1. Introduction
In the new global agricultural system, biofertilizer technology has become a pivotal subject in crop productivity enrichment. Aside from the potential of reducing synthetic agrochemicals input, the technology is profoundly tailored with specific plant beneficial microorganisms which have been exploited for crop nutrition and protection. Our previous report has demonstrated the potential of
Enterobacter hormaechei 40a with multiple plant growth-promoting (PGP) traits particularly phosphate (P) solubilizing activity to improve the early vegetative growth of okra [
1]. However, the efficacy of strain 40a in a long-term greenhouse setting and on the field is still unknown. It has become a real prospective challenge particularly concerning its non-spore forming characteristic. Contrary to its sporulating counterparts, strain 40a such as other Gram-negative bacteria is more susceptible to detrimental conditions occurring during storage, processing, handling and field application [
2]. Therefore, an ideal and protective formulation of biofertilizer enriched with strain 40a is of paramount importance.
In many cases, low-grade formulations and the concurrent poor inoculant quality are still considered as the main constraints to the effective, extensive use of biofertilizers [
3]. The supply of a high viable cell density to the plant is a prerequisite to achieve an effective colonization rate, which in turn boosts the anticipated PGP effects. Any stages before and during the application of biofertilizers may compromise the viable cell density of inoculants. For instance, a biofertilizer product should exhibit a sufficiently long shelf life, which translates its stability during the whole process of production, packaging, storage and transportation [
4]. Obviously, during soil application, inoculant strains are exposed to various biotic (e.g., native microflora and microfauna) and abiotic stress conditions (e.g., soil pH, temperature, salinity) as compared to during pre-application stages [
5]. In view of these limitations, the pre-application stages of biofertilizers, i.e., formulation and storage, should be prioritized since the quality of products in this stage can be controlled and monitored periodically. This is especially important to maintain the quality and sustain the efficacy of the establishment of biofertilizers at the target site and to facilitate the optimal application by farmers.
A high-quality biofertilizer formulation must be able to enhance inoculant cell density during production and maintain an adequately prolonged shelf life during storage [
6]. In addition to cell growth and preservation, the high nutrient constituents in the formulation are valuable to crop and soil application. Inoculant strains highly depend on the carbon (C) and nitrogen (N) sources included in the growth medium for cell maintenance, reproduction and product synthesis, particularly during the storage phase. Among the most common sources used to produce high inoculant strain biomass are semi-synthetic media such as skim-milk, yeast extract, carboxymethylcellulose, etc. Despite the fact that those commercial sources are inextricably effective and have been widely used, they add up to the capital cost required for production [
7]. Alternatively, the C and N elements can be obtained from a wide array of inexpensive and natural biomass including agricultural residues [
8,
9], industrial waste [
10], animal manure [
11], etc. While the utilization of various raw materials offers an ecologically sustainable solution to natural waste management, it also represents a tremendous opportunity to recycle nutrients that can nourish the soils, especially those of tropical regions that present low fertility conditions [
12].
Molasses is a viscous and dark syrup generated abundantly as a by-product in the sugar beet and sugarcane processing industry [
7]. It is a renewable resource that consists of 40 to 60% complex sugars, hence suitable as a C source formulation of biofertilizers. Additionally, it also contains humic substances which are the major components of organic matter beneficial for plant growth [
13]. Previously, it has shown potential as a component of biofertilizer for
Rhizobium sp. production for leguminous crops [
7], apart from the production of high-cell density fed-batch of
Saccharomyces cerevisiae culture for bioethanol synthesis [
14]. Defatted soybean meal (DSM), an abundant by-product in the soybean oil industry, has been extensively used as protein diets for animals, particularly for poultry and swine [
15]. DSM is comprised of 50% protein, 25–30% carbohydrate and other micronutrients [
16] which makes it an excellent source of organic N composition in biofertilizers. Recently, it has been utilized in myriads biotechnological application in-vitro, i.e., nisin production by a genetically engineered proteolytic system of
Lactococcus lactis, fermented soybean meal production as animal feed using
Lactobacillus sp. and
Clostridium butyricum, fumaric acid production by
Rhizopus oryzae [
15,
17,
18].
