
Responses of Peppermint and Spearmint Crops to Excessive Biostimulant Application and Increased Salinity in a Closed Soilless Production System

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth
2.2. Antioxidant Capacity and Soluble Phenolic Content
2.3. Lipid Peroxidation
2.4. Hydrogen Peroxide
2.5. Proline
2.6. Antioxidant Enzyme Activities
2.7. Experimental Design and Statistical Analysis
3. Results and Discussion
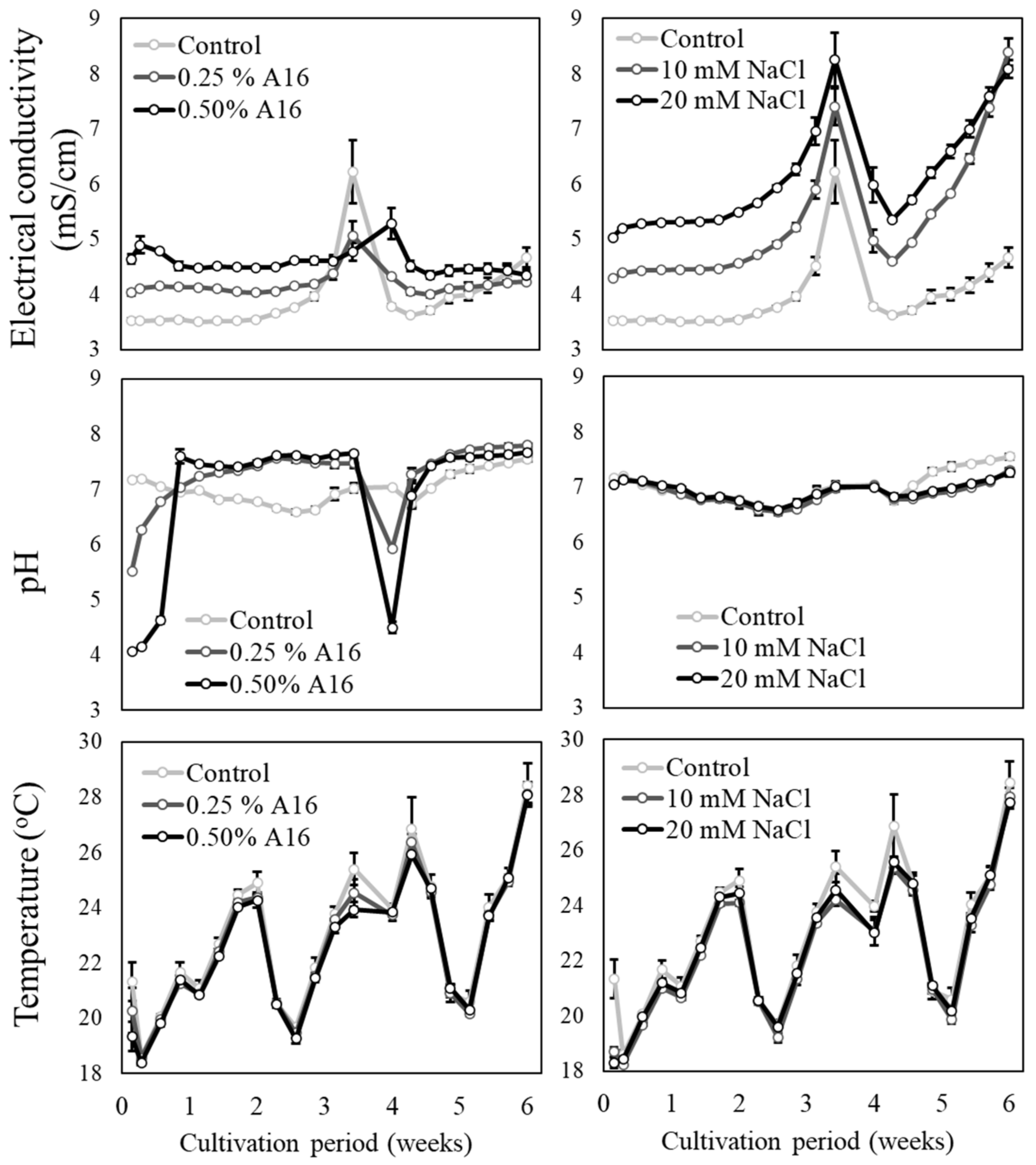
3.1. EC, pH, and Temperature of the Nutrient Solution
3.2. Plant Growth
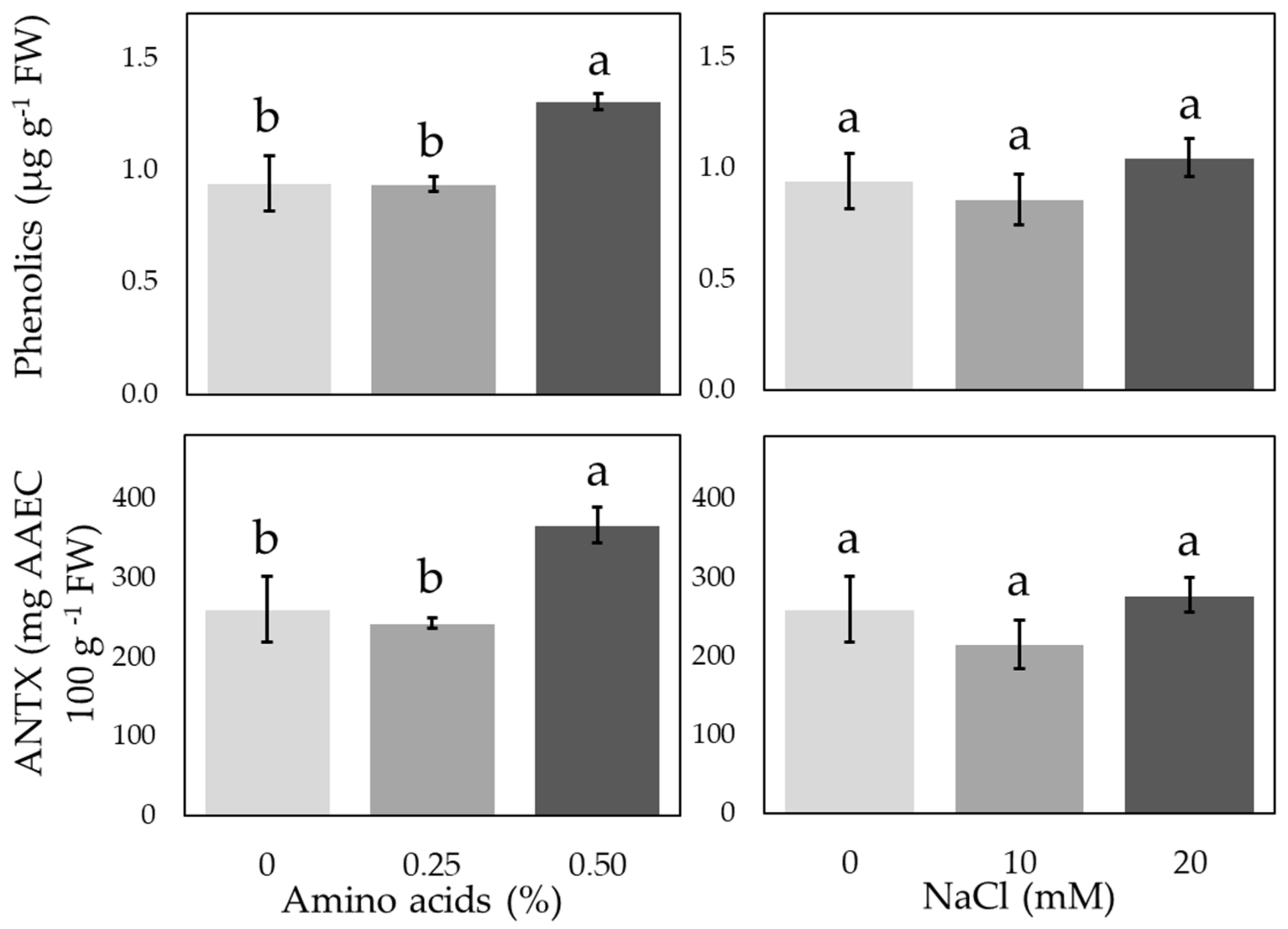
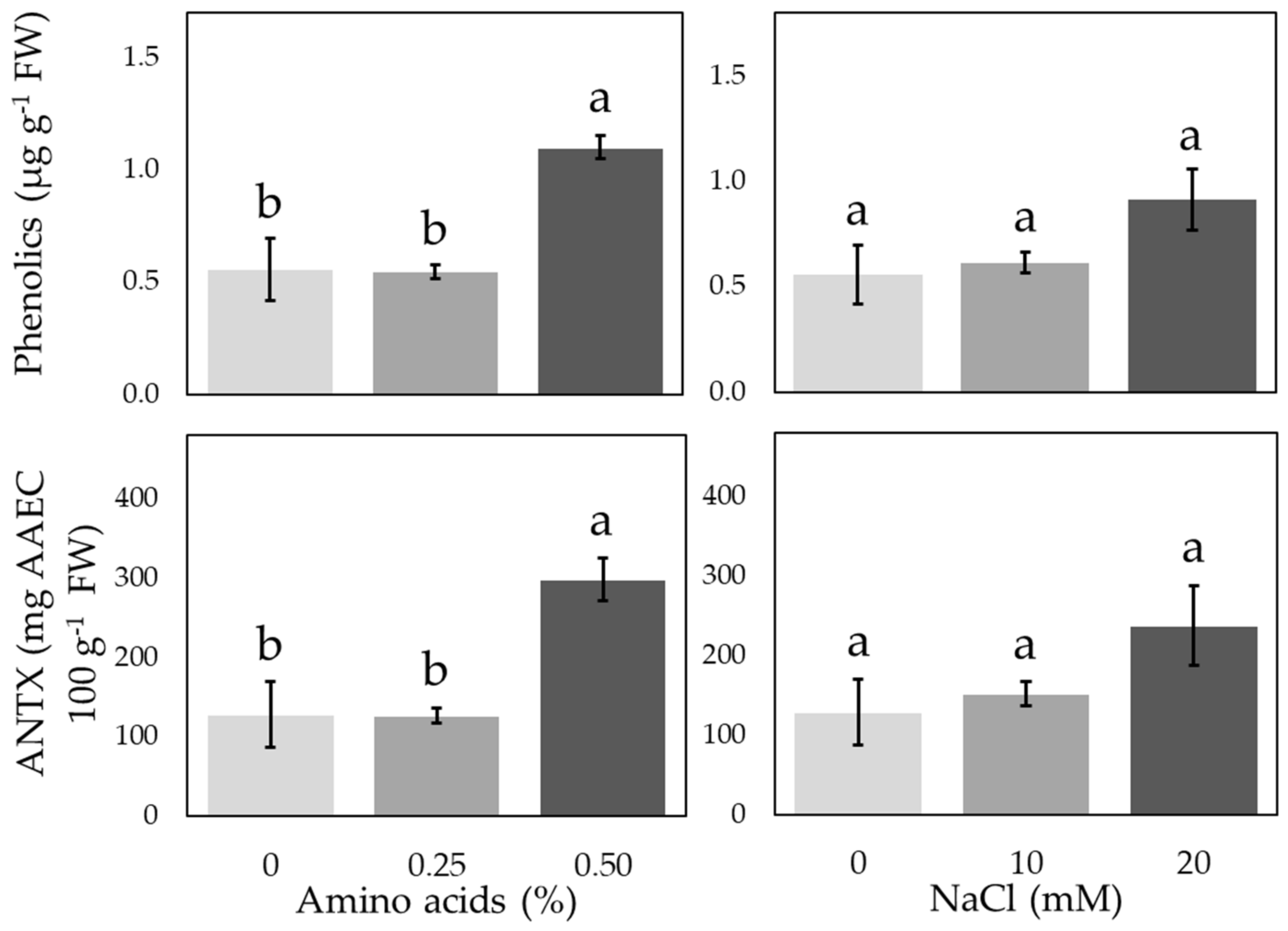
3.3. Antioxidant Compounds and Enzymatic Activities
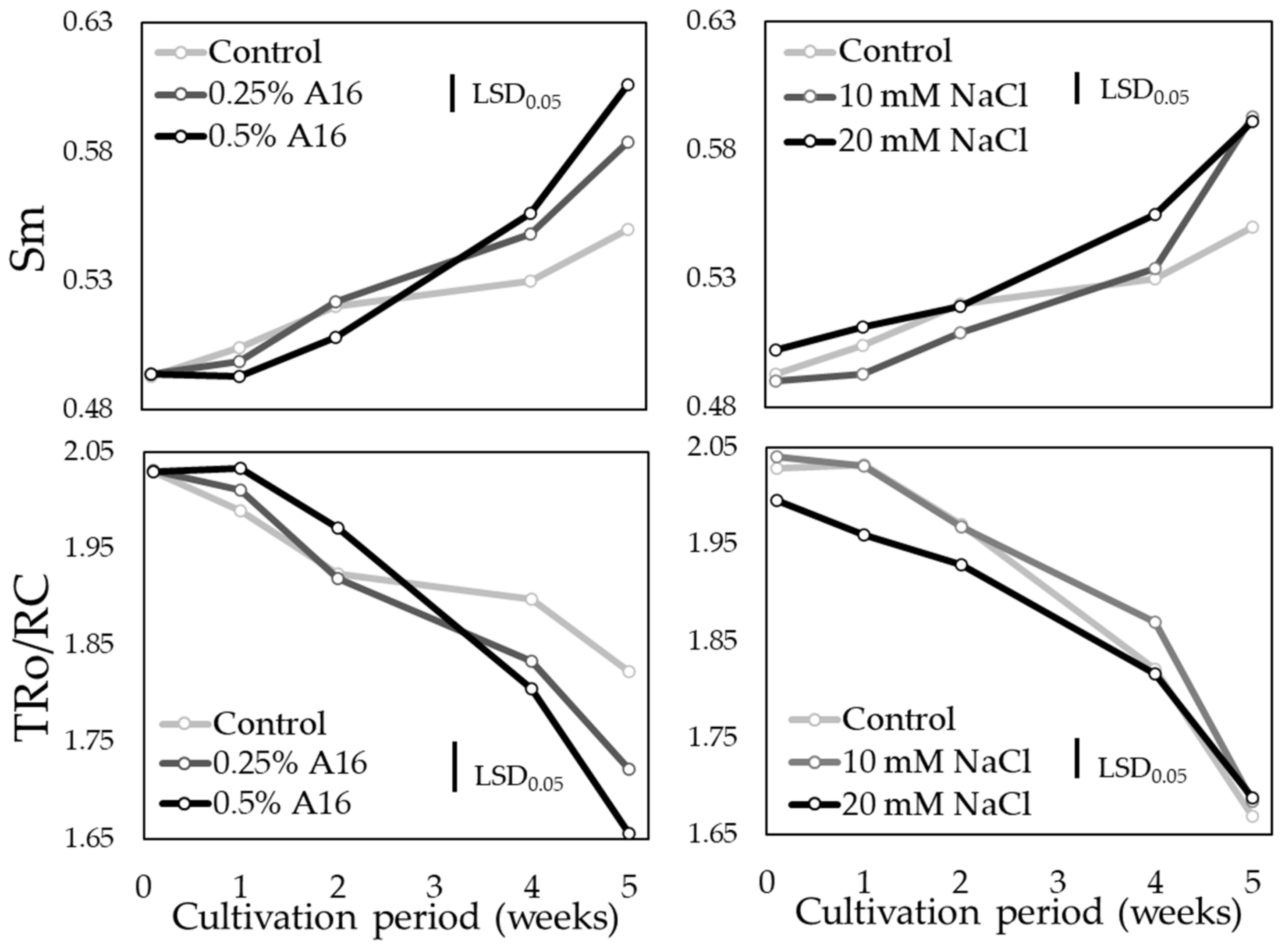
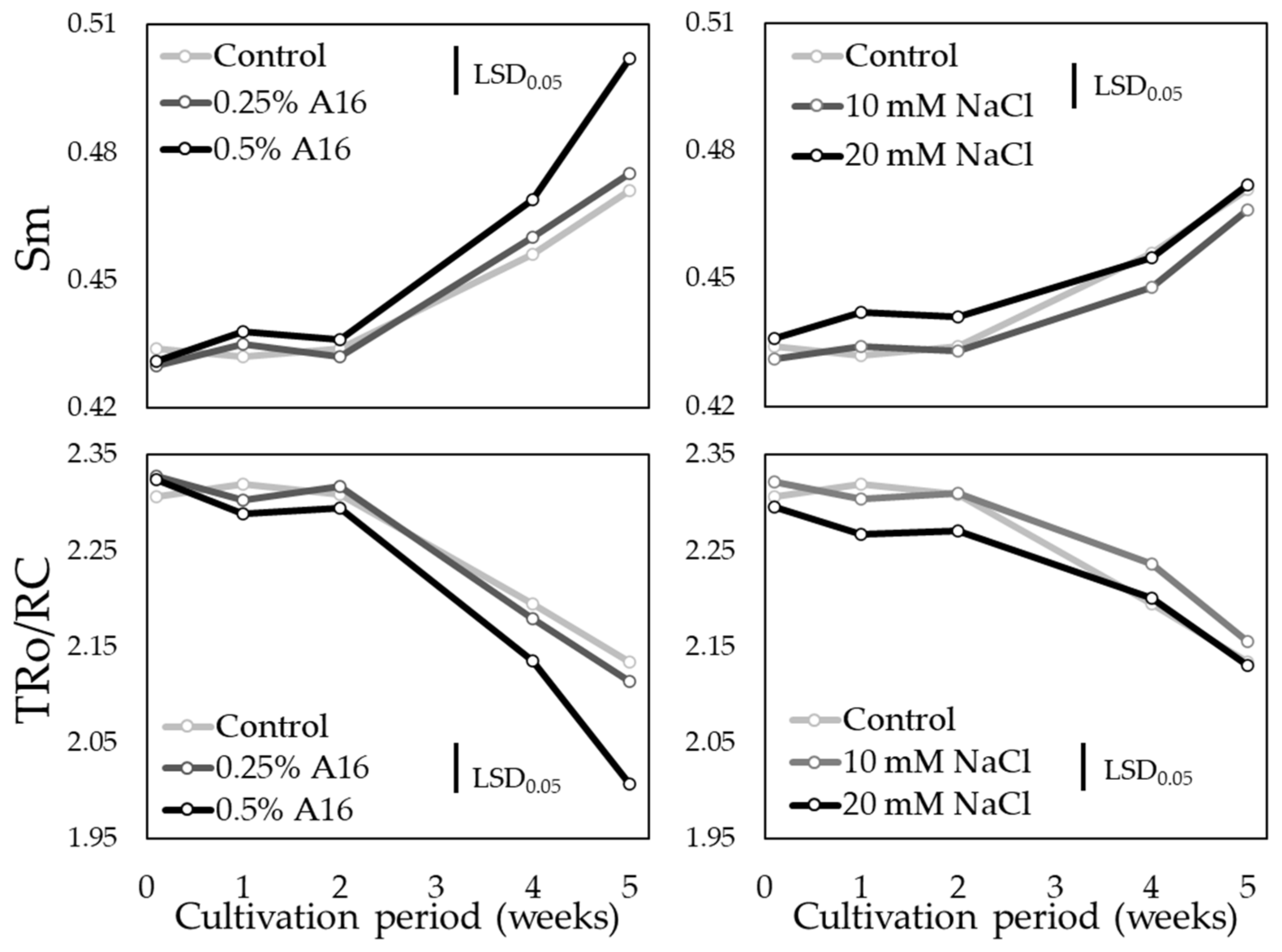
3.4. Chlorophyll Fluorescence
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bahadori, M.B.; Zengin, G.; Bahadori, S.; Dinparast, L.; Movahhedin, N. Phenolic composition and functional properties of wild mint (Mentha longifolia var. calliantha (Stapf) Briq.). Int. J. Food Prop. 2018, 21, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Pavlić, B.; Teslić, N.; Zengin, G.; Đurović, S.; Rakić, D.; Cvetanović, A.; Gunes, A.K.; Zeković, Z. Antioxidant and enzyme-inhibitory activity of peppermint extracts and essential oils obtained by conventional and emerging extraction techniques. Food Chem. 2021, 338, 127724. [Google Scholar] [CrossRef] [PubMed]
- Cam, M.; Basyigit, B.; Alasalvar, H.; Yilmaztekin, M.; Ahhmed, A.; Sagdic, O.; Konca, Y.; Telci, I. Bioactive properties of powdered peppermint and spearmint extracts: Inhibition of key enzymes linked to hypertension and type 2 diabetes. Food Biosci. 2020, 35, 100577. [Google Scholar] [CrossRef]
