
Agronomic Biofortification of Significant Cereal Crops with Selenium—A Review




1
Department of Agroecology and Environment Protection, Faculty of Agrobiotechnical Sciences Osijek, Josip Juraj Strossmayer University of Osijek, Vladimira Preloga 1, 31000 Osijek, Croatia
2
Department of Plant Production and Biotechnology, Faculty of Agrobiotechnical Sciences Osijek, Josip Juraj Strossmayer University of Osijek, Vladimira Preloga 1, 31000 Osijek, Croatia
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(5), 1015; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11051015
Submission received: 18 March 2021
/
Revised: 6 May 2021
/
Accepted: 19 May 2021
/
Published: 20 May 2021
(This article belongs to the Section Soil and Plant Nutrition)
Abstract
:Selenium (Se) is an important micronutrient which is essential for most living organisms and occurs in both organic and inorganic forms in the water system, soils, biomass, and the atmosphere. In addition to being essential for humans and animals, Se is beneficial for plants and is mostly involved in antioxidant activity/response, as well as a growth promoter. Se deficiency in the diet is a global problem, and Se levels in soils generally reflect its presence in food and, thus, availability to humans. Se participates in the antioxidant response mechanisms of the organism, heavy-metal detoxification, and regulation of the reproductive and immune system, as well as ensures the proper function of the thyroid gland. Plants are the main dietary source of Se for humans. Biofortification is a key strategy to increase Se in edible parts of plants. Agronomic biofortification provides an effective route to increase Se content in edible crop products via application of Se-enriched fertilizers to soil or by foliar application. The most common cereals in the human diet are wheat, rice, maize, and barley, making them the most suitable targets for agronomic biofortification. This review focuses on summarizing the most efficient form and method of Se application via agronomic biofortification corroborated by a meta-analysis of the literature reports. In the assessed literature, foliar application showed better results compared to application in soil. The selenate form appears to be the more efficient form of Se for biofortification than selenite in the most common cereals in human diet: wheat, rice, maize, and barley.
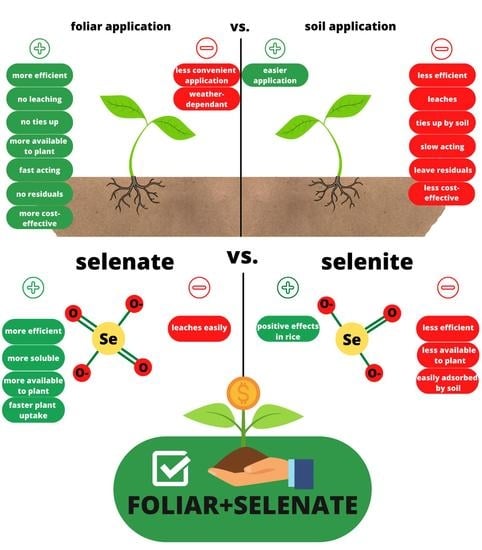
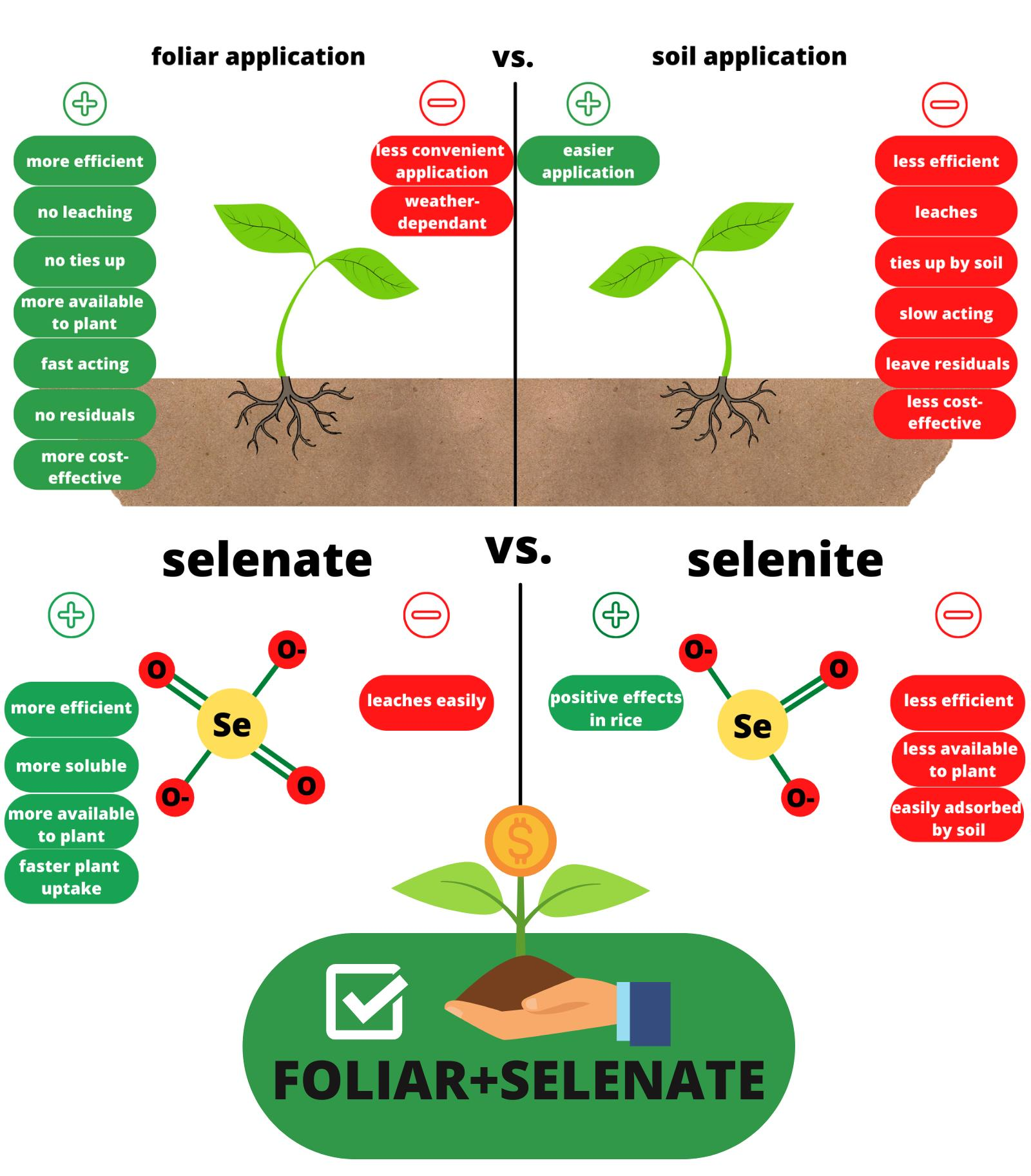
1. Introduction
Selenium (Se) is a nonmetal with atomic number 34 in group 16 [1]. The Se atom is slightly larger than sulfur (S) (the radius of Se2+ is 0.5 Å whereas the radius of S2+ is 0.37 Å), and, like S, Se can exist in five valence states, selenide (2−), elemental Se (0), thioselenate (2+), selenite (4+), and selenate (6+) [2,3]. Se is a mineral micronutrient essential for the adequate and healthy life of humans, animals, archaea, and some other microorganisms [4]. Se exists in the lithosphere layer of the earth crust, i.e. water, soil, and in open environments; therefore, its distribution differs throughout the globe [5,6]. Se levels in European soils are low, particularly in eastern Europe [6]. Plants absorb Se from soil primarily as selenate and translocate it to the chloroplasts, where it follows the sulfur assimilation pathway [7]. Se is essential for lower plants, but its essentiality in higher plants is still under investigation [8]. Se can have physiological benefits for plants, which may be related to its tendency to upregulate plant antioxidant metabolites and enzymes, leading to a better capacity to scavenge reactive oxygen species (ROS) that impede plant performance, especially under stress conditions [9].
Dietary Se deficiency has negative effects for human health, and more than 40 types of diseases have been associated with Se deficiency, such as Keshan disease, Kashin–Beck disease, some types of cancer, cardiovascular diseases, liver diseases, and cataracts [10]. Selenium deficiency occurs in several parts of the world, especially where Se concentration in soils is low, leading to poor concentration in agricultural plants [11]. The process of increasing the bioavailable concentrations of essential elements in edible parts of crop plants through agricultural intervention or genetic selection through breeding process is called biofortification [12]. Se biofortification substantially increases Se contents in agricultural food products and can help alleviate Se malnutrition, affecting more than one billion people worldwide [13]. Dietary deficiencies of Se can be improved through dietary diversification, food fortification, supplementation, or crop biofortification, including fertilization (agronomic biofortification) or crop improvement (genetic biofortification) as different approaches [14]. Cereal grains are grown and consumed by humans in higher quantities than any other crop and provide most of the food energy consumed worldwide [15], thus representing the most significant plant candidates suitable for biofortification with additional possible positive effects on plant yield [16], although differences in nutritional profiles and selenium content between wheat species were not observed [17].
Cereals are the most important source of Se in the Western diet with major representatives being rice, wheat, maize, and barley [18], while Se-enriched wheat has long been recognized as a source of supplemental Se for Se-deficient populations [7]. Many studies confirmed the positive effects of Se agronomic biofortification on Se content in rice, wheat, maize, and barley grains [16,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35]. Agronomic biofortification is based on applying fertilizers with mineral elements lacking in diet to increase their concentrations in crops through soil or foliar application [30]. The two most important factors in achieving the increased Se content in grains are the type of application and the form of Se, although systematic reports on these questions are still missing [36]. Plants uptake Se more readily when applied in a foliar manner, compared to soil application, with the additional benefit of having no residual effects in the soils [37]. The foliar technique includes minimal consumption of Se salts and represents the most effective, safe, and economically justified measure of improving Se content in agricultural crops [38]. The reason for the higher efficiency of foliar application is that there is no root-to-shoot translocation, accompanied by the finding that soils can act as considerable sinks for Se [39]. The form of applied Se is the second most important factor for effective agronomic biofortification [40], with the most commonly used forms of Se being selenate and selenite. However, nanoselenium (nSe) is being used more and more frequently [8]. The synthesis and use of nSe as a nutrient and biofortifier has been proven as an interesting strategy [40], although reports of wider use are still missing. Studies have shown selenate (Se(VI)) to be the most effective form of Se when applied to the soil and usually more effective compared to selenite (Se(IV)) when applied in a foliar manner [41]. Selenate is more efficient due to the more rapid uptake of Se (VI) and translocation from root to stem and leaves, as well as quicker transformation into its organic form, while selenite is more easily adsorbed to the soil, which makes its uptake by roots more difficult [42]. Selenite has characteristics similar to phosphate, corroborating its easy adsorption to the soil surfaces compared to selenite [43].