Apart from the sufficient cell counts of desired inoculant strains and the nutrient-rich components, liquid biofertilizers also incorporate essential cell protectants or additives in the formulation that promote prolonged shelf-life and protect against adverse conditions [
5,
19]. The common cell protectants include natural polymers, e.g., Arabic gum, xanthan gum, gelatin, sodium alginate (SA), and synthetic polymers, e.g., polyvinyl alcohol, horticultural oil, glycerol (Gly), monosaccharides and disaccharides such as lactose [
20,
21]. Thirumal et al. [
22] reported that the high viable cell of
Rhizobium biofertilizer was observed in SA formulation at room temperature after eight months of storage. Similarly,
Rhodopseudomonas palustris was also found to survive well and maintain at around 10 log CFU/mL in a growth medium amended with 0.5% SA [
23]. They also reported that the amendment with 1% Gly provided extra protection for bacterial cells against temperature stress.
Therefore, the objectives of this research were to: (1) investigate the biocompatibility of molasses as a basal nutrient composition and screen for supplementary C and N sources required to enhance cell density of strain 40a, (2) develop and optimize a medium using molasses and DSM to achieve maximum cell density of strain 40a, (3) evaluate the P solubilizing activity of strain 40a in the optimized medium, (4) amend the optimized medium with suitable cell protectants to improve shelf-life of strain 40a.
2. Materials and Methods
2.1. Characterization of Molasses and DSM
Unsulfured molasses and DSM were purchased from Anizam Resources Sdn. Bhd., Shah Alam, Selangor, Malaysia. The raw samples were transported to the laboratory and stored in a tight container at room temperature prior to analysis. The pH of the sample was determined using a pH-meter (Sartorius PB-10, Göttingen, Germany) by diluting at a ratio of 1:1 v/v with distilled water at ambient temperature. The moisture content was determined by oven-drying at 70 °C until a constant dry weight was obtained. Sugar profile was determined using High-Performance Liquid Chromatography (HPLC) Waters Alliance 2695 HPLC Separations Module (ELS 2410) System (Waters Corporation, Milford, MA, USA) with Waters 2410 RI detector against standards of 1 mg/mL sucrose, glucose, fructose, galactose, mannose solution (Sigma-Aldrich, Darmstadt, Germany). The dried sample was digested for elemental analysis using concentrated nitric acid at a ratio of 1:10 w/v on a digester for about 10 min until brown fume disappeared. The trace elements were determined using inductively coupled plasma-optical emission spectrometry (ICP–OES) Optima 7300 DV (Perkin Elmer, Waltham, MA, USA). The organic C, N and sulfur (S) content of the dried sample were determined using TruMac CNS Analyzer (Leco Corporation, St. Joseph, MI, USA).
2.2. Test Bacteria and Fermentation Condition
E. hormaechei 40a strain was obtained from the bacterial culture collection at Research Laboratory 1.6, Bioprocessing and Biomanufacturing Research Centre, Universiti Putra Malaysia. The growth of strain 40a was maintained by routine subculture on nutrient agar to acquire a pure single colony. Unless stated otherwise, the fermentation condition of the starter culture (nutrient broth) and other fermentation media were prepared in 250 mL conical flasks containing 100 mL liquid medium at neutral pH (6.8–7.0) and the bacteria was grown at 37 °C, shaking at 200 rpm for 24 h.