- Teng, Z.Q.; Shen, Y. Research progress of genetic engineering on medicinal plants. China J. Chin. Mater. Med. 2015, 40, 594–601. [Google Scholar]
- Aktsoglou, D.-C.; Kasampalis, D.S.; Sarrou, E.; Tsouvaltzis, P.; Chatzopoulou, P.; Martens, S.; Siomos, A.S. Protein hydrolysates supplement in the nutrient solution of soilless grown fresh peppermint and spearmint as a tool for improving product quality. Agronomy 2021, 11, 317. [Google Scholar] [CrossRef]
- Ceccarelli, A.V.; Miras-Moreno, B.; Buffagni, V.; Senizza, B.; Pii, Y.; Cardarelli, M.; Rouphael, Y.; Colla, G.; Lucini, L. Foliar application of different vegetal-derived protein hydrolysates distinctively modulates tomato root development and metabolism. Plants 2021, 10, 326. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Tsouvaltzis, P.; Kasampalis, D.S.; Aktsoglou, D.C.; Barbayiannis, N.; Siomos, A.S. Effect of reduced nitrogen and supplemented amino acids nutrient solution on the nutritional quality of baby green and red lettuce grown in a floating system. Agronomy 2020, 10, 922. [Google Scholar] [CrossRef]
- Sitohy, M.Z.; Desoky, E.-S.M.; Osman, A.; Rady, M.M. Pumpkin seed protein hydrolysate treatment alleviates salt stress effects on Phaseolus vulgaris by elevating antioxidant capacity and recovering ion homeostasis. Sci. Hortic. 2020, 271, 109495. [Google Scholar] [CrossRef]
- Boselli, M.; Bahouaoui, M.A.; Lachhab, N.; Sanzani, S.M.; Ferrara, G.; Ippolito, A. Protein hydrolysates effects on grapevine (Vitis vinifera L., cv. Corvina) performance and water stress tolerance. Sci. Hortic. 2019, 258, 108784. [Google Scholar] [CrossRef]
