
Identification of Candidate Genes and Genomic Regions Associated with Adult Plant Resistance to Stripe Rust in Spring Wheat

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Evaluation of Stripe Rust Resistance
2.3. Statistical Analysis of Stripe Rust Resistance
2.4. Genotyping of the Tested Genotypes Using DArT Markers
2.5. Single Marker Analysis (SMA) of Stripe Rust Adult Plant Resistance
2.6. Gene Models Underlying Each Significant DArT Marker and Their Annotations
3. Results
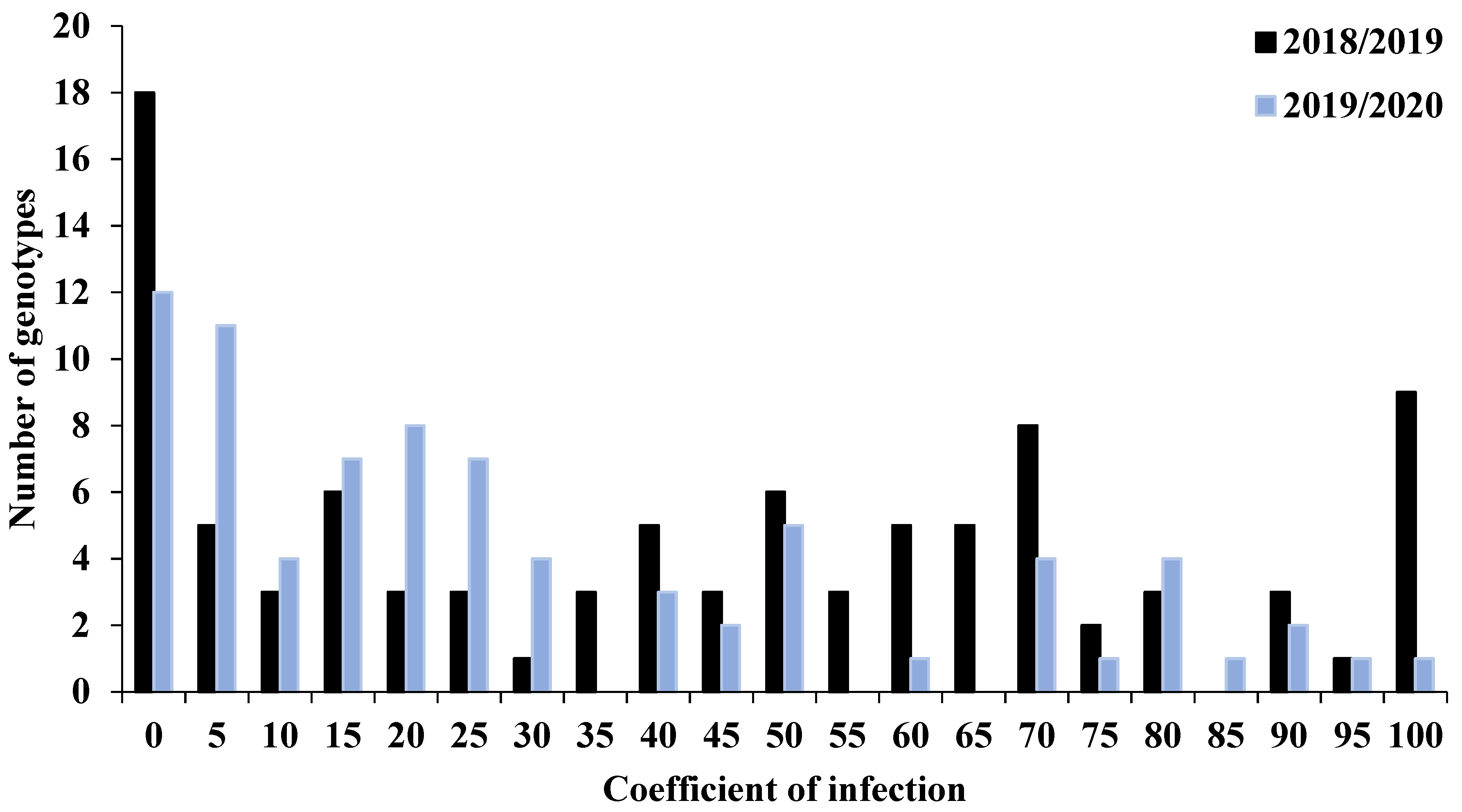
3.1. Phenotypic Variation for Stripe Rust Resistance in the Tested Genotypes
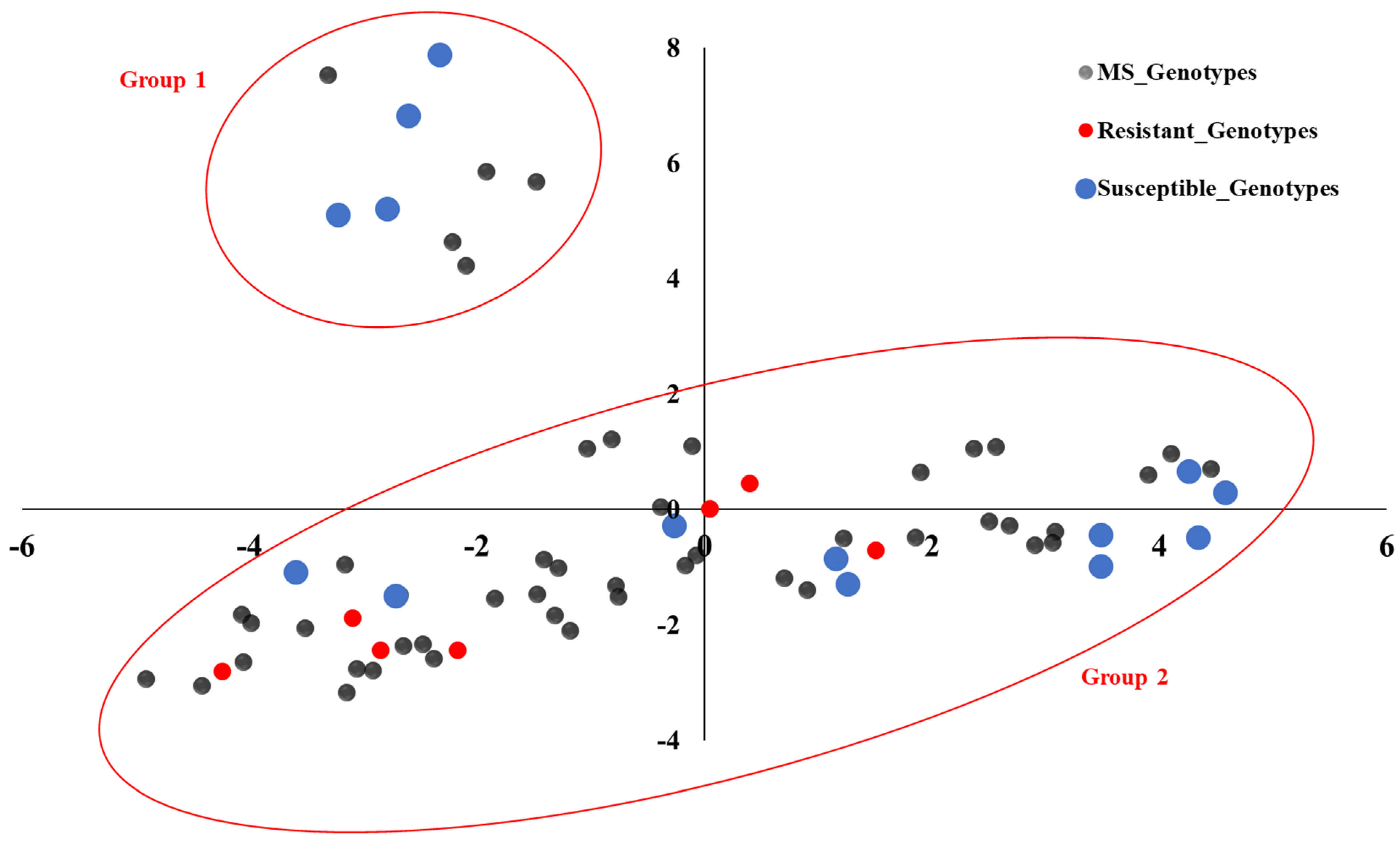