In the context of the Se biofortification complexity and differences in the methodology found in literature survey, the aim of this study was to review the present knowledge on the basis of studies dealing with Se agronomic biofortification as the most feasible way of increasing the Se content in the food, as well as to identify the best practices for accomplishing efficient Se biofortification. The former aim was supported by a meta-analysis of the data found in different research reports by using a linear mixed modeling approach. Research reports for meta-analysis were selected on the basis of consistency in methodology and availability of important data needed for adjustment in the linear mixed model. Generally, the methodology found in the research articles addressing Se biofortification was not harmonized and uniform.
2. Selenium in the Environment
Selenium (Se) is a nonmetal belonging to the same group of elements (group 16) as oxygen and sulfur (S). Se is rarely considered a metalloid due to its intermediate properties between a metal and a nonmetal. It is stable and does not oxidize at ordinary temperatures [2,3]. Naturally, it exists in five different oxidation forms in the environment, such as Se(VI) (selenate), Se(IV) (selenite), Se(0) (elemental Se), and Se(−II) (selenide), along with various organic species [1]. Many elements can be combined with Se, such as hydrogen, fluorine, chlorine, bromine, and phosphorus, and Se can form compounds analogous to sulfur [3]. The distribution of different species of Se may vary in the environment depending on the prevailing redox conditions. Principally, selenium oxyanions (SeO42− and SeO32−) are highly soluble, stable, and potentially mobile in oxic natural environments [44]. Se can be found in all components of the agroecosystem including soils, plants, rocks, and water [5,28]. The biogeochemical cycle of Se begins from weathering of Se-rich rocks, soils, and sediments, moving toward the different water bodies. Later on, from water, it arrives in plants, animals, or humans by various means. The Se cycle is completed by the degradation and different activities of organisms, which allow Se accessibility in the soil sediments and, ultimately, the rock depositaries [1]. Distribution processes of Se through the environment, thus, involve a variety of physical, chemical and biological activities [4]. The presence of Se is linked to natural activities such as soil erosion, volcanism, and forest fires, with the atmosphere playing an important role in the biogeochemical cycle of Se. The Se content in ambient air is mainly low, and it varies from 1 to 10 ng Se·m−3 [3], while coal and oil burning are the primary sources of considerable emissions of Se compounds in the air. In natural waters, the dissolved selenium concentrations are reported to be in the range of <0.1 to 100 µg·L−1 [44], and the average Se concentration in the Earth’s crust is around 0.05 mg·kg−1 [39].
2.1. Selenium in Soils
Se is a rare element, with an average concentration in igneous bedrock of only 0.05 mg·kg-1, less than any other nutrient element [45]. The largest reservoirs of Se are sulfide ores, pyrite, and high-sulfur coals [44]. Se occurs naturally in soils, at highly variable concentrations dependent on soil type [46]. Most soils have Se concentrations between 0.01 and 2.0 mg·kg−1 [47] with a worldwide mean of 0.4 mg·kg−1 [48]. However, in some soils associated with particular geological formations or climatic conditions, concentrations of Se up to 1200 mg·kg−1 have been reported [47]. Water-soluble selenate has been reported in seleniferous areas of the world at concentrations of around 38 mg Se·kg−1 [49] In such soils, Se is often lacking (for example, in China and USA, mainly derived from sedimentary rocks originating from the Cretaceous Period), while other soils are thought to derive much of their Se from atmospheric depositions [47]. The soils originating from sedimentary rocks, with high organic matter content, can contain high and potentially toxic concentrations of Se. In contrast, soils formed from magmatic rocks typically have low Se concentrations [48]. Generally, Se levels in European soils are low, particularly in eastern Europe [6]. Mountainous countries such as Finland, Sweden, and Scotland are generally deficient in soil Se content, while countries such as UK, France, India, Belgium, Brazil, Serbia, Slovenia, Spain, Portugal, Turkey, Poland, Germany, Denmark, Slovakia, Austria, Ireland, Greece, Netherlands, Italy, China, Nepal, Saudi Arabia, Czech Republic, Croatia, Egypt, Burundi, and New Guinea are reported to have Se-deficient areas. Some known Se-rich regions are the northeast region of Punjab in India, the Enshi district in Hubei province in China, the state of Para in the Brazilian Amazon, Japan, Greenland, USA, Venezuela, and Canada [13]. Se exists in soil in different forms including selenate (Se6+), selenite (Se4+), elemental selenium (Se0), selenide (H2Se), and organic selenide, among which selenate, partially selenite, and organic selenide are per se available for plant uptake [50]. Inorganic Se occurs in three phases: soil-phase-fixed, adsorbed to soil, and soluble, with only adsorbed/soluble forms of Se being available for plant uptake [51]. Se bioavailability in soil varies greatly with different soil properties and composition. Soil pH and redox potential are the key factors reflecting Se bioavailability. Principally, Se is more strongly immobilized in acid and reductive soils [52]. In acid soils, Se is immobilized by sesquioxides [53], and, in organic soil fractions, it is more weakly bound to fulvic acids (FA) compared to humic acids (HA) [54]. Under strongly reductive soil conditions (pH < 4 and Eh < 0), selenate and selenite are easily reduced to selenide or even to elemental selenium, which are less available to plants [55] and show lower mobility [48]. Adsorption of selenite on goethite produces two type of complexes: the protonated selenite anion (HSeO3−) with the active site on the goethite surface, and the bivalent selenite anion (SeO32−) reacting with the surface site. The proportion of each complex depends on the pH of the suspension [53]. Se speciation in the soil is basically controlled by three main mechanisms: oxidation vs. reduction, mineralization vs. immobilization, and volatilization. The rate coefficients of these processes vary depending on Se species, microbial activity, pH, and redox conditions, along with other soil properties [52]. Under aerobic conditions, selenium slowly oxidizes to selenite or selenate, depending on the soil pH. Alkaline soils favor the formation of selenate; however, in moist soils, most of the selenate is readily leached from the surface layers. In acidic soils, iron in colloidal material and sesquioxides render the selenite relatively unavailable as ferric selenite [56,57,58]. There is a general consensus that organic matter interacts with Se via a variety of mechanisms that can immobilize or release Se in soil [59] depending on the type of organic compounds present in soil, with some organic acids having the opposite effect on plant Se availability [60]. In addition to leaching, Se can be lost by volatilization in the form of dimethylselenide (DMSe), dimethyldiselenide (DMDSe), and dimethylselenone or methylmmethylselenite [61]. Strategies for crop biofortification with essential elements for humans aim to increase their accessibility in the soil. The soils lacking in essential elements are usually supplemented by the use of fertilizers. The amount of Se in the rhizosphere, its availability for plant uptake, and the physicochemical soil properties are usually considered for efficient agronomic biofortification.
2.2. Selenium in Plants
Plants mainly take up nutrients via their roots, and the local conditions in the rhizosphere can influence the bioavailability of Se to the plants [62]. The uptake of Se by plants is governed by many factors in the soil and plants. The most important factors determining uptake are the form and concentration of Se in the soil. Other important factors in determining the accumulation of Se by plants include soil properties such as pH, clay content, soil mineralogy, and the concentration of competitive anions [57]. Moreover, the uptake of Se by the plant can be greatly inhibited by the simultaneous occurrence of a high soil content of organic matter, Fe hydroxides, and clay minerals, all of which can adsorb or bind Se [63]. Se can adsorb on positively charged sites of Al-octahedral sheets in clay minerals, such as kaolinite, and it varies greatly with soil pH. Interactions of Se with soil components are either via electrostatic attraction or via complex formation on soil mineral surface [48]. Therefore, the plant-available Se in the soil, such as water-soluble and exchangeable (adsorbed) Se, consists of mobile fractions that are readily taken up by the plants. Plants are the main driving force for nutrient movement from the non-rhizosphere to rhizosphere soil layer [64]. There are no studies describing Se essentiality in plants, although numerous researches have reported the beneficial effects of Se on plant growth especially under biotic and abiotic stress conditions [65,66]. Se and sulfur (S) compete for the same transporters, and Se uptake is generally limited by high S levels [65]. Plants can be classified as hyperaccumulators, secondary-accumulators, and non-accumulators depending on Se accumulation inside their cells [13]. Se non-accumulators act against Se uptake relative to sulfate, whereas Se hyperaccumulator species preferentially absorb Se over S [67], such as Stanleya pinnata [68], Xylorhiza spp., and Symphyotrichum spp. [69]. Hyperaccumulators accumulate higher amounts of Se in their cells (i.e., >1000 mg Se·kg−1 dry weight (DW)) and thrive well in Se-rich regions of the world. Secondary accumulators accumulate Se in the range of 100–1000 mg Se·kg−1 DW, such as Brassica juncea [69], and non-accumulators, such as tobacco (Nicotiana tabacum) and tomato (Solanum lycopersicum) [68], are those plants which accumulate less than 100 mg Se·kg−1 DW [13]. Most crop plants are low rather than high Se accumulators [70]. Se typically stimulates growth and stress resistance at 1–10 mg Se·kg−1 DW, while the tissue concentration at which toxicity occurs is over 100 mg Se·kg−1 DW [9]. Of the two inorganic forms of Se, selenate and selenite, selenate is much more mobile and, thus, more plant-available in soils compared to selenite, which is tightly bound to positively charged binding sites in the soil [60]. Inorganic forms of Se absorbed by plants are transported from the root to the shoot through the xylem with the transport process dependent of the form of externally supplied Se. Se(IV) can be easily absorbed and transported by the xylem, before being distributed further to the reproductive organs by the phloem [62]. There are considerable differences present in the mechanisms of uptake and transport of selenate and selenite in plants [71]. Selenate, which is more soluble than selenite, can pass directly into plant roots [72], whereas selenite is probably transported by phosphate transporters [73]. Selenate is accumulated in plant cells against the gradient of electrochemical potential through a process of active transport [2]. The uptake of selenate across the plasma membrane of root cells is catalyzed by the high-affinity H+/sulfate symporters, homologous to AtSULTR1;1 and AtSULTR1;2 of the model brassicaceous plant Arabidopsis thaliana L. [47]. On the contrary, selenite uptake is carried out through passive diffusion. Moreover, it was reported that it is mediated by active transport, as the uptake of selenite was significantly inhibited by a metabolic inhibitor [13]. Plants cannot directly take up metallic selenide and elemental Se because these forms of Se are water-insoluble, while organo-Se compounds such as seleno-amino acids have relatively higher phytoavailability [48]. The two Se-amino acids produced in the S assimilation pathway are selenocysteine (SeCys) and selenomethionine (SeMet), which are analogues of the S-amino acids cysteine (Cys) and methionine (Met) [74]. Selenocysteine (SeCys) and Selenomethionine (SeMet) are both taken up at rates that were up to 20-fold higher than those observed for selenate or selenite [62]. SeMet is one of the most effectively accumulated Se species in different organs [75]. The distribution of Se in various parts of the plant differs according to species, phase of plant development, and plant physiological condition [2]. Se concentrations tend to be the greatest in the younger leaves of plants and generally increase to a maximum during seedling growth, prior to declining before or upon flowering, when Se is translocated from leaves to reproductive organs [76]. In Se accumulators, Se is accumulated in young leaves during the early vegetative stage of growth. During the reproductive stage, high levels of Se are found in seeds, while the Se content in leaves is drastically reduced. Non-accumulator cereal crop plants, when mature, often show about the same Se content in grain and roots, with smaller amounts in the stem and leaves [2]. Following uptake by root cells, selenate moves rapidly through the root symplast to the stele and is translocated to the shoot, whereas selenite is converted to organoselenium compounds, which often remain within