- Cholakova-Bimbalova, R.; Petrov, V.; Vassilev, A. Photosynthetic performance of young maize (Zea mays L.) plants exposed to chilling stress can be improved by the application of protein hydrolysates. Acta Agrobot. 2019, 72, 1769. [Google Scholar] [CrossRef]
- Thornton, B.; Robinson, D. Uptake and assimilation of nitrogen from solutions containing multiple N sources. Plant Cell Environ. 2005, 28, 813–821. [Google Scholar] [CrossRef]
- Sonneveld, C.; Voogt, W. (Eds.) Plant Nutrition of Greenhouse Crops; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Dat, J.; Vandenabeele, S.; Vranová, E.; Van Montagu, M.; Inzé, D.; Van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell Mol. Life Sci. 2000, 57, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Chen, C.; Dickman, M.B. Proline suppresses apoptosis in the fungal pathogen Colletotrichum trifolii. Proc. Natl. Acad. Sci. USA 2005, 102, 3459–3464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.-Y.; Yang, J.-Y.; Lu, H.-B.; Wang, S.-S.; Yang, J.; Yang, X.-C.; Chai, M.; Li, L.; Cao, J.-X. Effect of chitosan film incorporated with tea polyphenol on quality and shelf life of pork meat patties. Int. J. Biol. Macromol. 2013, 61, 312–316. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, U.; Bilger, W. Rapid assessment of stress effects on plant leaves by chlorophyll fluorescence measurements. In Plant Response to Stress; NATO ASI Series (Series G: Ecological Sciences); Tenhunen, J.D., Catarino, F.M., Lange, O.L., Oechel, W.C., Eds.; Springer: Berlin/Heidelberg, Germany, 1987; Volume 15, pp. 27–53. [Google Scholar]