2.3. C and N Sources Supplementation
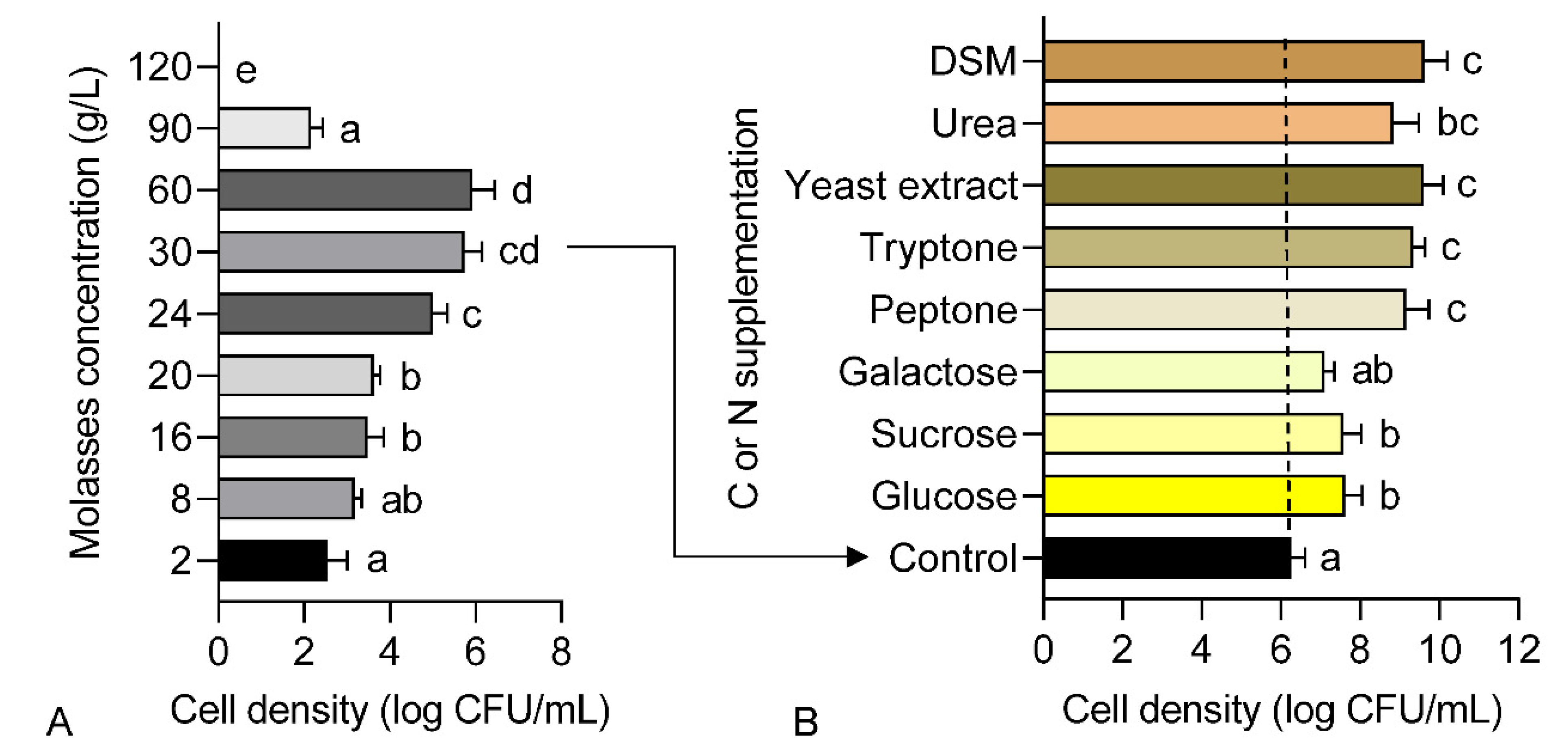
The unsulfured molasses was used as a basic nutrient source for growth culture throughout the whole experiment. A preliminary test was performed beforehand to evaluate the biocompatibility of molasses as the sole nutrient source to culture strain 40a with different concentrations ranging from 2 to 120 g/L. Nutrient broth was used as a starter culture and a standard inoculum size (~1 × 1010 CFU/mL) was used to inoculate the fermentation media unless stated otherwise. For single-factor screening of C and N supplementation, the molasses-based media were separately supplemented with different C sources, i.e., glucose, sucrose, galactose and N sources, i.e., peptone, tryptone, yeast extract and urea at a fixed concentration of 10 g/L respectively.