the root [77]. The conversion of selenate to selenite involves the consecutive action of two enzymes. ATP sulfurylase (APS) couples selenite to ATP, forming adenosine phosphoselenate (APSe). APSe is subsequently reduced to selenite by APS reductase (APR). There are isozymes for APS and APR in both chloroplast and cytosol, but most of the selenite reduction likely takes place in the chloroplasts. The further reduction of selenite to selenide may happen exclusively in the chloroplast if it is mediated by sulfite reductase, in analogy with sulfite reduction [78]. Se has dual effects on plant physiology depending on its concentration in plant tissues. At low doses, it can stimulate the growth of plants and counteract many types of environmental stresses [10]. At low concentration, Se enhances plant growth and can act as an antisenescent agent, assisting the upholding of cellular constituents and activities, thus helping to improve plant performance [5]. Se from plants has a negative effect on Se-sensitive ecological partners, which may protect plants from pathogens and herbivores, and which have allelopathic effects on neighboring plants [79]. Se was found to help to increase K+ accumulation in plants and also mediate the increase in chlorophyll a, chlorophyll b, and total chlorophyll content by 65%, 39%, and 56%, respectively, as Se uptake is linked to enhancing the uptake of Mg and Fe [5]. Many studies suggested the role of Se in the following plant physiological mechanisms: the regulation of reactive oxygen species (ROS) and antioxidants, the inhibition of uptake and translocation of heavy metals (HMs), changes in the speciation of HMs, rebuilding of the cell membrane and chloroplast structure, and recovery of the photosynthetic system [10]. Selenium addition to growing media or nutrient solution also increased the net photosynthesis rate, stomatal conductance, and transpiration rate of different plants [5]. Plants vary considerably in their physiological and biochemical response to Se, and a revision of the physiological responses of plants to Se was presented, especially in growth, uptake, transport, and interaction of Se with other minerals [80]. Beneficial soil microorganisms associated with plant roots via symbiotic association are rhizobia, mycorrhizal fungi, actinomycetes, and diazotrophic bacteria that protect plants by various means such as the promotion of nutrient mineralization and production of plant growth hormones [81]. The role of mycorrhizal fungi in enhancing Se uptake in plants [82], as well as the use of Se-tolerant bacteria (Pseudomonas aeruginosa, Bacillus spp., Enterobacter spp., Stenotrophomonas spp., Acinetobacter spp., and Klebsiella spp.) appears to be a possible alternative for Se enhancement of cereals grown on soils with low Se concentration [83]. The uptake of Se by agricultural crops is also dependent on the plant species [57]. The biogeochemical behavior of Se in soil–plant systems is considered to be the basis of Se cycling in living organisms. In fact, the Se content of edible plant parts is often closely related to soil Se content where the crops are being cultivated [48].
2.3. The Impact of Selenium Bioavailability on Human Health
Food safety and nutritional quality represent a priority to improve the health status of the global population [42]. Malnutrition is the main cause of global human mortality, with over 50% of deaths attributed to diet-related diseases [41]. Micronutrient deficiencies in human body mainly result from low concentrations and low availability of micronutrients in daily diet [84]. Although, Se is one of the most significant micronutrients for all forms of life, high levels of Se can be toxic, and the redox chemistry of Se can significantly influence its toxicity, mobility, and bioavailability [1]. Many agricultural areas in the world contain low Se levels, and it is estimated that more than one billion people are suffering from Se deficiency [46]. Se intake from drinking water and other nonfood sources is also minimal in most areas [14]. Individual dietary Se intakes across the world are estimated to range from 3 to 7000 µg per day, whereas, for European countries where estimates are available, mean intake is typically <50 µg·day−1 per person [51]. Daily intakes are high in Venezuela, Canada, USA, and Japan, but much lower in Europe, particularly in its eastern parts [85,86]. The minimal Se requirements depend on the form of Se ingested and the properties of the rest of the diet, as well as on the content of α-tocopherol, which seems to reduce the amount of required Se [87]. Recommended daily intake (following international guidelines) is 40 µg for adult women and 50 µg for men [45]. It has been suggested that approximately 100 selenoproteins may exist in mammalian systems [88]. Selenoproteins include glutathione peroxidases and thioredoxin reductases, which have a variety of functions including protection from oxidative damage, regulation of intracellular redox state, and thyroid hormone metabolism [39]. There is evidence that Se deficiency can adversely affect human health in a number of ways, such as suppressing immune functions, making the organism prone to viral infections, lowering reproduction success (especially male fertility), negatively regulating thyroid function, and causing asthma and various inflammatory conditions [65,89]. Additionally, selenium deficiency causes dilated cardiomyopathy (Keshan disease) and endemic osteoarthropathy (Kashin–Beck disease) [90]. A US study found Se-deficient HIV patients to be 20 times more likely to die from HIV-related causes than those with adequate levels of Se [91]. Se also acts as a cofactor of the GPx family of isoperoxidases which protect human cells against oxidative stress [92]. Recently, an epidemic caused by a novel coronavirus (COVID-19 or 2019-CoV) has spread worldwide while threating human health and threatening the world economy. Supplementary natural treatments should be considered to reduce the viral load in hosts and enhance their immune system, such as Se supplementation. Similar to Zn, Se supplementation to COVID-19 infected patients with low Se blood levels could be an option as a natural treatment against the virus [93]. Previous studies reported that Se concentrations in plants and animals are closely correlated to Se contents in soil. It is also true that, in natural environments, some crop wild relatives have a high percentage of microelements useful to humans, such as Zn and Fe, which are contained in high quantities, e.g., in Aegilops ventricose Tausch [94] which is a progenitor of cultivated wheat (Triticum sp.). Some authors [95] have proposed to start its cultivation in cooperation with local farmers, because, unlike cultivated wheat varieties, Ae. ventricose has a higher quantity of microelements such as Fe and Zn, and it can be use as a natural alternative to conventional medicine to help people with deficiencies in these microelements. Therefore, it should also be verified if some wild species are rich in Se. Humans and animals ingest Se by consuming agricultural products derived from plants that absorb Se from soil [59,63,96]. Generally, human blood Se levels follow the same geographical pattern around the world as those of livestock in the same regions [96]. Se content of foods varies as follows: organ meats and seafood, 0.4 to 1.5 µg·g−1; muscle meats, 0.1 to 0.4 µg·g−1; most agricultural crops, <1 µg·g−1 dry weight, cereals and grains, <0.1 to greater than 0.8 µg·g−1; dairy products, less than 0.1 to 0.3 µg·g−1; fruits and vegetables, less than 0.1 µg·g−1 [63]. In humans, Se absorption from products of plant origin is much easier than Se absorption from products of animal origin. Therefore, researchers are mostly interested in analyzing Se speciation in plant-derived fortified foods [97]. Se concentration in foods, such as rice and wheat grain, can vary greatly across countries and regions. Thus, to avoid Se deficiency and toxicity, it is important to monitor and optimize Se concentrations in various crops [98]. Improving Se uptake by different crops during growth through biofortification can provide additional supplementation of Se in the human diet [70].
3. Agronomic Biofortification
The efficacy of food fortification has been demonstrated consistently considering different micronutrients and different foods [99]. Biofortification was firstly defined by Bouis (1996) [84] and evolved to be thought of as a process of increasing the bioavailable concentrations of essential elements in edible portions of crop plants through agricultural intervention or genetic selection [12]. Different approaches can be explored to enrich plants with Se [93]. Dietary deficiencies of Se can be improved through dietary diversification, food fortification, supplementation, or crop biofortification, including fertilization (agronomic biofortification) and crop improvement (genetic biofortification) [14]. Agronomic biofortification consists of applying fertilizers of mineral elements lacking in the diet in order to increase their concentrations in crops through soil or foliar application [30]. Se-enriched fertilizers for soil or foliar application for agronomic biofortification of cereals provide the best short-term solution for increasing Se concentrations in crops [55]. Compared to foliar application, soil application of Se introduces confounding factors, such as adsorption to soil colloidal surfaces, resulting in less Se available to the plants [70]. Cereals are grown in the greatest quantities and provide more food energy worldwide than any other type of crops; they are, therefore, staple food crops [15]. The availability of soil Se to crops can be affected by irrigation, aeration, liming, and Se fertilization [91]. Generally, mean concentrations of Se in grains are higher in countries having arid climates than in countries having humid climates [80]. Se uptake also varies with the rate of plant growth, soil type, soil concentration of Se, and its oxidation state, along with the rate and method of Se application, as well as concentrations of other anions in soil, especially sulfate [100]. The theory of mineral nutrition suggests that differences in Se concentration in grains occur as a result of different transport and absorption mechanisms, and that the processes are mainly under genotypic influence [64]. Accordingly, plant breeding for improved Se uptake and/or retention by plants may be an effective and sustainable strategy. Numerous findings suggest that it should be possible to breed cultivars with enhanced Se uptake and/or retention or to use genetic engineering to enhance Se levels (and even specific Se metabolites) in food crops [91]. Plant breeding, as the most powerful agricultural approach, may not work effectively in regions where soils have very low plant-available pools of micronutrients due to very adverse soil chemical and physical conditions [101]. Another shortcoming of the breeding approach is the long breeding cycles combined with expensive phenotyping of segregating individuals. Maintaining the stability of target micronutrient traits across diverse types of environments and finding sufficient and promising genotypic variation may also be difficult [101]. Previous investigations suggested that genetic biofortification may be suitable for increasing the amounts of available Fe, carotenoids, and vitamin A, while agronomic biofortification is good for iodine (I) and Se [66]. Agronomic biofortification refers to the application of fertilizer through soil, as a foliar spray, or as seed treatment to enhance the status of a specific micronutrient in edible plant parts [66]. It is worth mentioning that the materials best suitable for Se fertilization include selenate, selenite, slow-release Se fertilizer, Se-enriched yeast, nano-Se, and amino acid-chelated Se [102]. Nano-Se is a relatively new approach to enrich food with Se. Selenium nanoparticles have been regarded as a promising material for many applications due to their unique properties such as high biological activity, bioavailability, low toxicity, high particle dispersion, and large surface area [103]. They represent a promising alternative to other forms of Se, where a reduction in application complexity may be achieved, and this leads to important results in the potentiation of antioxidant metabolism, the promotion of agronomic sustainability, and a reduction in waste [40]. The success of biofortification to enrich plants with Se depends on several factors, such as Se species, the mode of Se fertilization, and the crop species [93]. Investigations indicate that agronomic biofortification of wheat by Se fertilization may be the best approach to increase the Se intake by humans [23]. The average Se content in crop plants from non-seleniferous soils varies between about 0.01 and 1.0 mg·kg−1 DW [71]. Recommended Se contents in grain, used as fodder and food, are 0.2–0.3 mg Se·kg−1 dry matter (DM) and 01–0.2 mg Se·kg−1 DM, respectively [60]. Studies have shown that, in adequate concentrations, Se can be also beneficial to plants by increasing their productivity [104]. It has been reported that Se improves the yield of food crops such as wheat, barley, rice, and maize [16]. Therefore, type of application, form of Se, and time of application play a critical role in achieving prosperous biofortification in cereal crops.