- Guo, Y.; Tan, J. Recent advances in the application of chlorophyll a fluorescence from photosystem II. Photochem. Photobiol. 2015, 91, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zushi, K.; Matsuzoe, N. Using of chlorophyll a fluorescence OJIP transients for sensing salt stress in the leaves and fruits of tomato. Sci. Hortic. 2017, 219, 216–221. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Scalbert, A.; Monties, B.; Janin, G. Tannins in wood: Comparison of different estimation methods. J. Agric. Food Chem. 1989, 37, 1324–1329. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Troll, W.; Lindsley, J. A photometric method for the determination of proline. J. Biol. Chem. 1955, 215, 655–660. [Google Scholar] [CrossRef]
- Kato, M.; Shimizu, S. Chlorophyll metabolism in higher plants VI. Involvement of peroxidase in chlorophyll degradation. Plant Cell Physiol. 1985, 26, 1291–1301. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Ding, X.; Jiang, Y.; Zhao, H.; Guo, D.; He, L.; Liu, F.; Zhou, Q.; Nandwani, D.; Hui, D.; Yu, J. Electrical conductivity of nutrient solution influenced photosynthesis, quality, and antioxidant enzyme activity of pakchoi (Brassica campestris L. Ssp. Chinensis) in a hydroponic system. PLoS ONE 2018, 13, e0202090. [Google Scholar] [CrossRef] [PubMed]
- Imas, P.; Bar-Yosef, B.; Kafkafi, U.; Ganmore-Neumann, R. Release of carboxylic anions and protons by tomato roots in response to ammonium nitrate ratio and pH in nutrient solution. Plant Soil 1997, 191, 27–34. [Google Scholar] [CrossRef]
