Ludewig, F.; Krueger, S. Amino acids—A life between metabolism and signaling. Plant Sci. 2014, 229, 225–237. [Google Scholar] [CrossRef]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M.; Nesi, A.N.; Araujo, W.L.; Braun, H.-P. Amono acid catabolism in plants. Molecul. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Hendawy, S.F.; El-Din, A.A.E. Growth and yield of Foeniculum vulgare var. Azoricum as influenced by some vitamins and amino AIDS. Ozean. J. Appl. Sci. 2010, 3, 113–123. [Google Scholar]
- Mustafa, A.; Imran, M.; Ashraf, M.; Mahmood, K. Perspectives of using L-tryptophan for improving productivity of agricultural crops: A review. Pedosphere 2018, 28, 16–34. [Google Scholar] [CrossRef]
- Hassan, T.U.; Bano, A. The stimulatory effects of L-tryptophan and plant growth promoting rhizobacteria (PGPR) on soil health and physiology of wheat. J. Soil Sci. Plant Nutr. 2015, 15, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, A.; Hussain, A.; Naveed, M.; Ditta, A.; Nazli, Z.E.H.; Sattar, A. Response of okra (Abelmoschus esculentus L.) to soil and foliar applied L-tryptophan. Soil Environ. 2016, 35, 76–84. [Google Scholar]
- Khan, S.; Yu, H.; Li, Q.; Gao, Y.; Sallam, B.N.; Wang, H.; Liu, P.; Jiang, W. Exogenous application of amino acids improves the growth and yield of lettuce by enhancing photosynthetic Assimilation and nutrient availability. Agronomy 2019, 9, 266. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.L.; Madrid, R.; Gimeno, V.; Rodriguez-Ortega, W.M.; Nicolas, N.; Garcia-Sanchez, F. The effects of amino acids fertilization incorporated to the nutrient solution on mineral composition and growth in tomato seedlings. Span. J. Agricult. Res. 2011, 9, 852–861. [Google Scholar] [CrossRef] [Green Version]
- Popko, M.; Michalak, I.; Wilk, R.; Gramza, M.; Chojnacka, K.; Górecki, H. Effect of the new growth biostimulants based on amino acids on yield and grain quality of winter wheat. Molecules 2018, 23, 470. [Google Scholar] [CrossRef] [Green Version]
- Mohga, K.; Hosni, A.M.; Ibrahim, A.K.; Hewidy, M. Effect of amino Acids application on the growth and quality of octopus tree (Schefflera actinophylla L.). J. Biol. Chem. Environ. Sci. 2016, 11, 283–293. [Google Scholar]
- Sarwar, M.; Frankenberger, W.T., Jr. Influence of L-tryptophan and auxins applied to the rhizosphere on the vegetative growth of Zea mays L. Plant Soil 1994, 160, 97–104. [Google Scholar] [CrossRef]
- Kravchenko, L.V.; Azarova, T.S.; Makarova, N.M.; Tikhonovich, I.A. The effect of tryptophan of plant root metabolites on the phytostimulating activity of rhizobacteria. Mikrobiology 2004, 73, 195–198. [Google Scholar] [CrossRef]
- Oleszczuk, N.; Castro, J.T.; da Silva, M.M.; Korn, M.; Welz, B.; Vale, M.G. Method development for the determination of manganese, cobalt and copper in green coffee comparing direct solid sampling electrothermal atomic absorption spectrometry and inductively coupled plasma optical emission spectrometry. Talanta 2007, 73, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of diffrent resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Ostrowska, A.; Gawliński, A.; Szczubiałka, Z. Methods of Analysis and Evaluation of Soils and Plants; Institute of Environmental Protection: Warszawa, Poland, 1991. (In Polish) [Google Scholar]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Szparaga, A.; Kocira, S.; Kocira, A.; Czerwinska, E.; Swieca, M.; Lorencowicz, E.; Kornas, R.; Koszely, M.; Oniszczuk, T. Modification of growth, yield, and the nutraceutical and antioxidative potential of soybean through the use of synthetic biostimulants. Front. Plant Sci. 2018, 9, 1401. [Google Scholar] [CrossRef]
- Ahmad, R.; Khalid, A.; Arshad, M.; Zahir, Z.A.; Mahmood, T. Effect of compost enriched with N and L-tryptophan on soil and maize. Agron. Sustain. Dev. 2008, 28, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Souri, M.K.; Yaghoubi, F.; Moghadamyar, M. Growth and quality of cucumber, tomato, and green bean plants under foliar and soil applications of an aminochelate fertilizer. Horticult. Environ. Biotechnol. 2017, 58, 530–536. [Google Scholar] [CrossRef]
- Mohammandipour, N.; Souri, M.K. Effects of different levels of glycine in the nutrient solution on the growth, nutrirnt composition, and antioxidant activity of coriander (Coriandrum sativum L.). Acta Agrobot. 2019, 72, 1759. [Google Scholar] [CrossRef]
- Hammad, S.A.R.; Ali, O.A.M. Physiological and biochemical studies on drought tolerance of wheat plants by application of amino acids and yeast extract. Ann. Agricult. Sci. 2014, 59, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Filipek-Mazur, B.; Gorczyca, O.; Tabak, M. Effect of mineral containing sulfur on the content of nitrogen, sulfur and sulfur amono acids in spring rape seeds and winter wheat grains. Fragm. Agron. 2017, 34, 33–43. [Google Scholar]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxic. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Morkunas, I.; Woźniak, A.; Van Chung, M.; Rucińska-Sobkowiak, R.; Jeandet, P. The role of heavy metals in plant response to biotic stress. Molecules 2018, 23, 2320. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Amounts of dry matter in the aboveground parts and roots of maize. ± standard deviation, n = 3; means marked with the same letters do not differ significantly according to Duncan’ s test at p ≤ 0.05; factor: fertilization.
Figure 1.
Amounts of dry matter in the aboveground parts and roots of maize. ± standard deviation, n = 3; means marked with the same letters do not differ significantly according to Duncan’ s test at p ≤ 0.05; factor: fertilization.

