
Plant-Bacteria Interactions for the Elimination of Atmospheric Contaminants in Cities


Department of Environmental Protection, Estación Experimental del Zaidín, Consejo Superior de Investigaciones Científicas (CSIC), Calle Profesor Albareda 1, 18008 Granada, Spain
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(3), 493; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030493
Submission received: 18 January 2021
/
Revised: 18 February 2021
/
Accepted: 27 February 2021
/
Published: 6 March 2021
(This article belongs to the Special Issue Bacteria-Plant Interactions during Bioremediation)
Abstract
:One of the major health risks for humans, especially for those living in large cities, is air pollution. Air pollution consists mainly of emissions of particulate matter (PM), nitrogen oxides, sulphur dioxide, ammonia and volatile organic compounds (VOCs). The organic carbon fraction of particulate matter is a mixture of hundreds of organic compounds, such as polycyclic aromatic hydrocarbons (PAHs), or polychlorinated dibenzo-p-dioxins and dibenzofurans (PCDD/Fs), some of which are mutagenic and/or carcinogenic. Because this particulate matter represents a serious threat for human health, measures to reduce emissions and to eliminate contaminants need to be strongly reinforced, with a focus on novel biotechnologies. In this review, we will explore the possibilities that bacteria associated with plants may offer the amelioration of atmospheric contaminants in cities, and we will discuss this strategy in the context of “Green Architecture”.
1. Introduction
Around 55% of the global population (4.2 billion people) live in urban areas, and by 2050 it has been projected that 68% of the population will be urban (https://population.un.org/wup/ (accessed on 3 March 2021). The number of megacities (those with more than 10 million inhabitants) is predicted to increase from 33 to 43 by 2030. The management of urban growth in the context of sustainable development must, therefore, maximize the benefits of agglomeration whilst minimizing the potential adverse impacts. One of the major negative impacts for citizens living in large cities is air pollution. Metropolitan area emissions constitute 52% of total emissions by weight; however, they cause nearly 75% of the gross annual damage (GAD) in the USA [1]. Health impacts related to air pollution are predicted to increase worldwide, and by 2030 it is estimated that total premature deaths due to air pollution will reach 3.1 million annually (OECD Environment Outlook to 2030). The estimated cost to the National Health Service and social care of diseases associated with air pollution is estimated to be ₤5.56 billion between 2017 and 2025 in the UK [2]. In addition to the health problem, the dispersion and deposition of particulate matter (PM) also provoke negative effects on visibility, as well as on cultural heritage and agricultural and natural ecosystems [3,4,5,6].
The major source of air pollution in big cities is on-road vehicles [7], although waste incinerators, biomass burning, secondary aerosols, oil combustion, industrial exhaust and suspended road dust are also important sources of contamination [8,9]. Air pollution consists mainly of emissions of particulate matter (PM), nitrogen oxides, sulphur dioxide (SO2), ammonia (NH3) and volatile organic compounds (VOCs). Atmospheric PMs are complex mixtures of elemental carbon (EC), organic carbon (OC), ammonium, nitrates, sulphates, mineral dust, trace elements and water. Fine particles (PM2.5), NH3, SO2 and VOCs represent almost 50% of all emissions by weight [1]. Within PM2.5, OC, consisting of a mixture of hundreds of different organic compounds, may represent 10–70% of the dry mass [10]. Therefore, OC and VOCs are some of the main elements in air pollution. Aerosol particles, in particular PM2.5, have serious adverse effects on respiratory and cardiovascular systems, and although they represent only 6% of the emissions, they cause 23% of total damage [11,12,13]. PM2.5 concentrations in many metropolitan areas exceed the limits (15 μg m−3) recommended by the USA National Ambient Air Quality Standard [14]. Some of the VOCs and OC associated with PM2.5, such as benzene, toluene, ethylbenzene, and xylenes (BTEX), polycyclic aromatic hydrocarbons (PAHs), polychlorinated biphenyls (PCBs) and polychlorinated dibenzo–p–dioxins and dibenzofurans (PCDD/Fs) [15,16], are mutagenic and/or carcinogenic.
Consequently, air pollution has a clear negative effect on the economy; it decreases labour productivity and increases the costs of public health (OECD outlook for 2030). Activities to decrease emissions are being promoted by different national and international organizations, which include taxes and tradable permits, development of eco-innovation businesses and key actions in sectors such as energy, transport and agriculture. These actions should be complemented with new strategies to ameliorate air quality.
In this review, we will focus on the removal of the main OC and VOCs in air pollution by bioabsorbing and/or biodegrading and, therefore, detoxifying associations between plants and bacteria in the context of “Green Architecture”, which is based on the inclusion of green infrastructures to make citizen-friendly, energy-saving and climate-resilient cities.
2. BTEX, PAHs, PCBs and Dioxins: Origin and Toxicity
Among the most abundant contaminants in air pollution are BTEX, PAHs, PCBs and dioxins (Figure 1). BTEX and PAHs are petroleum-derived compounds that are released into the atmosphere as emissions from motor vehicles and aircraft exhaust systems, waste incinerator smoke, fossil fuel home-heating systems, losses during petrol manufacturing, accidental oil spills in gas stations or oil refineries and cigarette smoke, amongst others [9]. However, many are natural products that can also be released during volcanic eruptions or forest fires [17,18]. PAHs and BTEX are considered carcinogenic, mutagenic and teratogenic [15,16]. Global atmospheric emissions of the 16 PAHs listed as priority pollutants by the US EPA (United States Environmental Protection Agency) were estimated to be 520 kilotons per year during the first decade of the 21st century [19,20]. PAHs are potential carcinogens; in general, as the number of aromatic rings in the molecule increases, the risks for health increase concomitantly. PAHs induce reactive oxygen species and PAH-DNA adducts that are linked to lung, skin and bladder cancers in adult populations exposed to high levels of ambient PAHs [21]. BTEX compounds are among the most abundantly produced chemicals worldwide, with an annual production of 8–10 million tons of benzene, 5–10 million tons of toluene, 5–10 million tons of ethylbenzene and 10–15 million tons of xylenes. Benzene has been classified by the International Agency for Research on Cancer (IARC) as a class 1 carcinogen for humans. It causes acute and chronic leukaemia, although the toxicity mechanisms are not well understood [22,23,24]. Toluenes, ethylbenzenes and xylenes are toxic for humans (irritating to the skin, eyes and respiratory tract, and can cause systemic toxicity by unknown mechanisms).
PCBs were banned in 1978 in the US and later on in many other countries, but prior to banning they were produced in huge amounts (megatons) as insulators and dielectric or coolant fluids for cables, electrical apparatus and heat transfer systems [25]. Trade names of specific PCB mixtures include Aroclor, Clophens, Phenoclors and Pyralenes. Chlorinated dioxins are generated as unwanted by-products in the chemical syntheses of several pesticides, disinfectants and wood preservatives [26]. A predominant source of chlorodioxin-contamination is the incineration of PCB-containing plastic insulators of electrical cables or other halogenated organic compounds. Many municipal solid waste incinerators are still emitting dioxin-contaminated fly ash. Both chlorodioxins and co-planar PCBs cause immunotoxic and endocrine effects, as well as the induction of malign tumours, whilst non-coplanar PCBs can cause neurotoxic and/or immunotoxic effects. High concentrations of chlorodioxins and also of some PCB congeners cause chloracne and anaemia. Chronic or sub-chronic exposition to low concentrations may lead to liver damage and reproductive effects, as well as cognitive deficiencies, affect levels of thyroid hormones, and may cause several types of cancer [26,27].
The toxicity of PAHs, PCBs and chlorodioxins for animals and humans is based on their capacity to bind to the aryl hydrocarbon receptor (AhR), which leads to the transcription of target genes, among them, the genes of the cytochrome P450-monooxygenases (CYP) [27,28]. CYPs are generally found in the liver and are normally responsible for the detoxification of toxic compounds taken up by the body. CYPs activate these contaminants, including BTEX, by partial oxidation, transforming the poorly water-soluble compounds into easily secreted soluble ones or organo-soluble metabolites such as phenols, dihydrodiols, quinones or epoxides [29,30,31]. The epoxide intermediates can be converted through the action of an epoxide hydrolase into highly reactive diols that are mutagenic, carcinogenic and teratogenic [30,32]. These metabolites bind to and disrupt DNA and RNA, leading to tumour formation and genotoxic effects.
Not all the contaminants have the same capacity to produce deleterious effects; for example, the binding of chlorinated dioxins and PCBs to the AhR depends on the number of chlorines and their position in the molecule. 2,3,7,8-tetrachlorodibenzo-p-dioxin (Figure 1a) is the most toxic dioxin [33]; however, in molecules that contain the chlorine atoms in other positions of the aromatic ring structure (i.e., 1,2,3,4-tetrachlorodibenzo-p-dioxin), the resulting chemical compounds are less toxic. Similarly, the non-chlorinated dioxin molecule is nearly non-toxic. In both cases, the above-mentioned molecules are not able to bind or bind poorly to the AhR-receptor. The so-called co-planar PCBs that carry chlorine substituents on their lateral position, similar to the Seveso dioxin, as shown for 3,3′,4,4′-tetrachlorobiphenyl (Figure 1a), are toxic whilst, in general, the toxicity is reduced when the four chlorines are in other positions as they no longer bind to the Ah receptor, or if they do it is to a much lesser degree [34]. Similarly, if more chlorine atoms are present than only those of the depicted co-planar PCB molecule, a lower tendency exists to form a complex with the Ah receptor, and thereby reduce the induction of detrimental effects.
3. Deposition, Transport and Detoxification of Contaminants in Plants
Although contamination by these toxic compounds can occur locally at high concentrations, for instance, by leakages of PCBs from technical equipment or by accidental oil-spills, atmospheric contaminants mainly occur from a number of processes of technical usage, dumping, burning, etc., leading to distribution by wind, sometimes over long distances, and they are finally deposited as PMs on all the surfaces of the environment [35,36]. Transportation of these contaminants in PMs is different depending on their physico-chemical properties and on meteorological factors [15,37]. The vapour-pressure of the different PAHs determines their behaviour in the atmosphere; in general, low-molecular-weight PAHs (LMW-PAHs) remain in the gas phase, whilst medium- to high-molecular-weight PAHs (HMW-PAHs) tend to bind to small particles, mainly to PM2.5. PAHs, such as fluoranthene or pyrene (both of 4 rings, Figure 1a), are equally distributed between both phases, depending on ambient air temperature and on the carbonaceous composition of the particles [38,39,40].
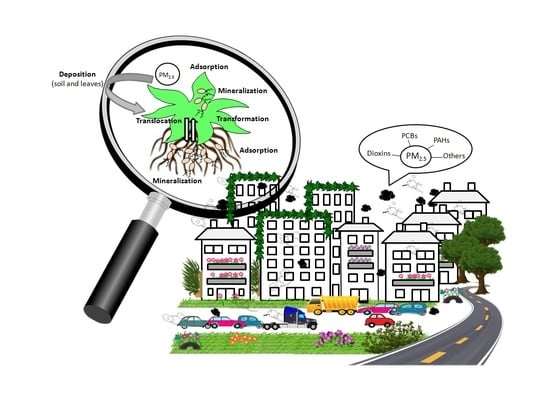
Atmospheric contaminants can be deposited directly from the air onto leaves or can be deposited in soils, and are then adsorbed to roots [41]; they can also be mobilized from soil to leaves by evaporation or wind, or be transported from roots to leaves (Figure 2) [42,43,44]. The direct relationship between PAH concentrations in soil and in plants suggests that soil-to-root transfer dominates over atmosphere-to-plant transfer [45]. Relatively volatile organic compounds, such as BTEX, anthracene and phenanthrene, are more likely to be deposited on the leaf cuticle through gaseous depositions whilst less volatile compounds, such as benzo[a]pyrene, are deposited by particulate depositions [46].
The amount of contaminants which finally accumulates in vegetation depends not only on the physico-chemical properties of the particular contaminant, but also on the characteristics of leaf surfaces and root architecture, as well as on many other environment-related parameters such as wind, rain, temperature, sorption to soils, organic content of soils, composition of root exudates and others [47,48,49].
After deposition on soils and because of the high lipophilicity and low solubility in water of PAHs, PCBs and chlorodioxins, some authors have indicated that they must be adsorbed to the epidermis of roots which are in contact with soil particles, without entering the inner root. However, there are many reports about the mobilization of contaminants from roots to shoots; i.e., for polychlorinated dibenzo-p-dioxins and dibenzofurans in Cucurbitaceae [50], total petroleum hydrocarbon in rice [51] or benzo[a]pyrene in ornamental plants [52]. Although the extent of contaminant uptake and transport from roots to shoots is still being debated in the literature, it is accepted that mobilization of contaminants through the plant is a consequence of two different processes: (i) the accumulation of contaminants in plant tissues mainly correlated to their hydrophobicity and plant lipid contents, and (ii) the transfer between plant tissues driven mainly by transpiration and the concentration gradient of the contaminant across plant-cell components [53]. Contaminant translocation is generally associated to the vascular system of the xylem with the transpiration stream acting as an important driving force, but it was recently demonstrated that PAHs are also acropetally transported via the phloem [54].
PAHs accumulate in different cells and plant tissues because of their varying solubility in the water of the tissues [55]. Although it is accepted that chemicals with logKow (logarithm of the n-octanol/water partition coefficient) between −1 and 4 are suitable for the transpiration stream, many contaminants are poorly transferred from the root to the shoot because of their high logKow values. In studies using different plants, it was demonstrated that pyrene (logKow = 5.32) accumulated in cell walls more than phenanthrene (logKow = 4.46), probably due to the higher logKow of pyrene [53,56,57]. LMW-PAHs seemed to show a stronger linear correlation between soil and shoot concentrations than HMW-PAHs, suggesting that the translocation of LMW-PAHs is faster from soil to shoots than the translocation of HMW-PAHs. Accordingly, translocation of LMW-PAHs and MMW-PAHs (pyrene, anthracene and phenanthrene) has amply been demonstrated [56], whilst there are few reports about the translocation of HMW-PAHs [55,58]. Although there is a correlation between the contaminant concentration in soil and in plants, some discrepancies have been observed between plant species.
The transport of contaminants can be symplastic (through the cell cytoplasm or vacuoles and to interconnected cells via the plasmodesmata) or apoplastic (diffusion between cell walls, without entering the cells) [59,60]. Wild et al. were pioneers in demonstrating the symplastic entrance of phenanthrene in spinach [59]. They observed that phenanthrene was mostly deposited on leaves via gas-phase transfer in maize (Zea mays L.) and spinach (Spinacia oleracea). After one or two days, phenanthrene diffused through the cuticles into the maize cell walls (indicative of apoplastic transport), being mainly located in the apoplast and moving through the cell walls and intercellular spaces of the epidermis. In spinach, they observed phenanthrene in the cellular cytoplasm (indicative of symplastic transport) and in the vacuole, with both acting as reservoirs for phenanthrene. After 12 days, most of the phenanthrene was located in the maize xylem (mostly bound to the walls), but it was not found in the spinach xylem. In soil, the symplastic uptake was also demonstrated to be the major entrance pathway of phenanthrene into wheat and of pyrene into Agropyron cristatum L. [61,62], whilst, in both cases, the apoplastic transport was also detected but at lower levels. The active absorption of contaminants involved H+ transport and energy conversion processes, and the passive transport, was associated with water led/directed through protein channels; i.e., there was a stronger correlation between the PCB concentration in soil and in carrots than in potatoes, probably because the carrots presented oil channels that could be used for PCB transport [63].
The distribution of contaminants within plants can determine their fate: to be metabolized, stored in the aerial parts of the plants or even returned (desorbed) to the atmosphere or soil. Once located in the plant interior, contaminants are mainly activated by cytochrome P450 monooxygenases (CYP). The resulting compounds are later conjugated with glucose, glucuronic acid or glutathione moieties. These conjugates are then sequestered in the cell wall or in vacuoles [64].
Despite evidence for the accumulation and subsequent biotransformation of organic contaminants in plants, it is believed that the contribution of plant uptake for their removal from the environment is very low. Soil-bound contaminants, such as PAHs, are strongly associated with soil organic matter and poorly transferred to plant roots. Furthermore, contaminants or derived products accumulating in plant cell walls or vacuoles may return to the environment after plant decay. However, plants may stimulate organic contaminant degradation through several processes such as by increasing the bioavailability of the contaminants, influencing desorption from soil particles and stimulating the biodegrading microbiota in the rhizosphere [65,66,67,68,69,70,71].