4. Selenium Content Affected by the Method and Form of Selenium: Meta-Analysis
The type of application is the first important factor that significantly increases Se content in plants, and most studies have demonstrated that foliar application is a more efficient method of fertilization, although soil fertilization is more popular [13,39,41,42,105]. Foliar spraying leads to more efficient uptake of Se compared to soil application corroborated by the lack of soil residuals. Foliar techniques use the minimum amount of Se salts and are the most effective, safe, and economically acceptable way of improving Se contents in crops such as wheat [37]. Foliar application of Se is a more efficient method of biofortification because there is no root-to-shoot translocation and soils can act as considerable sinks for Se [39]. Accordingly, the plant availability of applied Se can decrease rapidly in soils [106]. Se form is the second important parameter for effective biofortification. Most studies have shown selenate (where Se exists in its highest oxidation state, +6) to be the most effective form when applied to the soil and usually more effective than selenite (Se+4) when applied in a foliar manner [41]. The higher efficiency of selenate results from the more rapid uptake of Se(VI) and translocation from root to stem and leaves, as well as quicker transformation into the organic form, while selenite(IV) is more easily adsorbed to the soil, which makes its uptake difficult [42]. Selenate is easily distributed from roots to shoots, whereas selenite or its metabolic products tend to accumulate in roots [58]. It can be expected that most of the applied Se as selenate remains in an inorganic form in shoots, whereas most of the added selenite is incorporated as organic Se, i.e., in Se-amino acids and Se-proteins [43]. One study showed that the efficiency of foliar fertilization in rice plants becomes higher with sodium selenite compared to selenate [107]. The Se concentrations in rice grains tend to be higher compared to maize and wheat grains [108], possibly because rice cultivars can be differentiated into high-Se and low-Se cultivars [64]. As a staple food, rice is an excellent source of energy, with prevailing consumption in over 30 countries, providing about 80% of daily caloric intake to ca. three billion people [107], making it a suitable target for agronomic Se biofortification. Studies have shown that foliar application with selenate and selenite increased the Se content in wheat grains [20,33,34,35]. Foliar application of Se obtained good results, and it is a most commonly used method in wheat biofortification. The use of Se fertilizers in soil leads to low rates of Se enrichment in edible plant parts. Moreover, long-term use can be toxic to the nearby ecosystem; hence, the use of Se fertilizers should be carried out carefully to avoid toxicity [13]. Despite the cons of fertilizer application, agronomic biofortification is a justified measure to achieve increased Se content in maize grain [16,109,110]. The absorption, accumulation, distribution, and metabolization of Se in mature maize plants depend on the form of Se supplied [27]. Barley is the major small grain cereal after rice, wheat, and maize [18]. Two-rowed barley might be a suitable candidate to be included in Se biofortification programs [32], with foliar application of sodium selenate immediately after anthesis or during the germination stage of the malting process, resulting in the accumulation of Se in the foods [111]. The analysis of results of studies considering Se biofortification showed considerable variation in methods, times, and forms of application in the grains of four major cereal crops (Table 1). In rice, the distribution of values was skewed toward the right due to the limited number of studies after Premarantha et al. (2012). However, the increase varied from 0.025 µg Se per 1 g of applied Se [19] to 0.42 µg Se per 1 g of applied Se [28] in field experiments and up to 6.34 µg Se per 1 g of applied Se in a pot experiment in controlled conditions [29]. In maize, the reported values in µg Se per 1 g of applied Se varied from 0.091 [112] to 0.92 [30]. The results of barley showed considerably lower efficiencies when selenite was used [31,32] with values varying from 0.0422 to 0.78 µg Se per 1 g of applied Se. In wheat, the lowest value of 0.1125 µg Se per 1 g of applied Se was reported for selenite in a study by Ducsay et al. (2016), while the highest value of 0.853 was reported by De Vita et al. (2017).
To further analyze the data patterns from other studies, a meta-analysis was conducted by setting a liner mixed model [113]. Mixed linear models were set with normalized data of Se increase per 1 g of applied Se as response variable. Input data were values from Table 1. Eight models in total were set in the R lme4 library [113] with all random effects and intercepts for each term, assuming unstructured error variance. Models included eight combinations of the following factors: application type, species, form of Se, and their respective interactions. The final model was chosen on the basis of the lowest scores of Akaike and Bayesian information criteria (AIC and BIC), and it included three random terms. The first term was with the intercept varying as a function of application type and type of Se, the second was with the intercept varying as a function of species and type of Se, and the third was the random intercept term type of experiment with a fixed mean. Model coefficients were extracted using the lme4 coef() command.
The variance component analysis (Table 2) showed the lowest amount of variance explained by the application type, followed by the species × Se form interaction, while the type of experiment explained the most considerable portion of variance. Expectedly, a low value of residual variance was determined. The analysis of coefficients of random effects showed lower efficiency of Se application in soil compared to foliar application. In the analysis of species × form of application interaction, lower efficiencies of selenite were determined in all crop species, while the sizes of calculated coefficients were comparable only in rice. A plastic pot experiment in a growth chamber showed a multifold increase in Se accumulation, probably due to the lack of atmospheric factors affecting the leaching and volatilization of Se [39]. Field growing represents a more feasible biofortification method compared to plastic pots for growing cereals due to the low amount of food that can be produced. Plants, when supplied with selenite, had selenium concentrations in the xylem exudate lower than selenate [114]. The same pattern was confirmed by the mixed model analysis (Table 2) for grains. The second largest proportion of variance was explained by the interaction of application type × Se form with the foliar application of selenate being the best combination to achieve successful agronomic biofortification. Application of selenate to soil represents the second-best option to increase Se level in cereals. Application of selenite to the soil or in a foliar manner represents the least effective options, and foliar application of selenite is a better choice for increasing Se in plants. Wheat is the most important cereal for agronomic biofortification because of its importance for human diet [115], and it had a higher uptake of Se in the grain compared to all other studied cereals (Table 2). Generally, selenite is more strongly adsorbed by the soil solid phase and, thus, less soluble than selenate in soil solutions [58], while genotype also influences the increase in Se in the grain [116]. The best results observed in the species × Se form analysis were wheat/selenate > barley/selenate > maize/selenate > maize/selenite > wheat/selenite > barley/selenite > rice/selenite > rice/selenate, in the order from more efficient to less efficient. The higher translocation of selenate into grains (Table 2) might, therefore, be due to its higher bioavailability to plants than selenite, which is more strongly adsorbed to the soil surfaces [43]. Furthermore, in plants, selenite and phosphate compete for uptake because they share common transporters [69], while the translocation of selenate from roots to shoots occurs more readily than selenite [67]. Barley was also shown to be efficient in Se uptake [111], and selenate application resulted in enhanced accumulation of Se in barley grains [18] compared to selenite. Chilimba et al. [109] reported that agronomic biofortification of maize with Se appears to be a feasible option for increasing dietary Se [109]. Foliar application of selenite in rice plants resulted in higher total Se content compared to soil application [117], and selenite is a more effective Se form for rice biofortification with differing accumulation across genotypes [118], as discussed in this paper. Considering the analyzed data and reviewed literature, we can see that wheat is the most cultivated cereal and has the highest efficiency of Se uptake and translocation toward its edible parts. Barley showed higher efficiency regarding Se biofortification compared to rice and maize, although it is not cultivated as much. From the studied cereals, selenite was shown only in rice to be a more efficient Se form than selenate. The largest proportion of variance in the mixed model analysis was attributed to application type × Se form (Table 2), representing the most important factor in choosing the right strategy of Se agronomic biofortification, along with the choice of right plant species.
5. Conclusions and Future Perspectives
Cereals are the most common food in the human diet worldwide. Over one billion people have an Se deficiency, which leads to various disorders in human health. Recently, the necessity of a functioning immune system has been emphasized. Connecting these facts, we conclude that selenium agronomic biofortification should be done on the most widely used foodstuff to reduce Se malnutrition. According to a meta-analysis, foliar application has been shown to be a more efficient method than soil application. Foliar application is also a more cost-effective method with numerous advantages. The form of applied Se also plays an important role in increasing Se content in the cereal grain, with the selenate form being more effective for agronomic biofortification. For further research, it is necessary to determine the exact doses of selenium for individual crops that will ultimately have the best outcome for plants, humans, and soil. According to the available data and results of the meta-analysis, there is a possibility to design a computational model for predicting the efficiency of applied selenium with respect to crop and soil type, selenium form, and method of application. There are several papers that do not have yield data, and this would be crucial in the development of a model; thus, there is a niche for the improvement of scientific papers on agronomic Se biofortification. An interesting challenge would be to determine if some wild plant species have high concentrations of Se, starting from segetal weed species, which are often exclusive to these environments, sometimes have medicinal properties, and may be at risk of extinction due to intensive agricultural practices [119,120].