3.2. Marker Distribution and Principal Coordinate Analysis
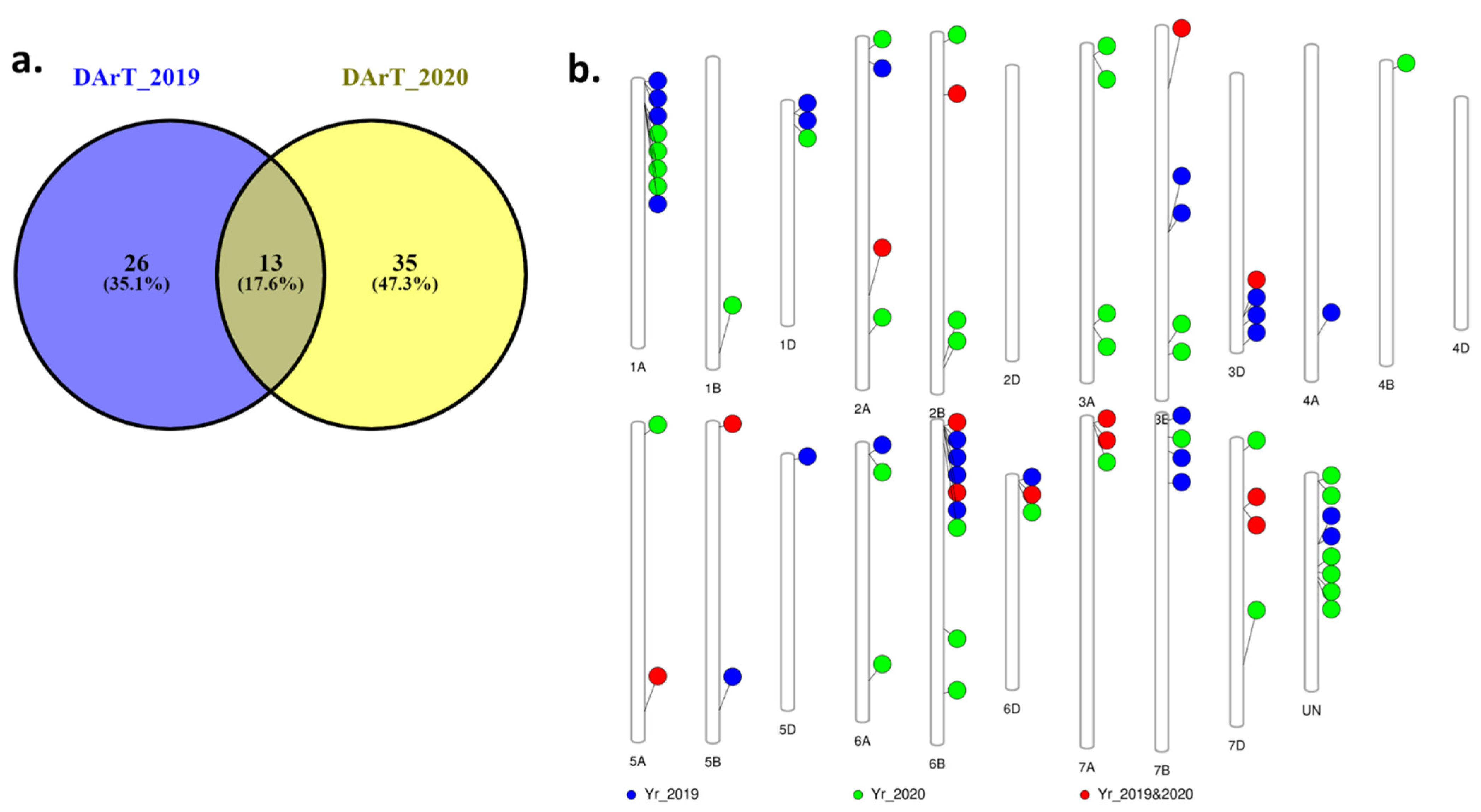
3.3. Single Marker Analysis (SMA) of Adult Plant Resistance to Stripe Rust
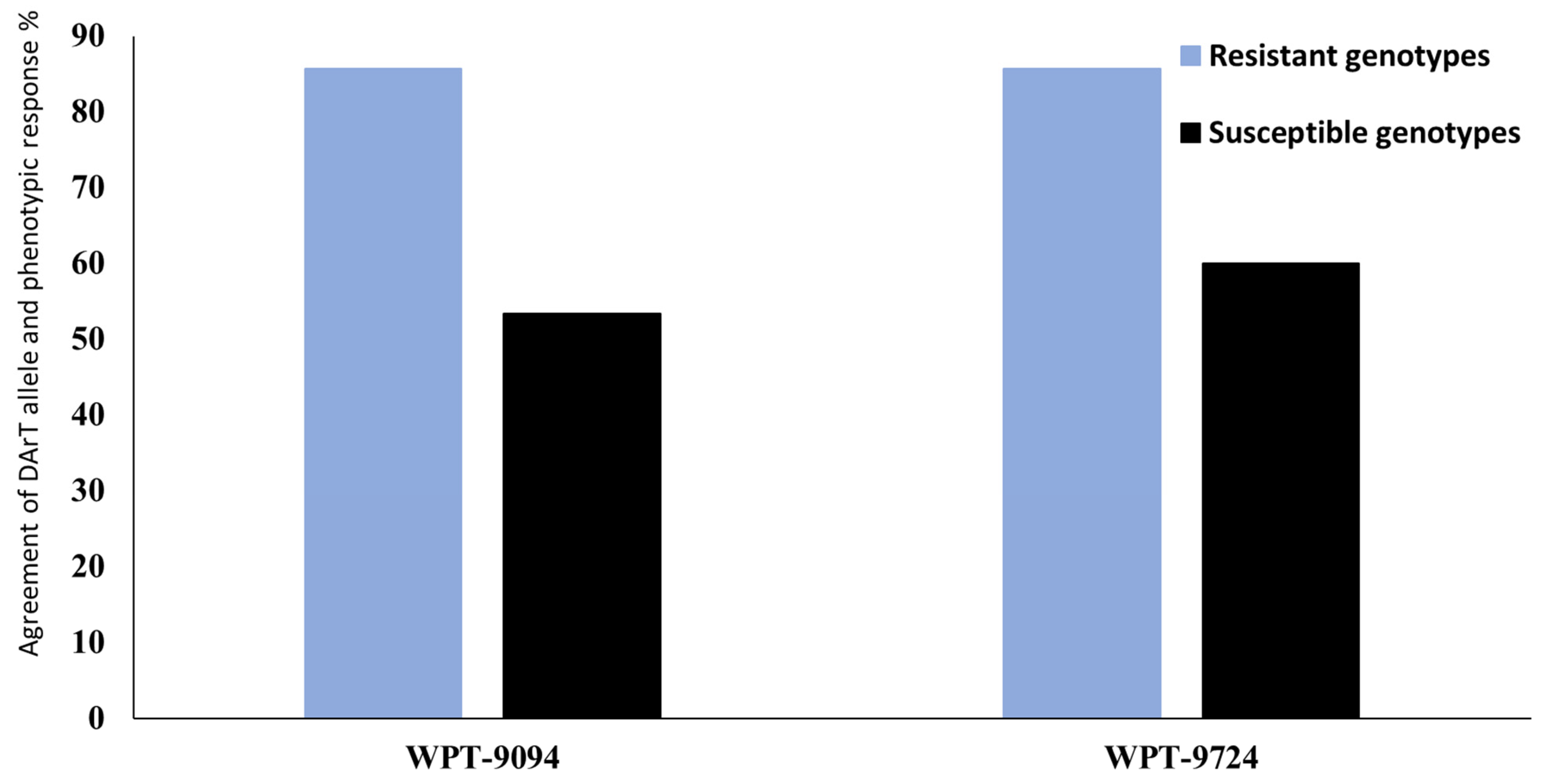
3.4. Validation of the Identified Genomic Regions Associated with Stripe Rust Resistance
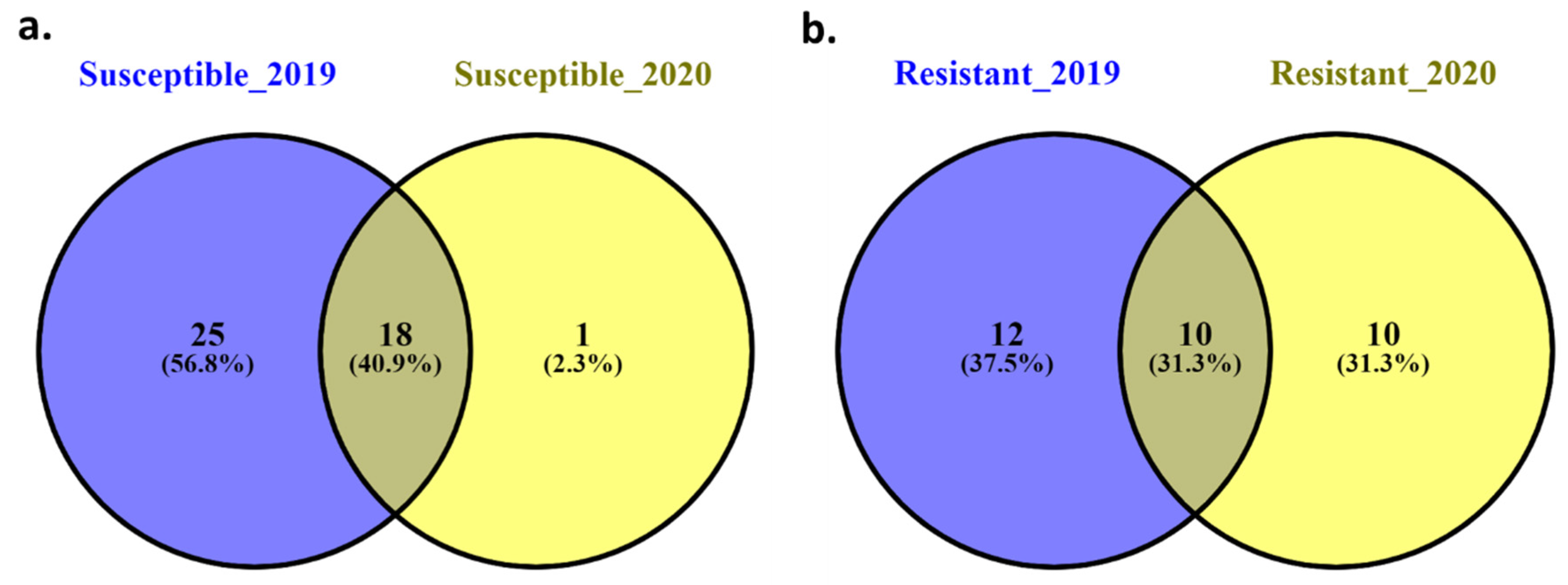
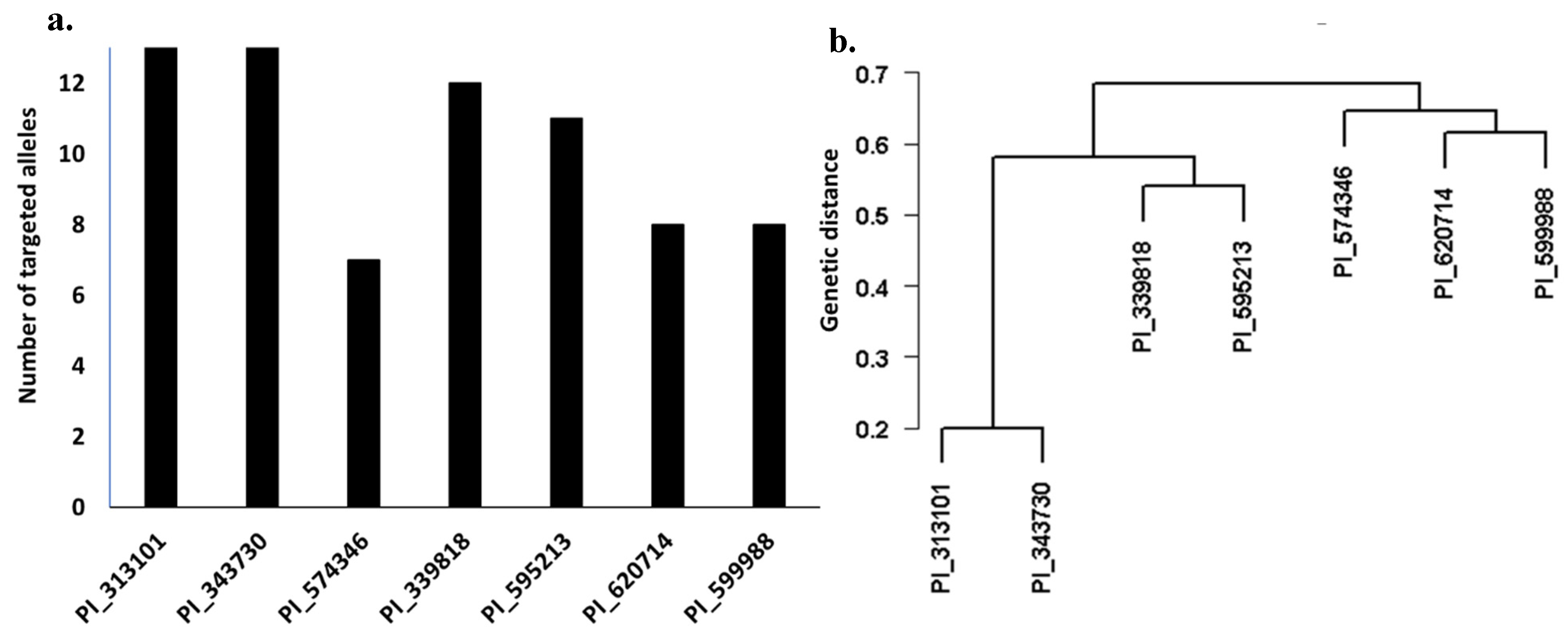
3.5. Selection of Superior Genotypes for Stripe Rust Resistance in the Studied Materials
4. Discussion
4.1. Genetic Variation of Stripe Rust Resistance in the Tested Genotypes
4.2. Genetic Analysis of Adult Plant Resistance to Stripe Rust in the Tested Genotypes
4.3. Identification of Genomic Regions Associated with Stripe Rust Adult Plant Resistance
4.4. Selection of Superior Genotypes to Stripe Rust Resistance in the Currently Studied Genotypes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Juliana, P.; Singh, R.P.; Singh, P.K.; Poland, J.A.; Bergstrom, G.C.; Huerta, J.; Bhavani, S.; Crossa, J.; Sorrells, M.E. Genome-wide association mapping for resistance to leaf rust, stripe rust and tan spot in wheat reveals potential candidate genes. Theor. Appl. Genet. 2018, 131, 1405–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of the United Nations. FAO FAOSTAT Database. 2017. Available online: https://www.fao.org/home/en (accessed on 9 December 2021).