4. Degradation of BTEX, PAHs, PCBs and Dioxins by Bacteria
Biodegradation of BTEX, PAHs and, to lesser extent, PCBs is performed by a large number of different microorganisms, including bacteria, fungi and algae that have been isolated from very diverse edaphic and aquatic, aerobic and anaerobic environments, and many reviews have been published on the subject since the 1980s [72,73,74,75]. Among BTEX- and PAH-degrading bacteria, Pseudomonas, Sphingomonas, Burkholderia, Mycobacterium, Rhodococcus and Bacillus have been extensively studied. Stenotrophomonas and Pasteurella species, and some anoxic PAH-degrading bacteria belonging to the Delta-proteobacteria, have also been reported. Aerobic bacterial biodegradation of these compounds requires the presence of O2 to initiate the enzymatic attack on the aromatic rings of PAHs [76,77]. In the initial step, dioxygenase catalyses the oxidation of an aromatic ring to yield non-aromatic cis-dihydrodiols. These initial dioxygenases are multi-component enzyme systems that catalyse the dihydroxylation of PAHs ranging in size from two to five fused rings. These dihydroxylated intermediates are then, upon enzymatic rearomatization to diols, cleaved by monocomponent intradiol or extradiol ring-cleaving dioxygenases (through either an ortho-cleavage pathway or a meta-cleavage pathway). Further breakdown then leads to central intermediates such as cis,cis-muconic acids or hydroxymuconic semi-aldehydes in the case of the degradation of monoaromatic rings, or to salicylates, catechol-like structures or protocatechuates in the degradation of higher annelated aromatics. All these intermediates are then further converted to tricarboxylic acid (TCA) cycle intermediates, used for anabolic biomass formation and mineralized to CO2 (Figure 1b) [73,76]. Compounds such as naphthalene (two aromatic rings), anthracene or phenanthrene (three aromatic rings) can be completely mineralized by many of these bacteria; however, the mineralization of HMW-PAHs has not completely been elucidated, and in some bacteria, only partially degraded, dead-end products have been detected [78].
Only a single specialized bacterium, Sphingomonas wittichii strain RW1, can use the non-chlorinated dibenzo-p-dioxin molecule as its sole carbon- and energy source [79] and co-metabolize several of its chlorinated derivatives [80,81]. Other bacterial isolates, with highly similar catabolic genes to that of the Sphingomonas wittichii strain RW1, are capable of growing with diphenyl ether, dibenzofuran, carbazol or structurally similar compounds, and they can also attack the dibenzo-p-dioxin molecule, including low- to mid-chlorinated dibenzo-p-dioxins and dibenzofurans [82,83]. All these strains possess a special angular dioxygenase system [84,85].
For the biodegradation of other haloorganic contaminants (e.g., polyhalogenated benzenes or toluenes) of the ecosphere, numerous bacteria have been described, even when a single aromatic benzene ring system carries up to four chlorine atoms. A few bacteria can dehalogenize such molecules aerobically with molecular oxygen [86,87]. A general scheme for the aerobic metabolism of a monochlorobenzene is shown in Figure 1c. Fully, or almost very highly chlorinated aromatic compounds, such as hexachlorobenzene, octachlorodibenzo-p-dioxin, octachlorodibenzofuran or decachlorobiphenyl are highly recalcitrant, and only a few aerobic bacterium have the potential to degrade hexachlorobenzene [88,89]. However, such completely or almost fully halogenated aromatics can be degraded anaerobically, under sequential dehalogenation, to the mono- or even non-halogenated aromatic or alkene structure. In this case, these compounds are being utilized by special anaerobic bacteria as electron acceptors in the presence of hydrogen, thereby releasing a benign chloride ion in every single reaction step [90,91,92]. Figure 1c shows such a simplified reaction sequence, where the model compound hexachlorobenzene is dehalogenated anoxically and the resulting benzene is then aerobically mineralized.
5. Plant-Bacteria Associations for the Elimination of Atmospheric Contaminants
Whilst bacteria are armoured with a battery of degradative genes encoding catabolic biocatalysts, plants offer a large surface area to collect air particles, and they are able to stimulate bacterial activities in their rhizosphere [93]. Therefore, the combination of both organisms could be a good solution for the elimination of contaminants [94]. Radwan et al. were pioneers in demonstrating the elimination of oil contaminants by a combination of plant and soil microorganisms under natural conditions [95]. However, this natural remediation, in general, takes a long time and, for this reason, there is a pressing need to engineer more efficient bio-systems for the elimination of contaminants.
5.1. Rhizoremediation
Rhizoremediation consists of the elimination of contaminants from the soil surrounding the plant root and is based on the fact that plant tissues, especially roots, secrete different metabolites that microbes can use as a nitrogen, carbon, sulphur or phosphorus source, and on the influence that root exudates exert over the bioavailability of contaminants. Bacteria which live near root surfaces may be attracted by these root exudates [96,97] and use such rhizospheric compounds as nutrients, thereby resisting the toxicity of certain secondary metabolites secreted by plants and the presence of reactive oxygen species produced by the respiration of root cells [98,99]. Plant roots provide a large surface area on which microorganisms can proliferate and reach high cell densities. Some roots can transport microorganisms through the soil to depths where aerobic microorganisms are normally unable to thrive, and provide oxygen for the initial attack of mono- and dioxygenases for (aromatic) contaminant degradation. Although many microorganisms exhibited specific biodegradation properties, their reintroduction into the soil to remove pollutants has sometimes been unsuccessful. It is thought that nutrient limitation, competition with indigenous microbiota or poor performance under environmental stressful conditions may limit the success of (re)introduced microorganisms in contaminated soils [100]. However, in the presence of contaminants, these microorganisms have a crucial advantage over indigenous microbiota; in general contaminant-degrading microorganisms are adapted to the toxic effects of the contaminant and they can use it as an additional carbon and energy source. There are, therefore, two crucial aspects in rhizoremediation: (i) the pathways for the degradation of contaminants have to be operative and free of catabolite repression effects [101,102,103,104,105], and (ii) the contaminant has to be bioavailable in a form that can be taken up by the bacterial cell [67,68,69,71].
The elimination of contaminants by bacteria decreases their concentration in the rhizosphere and, therefore, improves plant growth [106]. In addition, some rhizospheric bacteria can solubilize phosphate, facilitate iron uptake and/or synthetize plant hormones to promote plant growth. These two actions promote soil fertility during rhizoremediation [107]. Therefore, a mutual benefit for both organisms exists during the association of plants and bacteria for the elimination of contaminants (Figure 3). The identification of plant microbiomes has confirmed that microbial populations in the rhizosphere depend on the age of the plant, its physiological status and even the cultivar, supporting the idea that plant roots exert a certain selection toward the microorganisms living in their rhizosphere [108,109,110]. It has been demonstrated that plants can attract specific degradative bacteria toward their rhizosphere in the context of rhizoremediation [111,112,113]. It is becoming clear that plant-microbe interactions influence the effectiveness of rhizoremediation and, therefore, the selection of the best plant-bacteria combination for a specific contaminant is crucial for its effective elimination [114].
Prairie grasses, which have an extensive fibrous root system, are among the best studied plants for the remediation of hydrocarbon-polluted soils [115,116]. Although well-known bacteria could be used in rhizoremediation, in general they are not well-adapted to survive and compete in the rhizosphere. Bacteria isolated from contaminated soils have also been used for bioaugmentation, although there is, in general, a lack of information about the degradative abilities of these bacteria and about their ability to persist in specific plant environments [117,118]. Certain types of degradative bacteria that are commonly found in contaminated environments have been reported to be good candidates for rhizoremediation, amongst them Arthrobacter, Rhodobacter, Rhodococcus, Pseudomonas, Burkholderia, Bacillus, Mycobacterium or members of the Sphingomonadacea family [106,119,120,121]. Certain endophytes have also been reported as good choices for bioremediation as they are adapted to a specific plant and have fewer competitors in their natural environment. However, in this case, the transport of contaminants into the plant interior is crucial for effective bioremediation [122]. In the last five years, there have been many publications dealing with different aspects of rhizoremediation, suggesting the maturity of this research field [62,123,124,125,126,127,128,129,130,131]. As a result, a large number of different bacteria which are able to remove pollutants from soil have been described; Table 1 shows some examples of bacteria-plant combinations for the degradation of contaminants.
5.2. Phylloremediation
For the elimination of atmospheric contaminants, it is noteworthy that the surface area of leaves can be 6–14 times greater than the land the vegetation is growing upon and, therefore, the phyllosphere, which is the aerial part of plants consisting of the stems and leaves, is an important environment to study [164]. The phyllosphere is a hostile habitat for microorganisms; the microbial communities established on the phyllosphere are affected by environmental factors such as UV radiation, temperature variations, climatic factors (e.g., precipitation), low nutrients and pollution [165,166,167,168,169]. Generally, the most important factors determining the structures of phyllosphere bacterial communities are the season, the plant species and biogeography [166,170,171,172,173,174,175]. One of the factors by which plant species affect phyllosphere bacterial communities is the composition and type of waxy cuticle which covers the leaf surfaces. This waxy cuticle can be comprised of lipid components, cutin and waxes, but also contains hydrocarbons and amino acids [176]. However, the limited diffusion of nutrients and water, as well as the spatially heterogeneous nutrient availability, result in leaf surfaces being largely oligotrophic with localized “oases” of nutrients [166]. Nevertheless, leaves can support bacterial populations of 104–105 bacteria mm−2 of leaf surface or up to 108 bacteria g−1 leaf material together with smaller fungal populations [177]. Bacterial communities established on leaf surfaces of different plant species are typically dominated by Proteobacteria, especially Alphaproteobacteria such as Sphingomonas and Methylobacterium [178,179], and Gammaproteobacteria such as Pseudomonas [180,181].
The accumulation of air pollutants and airborne PMs on leaf surfaces is dependent on the plant species, leaf size and structure, but is also affected by the types of waxes which make up the cuticle, the hairs covering the leaf and leaf smoothness [46,47,182,183]. Both PMs and hydrocarbon contaminants accumulating on the leaf surface affect bacterial communities. For instance, the analyses of the microbial communities on the leaves of Platanus trees revealed that tree location (city vs. rural) explained the largest differences, and that the amounts of PM on the leaves acted as partial drivers of the bacterial community structure [184]. In another study, more particulate matter (PM) was found on hornbeam tree leaves in cities, but when the authors looked at the functional diversity of hydrocarbon degradation genes of the phyllosphere communities, they found statistically more of these type of genes in the phyllosphere of a natural forest [185]. Therefore, the response of bacterial communities to PM and hydrocarbon contaminant deposition may not always be linked. Nevertheless, in a study by Franzetti et al., in which they observed seasonal differences in PAH concentrations in Milan (with maxima in winter and minima in summer), the gene coverage of dioxygenases for the catabolism of PAHs and aromatic compounds found in metagenomic data of communities of the phyllosphere of magnolia was higher in winter than in summer [186]. Similarly, the diversity of bacterial communities and of the abundance of a gene encoding hydrocarbon degradation by the phyllospheric bacteria on the leaves of Platanus in different parts of Milan was affected more strongly by the season than by the sampling location (park/road) [187].
Other studies demonstrated that phyllospheric bacteria participate directly in the removal of hydrocarbon-related air pollutants [84,181]. For instance, Sandhu et al. [188,189] showed that phyllosphere communities mineralized significantly more phenol than bean leaves which had been surface sterilized. Ali and collaborators found that bacterial communities on legume leaves could consume more crude oil, phenanthrene and n-octadecane compared to sterilized leaves [190]. Similarly, Sorkhoh et al. observed that the leaves of broad beans and American grass removed 80% more volatile crude oil hydrocarbons than leaves which had been surface sterilized [191]. Waight et al. found that phenanthrene sprayed on the leaves of Ixora plants disappeared faster than on surface-sterilized leaves [192]. Sangthong et al. demonstrated the role of epiphytic bacteria in xylene degradation [193]. Similarly, phyllospheric bacteria on the unsterilized leaves of ornamental plants enhanced phenanthrene removal [194]. Endophytes have also been used for the elimination of gaseous ethylbenzene [195]. An extensive list of plant associated bacteria which are able to degrade or biotransform air pollutants can be found in the review by Wei et al. [181]. All these findings show the involvement of phyllospheric bacteria in the elimination of air pollutants (Table 2).
More importantly, these studies also open up the possibility that the manipulation of phyllosphere bacterial communities by bioaugmentation with air pollutant-degrading bacteria could be used to remove contaminants from the air [198]. The finding by Scheublin et al., that a chlorophenol degrading Arthrobacter strain originating from soil showed good phyllosphere colonization and that it expresses degradation genes, suggests that even non-phyllospheric bacteria may successfully be used for phylloremediation [199].
6. Removal of Air Pollutants and the Role of Green Architecture
Growing awareness about indoor and outdoor air contamination has promoted research in the phytoremediation of this type of pollution [198,200,201]. Furthermore, PAHs, PCBs and VOCs are now considered as re-emerging contaminants [202,203]. However, many of the studies dealing with this subject do not specifically investigate the plant-bacteria interactions involved, although the role of the bacteria is usually acknowledged [204,205]. For many authors, plants are passive accumulators of contaminants, and their role as biomarkers has been thoroughly investigated [206,207]. However, as mentioned above, the inoculation of plants with the appropriate microbes improves contaminant removal and mineralization.
The green infrastructures of traditional cities, which include gardens, parks, open corridors and ornamental vegetation, provide services that benefit urban populations [208,209]. These services include the psychological well-being of urbanites by providing space for sports activities, relaxation and socialization [210,211,212], noise attenuation and flood mitigation [213], and increase the value of cultural heritage [214]. They also contribute to the control and spread of air pollution and to the microclimate regulation of the urban environment [215,216,217]. However, growing urban populations have forced planners for urban environments to find new strategies for public transport, employment and urban amenities, as well as for the creation and maintenance of green infrastructures [218,219]. The current urbanization trend toward a more compact and dense urban form leaves less space for green infrastructures [220,221,222]. Green Architecture is a relatively new concept that involves the construction of eco-friendly buildings and infrastructures to minimize the harmful effects of urbanization on the environment, including outdoor and indoor air pollution. According to these practices, the elements that have traditionally been used for aesthetic reasons are now one of the best ecological methods for sustainable edification, with vegetation being incorporated in very innovative ways (i.e., incorporation of vegetation surrounding or connecting skyscrapers, in vertical gardens or roof gardens and botanical sculptures) [223]. Due to the importance of health problems caused by atmospheric contaminants and the increase in the number of citizens living in megacities, there is a growing interest in the utilization of green elements for contaminant dissipation. Several studies have demonstrated that the surface with green-coverage is inversely correlated with the amount of PM contamination in the air, although the type of green coverage is also important [205,224,225,226].
Some architectural elements to mitigate air pollution are being developed, such as green wall biofilters [227], or green belts, established around industrial areas [228]. At the laboratory scale, many studies have shown that biofilters are capable of removing indoor VOCs (such as formaldehyde, toluene, ethylbenzene and xylene) [229,230]. Two different biofiltration systems can be installed in green infrastructures. In passive biofiltration, the simple diffussion of air to green wall components is used to eliminate the contaminants. These systems consist of the use of potted plants, biocovers, green roofs and green walls [231,232,233,234]. One of the problems of these systems is their limited capacity for remediating high levels of air pollution, in particular PM, but their main advantage is their low cost [235]. In active botanical biofilters, the transfer of polluted air through active systems improves the amount of the contaminant elimination [231].
Green-walls are already being commercially installed in many buildings as biofiltration systems to alleviate indoor pollution. However, there are mixed results about the efficiency of green infrastructures such as green roofs, green-walls and others in the elimination of outdoor atmospheric contamination, at least, as passive biofilters [236,237]. In most of these studies, PM elimination or adsorption to plant material has been studied, but there are not many reports about the elimination of the specific contaminants contained in PM.
Although the importance of microorganisms in green infrastructures is becoming more evident [186,238,239,240,241], few examples of manipulated plant-microorganism combinations have been reported in the literature, and those that have mainly deal with microbes that improve plant growth [242,243]. Phyllo- and rhizoremediation are good strategies for air pollutant elimination and, therefore, improving rhizosphere and phyllosphere bacterial communities with tailor-made bacterial consortia, which attack diverse organic contaminants, could be considered a way of improving the capacity of plants to remove these air pollutants within a Green Architecture strategy [237,244,245].