Author Contributions
Conceptualization, Z.L. and L.G.; methodology, L.G. and Z.L.; formal analysis, L.G.; investigation, L.G., Z.L., T.V. and B.R.; data curation, L.G.; writing—original draft preparation, L.G.; writing—review and editing, Z.L. and T.V.; supervision, Z.L. and T.V.; project administration, T.V.; funding acquisition, T.V. and Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the project “Application of Nanobiotechnology for Nutritional Supplementation with Selenium—(grant HRZZ-IP-2018-01-8119)”, and work of doctoral student Lucija Galić has been fully supported by the “Young researches’ career development project–training of doctoral students” through grant HRZZ-DOK-2020-01-1288 both financed by the Croatian Science Foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ullah, H.; Liu, G.; Yousaf, B.; Ali, M.U.; Irshad, S.; Abbas, Q.; Ahmad, R. A Comprehensive Review on Environmental Transformation of Selenium: Recent Advances and Research Perspectives; Springer: Dordrecht, The Netherlands, 2019; Volume 41, ISBN 0123456789. [Google Scholar]
- Terry, N.; Zayed, A.M.; De Souza, M.P.; Tarun, A.S. Selenium in Higher Plants. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehdi, Y.; Hornick, J.L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef] [Green Version]
- El-Ramady, H.; Abdalla, N.; Alshaal, T.; Domokos-Szabolcsy, É.; Elhawat, N.; Prokisch, J.; Sztrik, A.; Fári, M.; El-Marsafawy, S.; Shams, M.S. Selenium in soils under climate change, implication for human health. Environ. Chem. Lett. 2014, 13, 1–19. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Al Mahmud, J.; Nahar, K.; Fujita, M. Selenium in plants: Boon or bane? Environ. Exp. Bot. 2020, 178, 104170. [Google Scholar] [CrossRef]
- dos Reis, A.R.; El-Ramady, H.; Santos, E.F.; Gratão, P.L.; Schomburg, L. Overview of Selenium Deficiency and Toxicity Worldwide: Affected Areas, Selenium-Related Health Issues, and Case Studies. In Selenium in Plants; Springer: Cham, Switzerland, 2017; pp. 209–230. [Google Scholar] [CrossRef]
- Finley, J.W. Selenium accumulation in plant foods. Nutr. Rev. 2005, 63, 196–202. [Google Scholar] [CrossRef] [PubMed]
- El-Ramady, H.; Faizy, S.E.D.; Abdalla, N.; Taha, H.; Domokos-Szabolcsy, É.; Fari, M.; Elsakhawy, T.; Omara, A.E.D.; Shalaby, T.; Bayoumi, Y.; et al. Selenium and nano-selenium biofortification for human health: Opportunities and challenges. Soil Syst. 2020, 4, 57. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.H. On the ecology of selenium accumulation in plants. Plants 2019, 8, 197. [Google Scholar] [CrossRef] [Green Version]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Schiavon, M.; Berto, C.; Malagoli, M.; Trentin, A.; Sambo, P.; Dall’Acqua, S.; Pilon-Smits, E.A.H. Selenium biofortification in radish enhances nutritional quality via accumulation of methyl-selenocysteine and promotion of transcripts and metabolites related to glucosinolates, phenolics amino acids. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Ríos, J.J.; Rosales, M.A.; Blasco, B.; Cervilla, L.M.; Romero, L.; Ruiz, J.M. Biofortification of Se and induction of the antioxidant capacity in lettuce plants. Sci. Hortic. 2008, 116, 248–255. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An overview of selenium uptake, metabolism, and toxicity in plants. Front. Plant Sci. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broadley, M.R.; Alcock, J.; Alford, J.; Cartwright, P.; Foot, I.; Fairweather-Tait, S.J.; Hart, D.J.; Hurst, R.; Knott, P.; McGrath, S.P.; et al. Selenium biofortification of high-yielding winter wheat (Triticum aestivum L.) by liquid or granular Se fertilisation. Plant. Soil 2010, 332, 5–18. [Google Scholar] [CrossRef]
- Sarwar, H. The importance of cereals (Poaceae: Gramineae) nutrition in human health: A review. J. Cereal. Oilseeds 2013, 4, 32–35. [Google Scholar] [CrossRef]
- Bocchini, M.; D’Amato, R.; Ciancaleoni, S.; Fontanella, M.C.; Palmerini, C.A.; Beone, G.M.; Onofri, A.; Negri, V.; Marconi, G.; Albertini, E.; et al. Soil selenium (Se) biofortification changes the physiological, biochemical and epigenetic responses to water stress in Zea mays L. by inducing a higher drought tolerance. Front. Plant Sci. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Abenavoli, L.; Milanovic, M.; Procopio, A.C.; Spampinato, G.; Maruca, G.; Perrino, E.V.; Mannino, G.C.; Fagoonee, S.; Luzza, F.; Musarella, C.M. Ancient wheats: Beneficial effects on insulin resistance. Minerva Med. 2021. [Google Scholar] [CrossRef]
- Narwal, S.; Kumar, D.; Kharub, A.S.; Verma, R.P.S. Barley Biofortification: Present Status and Future Prospects; Elsevier Inc.: Amsterdam, The Netherlands, 2020; ISBN 9780128184448. [Google Scholar]
- Premarathna, L.; McLaughlin, M.J.; Kirby, J.K.; Hettiarachchi, G.M.; Stacey, S.; Chittleborough, D.J. Selenate-enriched urea granules are a highly effective fertilizer for selenium biofortification of paddy rice grain. J. Agric. Food Chem. 2012, 60, 6037–6044. [Google Scholar] [CrossRef] [Green Version]
- De Vita, P.; Platani, C.; Fragasso, M.; Ficco, D.B.M.; Colecchia, S.A.; Del Nobile, M.A.; Padalino, L.; Di Gennaro, S.; Petrozza, A. Selenium-enriched durum wheat improves the nutritional profile of pasta without altering its organoleptic properties. Food Chem. 2017, 214, 374–382. [Google Scholar] [CrossRef]
- Xia, Q.; Yang, Z.; Shui, Y.; Liu, X.; Chen, J.; Khan, S.; Wang, J.; Gao, Z. Methods of Selenium Application Differentially Modulate Plant Growth, Selenium Accumulation and Speciation, Protein, Anthocyanins and Concentrations of Mineral Elements in Purple-Grained Wheat. Front. Plant Sci. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Zou, C.; Du, Y.; Rashid, A.; Ram, H.; Savasli, E.; Pieterse, P.J.; Ortiz-Monasterio, I.; Yazici, A.; Kaur, C.; Mahmood, K.; et al. Simultaneous Biofortification of Wheat with Zinc, Iodine, Selenium, and Iron through Foliar Treatment of a Micronutrient Cocktail in Six Countries. J. Agric. Food Chem. 2019, 67, 8096–8106. [Google Scholar] [CrossRef] [Green Version]
- Manojlović, M.S.; Lončarić, Z.; Cabilovski, R.R.; Popović, B.; Karalić, K.; Ivezić, V.; Ademi, A.; Singh, B.R. Biofortification of wheat cultivars with selenium. Acta Agric. Scand. Sect. B Soil Plant. Sci. 2019, 69, 715–724. [Google Scholar] [CrossRef]
- Ramkissoon, C.; Degryse, F.; da Silva, R.C.; Baird, R.; Young, S.D.; Bailey, E.H.; McLaughlin, M.J. Improving the efficacy of selenium fertilizers for wheat biofortification. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- de Lima Lessa, J.H.; Araujo, A.M.; Ferreira, L.A.; da Silva Júnior, E.C.; de Oliveira, C.; Corguinha, A.P.B.; Martins, F.A.D.; de Carvalho, H.W.P.; Guilherme, L.R.G.; Lopes, G. Agronomic biofortification of rice (Oryza sativa L.) with selenium and its effect on element distributions in biofortified grains. Plant. Soil 2019, 444, 331–342. [Google Scholar] [CrossRef]
- Huang, G.; Ding, C.; Yu, X.; Yang, Z.; Zhang, T.; Wang, X. Characteristics of Time-Dependent Selenium Biofortification of Rice (Oryza sativa L.). J. Agric. Food Chem. 2018, 66, 12490–12497. [Google Scholar] [CrossRef]
- Longchamp, M.; Castrec-Rouelle, M.; Biron, P.; Bariac, T. Variations in the accumulation, localization and rate of metabolization of selenium in mature Zea mays plants supplied with selenite or selenate. Food Chem. 2015, 182, 128–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, H.P.G.; de Queiroz Barcelos, J.P.; Junior, E.F.; Santos, E.F.; Silva, V.M.; Moraes, M.F.; Putti, F.F.; dos Reis, A.R. Agronomic biofortification of upland rice with selenium and nitrogen and its relation to grain quality. J. Cereal Sci. 2018, 79, 508–515. [Google Scholar] [CrossRef]
- Wang, Y.D.; Wang, X.; Wong, Y.S. Generation of selenium-enriched rice with enhanced grain yield, selenium content and bioavailability through fertilisation with selenite. Food Chem. 2013, 141, 2385–2393. [Google Scholar] [CrossRef]
- Ngigi, P.B.; Lachat, C.; Masinde, P.W.; Du Laing, G. Agronomic biofortification of maize and beans in Kenya through selenium fertilization. Environ. Geochem. Health 2019, 41, 2577–2591. [Google Scholar] [CrossRef]
- Winter, K.A.; Sanderson, J.B.; Gupta, U.C. Selenium content of barley as influenced by selenite- and selenate-enriched fertilizers. Commun. Soil Sci. Plant. Anal. 1993, 24, 1165–1170. [Google Scholar] [CrossRef]
- Rodrigo, S.; Santamaria, O.; Chen, Y.; McGrath, S.P.; Poblaciones, M.J. Selenium speciation in malt, wort, and beer made from selenium-biofortified two-rowed barley grain. J. Agric. Food Chem. 2014, 62, 5948–5953. [Google Scholar] [CrossRef] [PubMed]
- Poblaciones, M.J.; Rodrigo, S.; Santamaría, O.; Chen, Y.; McGrath, S.P. Agronomic selenium biofortification in Triticum durum under Mediterranean conditions: From grain to cooked pasta. Food Chem. 2014, 146, 378–384. [Google Scholar] [CrossRef]
- Ducsay, L.; Ložek, O.; Marček, M.; Varényiová, M.; Hozlár, P.; Lošák, T. Possibility of selenium biofortification of winter wheat grain. Plant. Soil Environ. 2016, 62, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Ali, F.; Wang, M.; Dinh, Q.T.; Zhou, F.; Bañuelos, G.S.; Liang, D. Understanding boosting selenium accumulation in Wheat (Triticum aestivum L.) following foliar selenium application at different stages, forms, and doses. Environ. Sci. Pollut. Res. 2020, 27, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Liu, K.; Li, M.; Zhang, W.; Zhao, X.; Zhao, Z. Field Crops Research Di ff erence of selenium uptake and distribution in the plant and selenium form in the grains of rice with foliar spray of selenite or selenate at di ff erent stages. Field Crop. Res. 2017, 211, 165–171. [Google Scholar] [CrossRef]