- Riaz, A.; Athiyannan, N.; Periyannan, S.K.; Afanasenko, O.; Mitrofanova, O.P.; Platz, G.J.; Aitken, E.A.B.; Snowdon, R.J.; Lagudah, E.S.; Hickey, L.T.; et al. Unlocking new alleles for leaf rust resistance in the Vavilov wheat collection. Theor. Appl. Genet. 2018, 131, 127–144. [Google Scholar] [CrossRef]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Kumar, S.; Archak, S.; Tyagi, R.K.; Kumar, J.; Vk, V.; Bhagat, I.; Meeta, M.; Bains, N.S.; Chowdhury, A.K.; Saha, B.C.; et al. Evaluation of 19,460 Wheat Accessions Conserved in the Indian National Genebank to Identify New Sources of Resistance to Rust and Spot Blotch Diseases. PLoS ONE 2016, 11, e0167702. [Google Scholar] [CrossRef]
- Chen, X.M. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Chen, X. High-temperature adult- plant resistance, key for sustainable control of stripe rust. Am. J. Plant Sci. 2013, 2013, 608–627. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.M. Integration of cultivar resistance and fungicide application for control of wheat stripe rust Symposium contribution/Contribution à un symposium Integration of cultivar resistance and fungicide application for control of wheat stripe rust. Can. J. Plant Pathol. 2014, 36, 311–326. [Google Scholar] [CrossRef]
- Markell, S.G.; Milus, E.A. Emergence of a novel population of Puccinia striiformis f. sp. tritici in eastern United States. Phytopathology 2008, 98, 632–639. [Google Scholar] [CrossRef] [Green Version]
- Elbasyoni, I.S.; El-Orabey, W.M.; Morsy, S.; Baenziger, P.S.; Al Ajlouni, Z.; Dowikat, I. Evaluation of a global spring wheat panel for stripe rust: Resistance loci validation and novel resources identification. PLoS ONE 2019, 14, e0222755. [Google Scholar] [CrossRef]
- Eltaher, S.; Baenziger, P.S.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.M.; Alqudah, A.M.; Sallam, A. GWAS revealed effect of genotype$\times$ environment interactions for grain yield of Nebraska winter wheat. BMC Genomics 2021, 22, 2. [Google Scholar] [CrossRef] [PubMed]
- Bhavani, S.; Singh, P.K.; Qureshi, N.; He, X.; Biswal, A.K.; Juliana, P.; Dababat, A.; Mourad, A.M.I. Globally Important Wheat Diseases: Status, Challenges, Breeding and Genomic Tools to Enhance Resistance Durability. In Genomic Designing for Biotic Stress Resistant Cereal Crops; Kole, C., Ed.; Springer: New Delhi, India, 2021; pp. 59–128. ISBN 9783030758790. [Google Scholar]
- Feng, J.; Wang, M.; See, D.R.; Chao, S.; Zheng, Y.; Chen, X. Characterization of novel gene Yr79 and four additional quantitative trait loci for all-stage and high-temperature adult-plant resistance to stripe rust in spring wheat PI 182103. Phytopathology 2018, 108, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Nsabiyera, V.; Bariana, H.S.; Qureshi, N.; Wong, D.; Hayden, M.J.; Bansa, U.K. Characterisation and mapping of adult plant stripe rust resistance in wheat accession Aus27284. Theor. Appl. Genet. 2018, 131, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Liu, S.; Zhang, Y.; Xie, Y.; Wang, X.; Jiao, H.; Wu, S.; Zeng, Q.; Wang, Q.; Singh, R.P.; et al. Genome-wide wheat 55K SNP-based mapping of stripe rust resistance loci in wheat cultivar shaannong 33 and their alleles frequencies in current chinese wheat cultivars and breeding lines. Plant Dis. 2021, 105, 1048–1056. [Google Scholar] [CrossRef]
- Maccaferri, M.; Zhang, J.; Bulli, P.; Abate, Z.; Chao, S.; Cantu, D.; Bossolini, E.; Chen, X.; Pumphrey, M.; Dubcovsky, J. A Genome-wide association study of resistance to stripe rust (Puccinia striiformis f. sp. tritici) in a worldwide collection of hexaploid spring wheat (Triticum aestivum L.). G3 Genes|Genomes|Genet. 2015, 5, 449–465. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Dundas, I.; Dong, C.; Li, G.; Trethowan, R.; Yang, Z.; Hoxha, S.; Zhang, P. Identification and characterization of a new stripe rust resistance gene Yr83 on rye chromosome 6R in wheat. Theor. Appl. Genet. 2020, 133, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Roland, F.L. Stripe rust of wheat and barley in North America: A retrospective historical review. Annu. Rev. Phytopathol. 2002, 40, 75–118. [Google Scholar]
- Myburg, A.A.; Griffein, A.R.; Sederoff, R.R.; Whetten, R.W. Comparative genetic linkage maps of Eucalyptus grandis, Eucalyptus globulus and their F1 hybrid based on a double pseudo-backcross mapping approach. Theor. Appl. Genet. 2003, 103, 1028–1042. [Google Scholar] [CrossRef]
- Wenzl, P.; Carling, J.; Kudrna, D.; Jaccoud, D.; Huttner, E.; Kleinhofs, A.; Kilian, A. Diversity Arrays Technology (DArT) for whole-genome profiling of barley. Proc. Natl. Acad. Sci. USA 2004, 101, 9915–9920. [Google Scholar] [CrossRef] [Green Version]