2.4. Cell Density Enumeration
The standard plate count method was used to determine the bacterial cell density. An aliquot of 100 μL culture was transferred into a microcentrifuge tube containing 900 μL of sterile phosphate-buffered saline (PBS). A routine serial dilution was performed and 10 μL of every dilution was pipetted onto nutrient agar. The inverted agar plate was incubated in a static incubator (Memmert GmbH + Co.KG, Schwabach, Germany) at 37 °C for 24 h. The bacterial colonies were counted manually by using a colony counter (Stuart, UK). Cell density was recorded in a log form (log CFU/mL).
2.5. Two-Level Factorial Design (TFLD)
A two-level factorial design (TFLD) was employed to screen for the significant factors [
24] influencing the cell growth of strain 40a. Nine factors (2
9-4 factorial design, 32 experimental runs) were evaluated in this test which were molasses, DSM, yeast extract, glucose, sucrose, pH, temperature, inoculum size, and agitation with a response measured in cell density. Each factor was evaluated at two levels coded as high levels (+1) and low levels (−1) in the experimental design using Design of Expert (DOE) software (Version 11, Stat-Ease, Inc., Minneapolis, MN, USA). The symbols, units and actual levels of the variables were shown in
Table 1.
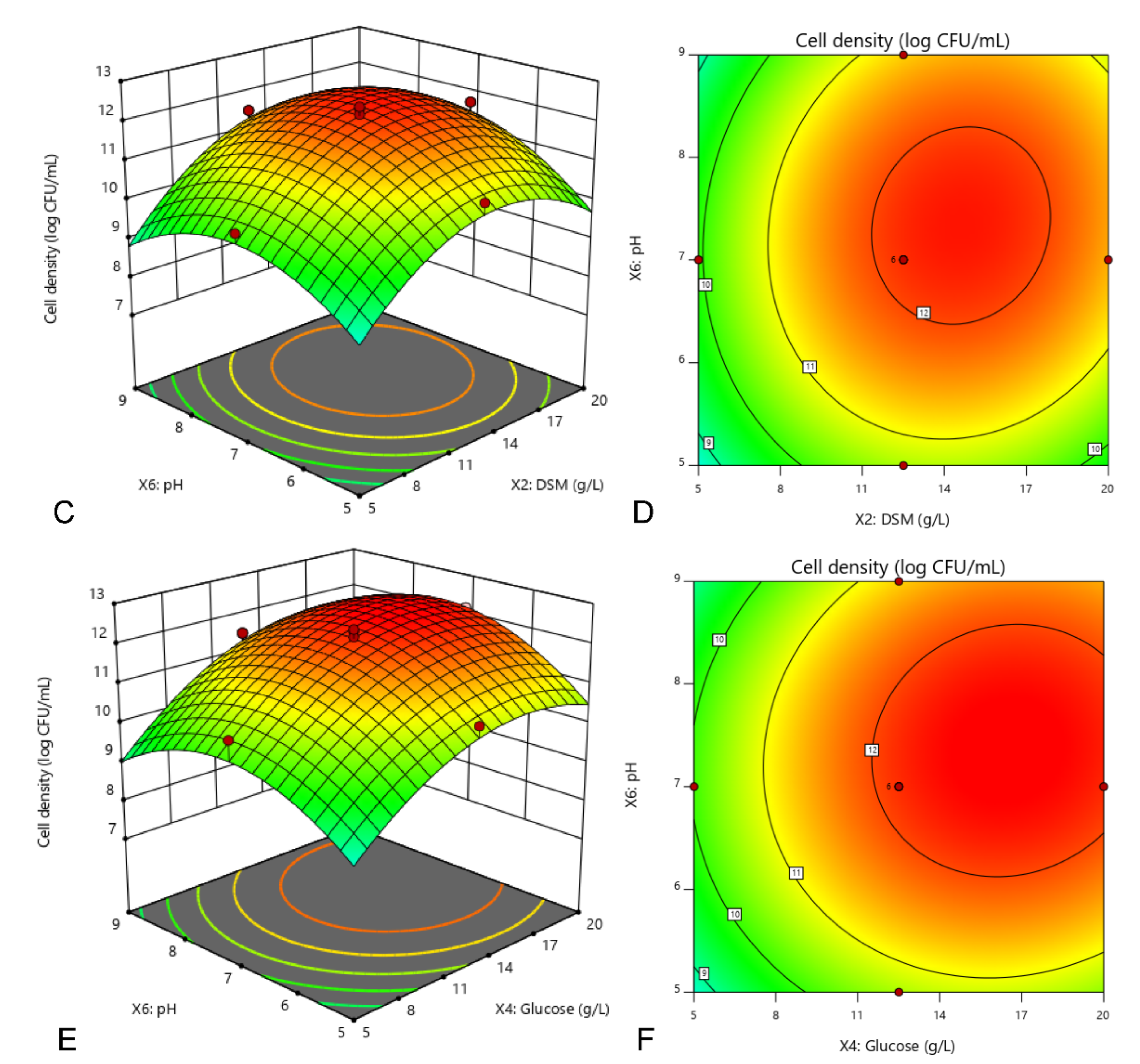
2.6. Central Composite Design (CCD) Using Response Surface Methodology (RSM)
A central composite design (CCD) [
25] of response surface methodology (RSM) was employed to optimize the culture conditions selected from the most significant (
p < 0.05) C source, N source and physical factor respectively, assessed in the previous TLFD test. In this study, the experimental design consisted of 20 experimental runs with 6 replicates at the center point. The behavior of the system was established by assuming a second-order polynomial function with linear, quadratic and interaction effects as shown in Equation (1).
where
Y is the predicted result,
β0 (offset term),
βi (linear effect),
βii (squared effect), and