- Pezzarossa, B.; Remorini, D.; Gentile, M.L.; Massai, R. Effects of foliar and fruit addition of sodium selenate on selenium accumulation and fruit quality. J. Sci. Food Agric. 2012, 92, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Kápolna, E.; Hillestrøm, P.R.; Laursen, K.H.; Husted, S.; Larsen, E.H. Effect of foliar application of selenium on its uptake and speciation in carrot. Food Chem. 2009, 115, 1357–1363. [Google Scholar] [CrossRef]
- Winkel, L.H.; Vriens, B.; Jones, G.D.; Schneider, L.S.; Pilon-Smits, E.; Bañuelos, G.S. Selenium cycling across soil-plant-atmosphere interfaces: A critical review. Nutrients 2015, 7, 4199–4239. [Google Scholar] [CrossRef] [Green Version]
- Márquez, V.G.; Moreno, Á.M.; Mendoza, A.B.; Macías, J.M. Ionic selenium and nanoselenium as biofortifiers and stimulators of plant metabolism. Agronomy 2020, 10, 1399. [Google Scholar] [CrossRef]
- Lyons, G. Biofortification of cereals with foliar selenium and iodine could reduce hypothyroidism. Front. Plant Sci. 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Izydorczyk, G.; Ligas, B.; Mikula, K.; Witek-Krowiak, A.; Moustakas, K.; Chojnacka, K. Biofortification of edible plants with selenium and iodine—A systematic literature review. Sci. Total Environ. 2020, 754, 141983. [Google Scholar] [CrossRef]
- Cartes, P.; Gianfreda, L.; Mora, M.L. Uptake of selenium and its antioxidant activity in ryegrass when applied as selenate and selenite forms. Plant. Soil 2005, 276, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Nancharaiah, Y.V.; Lens, P.N.L. Ecology and Biotechnology of Selenium-Respiring Bacteria. Microbiol. Mol. Biol. Rev. 2015, 79, 61–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haug, A.; Graham, R.D.; Christophersen, O.A.; Lyons, G.H. How to use the world’s scarce selenium resources efficiently to increase the selenium concentration in food. Microb. Ecol. Health Dis. 2007, 19, 209–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasin, M.; El Mehdawi, A.F.; Jahn, C.E.; Anwar, A.; Turner, M.F.S.; Faisal, M.; Pilon-Smits, E.A.H. Seleniferous soils as a source for production of selenium-enriched foods and potential of bacteria to enhance plant selenium uptake. Plant. Soil 2014, 386, 385–394. [Google Scholar] [CrossRef]
- White, P.J. Selenium in soils and crops. Mol. Integr. Toxicol. 2018, 29–50. [Google Scholar] [CrossRef]
- Natasha; Shahid, M.; Niazi, N.K.; Khalid, S.; Murtaza, B.; Bibi, I.; Rashid, M.I. A critical review of selenium biogeochemical behavior in soil-plant system with an inference to human health. Environ. Pollut. 2018, 234, 915–934. [Google Scholar] [CrossRef]
- Mayland, H.F.; James, L.F.; Panter, K.E.; Sonderegger, J.L. Selenium in Seleniferous Environments. Selenium Agric. Environ. 2015, 15–50. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Ren, J.; Xue, C.; Lin, E. Study on the relationship between soil selenium and plant selenium uptake. Plant. Soil 2005, 277, 197–206. [Google Scholar] [CrossRef]
- Hart, D.J.; Fairweather-Tait, S.J.; Broadley, M.R.; Dickinson, S.J.; Foot, I.; Knott, P.; McGrath, S.P.; Mowat, H.; Norman, K.; Scott, P.R.; et al. Selenium concentration and speciation in biofortified flour and bread: Retention of selenium during grain biofortification, processing and production of Se-enriched food. Food Chem. 2011, 126, 1771–1778. [Google Scholar] [CrossRef]
- Li, Z.; Liang, D.; Peng, Q.; Cui, Z.; Huang, J.; Lin, Z. Interaction between selenium and soil organic matter and its impact on soil selenium bioavailability: A review. Geoderma 2017, 295, 69–79. [Google Scholar] [CrossRef]
- Zhang, P.; Sparks, D.L. Kinetics of Selenate and Selenite Adsorption/Desorption at the Goethite/Water Interface. Environ. Sci. Technol. 1990, 24, 1848–1856. [Google Scholar] [CrossRef]
- Ramkissoon, C.; Degryse, F.; Young, S.; Bailey, E.H.; McLaughlin, M.J. Effect of soil properties on time-dependent fixation (ageing) of selenate. Geoderma 2021, 383, 114741. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, Y.; Li, J.; Wan, Y.; Huang, Q.; Guo, Y.; Li, H. Effects of Different Forms of Selenium Fertilizers on Se Accumulation, Distribution, and Residual Effect in Winter Wheat-Summer Maize Rotation System. J. Agric. Food Chem. 2017, 65, 1116–1123. [Google Scholar] [CrossRef] [PubMed]
- Girling, C.A. Selenium in agriculture and the environment. Agric. Ecosyst. Environ. 1984, 11, 37–65. [Google Scholar] [CrossRef]
- Mikkelsen, R.L.; Page, A.L.; Bingham, F.T. Factors Affecting Selenium Accumulation by Agricultural Crops. In Selenium in Agriculture and the Environment; Jacobs, L.W., Ed.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1989; pp. 65–94. [Google Scholar]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Dinh, Q.T.; Li, Z.; Tran, T.A.T.; Wang, D.; Liang, D. Role of organic acids on the bioavailability of selenium in soil: A review. Chemosphere 2017, 184, 618–635. [Google Scholar] [CrossRef] [PubMed]
- Eich-Greatorex, S.; Sogn, T.A.; Øgaard, A.F.; Aasen, I. Plant availability of inorganic and organic selenium fertiliser as influenced by soil organic matter content and pH. Nutr. Cycl. Agroecosyst. 2007, 79, 221–231. [Google Scholar] [CrossRef]
- Zieve, R.; Peterson, P.J. Factors influencing the volatilization of selenium from soil. Sci. Total Environ. 1981, 19, 277–284. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, J.; Kronzucker, H.J.; Shi, W. Selenium Biofortification and Interaction With Other Elements in Plants: A Review. Front. Plant Sci. 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Rayman, M.P. Food-chain selenium and human health: Emphasis on intake. Br. J. Nutr. 2008, 100, 254–268. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Xing, G.; Tang, S.; Pang, Y.; Yi, Q.; Huang, Q.; Huang, X.; Huang, J.; Li, P.; Fu, H. Improving soil selenium availability as a strategy to promote selenium uptake by high-Se rice cultivar. Environ. Exp. Bot. 2019, 163, 45–54. [Google Scholar] [CrossRef]
- Malagoli, M.; Schiavon, M.; Dall’Acqua, S.; Pilon-Smits, E.A.H. Effects of selenium biofortification on crop nutritional quality. Front. Plant Sci. 2015, 6, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, N.; Akhtar, M.; Kamran, M.A.; Imran, M.; Riaz, M.A.; Kamran, K.; Hussain, S. Selenium biofortification in food crops: Key mechanisms and future perspectives. J. Food Compos. Anal. 2020, 93, 103615. [Google Scholar] [CrossRef]
- Sors, T.G.; Ellis, D.R.; Salt, D.E. Selenium uptake, translocation, assimilation and metabolic fate in plants. Photosynth. Res. 2005, 86, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Wiesner-Reinhold, M.; Schreiner, M.; Baldermann, S.; Schwarz, D.; Hanschen, F.S.; Kipp, A.P.; Rowan, D.D.; Bentley-Hewitt, K.L.; McKenzie, M.J. Mechanisms of selenium enrichment and measurement in brassicaceous vegetables, and their application to human health. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, M.; Pilon-Smits, E.A.H. The fascinating facets of plant selenium accumulation—Biochemistry, physiology, evolution and ecology. New Phytol. 2017, 213, 1582–1596. [Google Scholar] [CrossRef] [Green Version]
- Newman, R.; Waterland, N.; Moon, Y.; Tou, J.C. Selenium Biofortification of Agricultural Crops and Effects on Plant Nutrients and Bioactive Compounds Important for Human Health and Disease Prevention—A Review. Plant. Foods Hum. Nutr. 2019, 74, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Läuchli, A. Selenium in Plants: Uptake, Functions, and Environmental Toxicity. Bot. Acta 1993, 106, 455–468. [Google Scholar] [CrossRef]
- Funes-Collado, V.; Morell-Garcia, A.; Rubio, R.; López-Sánchez, J.F. Selenium uptake by edible plants from enriched peat. Sci. Hortic. 2013, 164, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Hawrylak-Nowak, B.; Matraszek, R.; Pogorzelec, M. The dual effects of two inorganic selenium forms on the growth, selected physiological parameters and macronutrients accumulation in cucumber plants. Acta Physiol. Plant. 2015, 37. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, M.; Lima, L.W.; Jiang, Y.; Hawkesford, M.J. Effects of Selenium on Plant Metabolism and Implications for Crops and Consumers. In Selenium in Plants; Springer: Cham, Switzerland, 2017; pp. 257–275. [Google Scholar] [CrossRef]
- Pedrero, Z.; Madrid, Y. Novel approaches for selenium speciation in foodstuffs and biological specimens: A review. Anal. Chim. Acta 2009, 634, 135–152. [Google Scholar] [CrossRef]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, P.J. Selenium metabolism in plants. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.H.; Quinn, C.F. Selenium metabolism in plants. In Cell Biology of Metals and Nutrients; Springer: Berlin/Heidelberg, Germany, 2010; pp. 225–241. [Google Scholar]
- Pilon-Smits, E.A.H. Selenium in Plants. In Scientific American; Springer International Publishing: Cham, Switzerland, 2015; Volume 8, p. 2957. ISBN 9783319088075. [Google Scholar]