7. Research Needs
There are a number of issues that need to be solved in order to successfully implement the utilization of plant-bacteria combinations as a strategy to ameliorate air pollution in cities. First of all, although there has been a spectacular increase in the number of publications regarding the fate of contaminants in plants and the mechanisms of deposition on leaves and soil [246], more research will be necessary to improve our knowledge about the bioaccumulation and fate of contaminants in vegetation [247]. Depending on the location, the bioremediation strategy will require the utilization of endophytic, phyllospheric or rhizospheric bacteria capable of dealing with these target contaminants [248,249]. Because of the close proximity of green structures to citizens, detailed safety analyses of degrading microorganisms and studies about the accumulation of possible toxic intermediates from degradative pathways should be investigated [250]. The plant and its associated microbiome have co-evolved throughout time, establishing complex interrelationships to function almost as a single unit. The development and health of this supra-organism (holobiont) depends on the environmental conditions and the balance between its different components [251]. During bioremediation, the presence of pollutants and bioaugmentation with exogenous bacteria can affect the functioning of this holobiont [113,252]. Phenanthrene, for example, is able to induce the hypersensitive (HR) and systemic acquired responses (SAR), characterized by the overproduction of reactive oxidative species by the plant [253,254]. The study of these new interactions and how the degradation potential of contaminants could be altered by the complex signalization existing in these niches is a novel research field to be explored. Therefore, the molecular communication between the plant and its associated microbiota is crucial to ensure efficient contaminant elimination under changing environmental conditions [114]. Plants and their associated microbiota are able to produce multiple chemical signals (quorum sensing molecules, quorum quenching enzymes, VOCs, hormones and other secondary metabolites) that affect the equilibrium of the holobiont [165,255,256]. Unfortunately, little is known about the effects of these complex signalling cascades on the establishment and biodegradation potential of bacteria used in phyto-/rhizo-/phylloremediation. However, a better understanding of this environmental complexity, with the aid of high throughput –omics tools, is nowadays feasible [257,258].
Up to now, most of the research related to plants used in Green Architecture has focused on the tolerance of the plants toward atmospheric contaminants [259,260]. As mentioned previously, the associations of plants with contaminant-degrading bacteria promote plant growth by alleviating the toxicity of the contaminants and are increasing the number of plant species that could be used in Green Buildings. However, how to find the best plant-bacteria combinations for each location is a topic still to be resolved to achieve the successful elimination of contaminants. Moreover, these bacteria or bacterial consortia will need to be well adapted and robust, and this might be aided by a recently described adaptation strategy for the plant environment [261]. Improved engineering, architectural and agricultural practices to implement these strategies will also be required for successful implementation of this biotechnology.
8. Conclusions
The successful utilization of plant-bacteria combinations in Green Architecture is a promising technology that will have clear economic implications and lead to less public expenditure in citizens’ healthcare, and it could also lower the costs of contamination control.
Author Contributions
Conceptualization, A.S., writing—original draft preparation, L.M., R.-M.W., P.v.D. and A.S.; writing—review and editing, A.S., R.-M.W., L.M. and P.v.D.; funding acquisition, A.S. and P.v.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Ministry of Science and Innovation (grant number BIO2017-85994-P), the Junta de Andalucía (project P12-BIO-772) and FEDER funds. We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Muller, N.Z.; Mendelsohn, R. Measuring the Damages of Air Pollution in the United States. J. Environ. Econ. Manag. 2007, 54, 1–14. [Google Scholar] [CrossRef]
- Pimpin, L.; Retat, L.; Fecht, D.; de Preux, L.; Sassi, F.; Gulliver, J.; Belloni, A.; Ferguson, B.; Corbould, E.; Jaccard, A.; et al. Estimating the Costs of Air Pollution to the National Health Service and Social Care: An Assessment and Forecast up to 2035. PLoS Med. 2018, 15, e1002602. [Google Scholar] [CrossRef] [PubMed]
- Di Turo, F.; Proietti, C.; Screpanti, A.; Fornasier, M.F.; Cionni, I.; Favero, G.; De Marco, A. Impacts of Air Pollution on Cultural Heritage Corrosion at European Level: What Has Been Achieved and What Are the Future Scenarios. Environ. Pollut. Barking Essex 1987 2016, 218, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Jo, W.-K.; Chun, H.-H. Characteristics of Atmospheric Visibility and Its Relationship with Air Pollution in Korea. J. Environ. Qual. 2014, 43, 1519–1526. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.; Huang, X.; Song, Y. An Economic Assessment of the Health Effects and Crop Yield Losses Caused by Air Pollution in Mainland China. J. Environ. Sci. China 2017, 56, 102–113. [Google Scholar] [CrossRef]
- Stevens, C.J.; Bell, J.N.B.; Brimblecombe, P.; Clark, C.M.; Dise, N.B.; Fowler, D.; Lovett, G.M.; Wolseley, P.A. The Impact of Air Pollution on Terrestrial Managed and Natural Vegetation. Philos. Transact. A Math. Phys. Eng. Sci. 2020, 378, 20190317. [Google Scholar] [CrossRef]
- Dias, D.; Amorim, J.H.; Sá, E.; Borrego, C.; Fontes, T.; Fernandes, P.; Pereira, S.R.; Bandeira, J.; Coelho, M.C.; Tchepel, O. Assessing the Importance of Transportation Activity Data for Urban Emission Inventories. Transp. Res. Part. Transp. Environ. 2018, 62, 27–35. [Google Scholar] [CrossRef]
- Zhang, X.; Murakami, T.; Wang, J.; Aikawa, M. Sources, Species and Secondary Formation of Atmospheric Aerosols and Gaseous Precursors in the Suburb of Kitakyushu, Japan. Sci. Total Environ. 2020, 143001. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.T.; Liu, M.C.; Hung, P.C.; Chang, S.H.; Chang, M.B. PAH Emissions from Coal Combustion and Waste Incineration. J. Hazard. Mater. 2016, 318, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Turpin, B.J. Peer Reviewed: Options for Characterizing Organic Particulate Matter. Environ. Sci. Technol. 1999, 33, 76A–79A. [Google Scholar] [CrossRef]
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and Health Impacts of Air Pollution: A Review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krall, J.R.; Mulholland, J.A.; Russell, A.G.; Balachandran, S.; Winquist, A.; Tolbert, P.E.; Waller, L.A.; Sarnat, S.E. Associations between Source-Specific Fine Particulate Matter and Emergency Department Visits for Respiratory Disease in Four, U.S. Cities. Environ. Health Perspect. 2017, 125, 97–103. [Google Scholar] [CrossRef]
- Robinson, D.L. Composition and Oxidative Potential of PM2.5 Pollution and Health. J. Thorac. Dis. 2017, 9, 444–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murillo, J.H.; Rojas Marin, J.F.; Roman, S.R.; Beita Guerrero, V.H.; Arias, D.S.; Ramos, A.C.; Gonzalez, B.C.; Baumgardner, D.G. Temporal and Spatial Variations in Organic and Elemental Carbon Concentrations in PM10/PM2.5 in the Metropolitan Area of Costa Rica, Central America. Atmos. Pollut. Res. 2013, 4, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Bi, C.; Chen, Y.; Zhao, Z.; Li, Q.; Zhou, Q.; Ye, Z.; Ge, X. Characteristics, Sources and Health Risks of Toxic Species (PCDD/Fs, PAHs and Heavy Metals) in PM2.5 during Fall and Winter in an Industrial Area. Chemosphere 2020, 238, 124620. [Google Scholar] [CrossRef]
- Idris, S.A.A.; Hanafiah, M.M.; Khan, M.F.; Hamid, H.H.A. Indoor Generated PM2.5 Compositions and Volatile Organic Compounds: Potential Sources and Health Risk Implications. Chemosphere 2020, 255, 126932. [Google Scholar] [CrossRef] [PubMed]
- Remizovschi, A.; Carpa, R.; Forray, F.L.; Chiriac, C.; Roba, C.-A.; Beldean-Galea, S.; Andrei, A.-Ș.; Szekeres, E.; Baricz, A.; Lupan, I.; et al. Mud Volcanoes and the Presence of PAHs. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Han, Y.; Zhao, Y.; Huang, Y.; Liu, X.; Tao, S.; Liu, J.; Huang, T.; Wang, L.; Chen, K.; et al. Effect of Northern Boreal Forest Fires on PAH Fluctuations across the Arctic. Environ. Pollut. Barking Essex 1987 2020, 261, 114186. [Google Scholar] [CrossRef]
- Samburova, V.; Zielinska, B.; Khlystov, A. Do 16 Polycyclic Aromatic Hydrocarbons Represent PAH Air Toxicity? Toxics 2017, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Huang, Y.; Wang, R.; Zhu, D.; Li, W.; Shen, G.; Wang, B.; Zhang, Y.; Chen, Y.; Lu, Y.; et al. Global Atmospheric Emissions of Polycyclic Aromatic Hydrocarbons from 1960 to 2008 and Future Predictions. Environ. Sci. Technol. 2013, 47, 6415–6424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewa, B.; Danuta, M.-Š. Polycyclic Aromatic Hydrocarbons and PAH-Related DNA Adducts. J. Appl. Genet. 2017, 58, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Fenga, C.; Gangemi, S.; Costa, C. Benzene Exposure Is Associated with Epigenetic Changes (Review). Mol. Med. Rep. 2016, 13, 3401–3405. [Google Scholar] [CrossRef] [Green Version]
- Galbraith, D.; Gross, S.A.; Paustenbach, D. Benzene and Human Health: A Historical Review and Appraisal of Associations with Various Diseases. Crit. Rev. Toxicol. 2010, 40 (Suppl. S2), 1–46. [Google Scholar] [CrossRef]
- Lin, C.-K.; Hsu, Y.-T.; Brown, K.D.; Pokharel, B.; Wei, Y.; Chen, S.-T. Residential Exposure to Petrochemical Industrial Complexes and the Risk of Leukemia: A Systematic Review and Exposure-Response Meta-Analysis. Environ. Pollut. 2020, 258, 113476. [Google Scholar] [CrossRef] [PubMed]
- Vorkamp, K. An Overlooked Environmental Issue? A Review of the Inadvertent Formation of PCB-11 and Other PCB Congeners and Their Occurrence in Consumer Products and in the Environment. Sci. Total Environ. 2016, 541, 1463–1476. [Google Scholar] [CrossRef] [PubMed]
- Eskenazi, B.; Warner, M.; Brambilla, P.; Signorini, S.; Ames, J.; Mocarelli, P. The Seveso Accident: A Look at 40 years of Health Research and Beyond. Environ. Int. 2018, 121, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Larigot, L.; Juricek, L.; Dairou, J.; Coumoul, X. AhR Signaling Pathways and Regulatory Functions. Biochim. Open 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Jayasundara, N.; Van Tiem Garner, L.; Meyer, J.N.; Erwin, K.N.; Di Giulio, R.T. AHR2-Mediated Transcriptomic Responses Underlying the Synergistic Cardiac Developmental Toxicity of PAHs. Toxicol. Sci. 2015, 143, 469–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, C.J.; Hannigan, J.H.; Bowen, S.E. Effects of Inhaled Combined Benzene, Toluene, Ethylbenzene, and Xylenes (BTEX): Toward an Environmental Exposure Model. Environ. Toxicol. Pharmacol. 2021, 81, 103518. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, B.; Chu, C.; Carlin, D.J. Polycyclic Aromatic Hydrocarbons: From Metabolism to Lung Cancer. Toxicol. Sci. Off. J. Soc. Toxicol. 2015, 145, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, T.; Fujii-Kuriyama, Y. Metabolic Activation of Polycyclic Aromatic Hydrocarbons to Carcinogens by Cytochromes P450 1A1 And1B. Cancer Sci. 2004, 95, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Warshawsky, D. Metabolic Activation of Polycyclic and Heterocyclic Aromatic Hydrocarbons and DNA Damage: A Review. Toxicol. Appl. Pharmacol. 2005, 206, 73–93. [Google Scholar] [CrossRef] [PubMed]
- Croutch, C.R.; Lebofsky, M.; Schramm, K.-W.; Terranova, P.F.; Rozman, K.K. 2,3,7,8-Tetrachlorodibenzo-p-Dioxin (TCDD) and 1,2,3,4,7,8-Hexachlorodibenzo-p-Dioxin (HxCDD) Alter Body Weight by Decreasing Insulin-Like Growth Factor I (IGF-I) Signaling. Toxicol. Sci. 2005, 85, 560–571. [Google Scholar] [CrossRef] [Green Version]
- Hashmi, M.Z.; Zhang, J.; Li, B.; Su, X.; Tariq, M.; Ahmad, N.; Malik, R.N.; Ullah, K.; Chen, C.; Shen, C. Effects of Structurally Different Noncoplanar and Coplanar PCBs on HELF Cell Proliferation, Cell Cycle, and Potential Molecular Mechanisms. Environ. Toxicol. 2017, 32, 1183–1190. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, M.; Liu, B.; Wu, J.; Peng, X.; Zhang, Y.; Han, S.; Feng, Y.; Zhu, T. Characterization and Source Apportionment of Size-Segregated Atmospheric Particulate Matter Collected at Ground Level and from the Urban Canopy in Tianjin. Environ. Pollut. 2016, 219, 982–992. [Google Scholar] [CrossRef]
- Mei, D.; Wen, M.; Xu, X.; Zhu, Y.; Xing, F. The Influence of Wind Speed on Airflow and Fine Particle Transport within Different Building Layouts of an Industrial City. J. Air Waste Manag. Assoc. 1995 2018, 68, 1038–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, J.; Yang, H.; Li, R.; Yu, Q. Seasonal Variation and Potential Source Regions of PM2.5-Bound PAHs in the Megacity Beijing, China: Impact of Regional Transport. Environ. Pollut. 2017, 231, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, T.; Huang, Y.; Mao, T.; Zhong, N. Size Distribution of Polycyclic Aromatic Hydrocarbons in Urban and Suburban Sites of Beijing, China. Chemosphere 2005, 61, 792–799. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, R.; Lammel, G. Adsorptive and Absorptive Contributions to the Gas-Particle Partitioning of Polycyclic Aromatic Hydrocarbons: State of Knowledge and Recommended Parametrization for Modeling. Environ. Sci. Technol. 2004, 38, 3793–3803. [Google Scholar] [CrossRef]
