
Sunflower Metabolites Involved in Resistance Mechanisms against Broomrape



Allelopathy Group, Department of Organic Chemistry, Institute of Biomolecules (INBIO), Campus de Excelencia Internacional (ceiA3), School of Science, University of Cadiz, C/República Saharaui nº 7, 11510 Puerto Real, Spain
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(3), 501; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030501
Submission received: 30 December 2020
/
Revised: 24 February 2021
/
Accepted: 3 March 2021
/
Published: 8 March 2021
(This article belongs to the Special Issue Sunflower for the 21st Century – Omics Era and Changing Environment)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Several strategies have been proposed to control parasitic plants and these include biological control, the use of herbicides, and suicidal germination or trap crops. In the case of sunflower, the most effective solution is genetic resistance. Nevertheless, the use of resistant cultivars is followed by the appearance of new and more virulent races of the parasite that overcome the existing sources of resistance. For this reason, it is necessary to increase our knowledge of the mechanisms involved in the resistant varieties. In this study, the presence of compounds involved in pre-attachment resistance mechanisms in sunflower varieties was analyzed. The production of stimulators of the germination of parasitic plant seeds and the concentration of the haustorial elongation inhibitor, scopoletin, were measured in exudates of resistant and susceptible sunflower varieties. It was found that dehydrocostuslactone and orobanchyl acetate are not involved in the resistance of the sunflower varieties studied. The results demonstrate that costunolide, heliolactone, and scopoletin are related to sunflower resistance. The sunflower varieties studied do not use all available options to fight parasitic plants. This could be one of the reasons that sunflower resistance is rapidly overcome by broomrape.
1. Introduction
Sunflower (Helianthus annuus L.) is one of the most important crops in many regions of the world, such as Ukraine, Russia, Argentina, and Romania, because it is one of the major sources of vegetable oil [1]. Sunflowers are one of the higher adaptation crops because their deep roots prevent erosion and allow them to survive drought and other unsuitable conditions, growing as they do in hot semiarid climatic regions [2]. However, the emergence of Orobanche cumana Wallr. (broomrape) in the field reduces the growth and productivity of sunflower crops because it competes for nutritional resources, damages the crop, and reduces the yield by up to 50% [3]. Orobanche species are root parasitic plants and holoparasites that depend entirely on the host for their supply of water and nutrients [4]. Their optimum germination temperature is between 15 and 25 °C. Therefore, most broomrapes are located in the temperate and warm regions of the northern hemisphere, particularly the Mediterranean region and the Middle East. They can also be found in a similar climate in Australia, California, and Cuba [5].
The life cycle of broomrape starts with a chemical signal from its host that induces the germination of the seeds. The seeds can survive in the soil for around 20 years prior to receiving the signal to recognize their host [6,7,8]. This strategy ensures the presence of a host in the vicinity of the seeds, because once the seed germinates, it only has a few days to find a host before the nutrients in the seed run out. The most important stimulators of the germination of parasitic seeds are strigolactones (SLs) [9,10,11,12]. However, other compounds have been found to stimulate the germination of parasitic plant seeds. For example, sunflower sesquiterpene lactones (STLs), such as dehydrocostuslactone or costunolide, are able to stimulate the germination of O. cumana seeds [13,14]. Once the seeds have germinated, they connect with their host through an organ called the haustorium, through which they take the nutrients needed to survive. The parasitic plant then grows and flowers, and each plant produces thousands of seeds, which infect the soil again [9]. This life cycle, the tiny size of the seeds, and the high quantity of seeds produced by each parasitic plant, make it very difficult to manage the infection [15].
Several strategies have been proposed to control parasitic plants and these include biological control, the use of herbicides, and suicidal germination or trap crops. However, in the case of sunflower, the most effective, feasible, economical, and environmentally friendly solution is genetic resistance [16]. Nevertheless, the use of resistant cultivars, which are usually monogenic in nature, is followed by the appearance of new and more virulent races of the parasite that overcome the existing sources of resistance [17,18]. In 1980, Vrânceanu et al. described five resistant races of sunflower broomrape and these were denoted as A–E [19]. In the 1990s, race F appeared in the south of Spain [3]. Sunflower populations that were resistant to race F broomrape were also found in 2002 [20]. However, new broomrape populations, named races G and H [21,22,23], that overcome race F resistance sunflower genes are currently present in Eastern countries such as Turkey, Romania, Bulgaria, and Russia [19]. The speed with which these races of parasitic plants overcame the sunflower resistance is a result of the existence of a single mechanism. The success of genetic resistance will arise with multicomponent mechanisms at different stages of the infection. The resistance mechanism must avoid some of the steps in the life cycle of parasitic plants and these are classified as follows: pre-attachment (reduction in the seed germination or the haustorium development), pre-haustorial (avoiding the penetration of the haustorium in the roots), and post-haustorial (once the haustorium establishes vascular connections, the host kills the parasite) [24].
Regarding the pre-attachment mechanisms, phytoalexins have been proposed as compounds involved in the resistance mechanisms of sunflower. Coumarins, such as scopoletin, are synthesized by sunflower under biotic stress. In this respect, Wegman et al. found that the quantity of scopoletin in resistant sunflower was double that in susceptible sunflower [25], and Gutiérrez-Mellado et al. found that ayapin and scopoletin inhibit the germination of broomrape seeds, previously stimulated by GR24 (synthetic SL commonly used as stimulator of parasitic plant germination in bioassays), and produce necrosis in the germinated seeds as a pre-attachment mechanism [26]. Moreover, accumulation and high levels of excretion of these coumarins have been described in resistant sunflower in response to broomrape infection [27].
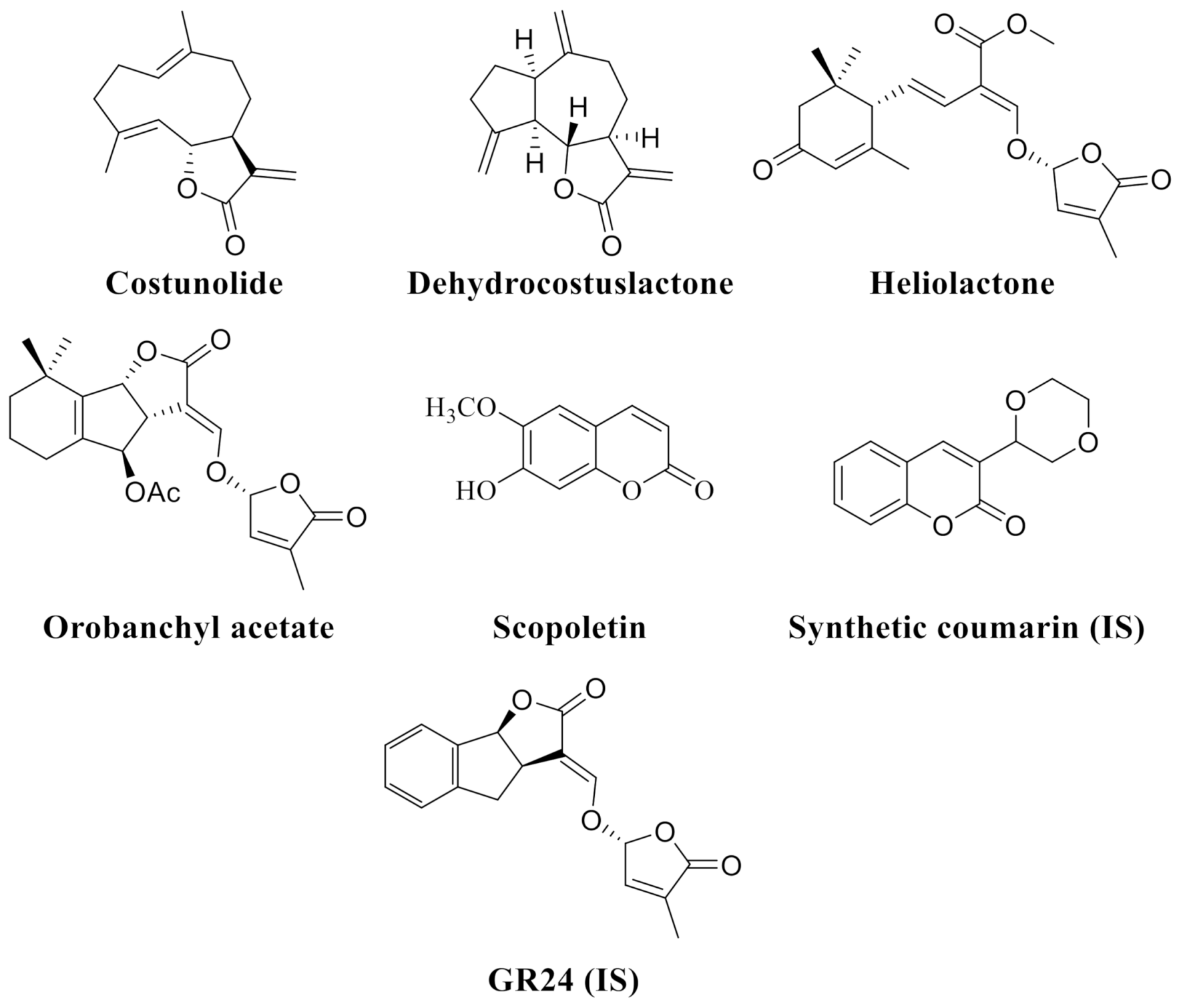
In addition to the above, a reduction in the levels of germination stimulants in the exudates could also help to decrease germination and the infestation ratio. Several resistant species of some host plants, such as vetches, peas, chickpeas, and sunflowers, have shown low induction of germination [28,29,30]. In addition, plants with low germination stimulant production have been used to successfully breed sorghum resistance to Striga [31]. Among the isolated compounds described in sunflower exudates that stimulate the germination of Orobanche and Phelipanche seeds are the following: costunolide [32] and dehydrocostuslactone [13], two STLs, and heliolactone, a non-canonical SL [33]. Moreover, Yoneyama et al. employed LC-MS/MS to identify the SLs 5-deoxystrigol and orobanchyl acetate in sunflower exudates (Figure 1) [10].