- Albornoz, F.; Lieth, H.J. Over fertilization limits lettuce productivity because of osmotic stress. Chil. J. Agric. Res. 2015, 75, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Vuletić, M.; Šukalović, V.H.-T.; Marković, K.; Maksimović, J.D. Antioxidative system in maize roots as affected by osmotic stress and different nitrogen sources. Biol. Plant 2010, 54, 530–534. [Google Scholar] [CrossRef]
- Kang, W.-H.; Sim, Y.M.; Koo, N.; Nam, J.-Y.; Lee, J.; Kim, N.; Jang, H.; Kim, Y.-M.; Yeom, S.-I. Transcriptome profiling of abiotic responses to heat, cold, salt, and osmotic stress of Capsicum annuum L. Sci. Data 2020, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.-K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Kravić, N.; Marković, K.; Andelković, V.; Šukalović, V.H.-T.; Babić, V.; Vuletić, M. Growth, proline accumulation and peroxidase activity in maize seedlings under osmotic stress. Acta Physiol. Plant. 2013, 35, 233–239. [Google Scholar] [CrossRef]
- Krauss, S.; Schnitzler, W.H.; Grassmann, J.; Woitke, M. The influence of different electrical conductivity values in a simplified recirculating soilless system on inner and outer fruit quality characteristics of tomato. J. Agric. Food Chem. 2006, 54, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Ünlükara, A.; Cemek, B.; Karaman, S.; Erşahin, S. Response of lettuce (Lactuca sativa var. Crispa) to salinity of irrigation water. N. Z. J. Crop Hortic. Sci. 2008, 36, 265–273. [Google Scholar] [CrossRef]
- Taïbi, K.; Taïbi, F.; Ait Abderrahim, L.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Eraslan, F.; Inal, A.; Savasturk, O.; Gunes, A. Changes in antioxidative system and membrane damage of lettuce in response to salinity and boron toxicity. Sci. Hortic. 2007, 114, 5–10. [Google Scholar] [CrossRef]