- Mu, Q.; Shiraiwa, M.; Octaviani, M.; Ma, N.; Ding, A.; Su, H.; Lammel, G.; Pöschl, U.; Cheng, Y. Temperature Effect on Phase State and Reactivity Controls Atmospheric Multiphase Chemistry and Transport of PAHs. Sci. Adv. 2018, 4. [Google Scholar] [CrossRef] [Green Version]
- Pullagurala, V.L.R.; Rawat, S.; Adisa, I.O.; Hernandez-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Plant Uptake and Translocation of Contaminants of Emerging Concern in Soil. Sci. Total Environ. 2018, 636, 1585–1596. [Google Scholar] [CrossRef]
- Collins, C.D.; Finnegan, E. Modeling the Plant Uptake of Organic Chemicals, Including the Soil-Air-Plant Pathway. Environ. Sci. Technol. 2010, 44, 998–1003. [Google Scholar] [CrossRef]
- Takaki, K.; Wade, A.J.; Collins, C.D. Assessment of Plant Uptake Models Used in Exposure Assessment Tools for Soils Contaminated with Organic Pollutants. Environ. Sci. Technol. 2014, 48, 12073–12082. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, S.; Zheng, L.; Li, T.; Sun, Y.; Ma, L.; Lin, Z.; Grathwohl, P.; Lohmann, R. Air-Soil Diffusive Exchange of PAHs in an Urban Park of Shanghai Based on Polyethylene Passive Sampling: Vertical Distribution, Vegetation Influence and Diffusive Flux. Sci. Total Environ. 2019, 689, 734–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fismes, J.; Perrin-Ganier, C.; Empereur-Bissonnet, P.; Morel, J.L. Soil-to-Root Transfer and Translocation of Polycyclic Aromatic Hydrocarbons by Vegetables Grown on Industrial Contaminated Soils. J. Environ. Qual. 2002, 31, 1649–1656. [Google Scholar] [CrossRef]
- Bakker, M.I.; Koerselman, J.W.; Tolls, J.; Kollöffel, C. Localization of Deposited Polycyclic Aromatic Hydrocarbons in Leaves of Plantago. Environ. Toxicol. Chem. 2001, 20, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Dai, C.; Zhou, Y.; Peng, H.; Yi, K.; Qin, P.; Luo, S.; Zhang, X. Comparisons of Three Plant Species in Accumulating Polycyclic Aromatic Hydrocarbons (PAHs) from the Atmosphere: A Review. Environ. Sci. Pollut. Res. 2018, 25, 16548–16566. [Google Scholar] [CrossRef]
- Howsam, M.; Jones, K.C.; Ineson, P. PAHs Associated with the Leaves of Three Deciduous Tree Species. II: Uptake during a Growing Season. Chemosphere 2001, 44, 155–164. [Google Scholar] [CrossRef]
- Howsam, M.; Jones, K.C.; Ineson, P. PAHs Associated with the Leaves of Three Deciduous Tree Species. I—Concentrations and Profiles. Environ. Pollut. 2000, 108, 413–424. [Google Scholar] [CrossRef]
- Huelster, A.; Mueller, J.F.; Marschner, H. Soil-Plant Transfer of Polychlorinated Dibenzo-p-Dioxins and Dibenzofurans to Vegetables of the Cucumber Family (Cucurbitaceae). Environ. Sci. Technol. 1994, 28, 1110–1115. [Google Scholar] [CrossRef]
- Patowary, R.; Patowary, K.; Devi, A.; Kalita, M.C.; Deka, S. Uptake of Total Petroleum Hydrocarbon (TPH) and Polycyclic Aromatic Hydrocarbons (PAHs) by Oryza sativa L. Grown in Soil Contaminated with Crude Oil. Bull. Environ. Contam. Toxicol. 2017, 98, 120–126. [Google Scholar] [CrossRef]
- Sun, Y.; Zhou, Q. Uptake and Translocation of Benzo[a]Pyrene (B[a]P) in Two Ornamental Plants and Dissipation in Soil. Ecotoxicol. Environ. Saf. 2016, 124, 74–81. [Google Scholar] [CrossRef]
- Kang, F.; Chen, D.; Gao, Y.; Zhang, Y. Distribution of Polycyclic Aromatic Hydrocarbons in Subcellular Root Tissues of Ryegrass (Lolium Multiflorum Lam.). BMC Plant. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Gu, R.; Sheng, Y.; Zeng, N.; Zhan, X. Acropetal Translocation of Phenanthrene in Wheat Seedlings: Xylem or Phloem Pathway? Environ. Pollut. 2020, 260, 114055. [Google Scholar] [CrossRef] [PubMed]
- Alves, W.S.; Manoel, E.A.; Santos, N.S.; Nunes, R.O.; Domiciano, G.C.; Soares, M.R. Detection of Polycyclic Aromatic Hydrocarbons (PAHs) in Medicago sativa L. by Fluorescence Microscopy. Micron 2017, 95, 23–30. [Google Scholar] [CrossRef]
- Gao, Y.; Zhu, L. Plant Uptake, Accumulation and Translocation of Phenanthrene and Pyrene in Soils. Chemosphere 2004, 55, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Houshani, M.; Salehi-Lisar, S.Y.; Motafakkerazad, R.; Movafeghi, A. Uptake and distribution of phenanthrene and pyrene in roots and shoots of maize (Zea mays L.). Environ. Sci. Pollut. Res. 2019, 26, 9938–9944. [Google Scholar] [CrossRef] [PubMed]
- Alagić, S.Č.; Jovanović, V.P.S.; Mitić, V.D.; Cvetković, J.S.; Petrović, G.M.; Stojanović, G.S. Bioaccumulation of HMW PAHs in the Roots of Wild Blackberry from the Bor Region (Serbia): Phytoremediation and Biomonitoring Aspects. Sci. Total Environ. 2016, 562, 561–570. [Google Scholar] [CrossRef]
- Wild, E.; Dent, J.; Thomas, G.O.; Jones, K.C. Direct Observation of Organic Contaminant Uptake, Storage, and Metabolism within Plant Roots. Environ. Sci. Technol. 2005, 39, 3695–3702. [Google Scholar] [CrossRef]
- Miller, E.L.; Nason, S.L.; Karthikeyan, K.G.; Pedersen, J.A. Root Uptake of Pharmaceuticals and Personal Care Product Ingredients. Environ. Sci. Technol. 2016, 50, 525–541. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Zhu, M.; Shen, Y.; Yue, L.; Li, J.; Gardea-Torresdey, J.L.; Xu, G. Apoplastic and Symplastic Uptake of Phenanthrene in Wheat Roots. Environ. Pollut. Barking Essex 1987 2018, 233, 331–339. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, J.; Liu, X.; Zhang, Y.; Zou, Y.; Yuan, J. Study on Removal of Pyrene by Agropyron cristatum L. in Pyrene–Ni Co-Contaminated Soil. Int. J. Phytoremediation 2020, 22, 313–321. [Google Scholar] [CrossRef]
- Kipopoulou, A.M.; Manoli, E.; Samara, C. Bioconcentration of Polycyclic Aromatic Hydrocarbons in Vegetables Grown in an Industrial Area. Environ. Pollut. 1999, 106, 369–380. [Google Scholar] [CrossRef]
- Pilon-Smits, E. Phytoremediation. Annu. Rev. Plant. Biol. 2005, 56, 15–39. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Garrido, B.; Balseiro-Romero, M.; Kidd, P.S.; Monterroso, C. Effect of Plant Root Exudates on the Desorption of Hexachlorocyclohexane Isomers from Contaminated Soils. Chemosphere 2020, 241, 124920. [Google Scholar] [CrossRef]
- Jia, H.; Lu, H.; Liu, J.; Li, J.; Dai, M.; Yan, C. Effects of Root Exudates on the Leachability, Distribution, and Bioavailability of Phenanthrene and Pyrene from Mangrove Sediments. Environ. Sci. Pollut. Res. Int. 2016, 23, 5566–5576. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.; Yan, W.; Ning, C.; Gsell, T. Both Artificial Root Exudates and Natural Koelreuteria paniculata Exudates Modify Bacterial Community Structure and Enhance Phenanthrene Biodegradation in Contaminated Soils. Chemosphere 2021, 263, 128041. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Sun, J.; Zhu, L. The Role of Artificial Root Exudate Components in Facilitating the Degradation of Pyrene in Soil. Sci. Rep. 2017, 7, 7130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Ren, L.; Ling, W.; Gong, S.; Sun, B.; Zhang, Y. Desorption of Phenanthrene and Pyrene in Soils by Root Exudates. Bioresour. Technol. 2010, 101, 1159–1165. [Google Scholar] [CrossRef]
- Balasubramaniyam, A. The Influence of Plants in the Remediation of Petroleum Hydrocarbon- Contaminated Sites. Pharm. Anal. Chem. Open Access 2015, 1. [Google Scholar] [CrossRef] [Green Version]
- Terzaghi, E.; Vitale, C.M.; Salina, G.; Di Guardo, A. Plants Radically Change the Mobility of PCBs in Soil: Role of Different Species and Soil Conditions. J. Hazard. Mater. 2020, 388, 121786. [Google Scholar] [CrossRef]
- Varjani, S.J. Microbial Degradation of Petroleum Hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Shuttleworth, K.L.; Cerniglia, E. Environmental Aspects of PAH Biodegradation. Appl. Biochem. Biotechnol. 1995, 54, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieper, D.H. Aerobic Degradation of Polychlorinated Biphenyls. Appl. Microbiol. Biotechnol. 2005, 67, 170–191. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.T.; Parales, R.E. Aromatic Hydrocarbon Dioxygenases in Environmental Biotechnology. Curr. Opin. Biotechnol. 2000, 11, 236–243. [Google Scholar] [CrossRef]
- Yavas, A.; Icgen, B. Diversity of the Aromatic-Ring-Hydroxylating Dioxygenases in the Monoaromatic Hydrocarbon Degraders Held by a Common Ancestor. Bull. Environ. Contam. Toxicol. 2018, 101, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Kanaly, R.A.; Harayama, S. Advances in the Field of High-Molecular-Weight Polycyclic Aromatic Hydrocarbon Biodegradation by Bacteria. Microb. Biotechnol. 2010, 3, 136–164. [Google Scholar] [CrossRef] [Green Version]
- Wittich, R.M.; Wilkes, H.; Sinnwell, V.; Francke, W.; Fortnagel, P. Metabolism of Dibenzo-p-Dioxin by Sphingomonas sp. Strain RWAppl. Environ. Microbiol. 1992, 58, 1005–1010. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, H.; Wittich, R.M.; Timmis, K.N.; Fortnagel, P.; Francke, W. Degradation of Chlorinated Dibenzofurans and Dibenzo-p-Dioxins by Sphingomonas sp. Strain RWAppl. Environ. Microbiol. 1996, 62, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Chai, B.; Tsoi, T.V.; Iwai, S.; Liu, C.; Fish, J.A.; Gu, C.; Johnson, T.A.; Zylstra, G.; Teppen, B.J.; Li, H.; et al. Sphingomonas wittichii Strain RW1 Genome-Wide Gene Expression Shifts in Response to Dioxins and Clay. PLoS ONE 2016, 11, e0157008. [Google Scholar] [CrossRef] [Green Version]
- Saibu, S.; Adebusoye, S.A.; Oyetibo, G.O.; Rodrigues, D.F. Aerobic Degradation of Dichlorinated Dibenzo-p-Dioxin and Dichlorinated Dibenzofuran by Bacteria Strains Obtained from Tropical Contaminated Soil. Biodegradation 2020, 31, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.; Hu, H.; Wang, W.; Zhou, Z.; Shah, S.B.; Xu, P.; Tang, H. Characterization of a Dibenzofuran-Degrading Strain of Pseudomonas aeruginosa, FA-HZ. Environ. Pollut. 2019, 250, 262–273. [Google Scholar] [CrossRef]
- Nojiri, H.; Omori, T. Molecular Bases of Aerobic Bacterial Degradation of Dioxins: Involvement of Angular Dioxygenation. Biosci. Biotechnol. Biochem. 2002, 66, 2001–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bünz, P.V.; Cook, A.M. Dibenzofuran 4,4a-Dioxygenase from Sphingomonas sp. Strain RW1: Angular Dioxygenation by a Three-Component Enzyme System. J. Bacteriol. 1993, 175, 6467–6475. [Google Scholar] [CrossRef] [Green Version]
- Sander, P.; Wittich, R.M.; Fortnagel, P.; Wilkes, H.; Francke, W. Degradation of 1,2,4-Trichloro- and 1,2,4,5-Tetrachlorobenzene by Pseudomonas strains. Appl. Environ. Microbiol. 1991, 57, 1430–1440. [Google Scholar] [CrossRef] [Green Version]
- Potrawfke, T.; Timmis, K.; Wittich, R. Degradation of 1,2,3,4-Tetrachlorobenzene by Pseudomonas chlororaphis RW. Appl. Environ. Microbiol. 1998, 64, 3798–3806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, K.; Takagi, K.; Iwasaki, A.; Tanaka, N.; Kanesaki, Y.; Martin-Laurent, F.; Igimi, S. Identification of the Hcb Gene Operon Involved in Catalyzing Aerobic Hexachlorobenzene Dechlorination in Nocardioides sp. Strain PD653. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.-Z.; Liu, H.; Zhou, N.-Y. Conversion of Sphingobium chlorophenolicum ATCC 39723 to a Hexachlorobenzene Degrader by Metabolic Engineering. Appl. Environ. Microbiol. 2006, 72, 2283–2286. [Google Scholar] [CrossRef] [Green Version]
- Tiedje, J.M.; Stevens, T.O. The Ecology of an Anaerobic Dechlorinating Consortium; Springer US: Boston, MA, USA, 1988. [Google Scholar] [CrossRef]
- Bunge, M.; Lechner, U. Anaerobic Reductive Dehalogenation of Polychlorinated Dioxins. Appl. Microbiol. Biotechnol. 2009, 84, 429–444. [Google Scholar] [CrossRef]
- Nijenhuis, I.; Kuntze, K. Anaerobic Microbial Dehalogenation of Organohalides—State of the Art and Remediation Strategies. Curr. Opin. Biotechnol. 2016, 38, 33–38. [Google Scholar] [CrossRef]
- Kuiper, I.; Lagendijk, E.L.; Bloemberg, G.V.; Lugtenberg, B.J.J. Rhizoremediation: A Beneficial Plant-Microbe Interaction. Mol. Plant. Microbe Interact. 2004, 17, 6–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyens, N.; Thijs, S.; Popek, R.; Witters, N.; Przybysz, A.; Espenshade, J.; Gawronska, H.; Vangronsveld, J.; Gawronski, S. The Role of Plant–Microbe Interactions and Their Exploitation for Phytoremediation of Air Pollutants. Int. J. Mol. Sci. 2015, 16, 25576–25604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radwan, S.; Sorkhoh, N.; EI-Nemr, I. Oil Biodegradation around Roots. Nature 1995, 376, 302. [Google Scholar] [CrossRef] [PubMed]
- Scharf, B.E.; Hynes, M.F.; Alexandre, G.M. Chemotaxis Signaling Systems in Model Beneficial Plant–Bacteria Associations. Plant. Mol. Biol. 2016, 90, 549–559. [Google Scholar] [CrossRef] [PubMed]
- López-Farfán, D.; Reyes-Darias, J.A.; Matilla, M.A.; Krell, T. Concentration Dependent Effect of Plant Root Exudates on the Chemosensory Systems of Pseudomonas putida KT. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the Root Microbiome by Plant Molecules: The Basis for Targeted Disease Suppression and Plant Growth Promotion. Front. Plant. Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
- Matilla, M.A.; Espinosa-Urgel, M.; Rodríguez-Herva, J.J.; Ramos, J.L.; Ramos-González, M. Genomic Analysis Reveals the Major Driving Forces of Bacterial Life in the Rhizosphere. Genome Biol. 2007, 8, R179. [Google Scholar] [CrossRef] [Green Version]
- Burken, J.G. Uptake and Metabolism of Organic Compounds: Green-Liver Model; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2004. [Google Scholar] [CrossRef]