Thus, it is necessary to increase our knowledge of the mechanism involved in the resistance of new varieties and to identify new mechanisms. The aim of this study was to analyze the presence of compounds involved in pre-attachment resistance mechanisms in resistant sunflower varieties. With this aim in mind, the production of stimulators of the germination of parasitic plant seeds was evaluated and the concentrations of the haustorial inhibitor, scopoletin, were measured by comparing resistant and susceptible sunflower varieties.
2. Materials and Methods
2.1. Plant Material and Chemicals
Sunflower seeds from susceptible varieties B117 and NR5 and resistant varieties BR4, P96, BR3, K96, and R96 were generously provided by Dr. Leonardo Velasco (Institute for Sustainable Agriculture (CSIC), Cordoba, Spain) [34,35]. Organic solvents were UHPLC (ultra high performance liquid chromatography) grade and were purchased from Fischer Chemicals (Geel, Belgium). Water was type I obtained from an Ultramatic system from Wasserlab (Barbatáin, Spain). Costunolide and dehydrocostuslactone were isolated from Saussurea lappa [36]. Scopoletin was purchased from Sigma-Aldrich (Steinheim, Germany). The synthetic coumarin was kindly provided by Dr. Francisco Javier Moreno-Dorado and Dr. Francisco Guerra-Martínez (Faculty of Sciences, University of Cadiz) [37].
2.2. Hydroponic Culture of Sunflower and Root Exudate Collection
Sunflower root exudates were obtained from 10 fresh plants (three biological repetitions per cultivar) using the procedure previously described by López-Ráez and co-workers [38]. Seeds were sterilized in an ultrasonic bath for 10 min using a sodium hypochlorite solution (0.1% v/v). The seeds were then rinsed with distilled water and germinated in Petri dishes (55 mm diameter and five seeds per dish) with moistened filter paper in darkness for 4 days at 27 °C. Germinated seeds were transferred into pots (6.5 × 6.5 × 9 cm) filled with a mixture of equal parts of vermiculite, sand, and clay. Plants were grown under 16/8 h day/night photoperiods in a climate chamber at 27 °C and 60% humidity. The plants were rinsed twice a week, once with half-strength Hoagland solution and once with distilled water.
As previously described in the literature [33,38,39], phosphate starvation was applied 7 weeks after sowing to promote the production of germination stimulants. Exudates were collected 1 week later by rinsing the pots with 100 mL of distilled water. The exudate was filtered through Whatman No.1 filter paper and purified using reverse phase solid phase extraction cartridges (StrataTM-X 33 µm Polymeric Reversed Phase 200 mg/3 mL, Phenomenex, Torrance, CA, USA). Firstly, cartridges were conditioned with 3 mL of MeOH, then, they were equilibrated with 6 mL of type I water. Next, the exudates were loaded into the cartridges. Regarding the washing step, 3 mL of type I water and 3 mL of acetone:water 40:60 were used to elute the impurities. Analytes were eluted with 6 mL of acetone:water 60:40, dried, and re-dissolved in MeOH to achieve a concentration of 1 mg/mL. Both of the internal standards, namely, GR24 and the synthetic coumarin, were added to achieve a final concentration of 10 µg L–1 for each of the samples. The samples were then filtered through a syringe filter (polytetrafluoroethylene (PTFE) 0.22 µm) and stored at −80 °C.
2.3. LC-MS/MS Analysis
The exudates were analyzed on an EVOQ Triple Quadrupole Mass Spectrometer from Bruker (Billerica, MA, USA) with an electrospray-ionization source (ESI) in positive mode following the procedures previously described by our research group [40,41] to analyze seven SLs (7-oxoorobanchyl acetate [42], solanacol [43], orobanchol [44], strigol [45,46], fabacyl acetate [47], orobanchyl acetate [48], 5-deoxystrigol [49]) and two STLs (costunolide and dehydrocostuslactone) (Table S1).
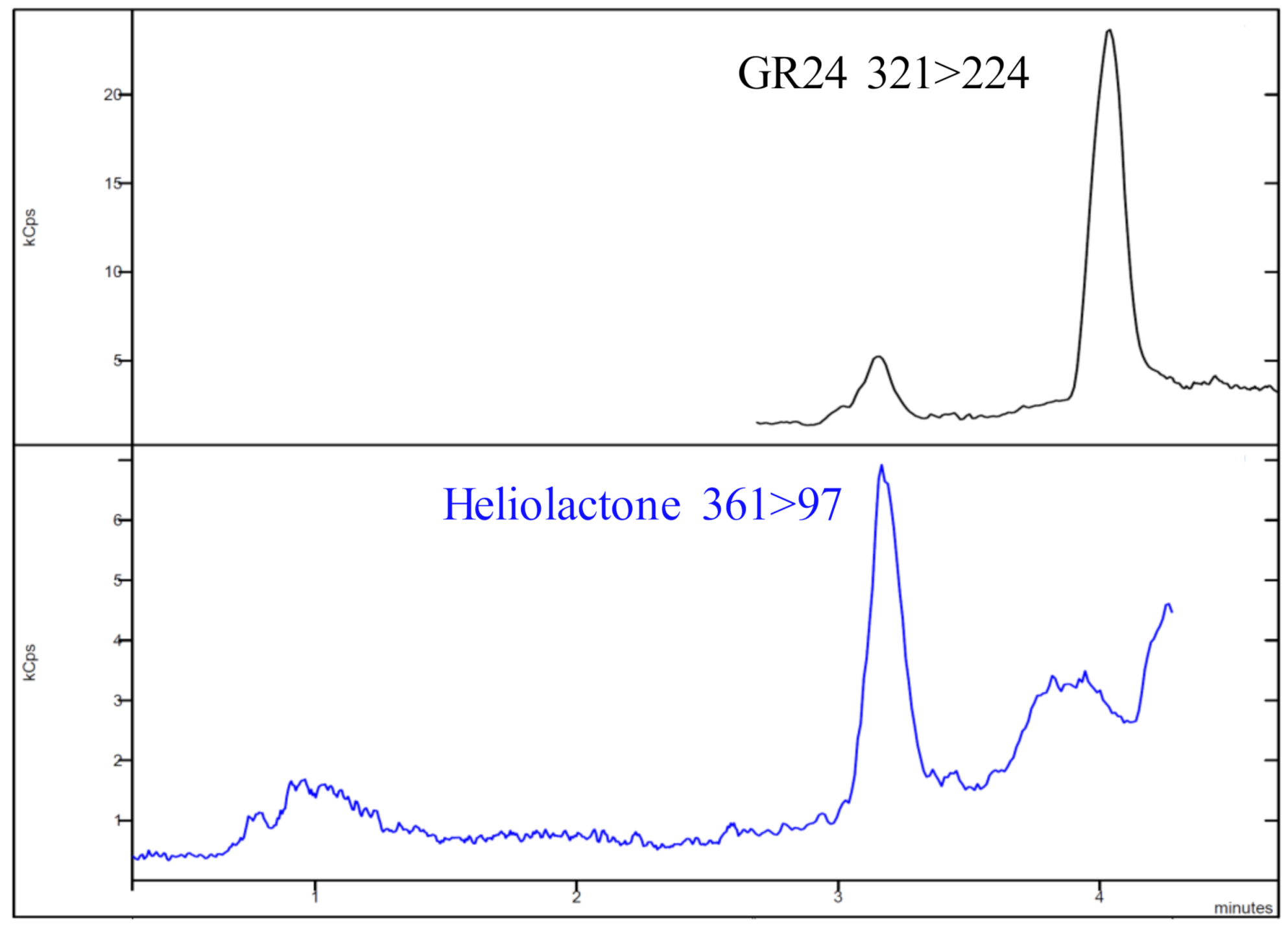
Heliolactone was determined in relative mode using GR24 to compare the concentration present in the sunflower varieties, as an analytical standard is not commercially available. The UHPLC-MS/MS (ultra high performance liquid chromatography tandem mass spectrometry) method described above was used for the determination of heliolactone using the most stable fragments previously reported in the literature. The precursor ion was m/z 361 and the quantifier and qualifier products were m/z 97 (collision energy (CE, collision energy) of 20 eV) and m/z 233 (CE of 17 eV), respectively [33]. Only one peak was observed for sunflower root exudates (at 3.18 min) and the collision energy of each transition was optimized to obtain the highest signal for this species.