- Wittenberg, A.H.J.; Van Der Lee, T.; Cayla, C.; Kilian, A.; Visser, R.G.F.; Schouten, H.J. Validation of the high-throughput marker technology DArT using the model plant Arabidopsis thaliana. Mol. Gen. Genom. 2005, 274, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Akbari, M.; Wenzl, P.; Caig, V.; Carling, J.; Xia, L.; Yang, S.; Uszynski, G.; Mohler, V.; Lehmensiek, A.; Kuchel, H.; et al. Diversity arrays technology (DArT) for high-throughput profiling of the hexaploid wheat genome. Theor. Appl. Genet. 2006, 113, 1409–1420. [Google Scholar] [CrossRef]
- Tinker, N.A.; Kilian, A.; Wight, C.P.; Heller-, K.; Wenzl, P.; Rines, H.W.; Bjørnstad, Å.; Howarth, C.J.; Jannink, J.; Anderson, J.M.; et al. New DArT markers for oat provide enhanced map coverage and global germplasm characterization. BMC Genomics 2009, 22, 39. [Google Scholar] [CrossRef] [Green Version]
- Jighly, A.; Oyiga, B.C.; Makdis, F.; Nazari, K. Genome-wide DArT and SNP scan for QTL associated with resistance to stripe rust (Puccinia striiformis f. sp. tritici) in elite ICARDA wheat (Triticum aestivum L.) germplasm. Theor. Appl. Genet. 2015, 128, 1277–1295. [Google Scholar] [CrossRef]
- Bemister, D.H.; Semagn, K.; Iqbal, M.; Randhawa, H.; Strelkov, S.E.; Spaner, D.M.; Bemister, D.H.; Semagn, K.; Iqbal, M.; Strelkov, S.E.; et al. Mapping QTL Associated with Stripe Rust, Leaf Rust, and Leaf Spotting in a Canadian Spring Wheat Population. Crop. Sci. 2019, 59, 650–658. [Google Scholar] [CrossRef]
- Chen, T.; Tantasawat, P.A.; Wang, W.; Gao, X.; Zhang, L. Population structure of Chinese southwest wheat germplasms resistant to stripe rust and powdery mildew using the DArT-seq technique. Ciência Rural. St. Maria 2018, 48, 1–9. [Google Scholar] [CrossRef]
- Ahmed, A.A.M.; Mohamed, E.A.; Hussein, M.Y.; Sallam, A. Genomic regions associated with leaf wilting traits under drought stress in spring wheat at the seedling stage revealed by GWAS. Environ. Exp. Bot. 2021, 184, 104393. [Google Scholar] [CrossRef]
- Peterson, R.; Campbell, A.; Hannah, A. A Diagrammitic scale for estimating rust intensity on leaves and stems of cereals. Can. J. Res. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Stakman, E.C.; Stewart, D.M.; Loegering, W.Q. Identification of Physiologic Races of Puccinia Graminis Var. Tritici; USDA_ARS: Washington, DC, USA, 1962. [Google Scholar]
- Roelfs, A.P.; Singh, R.P.; Saari, E.E. Rust Diseases of Wheat: Concepts and Methods of Disease Management; CYMMIT: Texcoco, Mexico, 1992; ISBN 968612747X. [Google Scholar]
- Abou-Zeid, M.A.; Mourad, A.M.I. Genomic regions associated with stripe rust resistance against the Egyptian race revealed by genome-wide association study. BMC Plant Biol. 2021, 21, 42. [Google Scholar] [CrossRef] [PubMed]
- Shewaye, Y.; Taddesse, W. Association mapping of seedling and adult plant resistance for stripe rust resistance in spring bread wheat (Triticum aestivum L.). Adv. Crop. Sci. Technol. 2018, 6, 2–11. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2020. Available online: https://www.R-project (accessed on 9 December 2021).
- Maccaferri, M.; Ricci, A.; Salvi, S.; Milner, S.G.; Noli, E.; Martelli, P.L.; Casadio, R.; Akhunov, E.; Scalabrin, S.; Vendramin, V.; et al. A high-density, SNP-based consensus map of tetraploid wheat as a bridge to integrate durum and bread wheat genomics and breeding. Plant Biotechnol. J. 2015, 13, 648–663. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omara, R.; Abu Aly, A.A.M.; Abou-zeid, M. Characterization of partial resistance to stripe rust (Puccinia striiformis f. sp. tritici) in some Egyptian wheat cultivars. J. Plant Prot. Path. Mansoura Univ. 2018, 9, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Minaas, E.S.; Walid, M.E.O.; Reda, I.O. Seedling and adult plant resistance to leaf rust in some Egyptian wheat genotypes. Afr. J. Agric. Res. 2016, 11, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Shahin, A.A.; Omar, H.A.; El-Sayed, A.B.B. Characterization of Yr18/Lr34 partial resistance gene to yellow rust in some Egyptian wheat cultivars. Egy. J. Plant Prot. Res. 2018, 6, 1–9. [Google Scholar]
- Mourad, A.M.I.; Sallam, A.; Belamkar, V.; Wegulo, S.; Bai, G.; Mahdy, E.; Bakheit, B.; El-wafa, A.A.; Jin, Y.; Baenziger, P.S. Molecular marker dissection of stem rust resistance in Nebraska bread wheat germplasm. Sci. Rep. 2019, 9, 11694. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Sallam, A.; Belamkar, V.; Wegulo, S.; Bowden, R.; Jin, Y.; Mahdy, E.; Bakheit, B.; El-Wafaa, A.A.; Poland, J.; et al. Genome-wide association study for identification and validation of novel SNP markers for Sr6 stem rust resistance gene in bread wheat. Front. Plant Sci. 2018, 9, 380. [Google Scholar] [CrossRef] [Green Version]