- Rojo, F. Carbon Catabolite Repression in Pseudomonas: Optimizing Metabolic Versatility and Interactions with the Environment. FEMS Microbiol. Rev. 2010, 34, 658–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rentz, J.A.; Alvarez, P.J.J.; Schnoor, J.L. Repression of Pseudomonas putida Phenanthrene-Degrading Activity by Plant Root Extracts and Exudates. Environ. Microbiol. 2004, 6, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Iyer, B.; Rajput, M.S.; Jog, R.; Joshi, E.; Bharwad, K.; Rajkumar, S. Organic Acid Mediated Repression of Sugar Utilization in Rhizobia. Microbiol. Res. 2016, 192, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Zhang, M.; Toyota, K. Biodegradation of Volatile Organic Compounds and Their Effects on Biodegradability under Co-Existing Conditions. Microbes Environ. 2017, 32, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Bharwad, K.; Rajkumar, S. Modulation of PQQ-Dependent Glucose Dehydrogenase (MGDH and SGDH) Activity by Succinate in Phosphate Solubilizing Plant Growth Promoting Acinetobacter sp. SK2. 3 Biotech. 2020, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Conde, S.; Molina, L.; González, P.; García-Puente, A.; Segura, A. Degradation of Phenanthrene by Novosphingobium sp. HS2a Improved Plant Growth in PAHs-Contaminated Environments. Appl. Microbiol. Biotechnol. 2016, 100, 10627–10636. [Google Scholar] [CrossRef] [PubMed]
- Bello-Akinosho, M.; Makofane, R.; Adeleke, R.; Thantsha, M.; Pillay, M.; Chirima, G.J. Potential of Polycyclic Aromatic Hydrocarbon-Degrading Bacterial Isolates to Contribute to Soil Fertility. BioMed Res. Int. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; Ver Loren van Themaat, E.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E.; et al. Revealing Structure and Assembly Cues for Arabidopsis Root-Inhabiting Bacterial Microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant. Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Cañizares, C.; Jorrín, B.; Poole, P.S.; Tkacz, A. Understanding the Holobiont: The Interdependence of Plants and Their Microbiome. Curr. Opin. Microbiol. 2017, 38, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xiang, Y.; Zhang, Z.; Ling, W.; Gao, Y. Inoculation of a Phenanthrene-Degrading Endophytic Bacterium Reduces the Phenanthrene Level and Alters the Bacterial Community Structure in Wheat. Appl. Microbiol. Biotechnol. 2017, 101, 5199–5212. [Google Scholar] [CrossRef]
- Siciliano, S.D.; Fortin, N.; Mihoc, A.; Wisse, G.; Labelle, S.; Beaumier, D.; Ouellette, D.; Roy, R.; Whyte, L.G.; Banks, M.K.; et al. Selection of Specific Endophytic Bacterial Genotypes by Plants in Response to Soil Contamination. Appl. Environ. Microbiol. 2001, 67, 2469–2475. [Google Scholar] [CrossRef] [Green Version]
- Jiao, S.; Chen, W.; Wei, G. Resilience and Assemblage of Soil Microbiome in Response to Chemical Contamination Combined with Plant Growth. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- Thijs, S.; Sillen, W.; Rineau, F.; Weyens, N.; Vangronsveld, J. Towards an Enhanced Understanding of Plant–Microbiome Interactions to Improve Phytoremediation: Engineering the Metaorganism. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Kaur, N.; Erickson, T.E.; Ball, A.S.; Ryan, M.H. A Review of Germination and Early Growth as a Proxy for Plant Fitness under Petrogenic Contamination—Knowledge Gaps and Recommendations. Sci. Total Environ. 2017, 603–604, 728–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyasi, R.O.; Atagana, H.I. Profiling of Plants at Petroleum Contaminated Site for Phytoremediation. Int. J. Phytoremediation 2018, 20, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Böltner, D.; Godoy, P.; Muñoz-Rojas, J.; Duque, E.; Moreno-Morillas, S.; Sánchez, L.; Ramos, J.L. Rhizoremediation of Lindane by Root-Colonizing Sphingomonas. Microb. Biotechnol. 2007, 1, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Zafra, G.; Absalón, Á.E.; Anducho-Reyes, M.Á.; Fernandez, F.J.; Cortés-Espinosa, D.V. Construction of PAH-Degrading Mixed Microbial Consortia by Induced Selection in Soil. Chemosphere 2017, 172, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Uhlik, O.; Wald, J.; Strejcek, M.; Musilova, L.; Ridl, J.; Hroudova, M.; Vlcek, C.; Cardenas, E.; Mackova, M.; Macek, T. Identification of Bacteria Utilizing Biphenyl, Benzoate, and Naphthalene in Long-Term Contaminated Soil. PLoS ONE 2012, 7, e40653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kappell, A.D.; Wei, Y.; Newton, R.J.; Van Nostrand, J.D.; Zhou, J.; McLellan, S.L.; Hristova, K.R. The Polycyclic Aromatic Hydrocarbon Degradation Potential of Gulf of Mexico Native Coastal Microbial Communities after the Deepwater Horizon Oil Spill. Front. Microbiol. 2014, 5, 205. [Google Scholar] [CrossRef]
- Vergani, L.; Mapelli, F.; Suman, J.; Cajthaml, T.; Uhlik, O.; Borin, S. Novel PCB-Degrading Rhodococcus Strains Able to Promote Plant Growth for Assisted Rhizoremediation of Historically Polluted Soils. PLoS ONE 2019, 14, e0221253. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Megharaj, M.; Wu, C.-Y.; Subashchandrabose, S.R.; Dai, C.-C. Endophyte-Assisted Phytoremediation: Mechanisms and Current Application Strategies for Soil Mixed Pollutants. Crit. Rev. Biotechnol. 2020, 40, 31–45. [Google Scholar] [CrossRef]
- Pino, N.J.; Múnera, L.M.; Peñuela, G.A. Phytoremediation of Soil Contaminated with PCBs Using Different Plants and Their Associated Microbial Communities. Int. J. Phytoremediation 2019, 21, 316–324. [Google Scholar] [CrossRef]
- Vergani, L.; Mapelli, F.; Zanardini, E.; Terzaghi, E.; Di Guardo, A.; Morosini, C.; Raspa, G.; Borin, S. Phyto-Rhizoremediation of Polychlorinated Biphenyl Contaminated Soils: An Outlook on Plant-Microbe Beneficial Interactions. Sci. Total Environ. 2017, 575, 1395–1406. [Google Scholar] [CrossRef]
- Correa-García, S.; Pande, P.; Séguin, A.; St-Arnaud, M.; Yergeau, E. Rhizoremediation of Petroleum Hydrocarbons: A Model System for Plant Microbiome Manipulation. Microb. Biotechnol. 2018, 11, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Hoang, S.A.; Lamb, D.; Seshadri, B.; Sarkar, B.; Choppala, G.; Kirkham, M.B.; Bolan, N.S. Rhizoremediation as a Green Technology for the Remediation of Petroleum Hydrocarbon-Contaminated Soils. J. Hazard. Mater. 2021, 401, 123282. [Google Scholar] [CrossRef]
- Tu, C.; Ma, L.; Guo, P.; Song, F.; Teng, Y.; Zhang, H.; Luo, Y. Rhizoremediation of a Dioxin-like PCB Polluted Soil by Alfalfa: Dynamic Characterization at Temporal and Spatial Scale. Chemosphere 2017, 189, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Terzaghi, E.; Vergani, L.; Mapelli, F.; Borin, S.; Raspa, G.; Zanardini, E.; Morosini, C.; Anelli, S.; Nastasio, P.; Sale, V.M.; et al. Rhizoremediation of Weathered PCBs in a Heavily Contaminated Agricultural Soil: Results of a Biostimulation Trial in Semi Field Conditions. Sci. Total Environ. 2019, 686, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Ancona, V.; Barra Caracciolo, A.; Grenni, P.; Di Lenola, M.; Campanale, C.; Calabrese, A.; Uricchio, V.F.; Mascolo, G.; Massacci, A. Plant-Assisted Bioremediation of a Historically PCB and Heavy Metal-Contaminated Area in Southern Italy. New Biotechnol. 2017, 38, 65–73. [Google Scholar] [CrossRef]
- Sivaram, A.K.; Subashchandrabose, S.R.; Logeshwaran, P.; Lockington, R.; Naidu, R.; Megharaj, M. Rhizodegradation of PAHs Differentially Altered by C3 and C4 Plants. Sci. Rep. 2020, 10, 16109. [Google Scholar] [CrossRef]
- Balseiro-Romero, M.; Gkorezis, P.; Kidd, P.S.; Vangronsveld, J.; Monterroso, C. Enhanced Degradation of Diesel in the Rhizosphere of after Inoculation with Diesel-Degrading and Plant Growth-Promoting Bacterial Strains. J. Environ. Qual. 2016, 45, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Nichols, T.D.; Wolf, D.C.; Rogers, H.B.; Beyrouty, C.A.; Reynolds, C.M. Rhizosphere Microbial Populations in Contaminated Soils. Water. Air. Soil Pollut. 1997, 95, 165–178. [Google Scholar] [CrossRef]
- Jacobsen, C.S. Plant Protection and Rhizosphere Colonization of Barley by Seed Inoculated Herbicide Degrading Burkholderia (Pseudomonas) cepacia DBO1(PRO101) in 2,4-D Contaminated Soil. Plant. Soil 1997, 189, 139–144. [Google Scholar] [CrossRef]
- Liste, H.-H.; Alexander, M. Plant-Promoted Pyrene Degradation in Soil. Chemosphere 2000, 40, 7–10. [Google Scholar] [CrossRef]
- Kelley, S.L.; Aitchison, E.W.; Deshpande, M.; Schnoor, J.L.; Alvarez, P.J.J. Biodegradation of 1,4-Dioxane in Planted and Unplanted Soil: Effect of Bioaugmentation with Amycolata Sp. CB1190. Water Res. 2001, 35, 3791–3800. [Google Scholar] [CrossRef]
- Kuiper, I.; Bloemberg, G.V.; Lugtenberg, B.J.J. Selection of a Plant-Bacterium Pair as a Novel Tool for Rhizostimulation of Polycyclic Aromatic Hydrocarbon-Degrading Bacteria. Mol. Plant. Microbe Interact. 2001, 14, 1197–1205. [Google Scholar] [CrossRef] [Green Version]
- Pai, S.G.; Riley, M.B.; Camper, N.D. Microbial Degradation of Mefenoxam in Rhizosphere of Zinnia angustifolia. Chemosphere 2001, 44, 577–582. [Google Scholar] [CrossRef]
- Anokhina, T.O.; Kochetkov, V.V.; Zelenkova, N.F.; Balakshina, V.V.; Boronin, A.M. Biodegradation of Phenanthrene by Pseudomonas Bacteria Bearing Rhizospheric Plasmids in Model Plant–Microbial Associations. Appl. Biochem. Microbiol. 2004, 40, 568–572. [Google Scholar] [CrossRef]
- Johnson, D. Enhanced Dissipation of Chrysene in Planted Soil: The Impact of a Rhizobial Inoculum. Soil Biol. Biochem. 2004, 36, 33–38. [Google Scholar] [CrossRef]
- Sheng, X.; Gong, J. Increased Degradation of Phenanthrene in Soil by Pseudomonas sp. GF3 in the Presence of Wheat. Soil Biol. Biochem. 2006, 38, 2587–2592. [Google Scholar] [CrossRef]
- Child, R.; Miller, C.D.; Liang, Y.; Narasimham, G.; Chatterton, J.; Harrison, P.; Sims, R.C.; Britt, D.; Anderson, A.J. Polycyclic Aromatic Hydrocarbon-Degrading Mycobacterium Isolates: Their Association with Plant Roots. Appl. Microbiol. Biotechnol. 2007, 75, 655–663. [Google Scholar] [CrossRef]
- Muratova, A.; Pozdnyakova, N.; Golubev, S.; Wittenmayer, L.; Makarov, O.; Merbach, W.; Turkovskaya, O. Oxidoreductase Activity of Sorghum Root Exudates in a Phenanthrene-Contaminated Environment. Chemosphere 2009, 74, 1031–1036. [Google Scholar] [CrossRef]
- Andria, V.; Reichenauer, T.; Sessitsch, A. Expression of Alkane Monooxygenase (AlkB) Genes by Plant-Associated Bacteria in the Rhizosphere and Endosphere of Italian Ryegrass (Lolium multiflorum L.) Grown in Diesel Contaminated Soil. Environ. Pollut. Barking Essex 1987 2009, 157, 3347–3350. [Google Scholar] [CrossRef] [PubMed]
- Muratova, A.Y.; Bondarenkova, A.D.; Panchenko, L.V.; Turkovskaya, O.V. Use of Integrated Phytoremediation for Cleaning-up of Oil-Sludge-Contaminated Soil. Appl. Biochem. Microbiol. 2010, 46, 789–794. [Google Scholar] [CrossRef]
- Afzal, M.; Yousaf, S.; Reichenauer, T.G.; Sessitsch, A. The Inoculation Method Affects Colonization and Performance of Bacterial Inoculant Strains in the Phytoremediation of Soil Contaminated with Diesel Oil. Int. J. Phytoremediation 2012, 14, 35–47. [Google Scholar] [CrossRef]
- Chouychai, W.; Thongkukiatkul, A.; Upatham, S.; Pokethitiyook, P.; Kruatrachue, M.; Lee, H. Effect of Corn Plant on Survival and Phenanthrene Degradation Capacity of Pseudomonas sp. UG14Lr in Two Soils. Int. J. Phytoremediation 2012, 14, 585–595. [Google Scholar] [CrossRef]
- Yu, X.Z.; Wu, S.C.; Wu, F.Y.; Wong, M.H. Enhanced Dissipation of PAHs from Soil Using Mycorrhizal Ryegrass and PAH-Degrading Bacteria. J. Hazard. Mater. 2011, 186, 1206–1217. [Google Scholar] [CrossRef]
- Teng, Y.; Shen, Y.; Luo, Y.; Sun, X.; Sun, M.; Fu, D.; Li, Z.; Christie, P. Influence of Rhizobium meliloti on Phytoremediation of Polycyclic Aromatic Hydrocarbons by Alfalfa in an Aged Contaminated Soil. J. Hazard. Mater. 2011, 186, 1271–1276. [Google Scholar] [CrossRef]
- Hong, S.H.; Ryu, H.; Kim, J.; Cho, K.-S. Rhizoremediation of Diesel-Contaminated Soil Using the Plant Growth-Promoting Rhizobacterium Gordonia sp. S2RP-17. Biodegradation 2011, 22, 593–601. [Google Scholar] [CrossRef]
- Afzal, M.; Yousaf, S.; Reichenauer, T.G.; Kuffner, M.; Sessitsch, A. Soil Type Affects Plant Colonization, Activity and Catabolic Gene Expression of Inoculated Bacterial Strains during Phytoremediation of Diesel. J. Hazard. Mater. 2011, 186, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Gałązka, A.; Król, M.; Perzyński, A. The Efficiency of Rhizosphere Bioremediation with Azospirillum sp. and Pseudomonas stutzeri in Soils Freshly Contaminated with PAHs and Diesel Fuel. Pol. J. Environ. Stud. 2012, 21, 345–353. [Google Scholar]
- Bisht, S.; Pandey, P.; Kaur, G.; Aggarwal, H.; Sood, A.; Sharma, S.; Kumar, V.; Bisht, N.S. Utilization of Endophytic Strain Bacillus sp. SBER3 for Biodegradation of Polyaromatic Hydrocarbons (PAH) in Soil Model System. Eur. J. Soil Biol. 2014, 60, 67–76. [Google Scholar] [CrossRef]
- Khan, Z.; Roman, D.; Kintz, T.; Alas, M.; Yap, R.; Doty, S. Degradation, Phytoprotection and Phytoremediation of Phenanthrene by Endophyte Pseudomonas putida, PD. Environ. Sci. Technol. 2014, 48. [Google Scholar] [CrossRef]
- Baneshi, M.M.; Rezaei Kalantary, R.; Jonidi Jafari, A.; Nasseri, S.; Jaafarzadeh, N.; Esrafili, A. Effect of Bioaugmentation to Enhance Phytoremediation for Removal of Phenanthrene and Pyrene from Soil with Sorghum and Onobrychis sativa. J. Environ. Health Sci. Eng. 2014, 12, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizarro-Tobías, P.; Fernández, M.; Niqui, J.L.; Solano, J.; Duque, E.; Ramos, J.-L.; Roca, A. Restoration of a Mediterranean Forest after a Fire: Bioremediation and Rhizoremediation Field-Scale Trial. Microb. Biotechnol. 2015, 8, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Gong, Z.; Miao, R.; Rookes, J.; Cahill, D.; Zhuang, J. Microbial Mechanisms Controlling the Rhizosphere Effect of Ryegrass on Degradation of Polycyclic Aromatic Hydrocarbons in an Aged-Contaminated Agricultural Soil. Soil Biol. Biochem. 2017, 113, 130–142. [Google Scholar] [CrossRef]
- Dealtry, S.; Ghizelini, A.M.; Mendonça-Hagler, L.C.S.; Chaloub, R.M.; Reinert, F.; devCampos, T.M.P.; Gomes, N.C.M.; Smalla, K. Petroleum Contamination and Bioaugmentation in Bacterial Rhizosphere Communities from Avicennia schaueriana. Braz. J. Microbiol. 2018, 49, 757–769. [Google Scholar] [CrossRef]