- Huang, Y.; Bie, Z.; Liu, Z.; Zhen, A.; Wang, W. Protective role of proline against salt stress is partially related to the improvement of water status and peroxidase enzyme activity in cucumber. Soil Sci. Plant Nutr. 2009, 55, 698–704. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, M.; Souri, M.K. Nutrient uptake, proline content and antioxidant enzymes activity of pepper (Capsicum annuum L.) under higher electrical conductivity of nutrient solution created by nitrate or chloride salts of potassium and calcium. Acta Sci. Pol. Hortorum Cultus 2019, 18, 113–122. [Google Scholar] [CrossRef]
- Sánchez, E.; López-Lefebre, L.R.; García, P.C.; Rivero, R.M.; Ruiz, J.M.; Romero, L. Proline metabolism in response to highest nitrogen dosages in green bean plants (Phaseolus vulgaris L. cv. Strike). J. Plant Physiol. 2001, 158, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Appenroth, K.-J.; Stöckel, J.; Srivastava, A.; Strasser, R. Multiple effects of chromate on the photosynthetic apparatus of Spirodela polyrhiza as probed by OJIP chlorophyll a fluorescence measurements. Environ. Pollut. 2001, 115, 49–64. [Google Scholar] [CrossRef]
- Stirbet, A.; Govindjee. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Singh-Tomar, R.; Mathur, S.; Allakhverdiev, S.I.; Jajoo, A. Changes in PS II heterogeneity in response to osmotic and ionic stress in wheat leaves (Triticum aestivum). J. Bioenerg. Biomembr. 2012, 44, 411–419. [Google Scholar] [CrossRef]
- Havaux, M. Rapid photosynthetic adaptation to heat stress triggered in potato leaves by moderately elevated temperatures. Plant Cell Environ. 1993, 16, 461–467. [Google Scholar] [CrossRef]