- Letta, T.; Olivera, P.; Maccaferri, M.; Jin, Y.; Ammar, K.; Badebo, A.; Salvi, S.; Noli, E.; Crossa, J.; Tuberosa, R. Association mapping reveals novel stem rust resistance loci in durum wheat at the seedling stage. Plant Genome 2014, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cane, M.A.; Maccaferri, M.; Nazemi, G.; Salvi, S.; Francia, R.; Colalongo, C.; Tuberosa, R. Association mapping for root architectural traits in durum wheat seedlings as related to agronomic performance. Mol. Breed. 2014, 34, 1629–1645. [Google Scholar] [CrossRef] [Green Version]
- Marone, D.; Panio, G.; Ficco, D.B.M.; Russo, M.A.; De Vita, P.; Papa, R.; Rubiales, D.; Cattivelli, L.; Mastrangelo, A.M. Characterization of wheat DArT markers: Genetic and functional features. Mol. Gen. Genom. 2012, 287, 741–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollier, M.; Talle, V.; Laure, A.; Zoé, B.; Bihan, L.; Duerr, S.; Lemmens, M.; Goudemand, E.; Robert, O.; Louis, J.; et al. QTL mapping and successful introgression of the spring wheat-derived QTL Fhb1 for Fusarium head blight resistance in three European triticale populations. Theor. Appl. Genet. 2020, 133, 457–477. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Pratap, A.; Solanki, R.K.; Gupta, D.S.; Goyal, A.; Chaturvedi, S.K.; Nadarajan, N.; Kumar, S. Genomic resources for improving food legume crops. J. Agric. Sci. 2012, 150, 289–318. [Google Scholar] [CrossRef]
- Alqudah, A.M.; Sallam, A.; Baenziger, P.S.; Börner, A. GWAS: Fast-forwarding gene identification and characterization in temperate Cereals: Lessons from Barley–A review. J. Adv. Res. 2020, 22, 119–135. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Dvorak, J. Genome plasticicty a key factor in the success of polyploid wheat under domestication. Science 2007, 316, 1862–1866. [Google Scholar] [CrossRef] [Green Version]
- Eltaher, S.; Sallam, A.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.M.; Poland, J.; Baenziger, P.S. Genetic diversity and population structure of F3:6 Nebraska Winter wheat genotypes using genotyping-by-sequencing. Front. Genet. 2018, 9, 76. [Google Scholar] [CrossRef]
- Nielsen, N.H.; Backes, G.; Stougaard, J.; Andersen, S.U.; Jahoor, A. Genetic diversity and population structure analysis of European hexaploid bread wheat (Triticum aestivum L.) varieties. PLoS ONE 2014, 9, e94000. [Google Scholar] [CrossRef] [Green Version]
- Röder, M.S.; Korzun, V.; Wendehake, K.; Plaschke, J.; Tixier, M.H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-G.; Tsunewaki, K. Restriction Fragment Length Polymorphism(RFLP) analysis in wheat. II. Linkage maps of the RFLP sites in common wheat. Japanese J. Genet. 1991, 66, 617–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, J.; Korol, A.B.; Fahima, T.; Röder, M.S.; Ronin, Y.I.; Li, Y.C.; Nevo, E. Molecular genetic maps in wild emmer wheat, Triticum dicoccoides: Genome-wide coverage, massive negative interference, and putative quasi-linkage. Genome Res. 2000, 10, 1509–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alipour, H.; Bihamta, M.R.; Mohammadi, V.; Peyghambari, S.A.; Bai, G.; Zhang, G. Genotyping-by-sequencing (GBS) revealed molecular genetic diversity of Iranian wheat landraces and cultivars. Front. Plant Sci. 2017, 8, 1293. [Google Scholar] [CrossRef]
- Mourad, A.; Belamkar, V.; Baenziger, P.S. Molecular genetic analysis of spring wheat core collection using genetic diversity, population structure, and linkage disequilibrium. BMC Genom. 2020, 21, 434. [Google Scholar] [CrossRef]
- Allen, A.M.; Barker, G.L.A.; Berry, S.T.; Coghill, J.A.; Gwilliam, R.; Kirby, S.; Robinson, P.; Brenchley, R.C.; D’Amore, R.; Mckenzie, N.; et al. Transcript-specific, single-nucleotide polymorphism discovery and linkage analysis in hexaploid bread wheat (Triticum aestivum L.). Plant Biotechnol. J. 2011, 9, 1086–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, K.F.M.; Sallam, A. Analysis of population structure and genetic diversity of Egyptian and exotic rice (Oryza sativa L.) genotypes. C. R. Biol. 2015, 339, 1–9. [Google Scholar] [CrossRef]
- Rostoks, N.; Ramsay, L.; Mackenzie, K.; Cardle, L.; Bhat, P.R.; Roose, M.L.; Svensson, J.T.; Stein, N.; Varshney, R.K.; Marshall, D.F.; et al. Recent history of artificial outcrossing facilitates whole-genome association mapping in elite inbred crop varieties. Proc. Natl. Acad. Sci. USA 2006, 103, 18656–18661. [Google Scholar] [CrossRef] [Green Version]