The coumarin scopoletin was analyzed by injecting the samples into the same LC-MS/MS described above and species were separated using a Kinetex 1.7 µm C18 100 Å column (100 × 2.1 mm) (Phenomenex, Torrance, CA, USA) maintained at 40 °C. The mobile phase consisted of solvent A (water, 0.1% formic acid) and solvent B (MeOH, 0.1% formic acid), and the flow rate was set to 0.4 mL/min. The optimized linear gradient system was as follows: 0–1 min, 50% B; 1–2 min, to 80% B; 2–3 min, 80% B; 3–5 min, 100% B; 5–7, 100% B.
The synthetic coumarin was used as internal standard and was added to all samples to give a final concentration of 10 µg L−1. This coumarin was selected because it has a similar chemical structure to scopoletin and because it is not present in the sunflower exudate. The compound-dependent parameters were optimized by direct infusion on the mass spectrometer to achieve maximum multiple reaction monitoring (MRM) signal intensities using argon as the collision gas. For scopoletin (retention time (tR) 2.04 min), the precursor ion was m/z 193 and the quantifier and qualifier products were m/z 133 (collision energy (CE) of 22 eV) and m/z 178 (CE of 26 eV), respectively. For synthetic coumarin (tR 3.71 min), the precursor ion was m/z 233 and the quantifier and qualifier products were m/z 189 (CE of 15 eV) and m/z 133 (CE of 29 eV), respectively. Calibration curves were also prepared from 100 to 5 µg·L−1 (6 levels) by following the previously described procedure [40].
The levels of the internal standards were optimized to obtain values similar to those found in the samples. The peak area ratio (y) of each analyzed compound to the corresponding internal standard was plotted versus the ratio of their concentrations (x). The resulting square coefficients of regression were higher than 0.99 for all the standards. The peak area ratio with respect to the internal standard and by reference to the standard curve was used to determine the levels in the samples. The equation of the calibration curves and the resulting limits of detection (LOD), quantification (LOQ), repeatability, and intermediate precision are shown in Table S2.
Samples were prepared following the previously described procedure [40]. A stock standard solution of 10 mg L–1 was prepared in MeOH for all standards and the internal standard. The external standard calibration curves were prepared by serial dilution of the working standard solution. All standards and stock solutions were filtered through a polytetrafluoroethylene (PTFE) syringe filter (0.22 µm) prior to analysis and samples were stored at −80 °C. The autosampler was set at 5 °C to preserve the samples. The injection volume was 5 μL. The instrument parameters were as follows: spray voltage +4500 V, cone temperature 300 °C, cone gas flow 15 psi, heated probe temperature 400 °C, heated probe gas flow 15 psi, nebulizer gas flow 55 psi, and collision pressure 2.0 mTorr.
The non-canonical SL heliolactone was determined in relative mode using GR24 to compare the concentrations present in the sunflower varieties. To determine heliolactone, the most stable fragments previously reported in the literature were used [33]. Only one peak was observed for sunflower root exudates (at 3.18 min, Figure 2), and the collision energy of each transition was optimized to obtain the highest signal.
Scopoletin was identified in the exudates of all varieties studied previously (Figure 3) and determined in sunflower root by UHPLC-MS/MS with electrospray in positive mode as the ionization source and using a synthetic coumarin (Figure 1) as the internal standard.