- Mehta, P.; Jajoo, A.; Mathur, S.; Bharti, S. Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem II in wheat leaves. Plant Physiol. Biochem. 2010, 48, 16–20. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peppermint | Spearmint | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | Plant Weight (g) | Plant Height (cm) | Root Length (cm) | Plant Weight (g) | Plant Height (cm) | Root Length (cm) | ||||||
| Amino16 (%) | *** | * | *** | * | * | *** | ||||||
| 0 | 20.37 y | a z | 34.45 | a | 12.43 | a | 16.73 | a | 37.62 | a | 11.02 | a |
| 0.25 | 18.35 | a | 33.77 | a | 5.45 | b | 17.65 | a | 35.76 | a | 4.47 | b |
| 0.50 | 11.90 | b | 28.79 | b | 0.74 | c | 13.08 | b | 28.24 | b | 0.94 | c |
| NaCl (mM) | ns | ns | * | ns | ns | * | ||||||
| 0 | 20.37 | 34.45 | 12.43 | b | 16.73 | 37.62 | 11.02 | b | ||||
| 10 | 25.67 | 35.97 | 16.58 | a | 18.52 | 39.42 | 13.03 | a | ||||
| 20 | 24.98 | 33.37 | 16.28 | a | 16.19 | 33.47 | 12.22 | ab | ||||
| Peppermint | Spearmint | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Source of Variation | POD (U/100 g FW) | CAT (U/100 g FW) | APX (U/100 g FW) | POD (U/100 g FW) | CAT (U/100 g FW) | APX (U/100 g FW) | ||||
| Amino16 (%) | ns | ns | * | * | ns | * | ||||
| 0 | 106 y | 2.33 | 5.10 | b z | 111 | b | 1.87 | 5.93 | c | |
| 0.25 | 103 | 2.73 | 6.43 | ab | 125 | a | 3.67 | 8.87 | b | |
| 0.50 | 131 | 2.47 | 8.50 | a | 132 | a | 3.53 | 12.50 | a | |
| NaCl (mM) | ns | ns | ns | ns | * | ns | ||||
| 0 | 106 | 2.33 | 5.00 | 111 | 1.87 | b | 5.93 | |||
| 10 | 106 | 2.43 | 5.00 | 112 | 3.30 | a | 7.10 | |||
| 20 | 109 | 2.37 | 6.00 | 123 | 2.33 | b | 7.80 | |||