βij (interaction effect) are constant coefficients, and
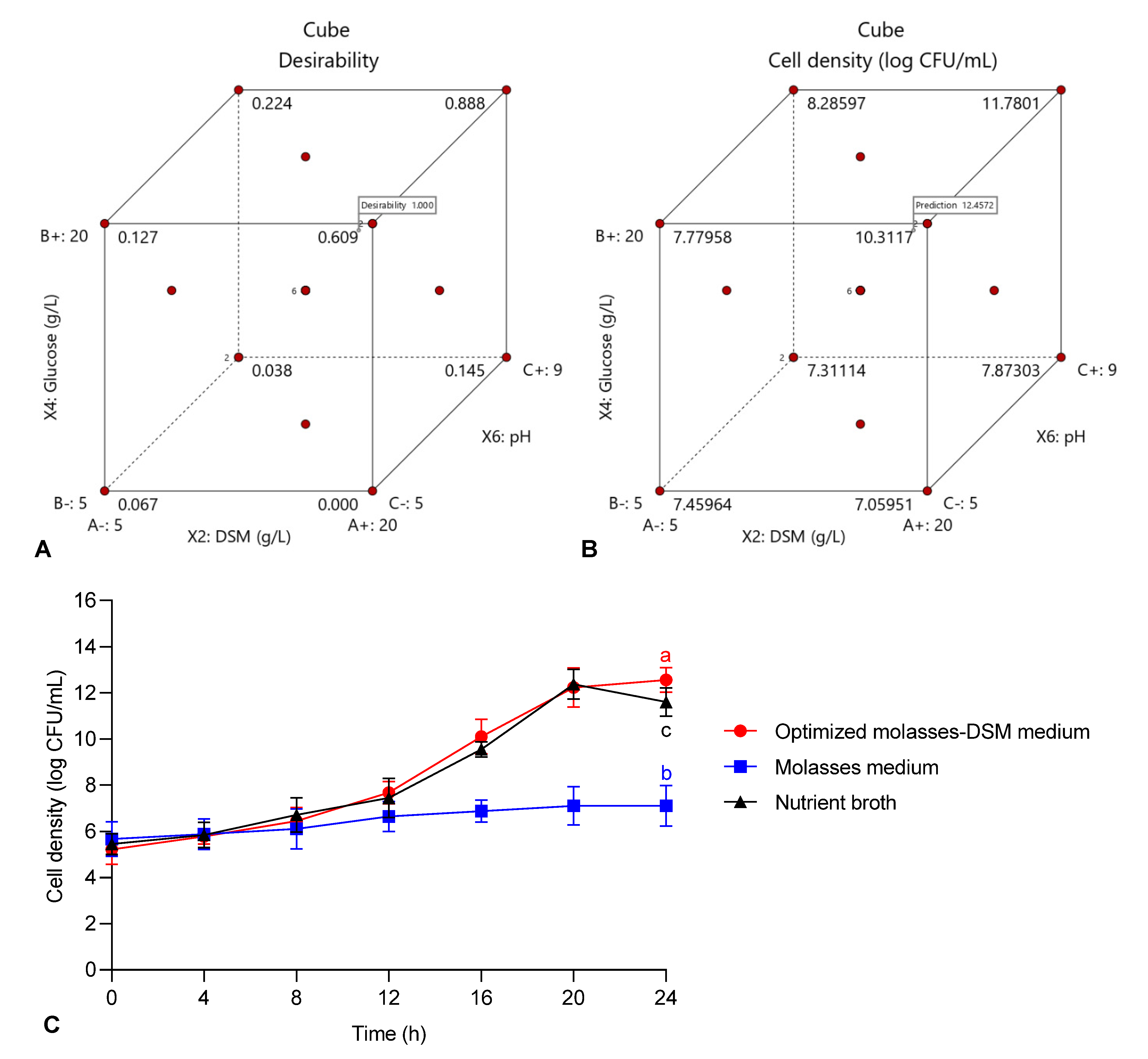
X shows the coded level of the independent variable. The regression analysis of variance and preparation of response surface graphs were implemented using Design of Expert (DOE) software (Version 11, Stat-Ease, Inc., Minneapolis, MN, USA). The optimal culture conditions for maximum cell density of
E. hormaechei 40a were estimated by statistical analysis.
2.7. Post-Analysis of the Optimized Model
The post-analysis of the optimized formulation model was performed according to the parameters suggested by the DOE algorithm to achieve maximum cell density of strain 40a. The bacterial cell density was grown in a basic molasses medium and a commercial nutrient broth were concurrently compared with the optimized molasses-DSM (OMD) medium (final composition: DSM, 13.8 g/L; glucose, 17.7 g/L; initial pH, 7.6). The samples were withdrawn every 4 h and analyzed for cell density using previously described standard plate count method.