Figure 2.
Chlorophyll a and b contents in maize leaves. ± standard deviation, n = 3; means marked with the same letters do not differ significantly according to Duncan’ s test at p ≤ 0.05; factor: fertilization.
Figure 2.
Chlorophyll a and b contents in maize leaves. ± standard deviation, n = 3; means marked with the same letters do not differ significantly according to Duncan’ s test at p ≤ 0.05; factor: fertilization.

Figure 3.
Value of the Chlorophyll a/chlorophyll b ratio. ± standard deviation, n = 3; means marked with the same letters do not differ significantly according to Duncan’ s test at p ≤ 0.05; factor: fertilization.
Figure 3.
Value of the Chlorophyll a/chlorophyll b ratio. ± standard deviation, n = 3; means marked with the same letters do not differ significantly according to Duncan’ s test at p ≤ 0.05; factor: fertilization.

Figure 4.
Utilization of nitrogen and sulfur from fertilization.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Selected chemical and physical properties of soil (means ± standard deviation).
| Determination | Unit | Value Determination (Mean ± S.D. 1) |
|---|---|---|
| pH H2O | - | 4.87 ± 0.04 |
| pH KCl | - | 3.79 ± 0.06 |
| Electrical conductivity (EC) | µS cm−1 | 68.7 ± 9.8 |
| Total carbon | g kg−1 DM 2 | 18.1 ± 2.0 |
| Total nitrogen | g kg−1 DM | 2.08 ± 0.07 |
| Total sulfur | g kg−1 DM | 0.15 ± 0.01 |
| Total magnesium | g kg−1 DM | 3.13 ± 0.02 |
| Total potassium | g kg−1 DM | 3.08 ± 0.25 |
| Total phosphorus | g kg−1 DM | 1.01 ± 0.01 |
| Total calcium | g kg−1 DM | 1.70 ± 0.01 |
| Total sodium | g kg−1 DM | 0.74 ± 0.04 |
| Sand | g kg−1 DM | 230 |
| Silt | g kg−1 DM | 630 |
| Clay | g kg−1 DM | 140 |
1 S.D. = Standard deviation, each value represents the mean of three replicates. 2 DM—dry matter.
Table 2.
Nitrogen and sulfur contents in the biomass of Zea mays L.
| Treatments | Aboveground Parts | Roots | ||||
|---|---|---|---|---|---|---|
| Ash | N | S | Ash | N | S | |
| g kg−1 DM | g kg−1 DM | |||||
| Control | 201.6 b | 49.3 a | 9.28 b | 51.1 a | 46.4 b | 11.54 b |
| NPKS | 163.9 a | 47.3 a | 6.11 b | 74.6 b | 34.3 a | 9.26 a |
| NPKS + L-TRP | 137.5 a | 46.7 a | 6.03 b | 73.5 b | 25.6 a | 13.35 b |
The mean values marked with the same letters do not differ significantly at p ≤ 0.05 (according to Duncan’s test).
Table 3.
Uptake of nitrogen and sulfur in the biomass of Zea mays L.
| Treatments | Nitrogen | Sulfur | ||||
|---|---|---|---|---|---|---|
| Aboveground Parts | Roots | Σ | Aboveground Parts | Roots | Σ | |
| g pot−1 | ||||||
| Control | 0.038 a | 0.011 b | 0.049 a | 0.007 a | 0.003 a | 0.010 a |
| NPKS | 0.077 b | 0.007 a | 0.084 b | 0.010 b | 0.002 a | 0.012 a |
| NPKS + L-TRP | 0.111 c | 0.011 b | 0.122 c | 0.014 c | 0.006 b | 0.020 b |
The mean values marked with the same letters do not differ significantly at p ≤ 0.05 (according to Duncan’s test).
Table 4.
The value of the mass and ionic ratios in the aboveground parts of maize.
| Treatments | N:S | N:P | K:Ca | K:Mg | K:(Ca + Mg) | (K + Na): (Ca + Mg) |
|---|---|---|---|---|---|---|
| Mass Rations | Ionic Rations | |||||
| Control | 5.3 a | 3.8 a | 14.6 b | 34.4 b | 10.3 b | 10.7 b |
| NPKS | 7.8 b | 5.5 ab | 14.3 b | 34.7 b | 10.1 b | 10.0 b |
| NPKS + L-TRP | 7.8 b | 7.7 c | 11.2 a | 27.2 a | 7.9 a | 7.7 a |
The mean values marked with the same letters do not differ significantly at p ≤ 0.05 (according to Duncan’s test).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gondek, K.; Mierzwa-Hersztek, M. Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize. Agronomy 2021, 11, 2582. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122582
AMA Style
Gondek K, Mierzwa-Hersztek M. Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize. Agronomy. 2021; 11(12):2582. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122582
Chicago/Turabian StyleGondek, Krzysztof, and Monika Mierzwa-Hersztek. 2021. "Effect of Soil-Applied L-tryptophan on the Amount of Biomass and Nitrogen and Sulfur Utilization by Maize" Agronomy 11, no. 12: 2582. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122582
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.