Data acquisition, calibration curves, and statistical analysis of the data from the determination were performed with the software MS Data Review (Bruker Chemical Analysis, Billerica, MA, USA). Significant differences were determined based on the Analysis of Variance (one-way ANOVA) followed by Newman–Keuls test at a probability of 95%.
2.4. Cluster Analysis
Cluster analyses were performed using Statistica v.7.0 software (Statistica 7.0, Tulsa, OK, USA). Euclidian distances and single linkage were used for the analysis.
3. Results
3.1. Stimulators of Parasitic Plant Germination
In an effort to ascertain whether a pre-attachment mechanism is involved in the resistance mechanism of resistant sunflower varieties, the concentrations of germination stimulants and inhibitors of haustorial elongation present in the root exudates were measured. For this purpose, five sunflower varieties that were resistant to O. cumana race F (K96, BR4, BR3, R96, and P96) were studied and compared with two susceptible sunflower varieties (B117 and NR5).
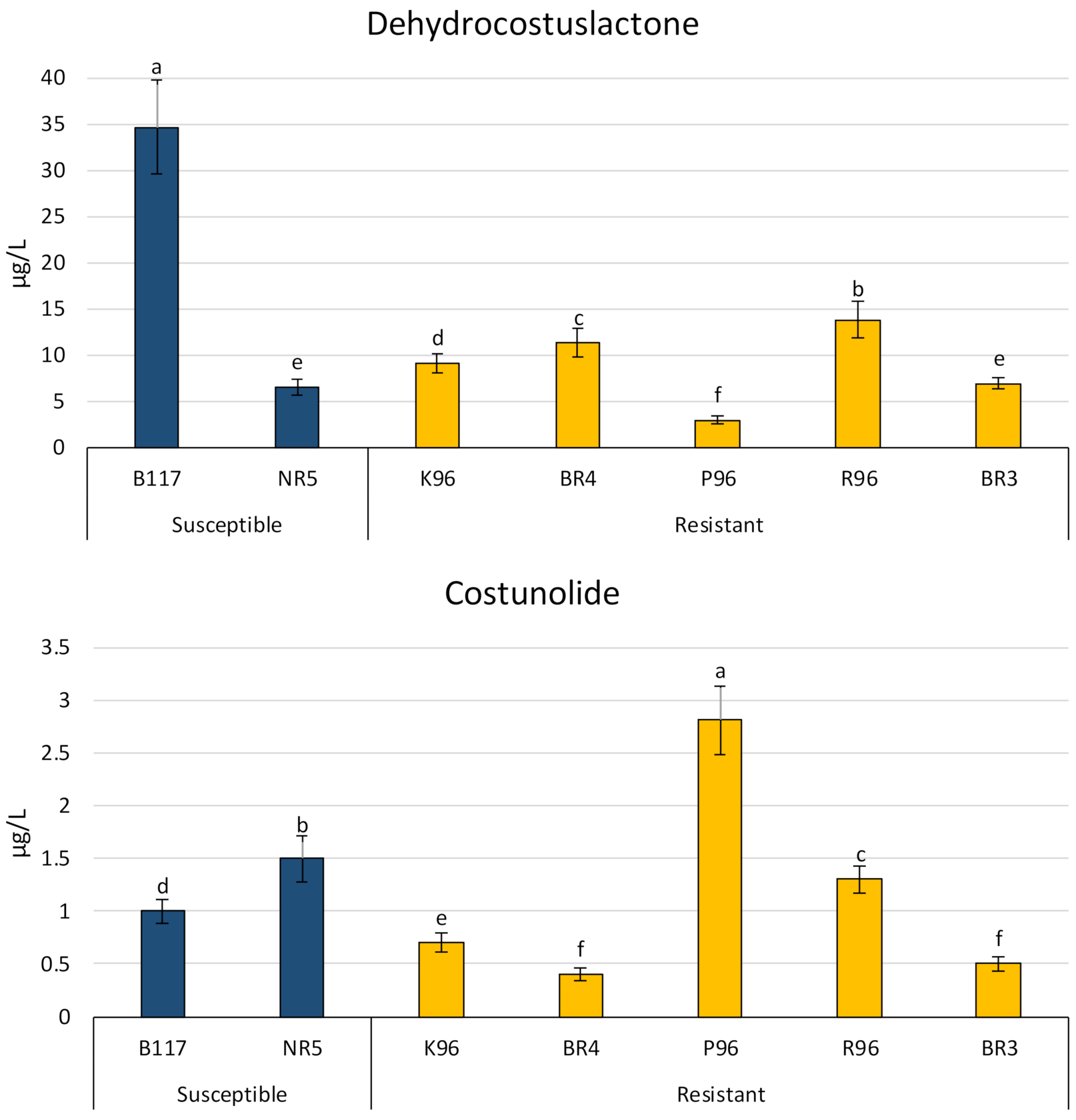
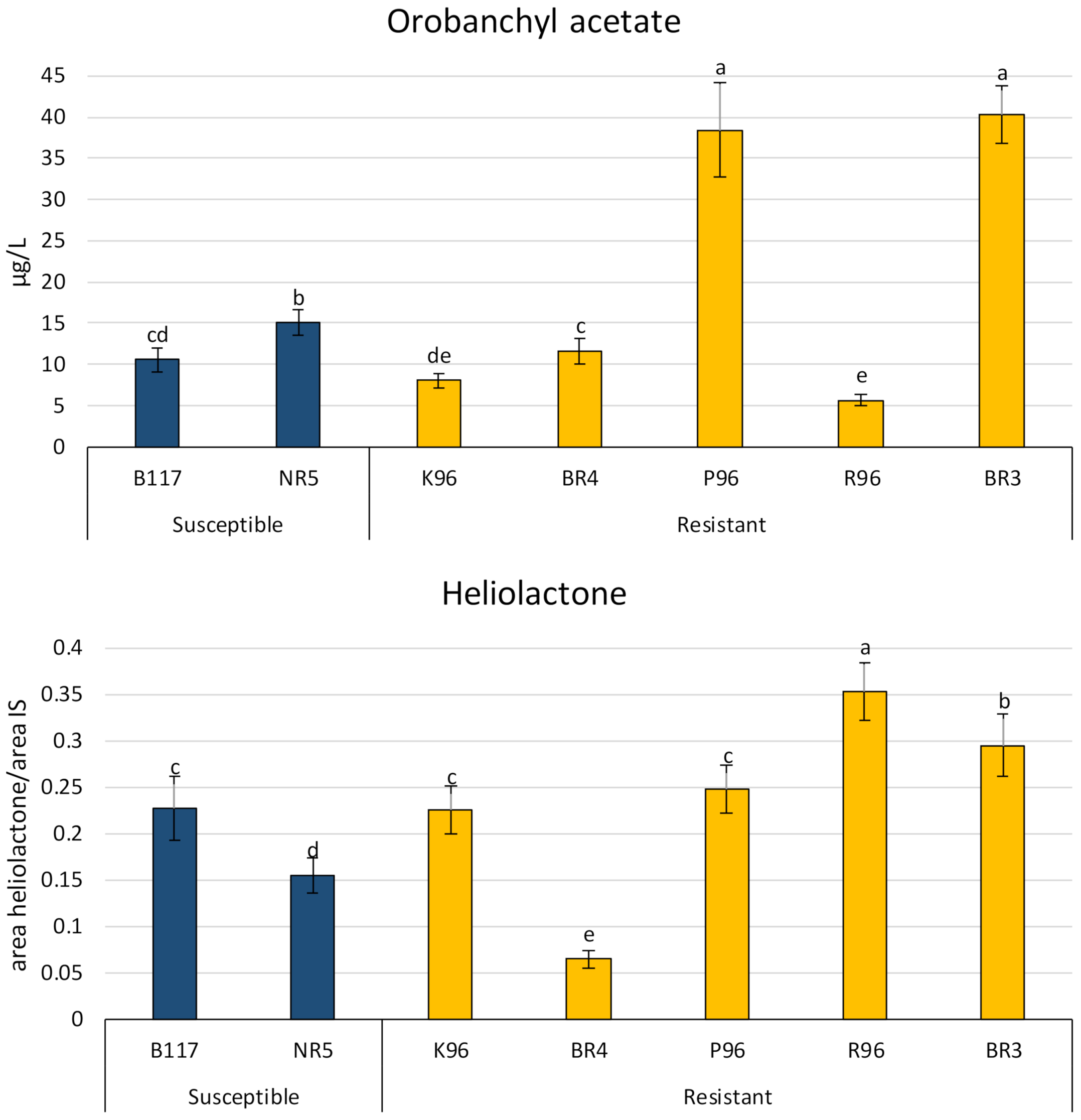
The STLs costunolide and dehydrocostuslactone, and the SLs heliolactone and orobanchyl acetate were identified in all varieties. The results are represented in Figure 4 and Figure 5.
Dehydrocostuslactone was the major STL identified in sunflower exudate, with a concentration of 30 µg L–1 in some varieties and more than 5 µg L–1 in all cases, except for P96. The highest level of dehydrocostuslactone was found in one of the susceptible varieties, namely, B117. This variety contained a concentration (34.7 µg L−1) that was almost three times higher than the highest one found in the resistant varieties (R96: 13.8 µg L−1). However, the other susceptible variety (NR5) showed a much lower level (6.5 µg L−1), i.e., a similar level to those in most of the resistant varieties.
In the case of costunolide, the highest concentration was found in the variety P96 (2.8 µg L−1). P96 was the only variety with values higher than 2.5 µg L−1 for costunolide, with the other varieties ranging from 0.4 µg L−1 (resistant BR4) to 1.5 µg L−1 (susceptible NR3).
Orobanchyl acetate was the major compound identified in the resistant varieties P96 and BR3, with concentrations of 38.4 and 40.0 µg L−1, respectively. The concentrations of orobanchyl acetate in the susceptible varieties ranged from 10 to 15 µg L−1. The remaining resistant varieties ranged from 5.6 to 11.8 µg L−1, i.e., slightly lower than the susceptible varieties.
Heliolactone was determined in relative mode using GR24 to compare the concentrations present in the sunflower varieties, as an analytical standard is not commercially available. The results of the analysis are shown in Figure 5 and are expressed as the area of heliolactone/area of GR24. Heliolactone was present at higher levels in all the resistant varieties than in the susceptible ones, except for the BR4 variety. The values for the susceptible varieties ranged from 0.15 to 0.22 µg L−1 and from 0.07 to 0.35 µg L−1 (area values relative to the internal standard (IS)) for the resistant ones.
3.2. Inhibitors of Haustorial Elongation
As mentioned above, coumarins, such as scopoletin, can inhibit the haustorial elongation of broomrape. Additionally, these compounds produce necrosis of germinated seeds, which is a pre-attached mechanism [26,27].
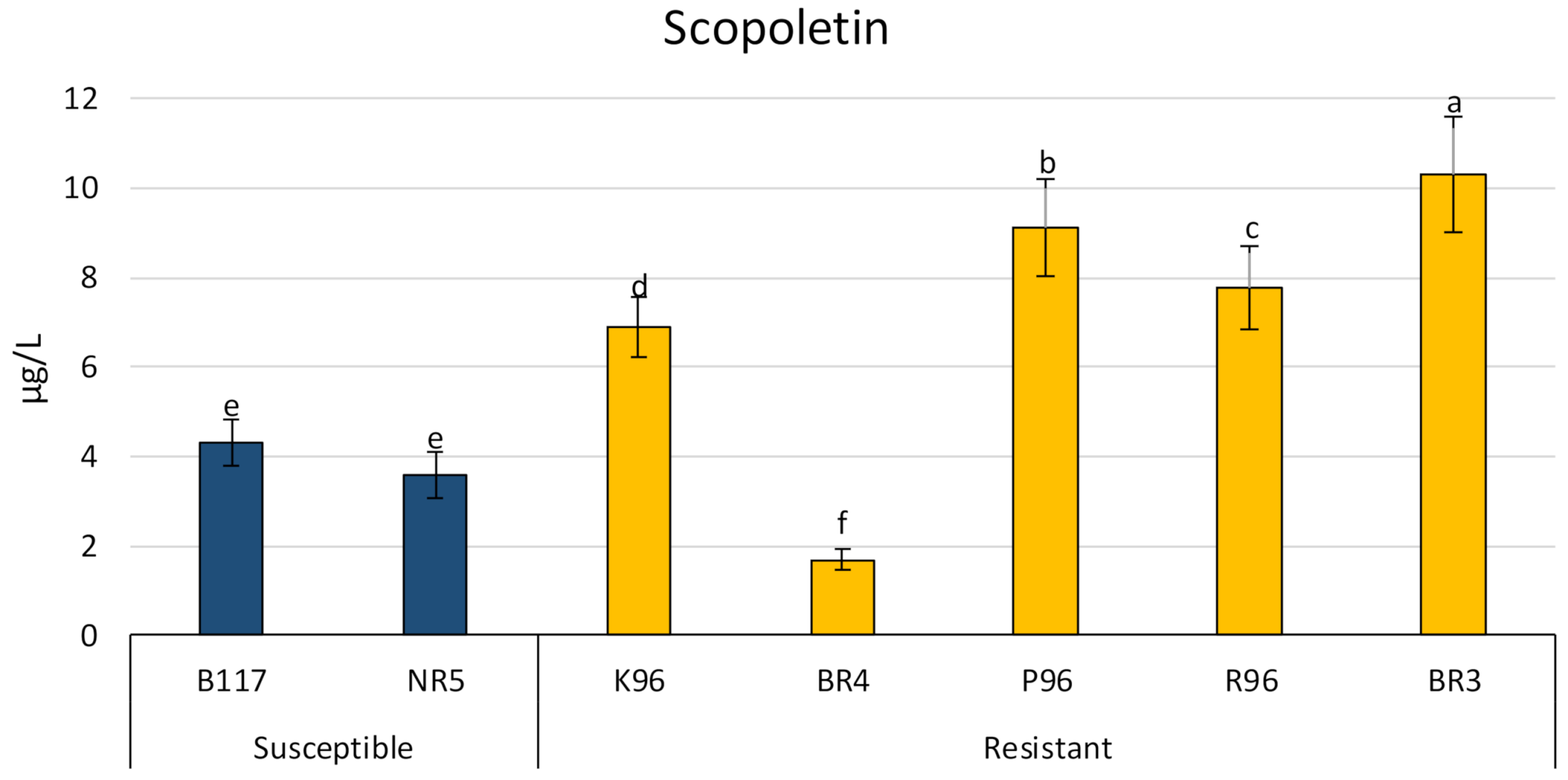
Scopoletin (Figure 6) showed similar behavior to heliolactone for both types of sunflower varieties (resistant and susceptible). All resistant varieties, except for BR4, showed higher levels of scopoletin than the susceptible ones, with values that were double those found in most cases. The values found for the susceptible varieties ranged from 3.60 to 4.32 µg L−1 and the resistant ones (not including BR4) ranged from 6.90 to 10.32 µg L−1.