2.8. P Solubilization Activity of E. hormaechei 40a in Different Media
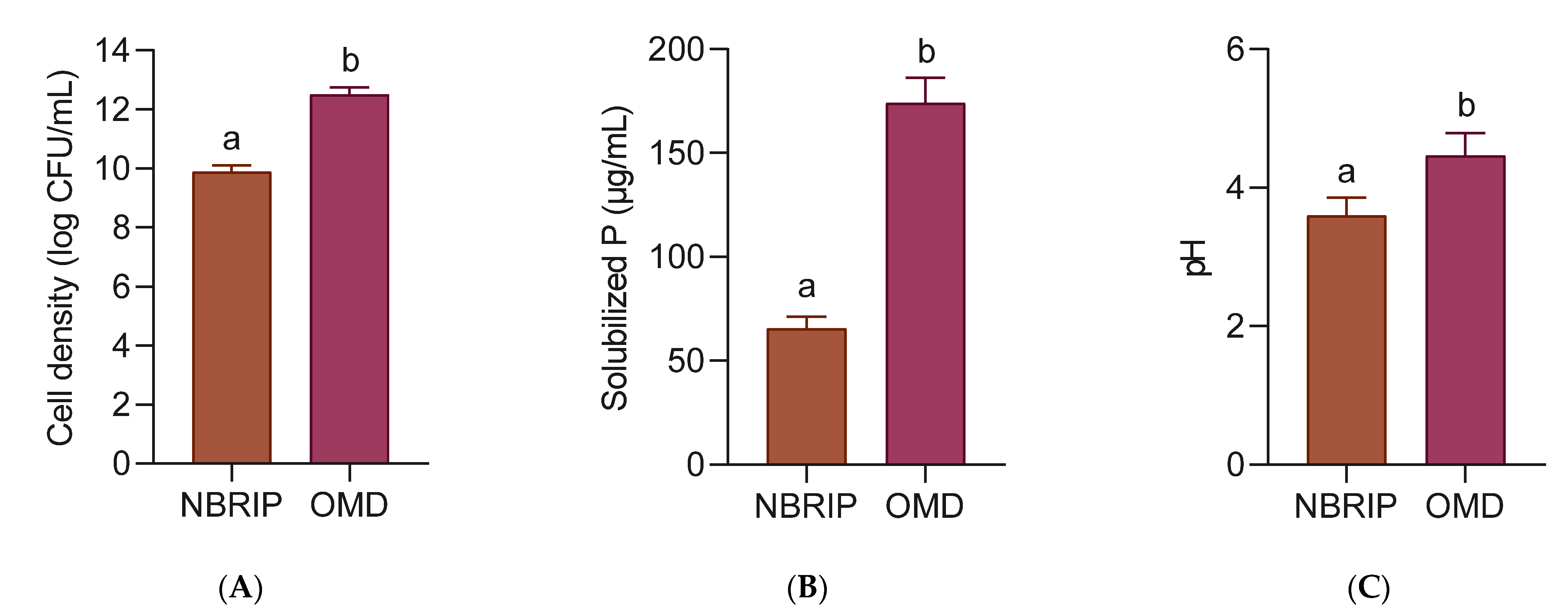
Strain 40a was grown in the OMD medium and the National Botanical Research Institute’s Phosphate (NBRIP) medium in a 250 mL Erlenmeyer flask separately for 24 h. NBRIP was prepared according to the standard composition excluding tricalcium phosphate [
26]. Both media were added with 5 g/L Christmas island rock phosphate (CIRP) before autoclaved. The soluble P was determined by the yellow phospho-molybdo-vanadate colorimetry [
27] using a UV-VIS spectrophotometer (Uviline 9400, Secomam, Alès France) against a KH
2PO
4 standard curve. The pH of the medium was measured using a benchtop pH meter (pH 700, Eutech Instruments, Ayer Rajah Crescent, Singapore). The viable cell density was determined using previously described standard plate count method.