- Iqbal, A.; Arshad, M.; Karthikeyan, R.; Gentry, T.J.; Rashid, J.; Ahmed, I.; Schwab, A.P. Diesel Degrading Bacterial Endophytes with Plant Growth Promoting Potential Isolated from a Petroleum Storage Facility. 3 Biotech. 2019, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Jampasri, K.; Pokethitiyook, P.; Poolpak, T.; Kruatrachue, M.; Ounjai, P.; Kumsopa, A. Bacteria-Assisted Phytoremediation of Fuel Oil and Lead Co-Contaminated Soil in the Salt-Stressed Condition by Chromolaena odorata and Micrococcus luteus. Int. J. Phytoremediation 2020, 22, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.C.; Cryder, Z.; Khoury, R.; Carlan, C.; Gan, J. Bioremediation of PAH-Contaminated Shooting Range Soil Using Integrated Approaches. Sci. Total Environ. 2020, 726, 138440. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, M.; Jha, P.; Paul, A.T.; Singh, R.P.; Jha, P.N. Evaluation of Biphenyl- and Polychlorinated-Biphenyl (PCB) Degrading Rhodococcus sp. MAPN-1 on Growth of Morus alba by Pot Study. Int. J. Phytoremediation 2020, 22, 1487–1496. [Google Scholar] [CrossRef]
- Simmer, R.; Mathieu, J.; da Silva, M.L.B.; Lashmit, P.; Gopishetty, S.; Alvarez, P.J.J.; Schnoor, J.L. Bioaugmenting the Poplar Rhizosphere to Enhance Treatment of 1,4-Dioxane. Sci. Total Environ. 2020, 744, 140823. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Miao, R.; Guo, M.; Zhou, Y. Effects of Fire Phoenix (a Genotype Mixture of Fesctuca arundinecea L.) and Mycobacterium sp. on the Degradation of PAHs and Bacterial Community in Soil. Environ. Sci. Pollut. Res. Int. 2021. [Google Scholar] [CrossRef] [PubMed]
- Simonich, S.L.; Hites, R.A. Vegetation-Atmosphere Partitioning of Polycyclic Aromatic Hydrocarbons. Environ. Sci. Technol. 1994, 28, 939–943. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial Life in the Phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Lindow, S.E.; Brandl, M.T. Microbiology of the Phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koskella, B. The Phyllosphere. Curr. Biol. CB 2020, 30, R1143–R1146. [Google Scholar] [CrossRef]
- Carvalho, S.D.; Castillo, J.A. Influence of Light on Plant-Phyllosphere Interaction. Front. Plant. Sci. 2018, 9, 1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leveau, J.H. A Brief from the Leaf: Latest Research to Inform Our Understanding of the Phyllosphere Microbiome. Curr. Opin. Microbiol. 2019, 49, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Redford, A.J.; Bowers, R.M.; Knight, R.; Linhart, Y.; Fierer, N. The Ecology of the Phyllosphere: Geographic and Phylogenetic Variability in the Distribution of Bacteria on Tree Leaves. Environ. Microbiol. 2010, 12, 2885–2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kembel, S.W.; O’Connor, T.K.; Arnold, H.K.; Hubbell, S.P.; Wright, S.J.; Green, J.L. Relationships between Phyllosphere Bacterial Communities and Plant Functional Traits in a Neotropical Forest. Proc. Natl. Acad. Sci. USA 2014, 111, 13715–13720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grady, K.L.; Sorensen, J.W.; Stopnisek, N.; Guittar, J.; Shade, A. Assembly and Seasonality of Core Phyllosphere Microbiota on Perennial Biofuel Crops. Nat. Commun. 2019, 10, 4135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laforest-Lapointe, I.; Messier, C.; Kembel, S.W. Tree Leaf Bacterial Community Structure and Diversity Differ along a Gradient of Urban Intensity. mSystems 2017, 2, e00087-17. [Google Scholar] [CrossRef] [Green Version]
- Shakir, S.; Zaidi, S.S.-A.; de Vries, F.T.; Mansoor, S. Plant Genetic Networks Shaping Phyllosphere Microbial Community. Trends Genet. 2020, S0168952520302468. [Google Scholar] [CrossRef]
- Stone, B.W.G.; Jackson, C.R. Biogeographic Patterns Between Bacterial Phyllosphere Communities of the Southern Magnolia (Magnolia grandiflora) in a Small Forest. Microb. Ecol. 2016, 71, 954–961. [Google Scholar] [CrossRef]
- Bakan, B.; Marion, D. Assembly of the Cutin Polyester: From Cells to Extracellular Cell Walls. Plants 2017, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Remus-Emsermann, M.N.P.; Schlechter, R.O. Phyllosphere Microbiology: At the Interface between Microbial Individuals and the Plant Host. New Phytol. 2018, 218, 1327–1333. [Google Scholar] [CrossRef]
- Knief, C.; Ramette, A.; Frances, L.; Alonso-Blanco, C.; Vorholt, J.A. Site and Plant Species Are Important Determinants of the Methylobacterium Community Composition in the Plant Phyllosphere. ISME J. 2010, 4, 719–728. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Hiradate, S.; Koitabashi, M.; Kamo, T.; Tsushima, S. Phyllosphere Methylobacterium Bacteria Contain UVA-Absorbing Compounds. J. Photochem. Photobiol. B 2017, 167, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Vacher, C.; Hampe, A.; Porté, A.J.; Sauer, U.; Compant, S.; Morris, C.E. The Phyllosphere: Microbial Jungle at the Plant–Climate Interface. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 1–24. [Google Scholar] [CrossRef]
- Wei, X.; Lyu, S.; Yu, Y.; Wang, Z.; Liu, H.; Pan, D.; Chen, J. Phylloremediation of Air Pollutants: Exploiting the Potential of Plant Leaves and Leaf-Associated Microbes. Front. Plant. Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Łukowski, A.; Popek, R.; Karolewski, P. Particulate Matter on Foliage of Betula pendula, Quercus robur, and Tilia cordata: Deposition and Ecophysiology. Environ. Sci. Pollut. Res. Int. 2020, 27, 10296–10307. [Google Scholar] [CrossRef] [Green Version]
- Popek, R.; Łukowski, A.; Bates, C.; Oleksyn, J. Accumulation of Particulate Matter, Heavy Metals, and Polycyclic Aromatic Hydrocarbons on the Leaves of Tilia cordata Mill. in Five Polish Cities with Different Levels of Air Pollution. Int. J. Phytoremediation 2017, 19, 1134–1141. [Google Scholar] [CrossRef]
- Espenshade, J.; Thijs, S.; Gawronski, S.; Bové, H.; Weyens, N.; Vangronsveld, J. Influence of Urbanization on Epiphytic Bacterial Communities of the Platanus × Hispanica Tree Leaves in a Biennial Study. Front. Microbiol. 2019, 10, 675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imperato, V.; Kowalkowski, L.; Portillo-Estrada, M.; Gawronski, S.W.; Vangronsveld, J.; Thijs, S. Characterisation of the Carpinus betulus L. Phyllomicrobiome in Urban and Forest Areas. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzetti, A.; Gandolfi, I.; Bestetti, G.; Padoa Schioppa, E.; Canedoli, C.; Brambilla, D.; Cappelletti, D.; Sebastiani, B.; Federici, E.; Papacchini, M.; et al. Plant-Microorganisms Interaction Promotes Removal of Air Pollutants in Milan (Italy) Urban Area. J. Hazard. Mater. 2020, 384, 121021. [Google Scholar] [CrossRef]
- Gandolfi, I.; Canedoli, C.; Imperato, V.; Tagliaferri, I.; Gkorezis, P.; Vangronsveld, J.; Padoa Schioppa, E.; Papacchini, M.; Bestetti, G.; Franzetti, A. Diversity and Hydrocarbon-Degrading Potential of Epiphytic Microbial Communities on Platanus x Acerifolia Leaves in an Urban Area. Environ. Pollut. 2017, 220, 650–658. [Google Scholar] [CrossRef]
- Sandhu, A.; Halverson, L.J.; Beattie, G.A. Bacterial Degradation of Airborne Phenol in the Phyllosphere. Environ. Microbiol. 2007, 9, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, A.; Halverson, L.J.; Beattie, G.A. Identification and Genetic Characterization of Phenol-Degrading Bacteria from Leaf Microbial Communities. Microb. Ecol. 2009, 57, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.; Sorkhoh, N.; Salamah, S.; Eliyas, M.; Radwan, S. The Potential of Epiphytic Hydrocarbon-Utilizing Bacteria on Legume Leaves for Attenuation of Atmospheric Hydrocarbon Pollutants. J. Environ. Manag. 2012, 93, 113–120. [Google Scholar] [CrossRef]
- Sorkhoh, N.A.; Al-Mailem, D.M.; Ali, N.; Al-Awadhi, H.; Salamah, S.; Eliyas, M.; Radwan, S.S. Bioremediation of Volatile Oil Hydrocarbons by Epiphytic Bacteria Associated with American Grass (Cynodon sp.) and Broad Bean (Vicia faba) Leaves. Int. Biodeterior. Biodegrad. 2011, 65, 797–802. [Google Scholar] [CrossRef]
- Waight, K.; Pinyakong, O.; Luepromchai, E. Degradation of Phenanthrene on Plant Leaves by Phyllosphere Bacteria. J. Gen. Appl. Microbiol. 2007, 53, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Sangthong, S.; Suksabye, P.; Thiravetyan, P. Air-Borne Xylene Degradation by Bougainvillea buttiana and the Role of Epiphytic Bacteria in the Degradation. Ecotoxicol. Environ. Saf. 2016, 126, 273–280. [Google Scholar] [CrossRef]
- Yutthammo, C.; Thongthammachat, N.; Pinphanichakarn, P.; Luepromchai, E. Diversity and Activity of PAH-Degrading Bacteria in the Phyllosphere of Ornamental Plants. Microb. Ecol. 2010, 59, 357–368. [Google Scholar] [CrossRef]
- Daudzai, Z.; Treesubsuntorn, C.; Thiravetyan, P. Inoculated Clitoria ternatea with Bacillus cereus ERBP for Enhancing Gaseous Ethylbenzene Phytoremediation: Plant Metabolites and Expression of Ethylbenzene Degradation Genes. Ecotoxicol. Environ. Saf. 2018, 164, 50–60. [Google Scholar] [CrossRef]
- De Kempeneer, L.; Sercu, B.; Vanbrabant, W.; Van Langenhove, H.; Verstraete, W. Bioaugmentation of the Phyllosphere for the Removal of Toluene from Indoor Air. Appl. Microbiol. Biotechnol. 2004, 64, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Terzaghi, E.; De Nicola, F.; Cerabolini, B.E.L.; Posada-Baquero, R.; Ortega-Calvo, J.-J.; Di Guardo, A. Role of Photo- and Biodegradation of Two PAHs on Leaves: Modelling the Impact on Air Quality Ecosystem Services Provided by Urban Trees. Sci. Total Environ. 2020, 739, 139893. [Google Scholar] [CrossRef]
- Pettit, T.; Irga, P.J.; Torpy, F.R. Towards Practical Indoor Air Phytoremediation: A Review. Chemosphere 2018, 208, 960–974. [Google Scholar] [CrossRef] [PubMed]
- Scheublin, T.R.; Deusch, S.; Moreno-Forero, S.K.; Müller, J.A.; van der Meer, J.R.; Leveau, J.H.J. Transcriptional Profiling of Gram-Positive A Rthrobacter in the Phyllosphere: Induction of Pollutant Degradation Genes by Natural Plant Phenolic Compounds. Environ. Microbiol. 2014, 16, 2212–2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brilli, F.; Fares, S.; Ghirardo, A.; de Visser, P.; Calatayud, V.; Muñoz, A.; Annesi-Maesano, I.; Sebastiani, F.; Alivernini, A.; Varriale, V.; et al. Plants for Sustainable Improvement of Indoor Air Quality. Trends Plant. Sci. 2018, 23, 507–512. [Google Scholar] [CrossRef] [PubMed]
- González-Martín, J.; Kraakman, N.J.R.; Pérez, C.; Lebrero, R.; Muñoz, R. A State–of–the-Art Review on Indoor Air Pollution and Strategies for Indoor Air Pollution Control. Chemosphere 2021, 262, 128376. [Google Scholar] [CrossRef]
- Salthammer, T. Emerging Indoor Pollutants. Int. J. Hyg. Environ. Health 2020, 224, 113423. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Min, X.; Weng, M. A Review of Polychlorinated Biphenyls (PCBs) Pollution in Indoor Air Environment. J. Air Waste Manag. Assoc. 2016, 66, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Teiri, H.; Pourzamani, H.; Hajizadeh, Y. Phytoremediation of VOCs from Indoor Air by Ornamental Potted Plants: A Pilot Study Using a Palm Species under the Controlled Environment. Chemosphere 2018, 197, 375–381. [Google Scholar] [CrossRef]
- Hörmann, V.; Brenske, K.-R.; Ulrichs, C. Assessment of Filtration Efficiency and Physiological Responses of Selected Plant Species to Indoor Air Pollutants (Toluene and 2-Ethylhexanol) under Chamber Conditions. Environ. Sci. Pollut. Res. 2018, 25, 447–458. [Google Scholar] [CrossRef]
- Oishi, Y. Comparison of Moss and Pine Needles as Bioindicators of Transboundary Polycyclic Aromatic Hydrocarbon Pollution in Central Japan. Environ. Pollut. 2018, 234, 330–338. [Google Scholar] [CrossRef]
- Sari, M.F.; Esen, F.; Tasdemir, Y. Biomonitoring and Source Identification of Polycyclic Aromatic Hydrocarbons (PAHs) Using Pine Tree Components from Three Different Sites in Bursa, Turkey. Arch. Environ. Contam. Toxicol. 2020, 78, 646–657. [Google Scholar] [CrossRef] [PubMed]
- Nowak, D.J.; Crane, D.E.; Stevens, J.C. Air Pollution Removal by Urban Trees and Shrubs in the United States. Urban For Urban Green. 2006, 4, 115–123. [Google Scholar] [CrossRef]
- Lafortezza, R.; Davies, C.; Sanesi, G.; Konijnendijk, C. Green Infrastructure as a Tool to Support Spatial Planning in European Urban Regions. IForest-Biogeosciences and Forrestry. 2013, 6, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.R. Biodiversity Conservation and the Extinction of Experience. Trends Ecol. Evol. 2005, 20, 430–434. [Google Scholar] [CrossRef]
- Lafortezza, R.; Carrus, G.; Sanesi, G.; Davies, C. Benefits and Well-Being Perceived by People Visiting Green Spaces in Periods of Heat Stress. Urban For Urban Green. 2009, 8, 97–108. [Google Scholar] [CrossRef]
- Adinolfi, C.; Suárez-Cáceres, G.P.; Cariñanos, P. Relation between Visitors’ Behaviour and Characteristics of Green Spaces in the City of Granada, South-Eastern Spain. Urban For Urban Green. 2014, 13, 534–542. [Google Scholar] [CrossRef]
- Bolund, P.; Hunhammar, S. Ecosystem Services in Urban Areas. Ecol. Econ. 1999, 29, 293–301. [Google Scholar] [CrossRef]
- Morancho, A.B. A Hedonic Valuation of Urban Green Areas. Landsc. Urban. Plan. 2003, 66, 35–41. [Google Scholar] [CrossRef]
- Jim, C.Y.; Chen, W.Y. External Effects of Neighbourhood Parks and Landscape Elements on High-Rise Residential Value. Land Use Policy 2010, 27, 662–670. [Google Scholar] [CrossRef]
- Xu, T.; Sathaye, J.; Akbari, H.; Garg, V.; Tetali, S. Quantifying the Direct Benefits of Cool Roofs in an Urban Setting: Reduced Cooling Energy Use and Lowered Greenhouse Gas Emissions. Build. Environ. 2012, 48, 1–6. [Google Scholar] [CrossRef]
- Lehmann, I.; Mathey, J.; Rößler, S.; Bräuer, A.; Goldberg, V. Urban Vegetation Structure Types as a Methodological Approach for Identifying Ecosystem Services–Application to the Analysis of Micro-Climatic Effects. Ecol. Indic. 2014, 42, 58–72. [Google Scholar] [CrossRef]