3.3. Cluster Analysis
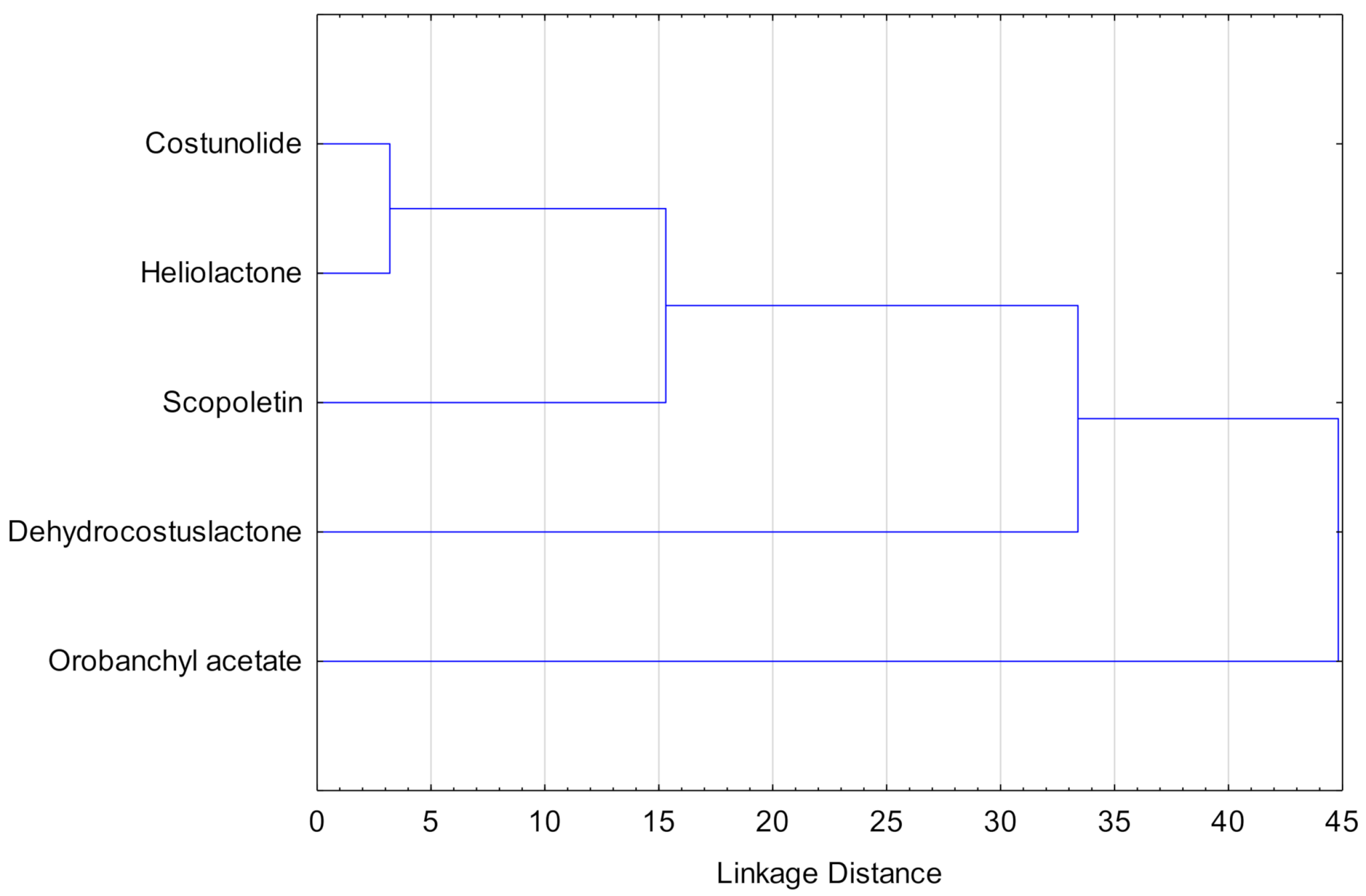
The study involved several varieties and several compounds per variety and as a consequence, a cluster analysis was performed to identify those compounds related to the resistant/susceptible properties of the varieties. The resulting dendrogram for compounds is shown in Figure 7. It can be seen that both costunolide and heliolactone were strongly related and that scopoletin is related to both of them. However, dehydrocostuslactone and orobanchyl acetate were located far from that cluster.
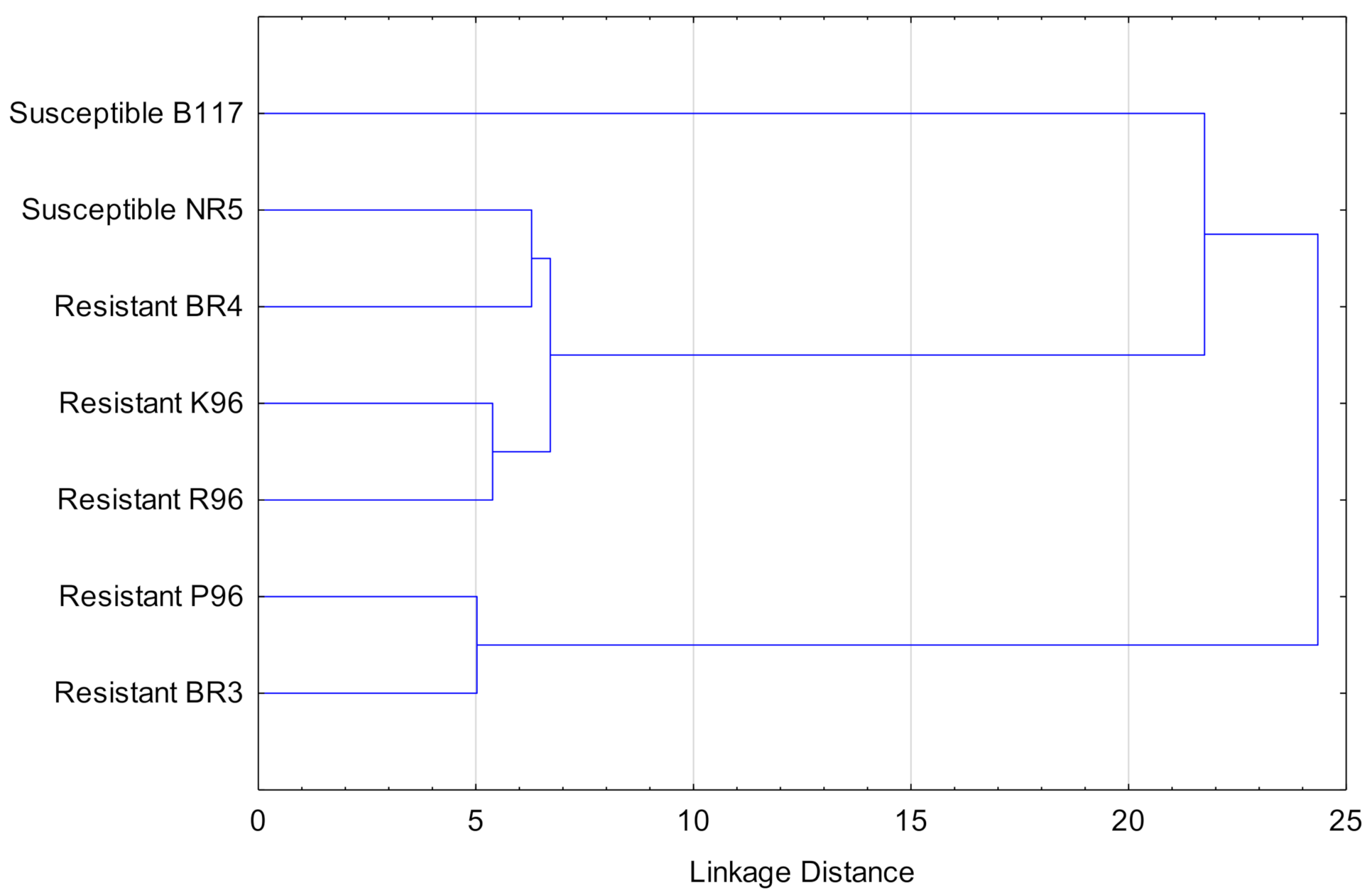
The resulting dendrogram for the different varieties is shown in Figure 8. It can be seen that resistant and susceptible varieties were not in the same cluster but they were allocated in heterogeneous clusters.