- Mourad, A.M.I.; Sallam, A.; Belamkar, V.; Mahdy, E.; Bakheit, B.; El-wafaa, A.A.; Baenziger, P.S. Genetic architecture of common bunt resistance in winter wheat using genome- wide association study. BMC Plant Biol. 2018, 18, 280. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Zhu, Y.; Sun, Z.; Yu, S.; Zhuang, J.; Fan, Y. Identification and validation of quantitative trait loci for grain number in rice (Oryza sativa L.). Agronomy 2020, 10, 180. [Google Scholar] [CrossRef] [Green Version]
- Hussain, W.; Baenziger, P.S.; Belamkar, V.; Guttieri, M.J.; Jorge, P.; Easterly, A.; Sallam, A.; Poland, J. Genotyping-by-Sequencing derived high-density linkage map and its application to QTL mapping of flag leaf traits in bread wheat. Sci. Rep. 2017, 7, 16394. [Google Scholar] [CrossRef] [PubMed]
- Klarquist, E.F.; Chen, X.; Carter, A. Novel QTL for stripe rust resistance on chromosomes 4A and 6B in soft white winter wheat cultivars. Agronomy 2016, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Lara, E.P. Mapping of Genomic Regions Associated with Agronomic Traits and Resistance to Diseases in Canadian Spring Wheat. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 2017. [Google Scholar]
- Randhawa, M.; Bansal, U.; Valárik, M.; Klocova, B.; Dolezel, J.; Bariana, H. Molecular mapping of stripe rust resistance gene Yr51 in chromosome 4AL of wheat. Theor. Appl. Genet. 2014, 127, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Lan, C.; Rosewarne, G.M.; Singh, R.P.; Sybil, A.; Huerta-Espino, J.; Basent, B.R.; Zhang, Y.; Yang, E. QTL characterization of resistance to leaf rust and stripe rust in the spring wheat line Francolin # 1. Mol. Breed. 2014, 34, 789–803. [Google Scholar] [CrossRef]
- Rosearne, G.M.; Herrera-Foessel, S.A.; Singh, R.; Huerta-Espine, J.; Lan, C.; He, Z. Quantitative trait loci of stripe rust resistance in wheat. Theor. Appl. Genet. 2013, 126, 2427–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, B.; Gao, X.; Cao, N.; Ding, Y.; Gao, Y.; Chen, T.; Xin, Z.; Zhang, L. Genome-wide association analysis of stripe rust resistance loci in wheat accessions from southwestern China. J. Appl. Genet. 2020, 61, 37–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qureshi, N.; Bariana, H.S.; Zhang, P.; McIntosh, R.; Bansal, U.K.; Wong, D.; Hayden, M.J.; Dubcovsky, J.; Shankar, M. Genetic relationship of stripe rust resistance genes Yr34 and Yr48 in wheat and identification of linked KASP markers. Plant Dis. 2018, 102, 413–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolser, D.; Staines, D.M.; Kersey, E.; Kersey, P. Ensembl Plants: Integrating tools for visualizing, mining, and analyzing plant genomics data. In Plant Bioinformatics: Mthods and Protocols; Edwards, D., Ed.; Humana Press: New York, NY, USA, 2007; Volume 406, pp. 115–140. ISBN 9781597455350. [Google Scholar]
- Quirino, B.F.; Normanly, J.; Amasino, R.M. Diverse range of gene activity during Arabidopsis thaliana leaf senescence includes pathogen-independent induction of defense-related genes. Plant Mol. Biol. 1999, 40, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Chekanova, J.A.; Shaw, R.J.; Wills, M.A.; Belostotsky, D.A. Poly(A) tail-dependent exonuclease AtRrp41p from Arabidopsis thaliana rescues 5.8 S rRNA processing and mRNA decay defects of the yeast ski6 mutant and is found in an exosome-sized complex in plant and yeast cells. J. Biol. Chem. 2000, 275, 33158–33166. [Google Scholar] [CrossRef] [Green Version]
- Stirpe, F.; Battelli, M.G. Ribosome-inactivating proteins: Progress and problems. Cell. Mol. life Sci. 2006, 63, 1850–1866. [Google Scholar] [CrossRef]
- Xi, L. A Negative Regulator of Ribosomal RNA Processing Specifies R-Gene-Independent Cell Death in Barley-Powdery Mildew Interactions. Diploma Thesis, Iowa State University, Ames, IA, USA, 2008. [Google Scholar]
- Rosa, S.B.; Zanella, C.M.; Hiebert, C.W.; Brûlé-Babel, A.L.; Randhawa, H.S.; Shorter, S.; Boyd, L.A.; McCallum, B.D. Genetic characterization of leaf and stripe rust resistance in the Brazilian wheat cultivar toropi. Phytopathology 2019, 109, 1760–1768. [Google Scholar] [CrossRef] [Green Version]
- Gullner, G.; Komives, T.; Király, L.; Schröder, P. Glutathione S-Transferase enzymes in plant-pathogen interactions. Font. Plant Sci. 2018, 9, 1836. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Knox, R.E.; DePauw, R.M.; Singh, A.K.; Cuthbert, R.D.; Campbell, H.L.; Shorter, S.; Bhavani, S. Stripe rust and leaf rust resistance QTL mapping, epistatic interactions, and co-localization with stem rust resistance loci in spring wheat evaluated over three continents. Theor. Appl. Genet. 2014, 127, 2465–2477. [Google Scholar] [CrossRef]