| Peppermint | Spearmint | ||||||
|---|---|---|---|---|---|---|---|
| Source of Variation | MDA (nmol/g FW) | H2O2 (μmol/g FW) | Proline (mmol/g FW) | MDA (nmol/g FW) | H2O2 (μmol/g FW) | Proline (mmol/g FW) | |
| Amino16 (%) | ns | ns | * | ns | ns | ns | |
| 0 | 0.529 y | 1.108 | 0.198 | b z | 0.442 | 0.399 | 0.197 |
| 0.25 | 0.558 | 1.127 | 0.259 | ab | 0.500 | 0.397 | 0.280 |
| 0.50 | 0.677 | 1.111 | 0.359 | a | 0.511 | 0.406 | 0.210 |
| NaCl (mM) | ns | ns | ns | ns | ns | ns | |
| 0 | 0.529 | 1.108 | 0.200 | 0.442 | 0.399 | 0.197 | |
| 10 | 0.400 | 0.740 | 0.250 | 0.433 | 0.440 | 0.280 | |
| 20 | 0.427 | 0.913 | 0.240 | 0.412 | 0.440 | 0.210 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasampalis, D.S.; Aktsoglou, D.-C.; Tsouvaltzis, P.; Siomos, A.S. Responses of Peppermint and Spearmint Crops to Excessive Biostimulant Application and Increased Salinity in a Closed Soilless Production System. Agronomy 2021, 11, 1168. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061168
Kasampalis DS, Aktsoglou D-C, Tsouvaltzis P, Siomos AS. Responses of Peppermint and Spearmint Crops to Excessive Biostimulant Application and Increased Salinity in a Closed Soilless Production System. Agronomy. 2021; 11(6):1168. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061168
Chicago/Turabian StyleKasampalis, Dimitrios S., Danai-Christina Aktsoglou, Pavlos Tsouvaltzis, and Anastasios S. Siomos. 2021. "Responses of Peppermint and Spearmint Crops to Excessive Biostimulant Application and Increased Salinity in a Closed Soilless Production System" Agronomy 11, no. 6: 1168. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11061168