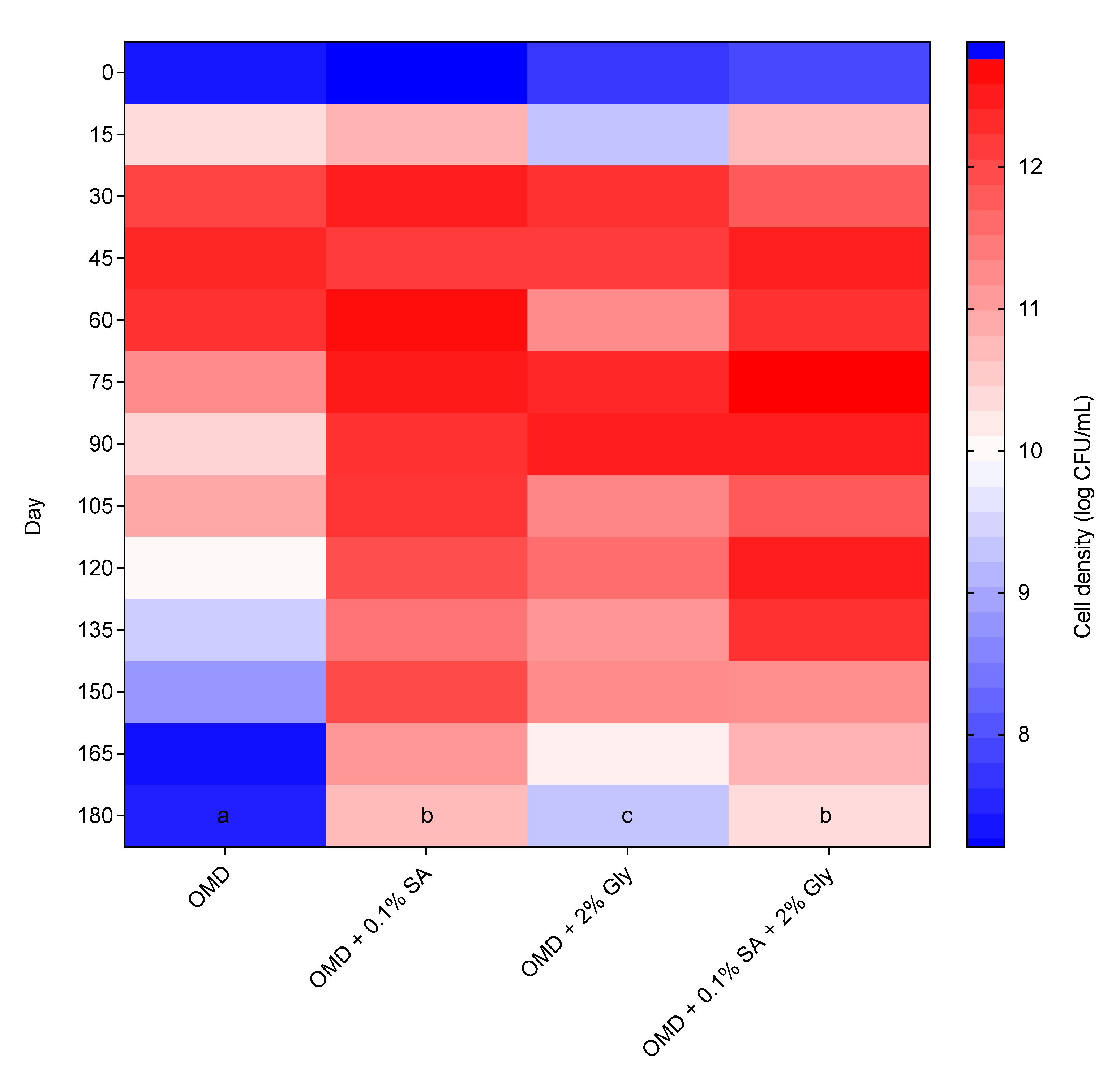
2.9. Shelf-Life Determination
The optimized culture formulation was prepared as stated previously and loaded half-full into a 100 mL UV-sterilized high-density polyethylene (HDPE) bottle. The effects of the addition of cell protectants, i.e., 0.1% sodium alginate (SA) [
19] and 2% glycerol (Gly) [
28] to the media were also investigated. The bottles were stored at room temperature (28 ± 2 °C) in a static condition. The shelf-life of the optimized culture was monitored by bimonthly interval up to 180 days of storage using the standard plate count method.
2.10. Statistical Analysis
Statistical tests were performed using one-way analysis of variance (ANOVA) to determine the significance of each variable employed in the preliminary single-factor screening and the shelf-life determination experiment. The regression coefficients were determined by Tukey-test as a post hoc pairwise comparison at p < 0.05. The significant difference between soluble P, pH and cell density between NBRIP and OMD media were determined using unpaired t-test at p < 0.05.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}