- Singh, A.; Knox, R.E.; DePauw, R.M.; Singh, A.K.; Cuthbert, R.D.; Campbell, H.L.; Singh, D.; Bhavani, S.; Fetch, T.; Clarke, F. Identification and mapping in spring wheat of genetic factors controlling stem rust resistance and the study of their epistatic interactions across multiple environments. Theor. Appl. Genet. 2013, 126, 1951–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertan, I.; De Carvalho, F.I.F.; Oliveira, A.C. De Parental selection strategies in plant breeding programs. J. Crop Sci. Biotechnol. 2007, 10, 211–222. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | d.f | M.S. |
|---|---|---|
| Years (Y) | 1 | 27122.56 ** |
| Replications (R) | 2 | 1839.41 |
| Genotypes (G) | 86 | 3799.96 ** |
| GY | 85 | 1402.18 ** |
| GYR | 289 | 941.92 |
| Heritability | 63.10 | |
| DArT Marker | Chromosome | p-Value | Allele Effect (%) | R2 (%) | |||
|---|---|---|---|---|---|---|---|
| 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | ||
| WPT-3976 | 2A | 0.015 | 0.044 | −24.36 | −19.21 | 11.55 | 9.80 |
| WPT-5736 | 2B | 0.037 | 0.048 | 1.66 | 13.89 | 4.55 | 0.09 |
| WPT-9088 | 3B | 0.000 | 0.002 | −36.09 | −27.63 | 17.21 | 15.00 |
| WPT-0485 | 3D | 0.005 | 0.027 | −23.89 | −20.14 | 10.50 | 10.24 |
| WPT-9094 | 5A | 0.010 | 0.004 | 19.02 | 24.98 | 7.58 | 16.81 |
| WPT-9724 | 5B | 0.030 | 0.046 | −16.66 | −13.81 | 6.38 | 5.90 |
| WPT-9256 | 6B | 0.033 | 0.001 | −17.84 | −26.54 | 6.95 | 19.42 |
| WPT-9589 | 0.001 | 0.000 | 30.85 | 32.14 | 13.38 | 20.30 | |
| RPT-7068 | 6D | 0.016 | 0.013 | 20.38 | 22.04 | 8.37 | 13.42 |
| WPT-4835 | 7A | 0.009 | 0.023 | −24.21 | −16.99 | 13.96 | 9.79 |
| WPT-6447 | 0.002 | 0.002 | −25.38 | −24.05 | 14.96 | 17.55 | |
| WPT-743380 | 7D | 0.009 | 0.015 | 19.10 | 13.37 | 4.38 | 10.98 |
| WPT-744976 | 0.008 | 0.018 | 15.26 | 19.11 | 5.59 | 10.71 | |
| Marker | Chro. | Marker Length (bp) | Blast Length (bp) | Blast Position | ID% | p-Value | Gene Model | Gene Position (bp) | Gene Annotation |
|---|---|---|---|---|---|---|---|---|---|
| WPT-5736 | 2B | 494 | 34 | 798127191-798127269 | 97.10% | 6.1 × 10−6 | TraesCS2B02G625200 | 798128178-798132084 | UniProtKB-W5B9H0 |
| TraesCS2B02G625300 | 798134799-798139436 | E3 ubiquitin-protein ligase | |||||||
| WPT-9088 | 3B | 518 | 510 | 47651816-47652325 | 100% | 0 | NA | NA | NA |
| WPT-0485 | 3D | 376 | 376 | 605639199-605639574 | 98.9 | 0 | NA | NA | NA |
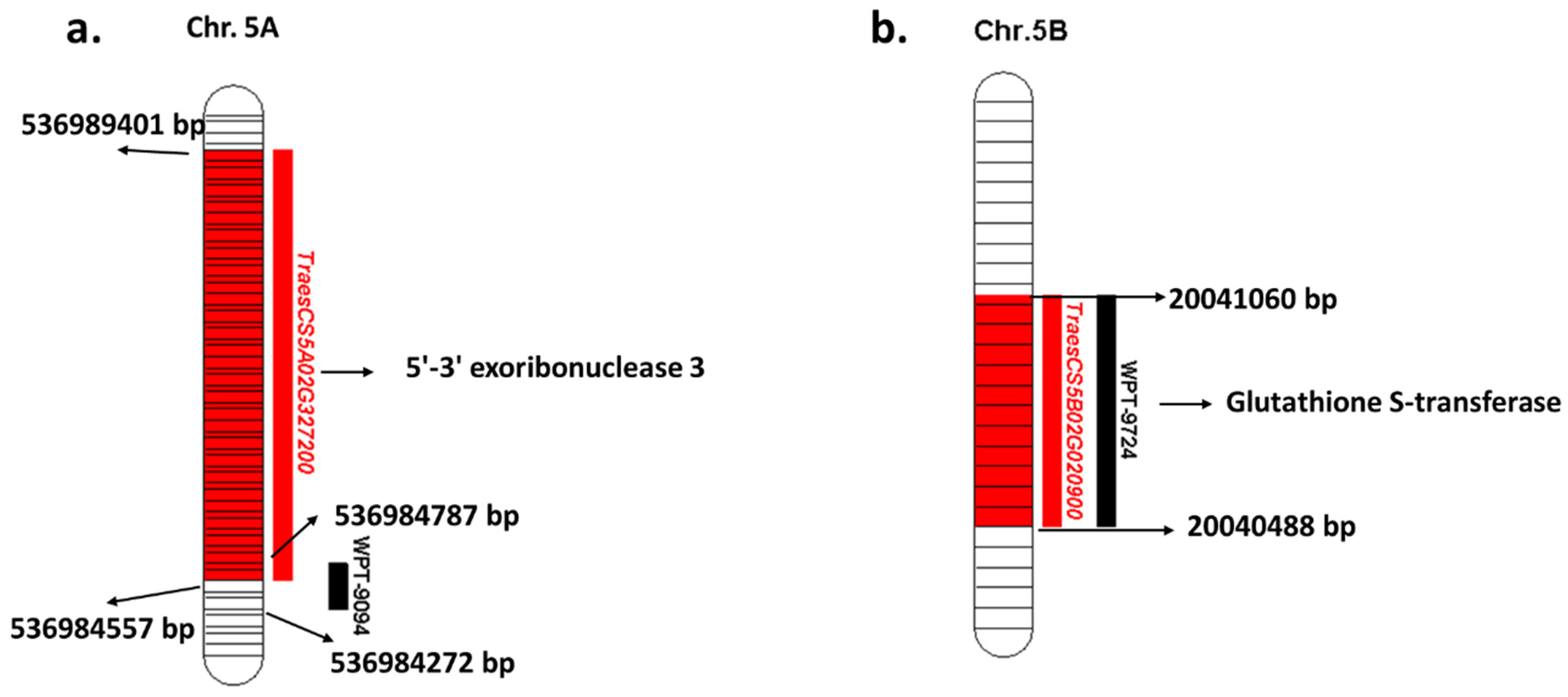
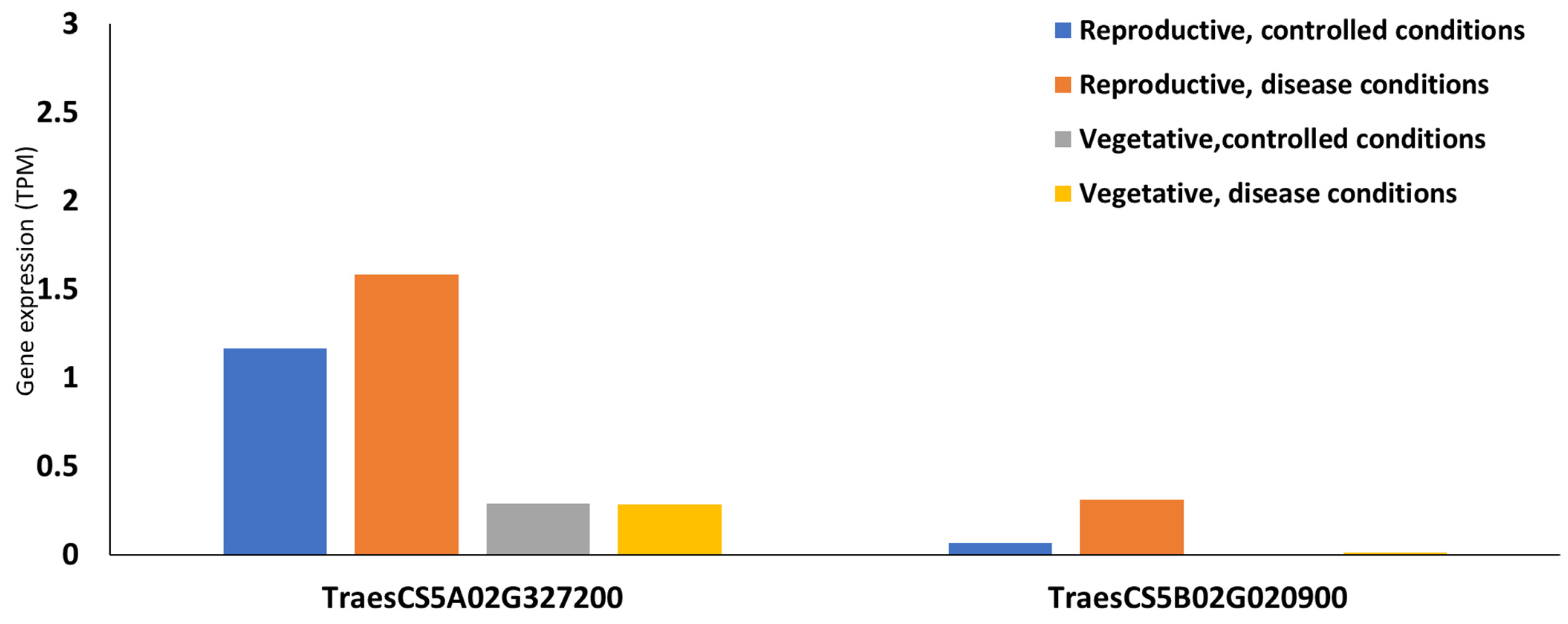
| WPT-9094 | 5A | 518 | 516 | 536984272-536984787 | 98.40% | 0 | TraesCS5A02G327200 | 536984557-536989401 | 5′-3′ exoribonuclease 3 |
| WPT-9724 | 5B | 573 | 573 | 20040488-20041060 | 99.8% | 0 | TraesCS5B02G020900 | 20040488-20041060 | Glutathione S-transferase |
| WPT-9256 | 6B | 748 | 143 | 711392100-711392242 | 97.2 | 1.20 × 10−63 | TraesCS6B02G452700 | 711372963-711375917 | Hepatocellular carcinoma-associated antigen 59 family protein, expressed |
| WPT-9589 | 572 | 101 | 715201767-715201867 | 96 | 1.1 × 10−38 | NA | NA | NA | |
| WPT-6447 | 7A | 568 | 120 | 691517984-691518114 | 85.5 | 5.30 × 10−19 | TraesCS7A02G502500 | 691521202-691522644 | Eukaryotic aspartyl protease family protein |
| WPT-4835 | 574 | 120 | 691517990-691518109 | 86.7 | 2.2 × 10−21 | NA | NA | NA | |
| WPT-743380 | 7D | 869 | 791 | 8220754-8221544 | 99.9 | 0 | TraesCS7D02G018400 | 8223605-8224570 | NBS-LRR disease resistance protein-like protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mourad, A.M.I.; Abou-Zeid, M.A.; Eltaher, S.; Baenziger, P.S.; Börner, A. Identification of Candidate Genes and Genomic Regions Associated with Adult Plant Resistance to Stripe Rust in Spring Wheat. Agronomy 2021, 11, 2585. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122585
Mourad AMI, Abou-Zeid MA, Eltaher S, Baenziger PS, Börner A. Identification of Candidate Genes and Genomic Regions Associated with Adult Plant Resistance to Stripe Rust in Spring Wheat. Agronomy. 2021; 11(12):2585. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122585
Chicago/Turabian StyleMourad, Amira M. I., Mohamed A. Abou-Zeid, Shamseldeen Eltaher, P. Stephen Baenziger, and Andreas Börner. 2021. "Identification of Candidate Genes and Genomic Regions Associated with Adult Plant Resistance to Stripe Rust in Spring Wheat" Agronomy 11, no. 12: 2585. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122585