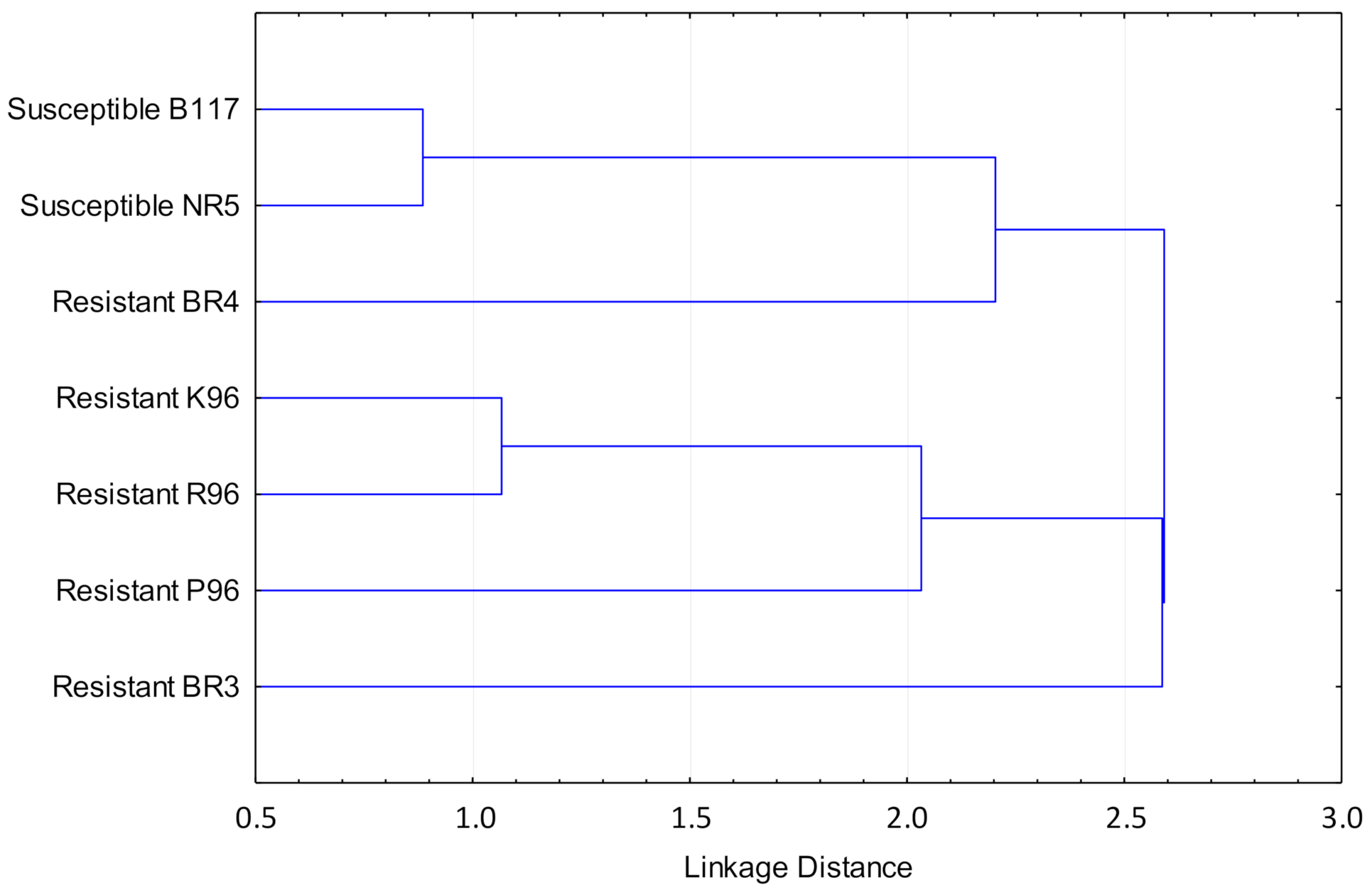
Given the results shown in Figure 8, an additional cluster analysis was performed using only the compounds that form a homogeneous cluster in the dendrogram in Figure 7, i.e., costunolide, heliolactone, and scopoletin. The resulting dendrogram is shown in Figure 9. In the dendrogram, the susceptible varieties form an isolated cluster from the resistant ones. It must be noted that resistant variety BR4 was not included in the cluster for resistant varieties, but in an intermediate situation. The cluster analysis performed based on the remaining compounds, i.e., orobanchyl acetate and dehydrocostuslactone, did not produce any specific cluster (results not shown).
4. Discussion
4.1. Stimulators of Parasitic Plant Germination
Firstly, the exuded stimulators of parasitic plant germination were studied. The STLs costunolide and dehydrocostuslactone and the SLs heliolactone and orobanchyl acetate were identified in all varieties.
The concentrations of these biocommunicators did not clearly decrease in the resistant varieties when compared with the susceptible varieties and they were even higher in some cases. For example, in susceptible varieties, dehydrocostuslactone gave the highest value (B117) and one of the lowest (NR5). A similar situation was found for orobanchyl acetate. The levels of this compound ranged from 10.5 to 15.0 µg L−1 for the susceptible varieties and from 5.6 to 40.0 µg L−1 for the resistant ones. Additionally, it can be concluded from the cluster analyses that the values for dehydrocostuslactone and orobanchyl acetate are not useful for grouping susceptible varieties vs. resistant ones. It can therefore be concluded that these compounds are not involved in the resistance mechanism of the resistant sunflower varieties studied.
Heliolactone is one of the compounds that allowed the varieties to be grouped in the cluster analysis in Figure 9. For most resistant varieties, the levels of the exuded heliolactone were higher than those produced by the susceptible varieties. Therefore, it can be concluded that a reduction in the production of heliolactone as a stimulator of the germination is not one of the mechanisms for sunflower plants to prevent the development of broomrape. However, the resistant variety BR4 exuded two and three times less heliolactone than the susceptible varieties NR5 and B117, respectively. This means that BR4 could reduce the percentage of germination, and consequently the infestation, to protect against broomrape.
Costunolide is an STL that has also been described as contributing to the germination of parasitic plants. Costunolide was found at lower levels in the resistant varieties than in the susceptible ones, with only the resistant variety P96 showing a much higher value (around double) than the susceptible varieties. Therefore, resistant varieties could be using costunolide to decrease the germination of broomrape seeds. However, there is one variety (P96) that produces much more costunolide than the other resistant varieties and even more than the susceptible ones. This finding indicates that P96 employs a different mechanism than other resistant varieties to prevent the development of broomrape.
Thus, it can be deduced that there are different behaviors in the resistant varieties to prevent the development of broomrape and that not all available strategies are employed.
4.2. Inhibitors of Haustorial Elongation
As discussed above, coumarins inhibit the haustorial elongation of broomrape and produce necrosis of the germinated seeds as a pre-attachment mechanism [26,27]. The coumarin scopoletin was isolated from sunflower roots [16] and it could be involved in the resistance mechanism of sunflower to inhibit the germination of broomrape.
Scopoletin was found in all sunflower varieties. Moreover, all resistant varieties, except for BR4, contained higher amounts of scopoletin in their exudates than the susceptible varieties, with double the amount present in some cases. These results suggest that resistant varieties K96, P96, R96, and BR3 could employ scopoletin as an inhibitor of haustorial elongation. As the levels found for BR4 were lower than those from susceptible varieties, one can conclude BR4 does not use this mechanism to inhibit the haustorial elongation. Although K96, P96, R96, and BR3 produced some compounds that stimulate broomrape seed germination, it should be noted that they also promote the synthesis of the haustorial elongation inhibitor, scopoletin.
In contrast to the above, BR4 uses the compounds related to the activation of germination rather than the inhibition of haustorial elongation used by the other varieties.
4.3. Cluster Analysis
Using the cluster analysis in Figure 7, it could be inferred that compounds from the same natural products family are not related because they are not grouped in the same cluster, i.e., STLs and SLs did not form specific clusters.
On the other hand, regarding cluster analysis in Figure 8 and using all available compounds, a discrimination between the varieties based on their properties against broomrape cannot be expected. However, using only the compounds that form a homogeneous cluster in the dendrogram in Figure 7, i.e., costunolide, heliolactone, and scopoletin, the susceptible varieties form an isolated cluster from the resistant ones (Figure 9). Therefore, these three compounds can be identified as responsible for the differences between the susceptible and the resistant varieties.
Therefore, these analyses indicate that resistant varieties are able to produce the chemical compounds that prevent the development of broomrape, but all of these compounds are not used collectively by each variety.
5. Conclusions
The results obtained confirm that the exuded concentrations of the STL dehydrocostuslactone and the canonical SL orobanchyl acetate are not involved in the resistance of the sunflower varieties studied. However, the STL costunolide, the SL heliolactone, and the coumarin scopoletin can be used by the resistant varieties to fight the parasitic plants. As far as the defense mechanisms are concerned, some varieties can produce compounds related to the resistance by reducing the germination or inhibiting the haustorial elongation. However, the sunflower varieties studied do not use all available options to fight the parasitic plants. This could be one of the reasons that sunflower resistance is rapidly overcome by broomrape [17,18].
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2073-4395/11/3/501/s1, Table S1: Mass pair (m/z) and compound-dependent parameters of the standards, Table S2: Analytical characteristics of the chromatographic methods.
Author Contributions
Conceptualization, C.R. and R.M.V.; Methodology, C.R. and R.M.V.; Validation, C.R., R.M.V. and J.M.G.M.; Formal Analysis, C.R., R.M.V. and J.M.G.M.; Investigation, C.R., R.M.V. and S.P.; Resources, J.M.G.M. and F.A.M.; Data Curation, C.R. and R.M.V.; Writing—Original Draft Preparation, C.R.; Writing—Review & Editing, R.M.V., J.M.G.M. and F.A.M.; Visualization, R.M.V., J.M.G.M. and F.A.M.; Supervision, R.M.V., J.M.G.M. and F.A.M.; Project Administration, J.M.G.M. and F.A.M.; Funding Acquisition, J.M.G.M. and F.A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministerio de Economía, Industria y Competitividad (MINEICO) Spain, grant number AGL2017-88083-R.