- El-Ramady, H.; Abdalla, N.; Alshaal, T.; El-Henawy, A.; Faizy, S.E.D.A.; Shams, M.S.; Shalaby, T.; Bayoumi, Y.; Elhawat, N.; Shehata, S.; et al. Selenium and its role in higher plants. In Pollutants in Buildings, Water and Living Organisms; Springer International Publishing: Cham, Switzerland, 2015; pp. 235–296. ISBN 9783319192765. [Google Scholar]
- Jha, A.B.; Warkentin, T.D. Biofortification of pulse crops: Status and future perspectives. Plants 2020, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Qu, J.; Pu, Y.; Rao, S.; Xu, F.; Wu, C. Selenium biofortification of crop food by beneficial microorganisms. J. Fungi 2020, 6, 59. [Google Scholar] [CrossRef]
- Patel, P.J.; Trivedi, G.R.; Shah, R.K.; Saraf, M. Selenorhizobacteria: As biofortification tool in sustainable agriculture. Biocatal. Agric. Biotechnol. 2018, 14, 198–203. [Google Scholar] [CrossRef]
- Yang, X.E.; Chen, W.R.; Feng, Y. Improving human micronutrient nutrition through biofortification in the soil-plant system: China as a case study. Environ. Geochem. Health 2007, 29, 413–428. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Stoffaneller, R.; Morse, N.L. A review of dietary selenium intake and selenium status in Europe and the Middle East. Nutrients 2015, 7, 1494–1537. [Google Scholar] [CrossRef]
- Shamberger, R.J. Selenium in the environment. Sci. Total Environ. 1981, 17, 59–74. [Google Scholar] [CrossRef]
- Lyons, G.H.; Stangoulis, J.C.R.; Graham, R.D. Exploiting micronutrient interaction to optimize biofortification programs: The case for inclusion of selenium and iodine in the HarvestPlus program. Nutr. Rev. 2004, 62, 247–252. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; Zhao, F.J. Strategies for increasing the selenium content of wheat. J. Cereal Sci. 2007, 46, 282–292. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błazejak, S. Current knowledge on the importance of selenium in food for living organisms: A review. Molecules 2016, 21, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, G.; Stangoulis, J.; Graham, R. High-selenium wheat: Biofortification for better health. Nutr. Res. Rev. 2003, 16, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Alarcon, M.; Cabrera-Vique, C. Selenium in food and the human body: A review. Sci. Total Environ. 2008, 400, 115–141. [Google Scholar] [CrossRef]
- Schiavon, M.; Nardi, S.; dalla Vecchia, F.; Ertani, A. Selenium biofortification in the 21st century: Status and challenges for healthy human nutrition. Plant. Soil 2020, 453, 245–270. [Google Scholar] [CrossRef]
- Rawat, N.; Tiwari, V.K.; Singh, N.; Randhawa, G.S.; Singh, K.; Chhuneja, P.; Dhaliwal, H.S. Evaluation and utilization of Aegilops and wild Triticum species for enhancing iron and zinc content in wheat. Genet. Resour. Crop. Evol. 2009, 56, 53–64. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P. Crop wild relatives (Cwr) priority in italy: Distribution, ecology, in situ and ex situ conservation and expected actions. Sustainability 2021, 13, 1682. [Google Scholar] [CrossRef]
- Brown, K.; Arthur, J. Selenium, selenoproteins and human health: A review. Public Health Nutr. 2001, 4, 593–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amato, R.; Regni, L.; Falcinelli, B.; Mattioli, S.; Benincasa, P.; Dal Bosco, A.; Pacheco, P.; Proietti, P.; Troni, E.; Santi, C.; et al. Current Knowledge on Selenium Biofortification to Improve the Nutraceutical Profile of Food: A Comprehensive Review. J. Agric. Food Chem. 2020, 68, 4075–4097. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Pilon-Smits, E.A.H.; Zhao, F.J.; Williams, P.N.; Meharg, A.A. Selenium in higher plants: Understanding mechanisms for biofortification and phytoremediation. Trends Plant. Sci. 2009, 14, 436–442. [Google Scholar] [CrossRef]
- Osendarp, S.J.M.; Martinez, H.; Garrett, G.S.; Neufeld, L.M.; De-Regil, L.M.; Vossenaar, M.; Darnton-Hill, I. Large-Scale Food Fortification and Biofortification in Low- and Middle-Income Countries: A Review of Programs, Trends, Challenges, and Evidence Gaps. Food Nutr. Bull. 2018, 39, 315–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephen, R.C.; Saville, D.J.; Watkinson, J.H. The effects of sodium selenate applications on growth and selenium concentration in wheat. N. Z. J. Crop. Hortic. Sci. 1989, 17, 229–237. [Google Scholar] [CrossRef]
- Mao, H.; Wang, J.; Wang, Z.; Zan, Y.; Lyons, G.; Zou, C. Using agronomic biofortification to boost zinc, selenium, and iodine concentrations of food crops grown on the loess plateau in China. J. Soil Sci. Plant. Nutr. 2014, 14, 459–470. [Google Scholar] [CrossRef] [Green Version]
- El-Ramady, H.; Abdalla, N.; Taha, H.S.; Alshaal, T.; El-Henawy, A.; Faizy, S.E.D.A.; Shams, M.S.; Youssef, S.M.; Shalaby, T.; Bayoumi, Y.; et al. Selenium and nano-selenium in plant nutrition. Environ. Chem. Lett. 2016, 14, 123–147. [Google Scholar] [CrossRef]
- Kumar, A.; Prasad, K.S. Role of nano-selenium in health and environment. J. Biotechnol. 2020, 325, 152–163. [Google Scholar] [CrossRef]
- Lara, T.S.; de Lima Lessa, J.H.; de Souza, K.R.D.; Corguinha, A.P.B.; Martins, F.A.D.; Lopes, G.; Guilherme, L.R.G. Selenium biofortification of wheat grain via foliar application and its effect on plant metabolism. J. Food Compos. Anal. 2019, 81, 10–18. [Google Scholar] [CrossRef]
- Petković, K.P.; Anojlović, M.M.; Abilovski, R.Č.; Rstić, Đ.K. Foliar Application Of Selenium, Zinc And Copper In Alfalfa (Medicago sativa L.) Biofortification. Turk. J. Filed Crop. 2019, 24, 81–90. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.; Mao, H.; Zhao, H.; Huang, D. Increasing Se concentration in maize grain with soil- or foliar-applied selenite on the Loess Plateau in China. Field Crop. Res. 2013, 150, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Lidon, F.C.; Oliveira, K.; Ribeiro, M.M.; Pelica, J.; Pataco, I.; Ramalho, J.C.; Leitão, A.E.; Almeida, A.S.; Campos, P.S.; Ribeiro-Barros, A.I.; et al. Selenium biofortification of rice grains and implications on macronutrients quality. J. Cereal Sci. 2018, 81, 22–29. [Google Scholar] [CrossRef]
- Dinh, Q.T.; Cui, Z.; Huang, J.; Tran, T.A.T.; Wang, D.; Yang, W.; Zhou, F.; Wang, M.; Yu, D.; Liang, D. Selenium distribution in the Chinese environment and its relationship with human health: A review. Environ. Int. 2018, 112, 294–309. [Google Scholar] [CrossRef]
- Chilimba, A.D.C.; Young, S.D.; Black, C.R.; Meacham, M.C.; Lammel, J.; Broadley, M.R. Agronomic biofortification of maize with selenium (Se) in Malawi. Field Crop. Res. 2012, 125, 118–128. [Google Scholar] [CrossRef]
- Nawaz, F.; Naeem, M.; Ashraf, M.Y.; Tahir, M.N.; Zulfiqar, B.; Salahuddin, M.; Shabbir, R.N.; Aslam, M. Selenium supplementation affects physiological and biochemical processes to improve fodder yield and quality of maize (Zea mays L.) under water deficit conditions. Front. Plant Sci. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, C.; Park, Y.H.; Myoung, K.H.; Suh, M.K.; McArthur, T.; Lyons, G.; Stewart, D. The Bio-fortification of Barley with Selenium Catherine. In Proceedings of the Institute of Brewery & Distillating (Asia-Pacific Section) Carventron, Hobart, Tasmania, 19–24 March 2006. [Google Scholar]
- Wang, S.; Liang, D.; Wang, D.; Wei, W.; Fu, D.; Lin, Z. Selenium fractionation and speciation in agriculture soils and accumulation in corn (Zea mays L.) under field conditions in Shaanxi Province, China. Sci. Total Environ. 2012, 427–428, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–51. [Google Scholar] [CrossRef]
- Shanker, K.; Mishra, S.; Srivastava, S.; Srivastava, R.; Daas, S.; Prakash, S.; Srivastava, M.M. Effect of selenite and selenate on plant uptake and translocation of mercury by tomato (Lycopersicum esculentum). Plant. Soil 1996, 183, 233–238. [Google Scholar] [CrossRef]
- Guerrero, B.; Llugany, M.; Palacios, O.; Valiente, M. Dual effects of different selenium species on wheat. Plant. Physiol. Biochem. 2014, 83, 300–307. [Google Scholar] [CrossRef]
- Cubadda, F.; Aureli, F.; Ciardullo, S.; D’Amato, M.; Raggi, A.; Acharya, R.; Reddy, R.A.V.; Prakash, N.T. Changes in selenium speciation associated with increasing tissue concentrations of selenium in wheat grain. J. Agric. Food Chem. 2010, 58, 2295–2301. [Google Scholar] [CrossRef]
- Yin, H.; Qi, Z.; Li, M.; Ahammed, G.J.; Chu, X.; Zhou, J. Selenium forms and methods of application differentially modulate plant growth, photosynthesis, stress tolerance, selenium content and speciation in Oryza sativa L. Ecotoxicol. Environ. Saf. 2019, 169, 911–917. [Google Scholar] [CrossRef]
- Lidon, F.C.; Oliveira, K.; Galhano, C.; Guerra, M.; Ribeiro, M.M.; Pelica, J.; Pataco, I.; Ramalho, J.C.; Leitaõ, A.E.; Almeida, A.N.A.S.; et al. Selenium biofortification of rice through foliar application with selenite and selenate. Exp. Agric. 2019, 55, 528–542. [Google Scholar] [CrossRef]
- Perrino, E.V.; Calabrese, G. Endangered segetal species in southern Italy: Distribution, conservation status, trends, actions and ethnobotanical notes. Genet. Resour. Crop. Evol. 2018, 65, 2107–2134. [Google Scholar] [CrossRef]
- Hammer, K.; Gladis, T.; Diederichsen, A. Weeds as genetic resources. Plant. Genet. Rersour. Newsl. 1997, 111, 33–39. [Google Scholar]

{kind=link}
Table 1.