- Eigenbrod, F.; Bell, V.A.; Davies, H.N.; Heinemeyer, A.; Armsworth, P.R.; Gaston, K.J. The Impact of Projected Increases in Urbanization on Ecosystem Services. Proc. R. Soc. B Biol. Sci. 2011, 278, 3201–3208. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Lin, J.; Cui, S.; Qiu, Q.; Zhao, Q. Exploring the Relationship between Urban Transportation Energy Consumption and Transition of Settlement Morphology: A Case Study on Xiamen Island, China. Habitat Int. 2013, 37, 70–79. [Google Scholar] [CrossRef]
- Bowler, D.E.; Buyung-Ali, L.; Knight, T.M.; Pullin, A.S. Urban Greening to Cool Towns and Cities: A Systematic Review of the Empirical Evidence. Landsc. Urban. Plan. 2010, 97, 147–155. [Google Scholar] [CrossRef]
- Byrne, J.; Sipe, N.; Searle, G. Green around the Gills? The Challenge of Density for Urban Greenspace Planning in SEQ. Aust. Plan. 2010, 47, 162–177. [Google Scholar] [CrossRef] [Green Version]
- Lin, B.; Meyers, J.; Barnett, G. Understanding the Potential Loss and Inequities of Green Space Distribution with Urban Densification. Urban. For. Urban. Green. 2015, 14, 952–958. [Google Scholar] [CrossRef]
- Pérez, G.; Coma, J.; Martorell, I.; Cabeza, L.F. Vertical Greenery Systems (VGS) for Energy Saving in Buildings: A Review. Renew. Sustain. Energy Rev. 2014, 39, 139–165. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.; Liu, F.; Zhang, X.; Gao, T. Difference of Airborne Particulate Matter Concentration in Urban Space with Different Green Coverage Rates in Baoji, China. Int. J. Environ. Res. Public. Health 2019, 16, 1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, T.; Liu, F.; Wang, Y.; Mu, S.; Qiu, L. Reduction of Atmospheric Suspended Particulate Matter Concentration and Influencing Factors of Green Space in Urban Forest Park. Forests 2020, 11, 950. [Google Scholar] [CrossRef]
- Yoo, S.-Y.; Kim, T.; Ham, S.; Choi, S.; Park, C.-R. Importance of Urban Green at Reduction of Particulate Matters in Sihwa Industrial Complex, Korea. Sustainability 2020, 12, 7647. [Google Scholar] [CrossRef]
- Irga, P.J.; Pettit, T.J.; Torpy, F.R. The Phytoremediation of Indoor Air Pollution: A Review on the Technology Development from the Potted Plant through to Functional Green Wall Biofilters. Rev. Environ. Sci. Biotechnol. 2018, 17, 395–415. [Google Scholar] [CrossRef]
- Guo, L.; Ma, S.; Zhao, D.; Zhao, B.; Xu, B.; Wu, J.; Tong, J.; Chen, D.; Ma, Y.; Li, M.; et al. Experimental Investigation of Vegetative Environment Buffers in Reducing Particulate Matters Emitted from Ventilated Poultry House. J. Air Waste Manag. Assoc. 2019, 69, 934–943. [Google Scholar] [CrossRef] [Green Version]
- Darlington, A.B.; Dat, J.F.; Dixon, M.A. The Biofiltration of Indoor Air: Air Flux and Temperature Influences the Removal of Toluene, Ethylbenzene, and Xylene. Environ. Sci. Technol. 2001, 35, 240–246. [Google Scholar] [CrossRef]
- Darlington, A.; Chan, M.; Malloch, D.; Pilger, C.; Dixon, M.A. The Biofiltration of Indoor Air: Implications for Air Quality. Indoor Air 2000, 10, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Paull, N.J.; Irga, P.J.; Torpy, F.R. Active Green Wall Plant Health Tolerance to Diesel Smoke Exposure. Environ. Pollut. 2018, 240, 448–456. [Google Scholar] [CrossRef]
- Lundholm, J.; MacIvor, J.S.; MacDougall, Z.; Ranalli, M. Plant Species and Functional Group Combinations Affect Green Roof Ecosystem Functions. PLoS ONE 2010, 5, e9677. [Google Scholar] [CrossRef] [Green Version]
- Irga, P.J.; Paull, N.J.; Abdo, P.; Torpy, F.R. An Assessment of the Atmospheric Particle Removal Efficiency of an In-Room Botanical Biofilter System. Build. Environ. 2017, 115, 281–290. [Google Scholar] [CrossRef]
- Torpy, F.R.; Irga, P.J.; Burchett, M.D. Reducing Indoor Air Pollutants Through Biotechnology. In Biotechnologies and Biomimetics for Civil Engineering; Pacheco Torgal, F., Labrincha, J.A., Diamanti, M.V., Yu, C.-P., Lee, H.K., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 181–210. [Google Scholar] [CrossRef]
- Soreanu, G.; Dixon, M.; Darlington, A. Botanical Biofiltration of Indoor Gaseous Pollutants–A Mini-Review. Chem. Eng. J. 2013, 229, 585–594. [Google Scholar] [CrossRef]
- Speak, A.F.; Rothwell, J.J.; Lindley, S.J.; Smith, C.L. Urban Particulate Pollution Reduction by Four Species of Green Roof Vegetation in a UK City. Atmos. Environ. 2012, 61, 283–293. [Google Scholar] [CrossRef]
- Paull, N.J.; Krix, D.; Torpy, F.R.; Irga, P.J. Can Green Walls Reduce Outdoor Ambient Particulate Matter, Noise Pollution and Temperature? Int. J. Environ. Res. Public. Health 2020, 17, 5084. [Google Scholar] [CrossRef]
- Russell, J.A.; Hu, Y.; Chau, L.; Pauliushchyk, M.; Anastopoulos, I.; Anandan, S.; Waring, M.S. Indoor-Biofilter Growth and Exposure to Airborne Chemicals Drive Similar Changes in Plant Root Bacterial Communities. Appl. Environ. Microbiol. 2014, 80, 4805–4813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reese, A.T.; Savage, A.; Youngsteadt, E.; McGuire, K.L.; Koling, A.; Watkins, O.; Frank, S.D.; Dunn, R.R. Urban Stress Is Associated with Variation in Microbial Species Composition-but Not Richness-in Manhattan. ISME J. 2016, 10, 751–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, A.S.; Purnell, K.; Palmer, M.I.; Stein, J.; McGuire, K.L. Microbial Composition and Functional Diversity Differ Across Urban Green Infrastructure Types. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Lymperopoulou, D.S.; Adams, R.I.; Lindow, S.E. Contribution of Vegetation to the Microbial Composition of Nearby Outdoor Air. Appl. Environ. Microbiol. 2016, 82, 3822–3833. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Ma, J.; Wei, J.; Gong, X.; Yu, X.; Guo, H.; Zhao, Y. Biochar Increases Plant Growth and Alters Microbial Communities via Regulating the Moisture and Temperature of Green Roof Substrates. Sci. Total Environ. 2018, 635, 333–342. [Google Scholar] [CrossRef]
- Molineux, C.J.; Connop, S.P.; Gange, A.C. Manipulating Soil Microbial Communities in Extensive Green Roof Substrates. Sci. Total Environ. 2014, 493, 632–638. [Google Scholar] [CrossRef]
- Kaur, M.; Nagpal, A.K. Evaluation of Air Pollution Tolerance Index and Anticipated Performance Index of Plants and Their Application in Development of Green Space along the Urban Areas. Environ. Sci. Pollut. Res. 2017, 24, 18881–18895. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, M.; Mortazavi, S.N.; Hajizadeh, Y. Evaluation of Anticipated Performance Index, Biochemical, and Physiological Parameters of Cupressus arizonica Greene and Juniperus excelsa Bieb for Greenbelt Development and Biomonitoring of Air Pollution. Int. J. Phytoremediation 2019, 21, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, H.; Bolinius, D.J.; Jahnke, A.; MacLeod, M. Mass Transfer of Hydrophobic Organic Chemicals between Silicone Sheets and through Plant Leaves and Low-Density Polyethylene. Chemosphere 2016, 164, 683–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fantke, P.; Arnot, J.A.; Doucette, W.J. Improving Plant Bioaccumulation Science through Consistent Reporting of Experimental Data. J. Environ. Manage. 2016, 181, 374–384. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, J.; Leglize, P.; Vincent, Q.; Zelko, I.; Mustin, C.; Ouvrard, S.; Sterckeman, T. Effect and Localization of Phenanthrene in Maize Roots. Chemosphere 2016, 149, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Cennerazzo, J.; de Junet, A.; Audinot, J.-N.; Leyval, C. Dynamics of PAHs and Derived Organic Compounds in a Soil-Plant Mesocosm Spiked with 13C-Phenanthrene. Chemosphere 2017, 168, 1619–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trine, L.S.D.; Davis, E.L.; Roper, C.; Truong, L.; Tanguay, R.L.; Simonich, S.L.M. Formation of PAH Derivatives and Increased Developmental Toxicity during Steam Enhanced Extraction Remediation of Creosote Contaminated Superfund Soil. Environ. Sci. Technol. 2019, 53, 4460–4469. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial Interactions within the Plant Holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ding, Q.; Chao, Y.; Wei, X.; Wang, S.; Qiu, R. Structural Development and Assembly Patterns of the Root-Associated Microbiomes during Phytoremediation. Sci. Total Environ. 2018, 644, 1591–1601. [Google Scholar] [CrossRef]
- Alkio, M.; Tabuchi, T.M.; Wang, X.; Colón-Carmona, A. Stress Responses to Polycyclic Aromatic Hydrocarbons in Arabidopsis Include Growth Inhibition and Hypersensitive Response-like Symptoms. J. Exp. Bot. 2005, 56, 2983–2994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisman, D.; Alkio, M.; Colón-Carmona, A. Transcriptional Responses to Polycyclic Aromatic Hydrocarbon-Induced Stress in Arabidopsis thaliana Reveal the Involvement of Hormone and Defense Signaling Pathways. BMC Plant. Biol. 2010, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Carvalhais, L.C.; Crawford, M.; Singh, E.; Dennis, P.G.; Pieterse, C.M.J.; Schenk, P.M. Inner Plant Values: Diversity, Colonization and Benefits from Endophytic Bacteria. Front. Microbiol. 2017, 8, 2552. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The Chemistry of Plant–Microbe Interactions in the Rhizosphere and the Potential for Metabolomics to Reveal Signaling Related to Defense Priming and Induced Systemic Resistance. Front. Plant. Sci. 2018, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Ghatak, A.; Chaturvedi, P.; Weckwerth, W. Metabolomics in Plant Stress Physiology. In Plant Genetics and Molecular Biology; Varshney, R.K., Pandey, M.K., Chitikineni, A., Eds.; Advances in Biochemical Engineering/Biotechnology; Springer International Publishing: Cham, Switzerland, 2018; Volume 164, pp. 187–236. [Google Scholar] [CrossRef]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic Stress Responses and Microbe-Mediated Mitigation in Plants: The Omics Strategies. Front. Plant. Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, K.; Adamec, V.; Bañuelos, A.; Bruse, M.; Bürger, M.; Damsgaard, O.; Dufek, J.; Hirsch, J.; Lefebre, F.; Pérez-Lacorzana, J.M.; et al. An Integrated Methodology to Assess the Benefits of Urban Green Space. Sci. Total Environ. 2004, 334–335, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Pandey, M.; Tripathi, B.D. Assessment of Air Pollution Tolerance Index of Some Plants to Develop Vertical Gardens near Street Canyons of a Polluted Tropical City. Ecotoxicol. Environ. Saf. 2016, 134, 358–364. [Google Scholar] [CrossRef]
- Morella, N.M.; Weng, F.C.-H.; Joubert, P.M.; Metcalf, C.J.E.; Lindow, S.; Koskella, B. Successive Passaging of a Plant-Associated Microbiome Reveals Robust Habitat and Host Genotype-Dependent Selection. Proc. Natl. Acad. Sci. USA 2020, 117, 1148–1159. [Google Scholar] [CrossRef]
Figure 1.
(a) Chemical structures of different polycyclic aromatic hydrocarbons (PAHs), polychlorinated biphenyls (PCBs), dioxin and dioxin-like molecules; (b) Scheme for the aerobic bacterial degradation of phenanthrene. (c) A general scheme for the aerobic bacterial degradation of monochlorobenzene and for the anoxic bacterial dehalogenation and degradation of hexachlorobenzene to benzene and its oxidative mineralization to CO2. Both contaminants end up in the central bacterial metabolism pathways from which energy and biomass are obtained. In both cases, chlorine is released as an environmentally friendly chloride ion.
Figure 1.
(a) Chemical structures of different polycyclic aromatic hydrocarbons (PAHs), polychlorinated biphenyls (PCBs), dioxin and dioxin-like molecules; (b) Scheme for the aerobic bacterial degradation of phenanthrene. (c) A general scheme for the aerobic bacterial degradation of monochlorobenzene and for the anoxic bacterial dehalogenation and degradation of hexachlorobenzene to benzene and its oxidative mineralization to CO2. Both contaminants end up in the central bacterial metabolism pathways from which energy and biomass are obtained. In both cases, chlorine is released as an environmentally friendly chloride ion.

Figure 2.
Schematic representation of the fate of contaminants. In green are shown plant-dependent processes; in pink, physical processes. Contaminants are deposited on surfaces (plants and soil) where they can be adsorbed. Parts of these contaminants are absorbed and then enter the plant structures. Contaminants can also be translocated from the roots to the shoots. Wind and evaporation can also mobilize contaminants from soil to leaves. BTEX (benzene, toluene, ethylbenzene and xylenes); PM2.5 (fine particulate matter); PCBs (polychlorinated biphenyls) and PAHs (polycyclic aromatic hydrocarbons).
Figure 2.
Schematic representation of the fate of contaminants. In green are shown plant-dependent processes; in pink, physical processes. Contaminants are deposited on surfaces (plants and soil) where they can be adsorbed. Parts of these contaminants are absorbed and then enter the plant structures. Contaminants can also be translocated from the roots to the shoots. Wind and evaporation can also mobilize contaminants from soil to leaves. BTEX (benzene, toluene, ethylbenzene and xylenes); PM2.5 (fine particulate matter); PCBs (polychlorinated biphenyls) and PAHs (polycyclic aromatic hydrocarbons).

Figure 3.
Schematic representation of bacterial activities in the rhizosphere and phyllosphere (a). ISR (induced systemic resistance); BTEX (benzene, toluene, ethylbenzene and xylenes); PM2.5 (fine particulate matter). The bioaugmentation of soil with Novosphingobium sp. HS2a, a bacterium able to mineralize phenanthrene (Phe), promotes clover growth on phenanthrene-contaminated soils (b) [106].
Figure 3.
Schematic representation of bacterial activities in the rhizosphere and phyllosphere (a). ISR (induced systemic resistance); BTEX (benzene, toluene, ethylbenzene and xylenes); PM2.5 (fine particulate matter). The bioaugmentation of soil with Novosphingobium sp. HS2a, a bacterium able to mineralize phenanthrene (Phe), promotes clover growth on phenanthrene-contaminated soils (b) [106].