Acknowledgments
The authors thank Leonardo Velasco (Institute of Sustainable Agriculture (CSIC), Córdoba, Spain) for supplying sunflower seeds. Moreover, we would like thank Miguel Palma from the University of Cadiz for his comments and help with the manuscript and Francisco Javier Moreno-Dorado from the University of Cadiz for supplying the synthetic coumarin 6.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Food and Agriculture Organization of the United Nations FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 6 January 2019).
- Kaya, Y. Sunflower. In Breeding Oilseed Crops for Sustainable Production; Gupta, S.K., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 55–88. [Google Scholar]
- Echevarría-Zomeño, S.; Pérez-De-Luque, A.; Jorrín, J.; Maldonado, A.M. Pre-haustorial resistance to broomrape (Orobanche cumana) in sunflower (Helianthus annuus): Cytochemical studies. J. Exp. Bot. 2006, 57, 4189–4200. [Google Scholar] [CrossRef]
- Heide-Jørgensen, H.S. The Parasitic syndrome in higher plants. In Parasitic Orobanchaceae—Parasitic Mechanisms and Control Strategies; Joel, D.M., Gressel, J., Musselman, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–18. [Google Scholar]
- Labrousse, P.; Delmail, D. Sunflower resistance to the vampire weed broomrape: A Van Helsing quest story. In Oilseed Crops: Yield and Adaptations under Environmental Stress; Parvaiz, A., Ed.; John Wiley: Hoboken, NJ, USA, 2017; pp. 123–151. ISBN 9781119048770. [Google Scholar]
- Joel, D.M.; Gressel, J.; Musselman, L.J. Parasitic Orobanchaceae. Parasitic Mechanisms and Control Strategies; Springer: Berlin/Heidelberg, Germany, 2012; ISBN 9783642381454. [Google Scholar]
- Brown, R.; Greenwood, A.D.; Johnson, A.W.; Long, A.G. The stimulant involved in the germination of Orobanche minor Sm. Chromatographic purification of crude concentrates. Biochem. J. 1951, 48, 564–568. [Google Scholar] [CrossRef] [Green Version]
- Yoneyama, K.; Awad, A.A.; Xie, X.; Takeuchi, Y. Strigolactones as germination stimulants for root parasitic plants. Plant Cell Physiol. 2010, 51, 1095–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Yoneyama, K.K.; Yoneyama, K.K. The strigolactone story. Annu. Rev. Phytopathol. 2010, 48, 93–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoneyama, K.K.; Xie, X.; Kisugi, T.; Nomura, T.; Sekimoto, H.; Yokota, T.; Yoneyama, K.K. Characterization of strigolactones exuded by Asteraceae plants. Plant Growth Regul. 2011, 65, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Yoneyama, K.; Xie, X.; Yoneyama, K.; Takeuchi, Y. Strigolactones: Structures and biological activities. Pest Manag. Sci. 2009, 65, 467–470. [Google Scholar] [CrossRef]
- Yoneyama, K.K.K.; Xie, X.; Yoneyama, K.K.K.; Kisugi, T.; Nomura, T.; Nakatani, Y.; Akiyama, K.; McErlean, C.S.P. Which are the major players, canonical or non-canonical strigolactones? J. Exp. Bot. 2018, 69, 2231–2239. [Google Scholar] [CrossRef]
- Raupp, F.M.; Spring, O. New sesquiterpene lactones from sunflower root exudate as germination stimulants for Orobanche cumana. J. Agric. Food Chem. 2013, 61, 10481–10487. [Google Scholar] [CrossRef]
- Cala, A.; Molinillo, J.M.G.; Fernández-Aparicio, M.; Ayuso, J.; Álvarez, J.A.; Rubiales, D.; Macías, F.A. Complexation of sesquiterpene lactones with cyclodextrins: Synthesis and effects on their activities on parasitic weeds. Org. Biomol. Chem. 2017, 15, 6500–6510. [Google Scholar] [CrossRef]
- Rubiales, D.; Verkleij, J.; Vurro, M.; Murdoch, A.J.; Joel, D.M. Parasitic plant management in sustainable agriculture. Weed Res. 2009, 49, 1–5. [Google Scholar] [CrossRef]
- Rodriguez-Ojeda, M.I. Estudios Genéticos y de Biología Reproductiva en Orobanche cumana Wallr; University of Cordoba: Andalusia, Spain, 2016. [Google Scholar]
- Velasco, L.; Pérez-Vich, B.; Yassein, A.A.M.; Jan, C.-C.; Fernández-Martínez, J.M. Inheritance of resistance to sunflower broomrape (Orobanche cumana Wallr.) in an interspecific cross between Helianthus annuus and Helianthus debilis subsp. tardiflorus. Plant Breed. 2012, 131, 220–221. [Google Scholar] [CrossRef] [Green Version]
- Hladni, N.; Jocic, S.; Miklic, V.; Saftic-Pankovic, D.; Skoric, D. Using new Rf inbred lines originating from an interspecific population with H. deserticola for development of sunflower hybrids resistant to broomrape. Helia 2009, 32, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Akhtouch, B.; del Moral, L.; Leon, A.; Velasco, L.; Fernández-Martínez, J.M.; Pérez-Vich, B. Genetic study of recessive broomrape resistance in sunflower. Euphytica 2016, 209, 419–428. [Google Scholar] [CrossRef]
- Akhtouch, B.; Muñoz-Cuz, J.; Melero-Vara, J.; Fernandez-Martinez, J.; Dominguez, J. Inheritance of resistance to race F of broomrape in sunflower lines of different origins. Plant Breed. 2002, 121, 266–268. [Google Scholar] [CrossRef]
- Shindrova, P.; Penchev, E. Race composition and distribution of broomrape (Orobanche cumana Wallr.) in Bulgaria during 2007–2011. Helia 2012, 57, 87–94. [Google Scholar] [CrossRef]
- Antonova, T.S.; Araslanova, N.M.; Strelnikov, E.A.; Ramazanova, S.A.; Guchetl, S.Z.; Chelyustnikova, T.A. Distribution of highly virulent races of sunflower broomrape (Orobanche cumana Wallr.) in the southern regions of the Russian Federation. Russ. Agric. Sci. 2013, 39, 46–50. [Google Scholar] [CrossRef]
- Kaya, Y.; Evci, G.; Pekcan, V.; Gucer, T.; Yilmaz, M.I. Evaluation of broomrape resistance in sunflower hybrids. Helia 2009, 51, 161–170. [Google Scholar] [CrossRef]
- Pérez-de-Luque, A.; Moreno, M.T.T.; Rubiales, D. Host plant resistance against broomrapes (Orobanche spp.): Defence reactions and mechanisms of resistance. Ann. Appl. Biol. 2008, 152, 131–141. [Google Scholar] [CrossRef]
- Wegmann, K.; Von Elert, E.; Harloff, H.J.; Stadler, M. Tolerance and resistance to Orobanche. In Progress in Orobanche Research, Proceedings International Workshop on Orobanche Research; Wegmann, K., Musselman, L.J., Eds.; Eberhard-Karls University: Tübingen, Germany, 1991; pp. 318–321. [Google Scholar]
- Gutiérrez-Mellado, C.; Parry, A.; Tena, M.; Jorrin, J.; Edwards, R. Abiotic elicitation of coumarin phytoalexins in sunflower. Phytochem. Resour. Med. Agric. 1995, 38, 1185–1191. [Google Scholar] [CrossRef]
- Serghini, K.; de Luque, A.P.; Castejón-Muñoz, M.; García-Torres, L.; Jorrín, J.V.; Pérez De Luque, A.; Castejón-Muñoz, M.; García-Torres, L.; Jorrín, J.V.; de Luque, A.P.; et al. Sunflower (Helianthus annuus L.) response to broomrape (Orobanche cernua Loefl.) parasitism: Induced synthesis and excretion of 7-hydroxylated simple coumarins. J. Exp. Bot. 2001, 52, 2227–2234. [Google Scholar] [CrossRef]
- Labrousse, P.; Arnaud, M.C.; Serieys, H.; Bervillé, A.; Thalouarn, P. Several mechanisms are involved in resistance of Helianthus to Orobanche cumana Wallr. Ann. Bot. 2001, 88, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Pérez-de-Luque, A.; Joel, D.M.; Alcántara, C.; Sillero, J.C. Characterization of resistance in chickpea to crenate broomrape (Orobanche crenata). Weed Sci. 2003, 51, 702–707. [Google Scholar] [CrossRef]
- Perez-de-Luque, A.; Jorri, J.N.; Cubero, J.I.; Rubiales, D. Orobanche crenata resistance and avoidance in pea ( Pisum spp.) operate at different developmental stages of the parasite. Weed Res. 2005, 45, 379–387. [Google Scholar] [CrossRef]
- Haussmann, B.I.G.; Hess, D.E.; Welz, H.G.; Geiger, H.H. Improved methodologies for breeding striga-resistant sorghums. Field Crop. Res. 2000, 66, 195–211. [Google Scholar] [CrossRef] [Green Version]
- Joel, D.M.; Chaudhuri, S.K.; Plakhine, D.; Ziadna, H.; Steffens, J.C. Dehydrocostus lactone is exuded from sunflower roots and stimulates germination of the root parasite Orobanche cumana. Phytochemistry 2011, 72, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Furumoto, T.; Umeda, S.; Mizutani, M.; Takikawa, H.; Batchvarova, R.; Sugimoto, Y. Heliolactone, a non-sesquiterpene lactone germination stimulant for root parasitic weeds from sunflower. Phytochemistry 2014, 108, 122–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Martínez, J.; Pérez-Vich, B.; Akhtouch, B.; Velasco, L.; Muñoz-Ruz, J.; Melero-Vara, J.M.; Domínguez, J. Registration of four sunflower germplasms resistant to race F of broomrape. Crop Sci. 2004, 44, 1033–1034. [Google Scholar] [CrossRef]