Different selenium (Se) forms and application methods in major cereal crops.
| Specie | Type of Experiment | Application | Time of Application | Se Form | g Se/ha | Control µg·kg−1 | Se Content in Grain µg·kg−1 DW | Increase | Increase by 1 g of Added Se | References |
|---|---|---|---|---|---|---|---|---|---|---|
| Rice | Field experiment | Soil | At heading | Selenite | 30 | 76 | 59 | 0.78 | 0.03 | [19] |
| Rice | Field experiment | Soil | At heading | Selenate | 30 | 76 | 79 | 1.04 | 0.03 | [19] |
| Rice | Field experiment | Soil | At heading | Selenite | 30 | 86 | 85 | 0.99 | 0.03 | [19] |
| Rice | Field experiment | Soil | At heading | Selenate | 30 | 86 | 92 | 1.07 | 0.04 | [19] |
| Rice | Field experiment | Soil | At heading | Selenite | 30 | 97 | 82 | 0.85 | 0.03 | [19] |
| Rice | Field experiment | Soil | At heading | Selenate | 30 | 97 | 92 | 0.95 | 0.03 | [19] |
| Rice | Field experiment | Foliar | At heading | Selenite | 30 | 76 | 273 | 3.59 | 0.12 | [19] |
| Rice | Field experiment | Foliar | At heading | Selenate | 30 | 76 | 150 | 1.97 | 0.07 | [19] |
| Rice | Field experiment | Foliar | At heading | Selenite | 30 | 86 | 122 | 1.42 | 0.05 | [19] |
| Rice | Field experiment | Foliar | At heading | Selenate | 30 | 86 | 105 | 1.22 | 0.04 | [19] |
| Rice | Field experiment | Foliar | At heading | Selenite | 30 | 97 | 136 | 1.4 | 0.05 | [19] |
| Rice | Field experiment | Foliar | At heading | Selenate | 30 | 97 | 176 | 1.81 | 0.06 | [19] |
| Rice | Field experiment, no till | Soil | At sowing | Selenate | 25 | 30 | 320 | 10.67 | 0.43 | [28] |
| Rice | Plastic pots in growth chamber | Foliar | Seven times through vegetation | Selenite | 0.53 | 30 | 100 | 3.33 | 6.35 | [29] |
| Rice | Plastic pots in growth chamber | Foliar | Seven times through vegetation | Selenite | 10.5 | 30 | 1540 | 51.33 | 4.89 | [29] |
| Rice | Plastic pots in growth chamber | Foliar | Seven times through vegetation | Selenite | 21 | 30 | 1560 | 52 | 2.48 | [29] |
| Maize | Field experiment | Soil | Before sowing | Selenite | 150 | 3.7 | 51 | 13.78 | 0.09 | [106] |
| Maize | Field experiment | Foliar | Tasseling and one week after silking | Selenite | 11 | 11 | 96 | 8.73 | 0.79 | [106] |
| Maize | Field experiment | Soil | Before seeding | Selenate | 5 | 34 | 41.66 | 1.23 | 0.25 | [30] |
| Maize | Field experiment | Soil | Before seeding | Selenate | 10 | 34 | 68.33 | 2.01 | 0.2 | [30] |
| Maize | Field experiment | Soil | Before seeding | Selenate | 20 | 34 | 92.66 | 2.73 | 0.14 | [30] |
| Maize | Field experiment | Foliar | During the stem elongation stage | Selenate | 5 | 34 | 156.66 | 4.61 | 0.92 | [30] |
| Maize | Field experiment | Foliar | During the stem elongation stage | Selenate | 10 | 34 | 205.33 | 6.04 | 0.6 | [30] |
| Maize | Field experiment | Foliar | During the stem elongation stage | Selenate | 20 | 34 | 305.66 | 8.99 | 0.45 | [30] |
| Barley | Field experiment | Soil | Before seeding | Selenite | 20 | 45 | 57 | 1.27 | 0.06 | [31] |
| Barley | Field experiment | Soil | Before seeding | Selenate | 20 | 33 | 391 | 11.85 | 0.59 | [31] |
| Barley | Field experiment | Soil | Before seeding | Selenite | 40 | 45 | 76 | 1.69 | 0.04 | [31] |
| Barley | Field experiment | Soil | Before seeding | Selenate | 40 | 33 | 959 | 29.06 | 0.73 | [31] |
| Barley | Field experiment | Foliar | End of tillering EC-39 | Selenate | 10 | 111.7 | 880 | 7.88 | 0.79 | [32] |
| Barley | Field experiment | Foliar | End of tillering EC-39 | Selenate | 20 | 111.7 | 1113.9 | 9.97 | 0.5 | [32] |
| Barley | Field experiment | Foliar | End of tillering EC-39 | Selenite | 10 | 111.7 | 270 | 2.42 | 0.24 | [32] |
| Barley | Field experiment | Foliar | End of tillering EC-39 | Selenite | 20 | 111.7 | 345.5 | 3.09 | 0.15 | [32] |
| Wheat | Field experiment | Foliar | Tillering state | Selenite | 10 | 66.6 | 153.6 | 2.31 | 0.23 | [33] |
| Wheat | Field experiment | Foliar | Tillering state | Selenite | 20 | 66.6 | 254.8 | 3.83 | 0.19 | [33] |
| Wheat | Field experiment | Foliar | Tillering state | Selenite | 40 | 66.6 | 430.4 | 6.46 | 0.16 | [33] |
| Wheat | Field experiment | Foliar | Tillering state | Selenate | 10 | 66.6 | 266.8 | 4.01 | 0.4 | [33] |
| Wheat | Field experiment | Foliar | Tillering state | Selenate | 20 | 66.6 | 820 | 12.31 | 0.62 | [33] |
| Wheat | Field experiment | Foliar | Tillering state | Selenate | 40 | 66.6 | 1383.2 | 20.77 | 0.52 | [33] |
| Wheat | Small-plot field experiment | Foliar | Growth stage of 2nd node on the main stem | Selenite | 10 | 32 | 51 | 1.59 | 0.16 | [34] |
| Wheat | Small-plot field experiment | Foliar | Growth stage of 2nd node on the main stem | Selenite | 20 | 32 | 72 | 2.25 | 0.11 | [34] |
| Wheat | Small-plot field experiment | Foliar | Growth stage of 2nd node on the main stem | Selenate | 10 | 32 | 190 | 5.94 | 0.59 | [34] |
| Wheat | Small-plot field experiment | Foliar | Growth stage of 2nd node on the main stem | Selenate | 20 | 32 | 350 | 10.94 | 0.55 | [34] |
| Wheat | Field experiment | Foliar | Preflowering | Selenite | 20 | 120 | 610 | 5.08 | 0.25 | [35] |
| Wheat | Field experiment | Foliar | Preflowering | Selenate | 20 | 120 | 1340 | 11.17 | 0.56 | [35] |
| Wheat | Field experiment | Foliar | Pre-grain filling stages | Selenite | 20 | 120 | 970 | 8.08 | 0.4 | [35] |
| Wheat | Field experiment | Foliar | Pre-grain filling stages | Selenate | 20 | 120 | 1590 | 13.25 | 0.66 | [35] |
| Wheat | Field experiment | Foliar | During GS 31 and GS 49 stage | Selenate | 5 | 150 | 640 | 4.27 | 0.85 | [20] |
| Wheat | Field experiment | Foliar | During GS 31 and GS 49 stage | Selenate | 25 | 150 | 2390 | 15.93 | 0.64 | [20] |
| Wheat | Field experiment | Foliar | During GS 31 and GS 49 stage | Selenate | 50 | 150 | 2820 | 18.8 | 0.38 | [20] |
| Wheat | Field experiment | Foliar | During GS 31 and GS 49 stage | Selenate | 80 | 150 | 3930 | 26.2 | 0.33 | [20] |
Table 2.
Variance explained by application type × Se form, species × Se form, and type of experiment, followed by coefficients of random effects (best linear unbiased predictions) for each level of factors from the 11 scientific papers evaluated (n.a., not applicable).
Table 2.
Variance explained by application type × Se form, species × Se form, and type of experiment, followed by coefficients of random effects (best linear unbiased predictions) for each level of factors from the 11 scientific papers evaluated (n.a., not applicable).
| Factor | Application Type × Se Form | Species × Se Form | Type of Experiment | Residual |
|---|---|---|---|---|
| Variance | 0.012352 | 0.006132 | 10.624367 | 0.213215 |
| Coefficients of random effects | ||||
| Soil/selenate | −0.007177254 | – | – | |
| Foliar/selenate | 0.095549031 | – | – | |
| Soil/selenite | −0.060080039 | – | – | n.a. |
| Foliar/selenite | −0.022608397 | – | – | |
| Barley/selenate | – | 0.031985788 | – | |
| Maize/selenate | – | 0.012493402 | – | |
| Rice/selenate | – | −0.039315129 | – | |
| Wheat/selenate | – | 0.038706521 | – | |
| Barley/selenite | – | −0.013442636 | – | |
| Maize/selenite | – | 0.010163323 | – | |
| Rice/selenite | – | −0.028004366 | – | |
| Wheat/selenite | – | −0.009765509 | – | |
| Field experiment | – | – | 0.2971243 | |
| Plastic pots in growth chamber | – | – | 4.5913276 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Galić, L.; Vinković, T.; Ravnjak, B.; Lončarić, Z. Agronomic Biofortification of Significant Cereal Crops with Selenium—A Review. Agronomy 2021, 11, 1015. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11051015
AMA Style
Galić L, Vinković T, Ravnjak B, Lončarić Z. Agronomic Biofortification of Significant Cereal Crops with Selenium—A Review. Agronomy. 2021; 11(5):1015. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11051015
Chicago/Turabian StyleGalić, Lucija, Tomislav Vinković, Boris Ravnjak, and Zdenko Lončarić. 2021. "Agronomic Biofortification of Significant Cereal Crops with Selenium—A Review" Agronomy 11, no. 5: 1015. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11051015
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.