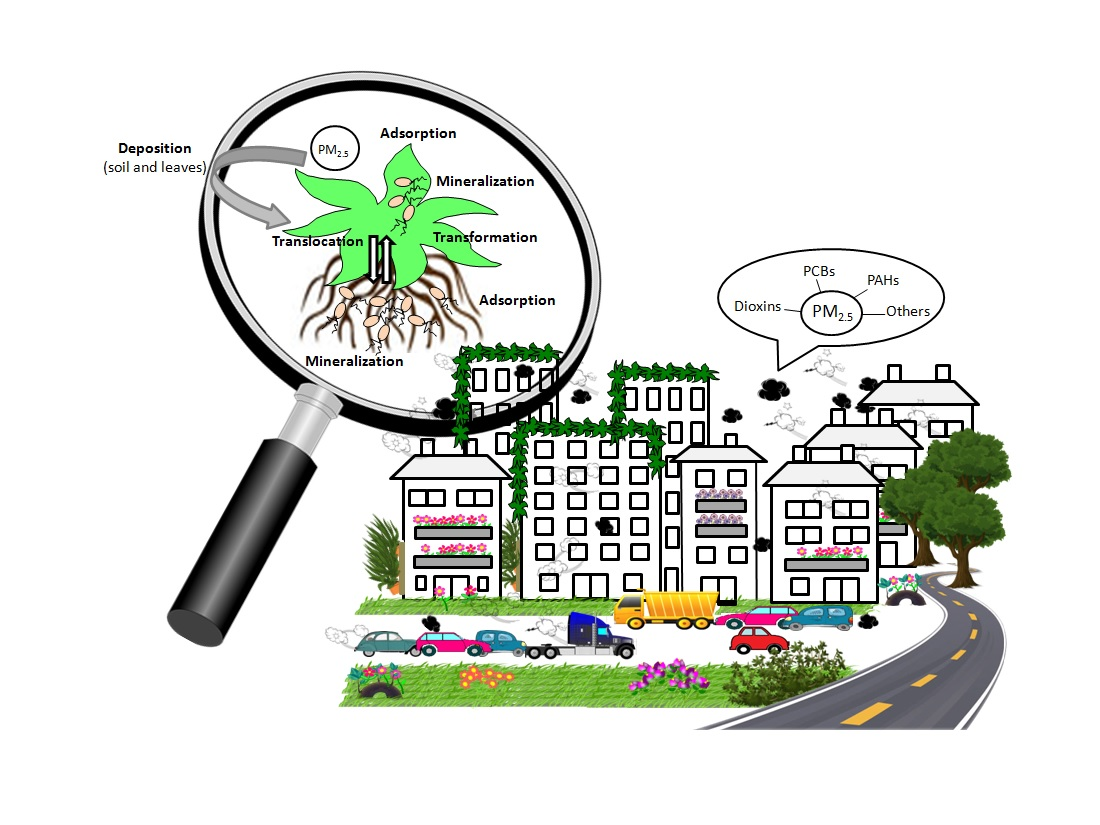{kind=link}
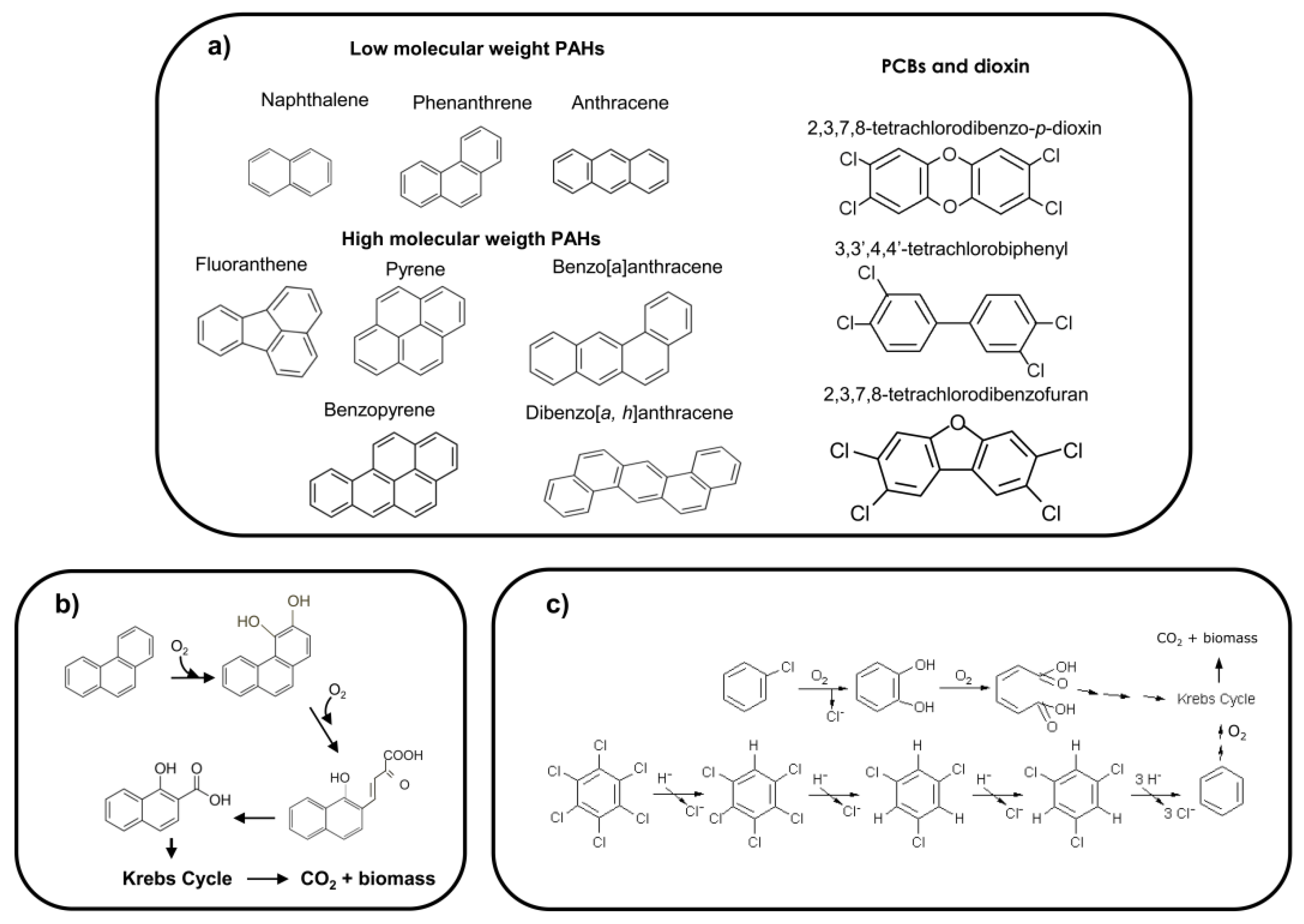{kind=link}
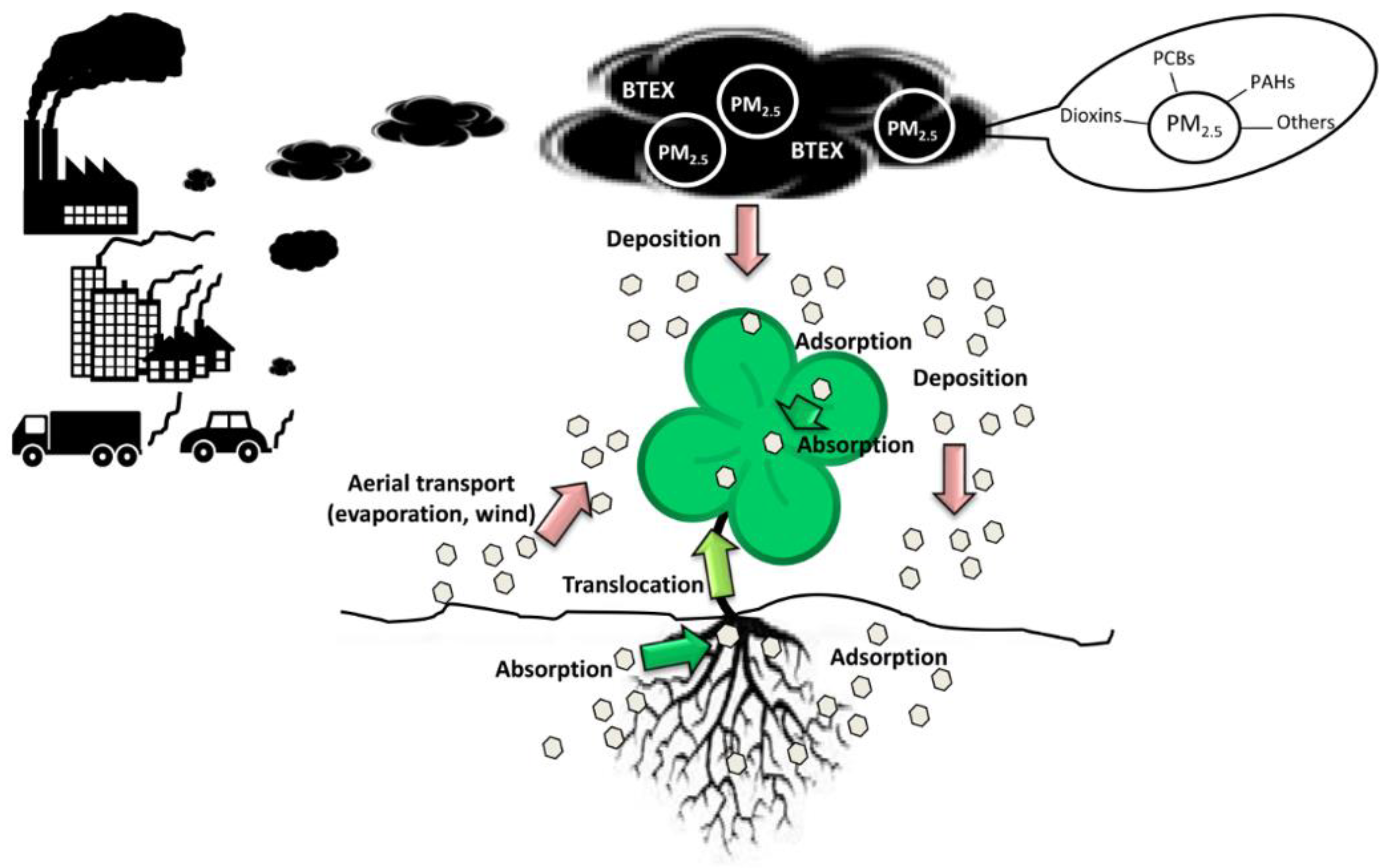{kind=link}
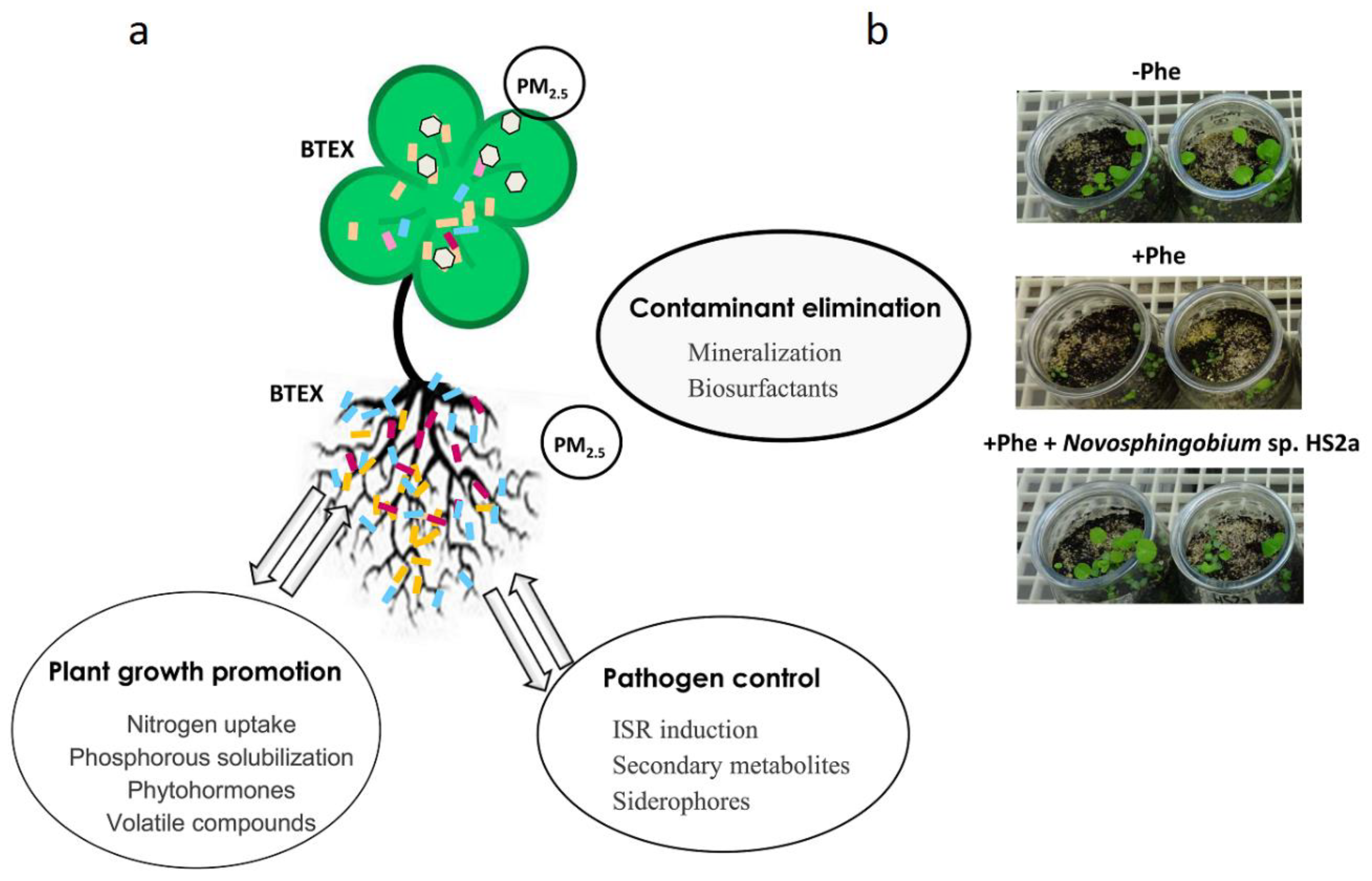{kind=link}
Table 1.
Examples of rhizoremediation of pollutants using plant-microbe pairs.
| Plants | Pollutants | Microbes | References |
|---|---|---|---|
| Senecus glaucus | Oil | Arthrobacter | [95] |
| Alfalfa | PAHs | Not identified | [132] |
| Barley | 2,4-D | Burkholderia cepacia | [133] |
| Oat, lupin, rape, pepper, radish, pine | Pyrene | Not identified | [134] |
| Poplar | 2,4-D | Amycolata sp. CB1190 | [135] |
| Lolium multiflorum | PAHs | Pseudmonas putida strain PCL1444 | [136] |
| Zinnia anguistifolia | Mefenoxam | Pseudomonas fluorescens Chrysobacterium indologenes | [137] |
| Hordeum vulgare | Phenanthrene | Pseudomonas fluorescens, Pseudomonas aureofaciens | [138] |
| Trifolium repens | Chrysene | Rhizobium leguminosarum | [139] |
| Triticum aestivum | Phenanthrene | Pseudomonas sp. strain GF3 | [140] |
| Hordeum vulgare | PAHs | Mycobacterium sps. | [141] |
| Sorghum bicolor | Phenanthrene | Sinorhizobium meliloti strain P221 | [142] |
| Lolium multiflorum | Diesel oil | Rhodococcus sp. strain ITRH43 | [143] |
| Secale cereale, medicago sativa | Crude oil | Azospirillum brasilense strain SR80 | [144] |
| Lotus corniculatus | Diesel oil | Pantoea sp. strain BTRH79 | [145] |
| Zea mays | Phenanthrene, pyrene | Pseudomonas sp. strain UG14Lr, Pseudomonas putida strain MUB1 | [146] |
| Lolium multiflorum | PAHs | Acinetobacter sp. | [147] |
| Medicago sativa | PAHs | Rhizobium meliloti strain ACCC 17519 | [148] |
| Zea mays | Diesel oil | Gordonia sp. strain S2RP-17 | [149] |
| Lolium perenne | Diesel oil | Pantoea sp. strain BTRH79 | [150] |
| Festuca | PAHs and diesel | Azospirillum sp. and Pseudomonas stutzeri | [151] |
| Populus deltoides | PAHs | Kurthia sp. Micrococcus sp. Bacillus sp. Dienococcus sp. Endophytic Bacillus sp. | [152] |
| Salix purpurea | Phenanthrene | Pseudomonas putida PD1 | [153] |
| Sorghum and onobrychis sativa | Phenanthrene, pyrene | Bacterial consortium | [154] |
| Annual grasses | Monoaromatics, PAHs | Pseudomonas putida strains | [155] |
| Clover | Phenanthrene | Novosphingobium sp. HS2a | [106] |
| Lolium multiflorum | PAHs | Mycobacterium gilvum | [156] |
| Avicennia schaueriana | Oil | Bacterial consortium | [157] |
| Arabidopsis thaliana | PCBs | Rhodococcus | [121] |
| Echinochloa crus-galli, cynodon dactylon | Monoaromatics | Pseudomonas sp. J10 | [158] |
| Chromolaena odorata | Lead, petroleum | Micrococcus luteus. | [159] |
| Grases | PAHs | Mycobacterium vanbaalenii PYR-1 | [160] |
| Morus alba | Biphenyl | Rhodococcus sp. MAPN-1 | [161] |
| Poplar | 1,4-Dioxane | Mycobacterium dioxanotrophicus PH-06, Pseudonocardia dioxanivorans CB1190 | [162] |
| Fesctuca arundinecea l. | PAHs | Mycobacterium sp. | [163] |
Table 2.
Phylloremediation of pollutants using plant—microbe pairs.
| Plants | Pollutants | Microbes | References |
|---|---|---|---|
| Azalea indica | Toluene | Pseudomonas putida TVA8 | [196] |
| Bean | Phenol | Pseudomonas sp. strain CF600 | [188] |
| Ixora sp | Phenanthrene | Pseudomonas oleovorans Mycobacterium sps. Rhizobium sps. Deinococcus sp. | [192] |
| Ornamental plants | Phenanthrene | Indigenous population | [194] |
| Beans and peas | Hydrocarbon vapors | Not identified | [190] |
| Bougainvillea buttiana | Xylene | Enterobacter cloacae LSRC11, Staphylococus sp. A1, Pseudomonas aeruginosa | [193] |
| Urban trees | Phenanthrene, benzo[a]pyrene | Non identified | [197] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Molina, L.; Wittich, R.-M.; van Dillewijn, P.; Segura, A. Plant-Bacteria Interactions for the Elimination of Atmospheric Contaminants in Cities. Agronomy 2021, 11, 493. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030493
AMA Style
Molina L, Wittich R-M, van Dillewijn P, Segura A. Plant-Bacteria Interactions for the Elimination of Atmospheric Contaminants in Cities. Agronomy. 2021; 11(3):493. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030493
Chicago/Turabian StyleMolina, Lázaro, Regina-Michaela Wittich, Pieter van Dillewijn, and Ana Segura. 2021. "Plant-Bacteria Interactions for the Elimination of Atmospheric Contaminants in Cities" Agronomy 11, no. 3: 493. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030493
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.