- Jan, C.C.; Fernández-Martínez, J.M.; Ruso, J.; Muñoz-Ruz, J. Registration of four sunflower germplasms with resistance to Orobanche cumana race F. Crop Sci. 2002, 42, 2217–2218. [Google Scholar] [CrossRef]
- Cala, A.; Zorrilla, J.G.; Rial, C.; Molinillo, J.M.G.; Varela, R.M.; Macías, F.A. Easy access to alkoxy, amino, carbamoyl, hydroxy, and thiol derivatives of sesquiterpene lactones and evaluation of their bioactivity on parasitic weeds. J. Agric. Food Chem. 2019, 67, 10764–10773. [Google Scholar] [CrossRef]
- Niu, B.; Zhao, W.; Ding, Y.; Bian, Z.; Pittman, C.U.; Zhou, A.; Ge, H. Regioselective cross-couplings of coumarins and flavones with ethers via C(sp 3 )–H functionalization. J. Org. Chem. 2015, 80, 7251–7257. [Google Scholar] [CrossRef]
- López-Ráez, J.A.; Charnikhova, T.; Gómez-Roldán, V.; Matusova, R.; Kohlen, W.; De Vos, R.; Verstappen, F.; Puech-Pages, V.; Bécard, G.; Mulder, P.; et al. Tomato strigolactones are derived from carotenoids and their biosynthesis is promoted by phosphate starvation. New Phytol. 2008, 178, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, K.; Xie, X.; Kim, H., II; Kisugi, T.; Nomura, T.; Sekimoto, H.; Yokota, T.; Yoneyama, K. How do nitrogen and phosphorus deficiencies affect strigolactone production and exudation? Planta 2012, 235, 1197–1207. [Google Scholar] [CrossRef] [Green Version]
- Rial, C.; Varela, R.M.; Molinillo, J.M.G.; López-Ráez, J.A.; Macías, F.A. A new UHPLC-MS/MS method for the direct determination of strigolactones in root exudates and extracts. Phytochem. Anal. 2019, 30, 110–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rial, C.; Tomé, S.; Varela, R.M.; Molinillo, J.M.G.; Macías, F.A. Phytochemical study of safflower roots (Carthamus tinctorius) on the induction of parasitic plant germination and weed control. J. Chem. Ecol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Yoneyama, K.K.K.; Kurita, J.; Harada, Y.; Yamada, Y.; Takeuchi, Y.; Yoneyama, K.K.K. 7-Oxoorobanchyl acetate and 7-Oxoorobanchol as germination stimulants for root parasitic plants from flax (Linum usitatissimum). Biosci. Biotechnol. Biochem. 2009, 73, 1367–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Kusumoto, D.; Takeuchi, Y.; Yoneyama, K.K.; Yamada, Y.; Yoneyama, K.K. 2′-Epi-orobanchol and solanacol, two unique strigolactones, germination stimulants for root parasitic weeds, produced by tobacco. J. Agric. Food Chem. 2007, 55, 8067–8072. [Google Scholar] [CrossRef]
- Yokota, T.; Sakai, H.; Okuno, K.; Yoneyama, K.; Takeuchi, Y. Alectrol and orobanchol, germination stimulants for Orobanche minor, from its host red clover. Phytochemistry 1998, 49, 1967–1973. [Google Scholar] [CrossRef]
- Cook, C.E.; Whichard, L.P.; Turner, B.; Wall, M.E.; Egley, G.H. Germination of witchweed (Striga lutea Lour.): Isolation and properties of a potent stimulant. Science 1966, 154, 1189–1190. [Google Scholar] [CrossRef]
- Cook, C.; Whichard, L.; Wall, M.; Egley, G. Germination stimulants. II. Structure of strigol, a potent seed germination stimulant for witchweed (Striga lutea). J. Am. Chem. Soc. 1972, 94, 6198–6199. [Google Scholar] [CrossRef]
- Xie, X.; Yoneyama, K.; Harada, Y.; Fusegi, N.; Yamada, Y.; Ito, S.; Yokota, T.; Takeuchi, Y.; Yoneyama, K. Fabacyl acetate, a germination stimulant for root parasitic plants from Pisum sativum. Phytochemistry 2009, 70, 211–215. [Google Scholar] [CrossRef]
- Xie, X.; Yoneyama, K.; Kusumoto, D.; Yamada, Y.; Yokota, T.; Takeuchi, Y.; Yoneyama, K. Isolation and identification of alectrol as (+)-orobanchyl acetate, a germination stimulant for root parasitic plants. Phytochemistry 2008, 69, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Matsuzaki, K.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 2005, 435, 824–827. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structures of costunolide, dehydrocostuslactone, heliolactone, orobanchyl acetate, scopoletin, the synthetic coumarin used as the internal standard (IS) and GR24 (synthetic SL used as internal standard).
Figure 1.
Structures of costunolide, dehydrocostuslactone, heliolactone, orobanchyl acetate, scopoletin, the synthetic coumarin used as the internal standard (IS) and GR24 (synthetic SL used as internal standard).

Figure 2.
Chromatogram of the sunflower exudate for heliolactone.

Figure 3.
Chromatogram of sunflower exudate, scopoletin, and the IS.

Figure 4.
Concentrations of dehydrocostuslactone and costunolide in sunflower root exudates of five resistant varieties (BR4, P96, BR3, K96, and R96) and two susceptible varieties (B117 and NR5). Different letters indicate significant difference (p < 0.05) according to Analysis of Variance (one-way ANOVA) followed by Newman–Keuls test. Similar letters indicate non-significant difference.
Figure 4.
Concentrations of dehydrocostuslactone and costunolide in sunflower root exudates of five resistant varieties (BR4, P96, BR3, K96, and R96) and two susceptible varieties (B117 and NR5). Different letters indicate significant difference (p < 0.05) according to Analysis of Variance (one-way ANOVA) followed by Newman–Keuls test. Similar letters indicate non-significant difference.

Figure 5.
Concentrations of orobanchyl acetate and heliolactone in sunflower root exudates of five resistant varieties (BR4, P96, BR3, K96, and R96) and two susceptible varieties (B117 and NR5). Different letters indicate significant difference (p < 0.05) according to one-way ANOVA followed by Newman–Keuls test. Similar letters indicate non-significant difference.
Figure 5.
Concentrations of orobanchyl acetate and heliolactone in sunflower root exudates of five resistant varieties (BR4, P96, BR3, K96, and R96) and two susceptible varieties (B117 and NR5). Different letters indicate significant difference (p < 0.05) according to one-way ANOVA followed by Newman–Keuls test. Similar letters indicate non-significant difference.

Figure 6.
Concentrations of scopoletin in sunflower root exudates of five resistant varieties (BR4, P96, BR3, K96, and R96) and two susceptible varieties (B117 and NR5). Different letters indicate significant difference (p < 0.05) according to one-way ANOVA followed by Newman–Keuls test. Similar letters indicate non-significant difference.
Figure 6.
Concentrations of scopoletin in sunflower root exudates of five resistant varieties (BR4, P96, BR3, K96, and R96) and two susceptible varieties (B117 and NR5). Different letters indicate significant difference (p < 0.05) according to one-way ANOVA followed by Newman–Keuls test. Similar letters indicate non-significant difference.

Figure 7.
Cluster analysis for the compounds based on their concentration in sunflower exudates of all varieties.
Figure 7.
Cluster analysis for the compounds based on their concentration in sunflower exudates of all varieties.

Figure 8.
Cluster analysis for the sunflower varieties studied based on the concentration of the analyzed compounds (costunolide, dehydrocostuslactone, heliolactone, orobanchyl acetate, and scopoletin).
Figure 8.
Cluster analysis for the sunflower varieties studied based on the concentration of the analyzed compounds (costunolide, dehydrocostuslactone, heliolactone, orobanchyl acetate, and scopoletin).

Figure 9.
Cluster analysis for the sunflower varieties studied based on the concentration of costunolide, heliolactone, and scopoletin.
Figure 9.
Cluster analysis for the sunflower varieties studied based on the concentration of costunolide, heliolactone, and scopoletin.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rial, C.; Varela, R.M.; Molinillo, J.M.G.; Peralta, S.; Macías, F.A. Sunflower Metabolites Involved in Resistance Mechanisms against Broomrape. Agronomy 2021, 11, 501. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030501
AMA Style
Rial C, Varela RM, Molinillo JMG, Peralta S, Macías FA. Sunflower Metabolites Involved in Resistance Mechanisms against Broomrape. Agronomy. 2021; 11(3):501. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030501
Chicago/Turabian StyleRial, Carlos, Rosa M. Varela, José M.G. Molinillo, Sara Peralta, and Francisco A. Macías. 2021. "Sunflower Metabolites Involved in Resistance Mechanisms against Broomrape" Agronomy 11, no. 3: 501. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030501
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.