
Two Distinct Soil Disinfestations Differently Modify the Bacterial Communities in a Tomato Field

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Disinfestations and Sampling
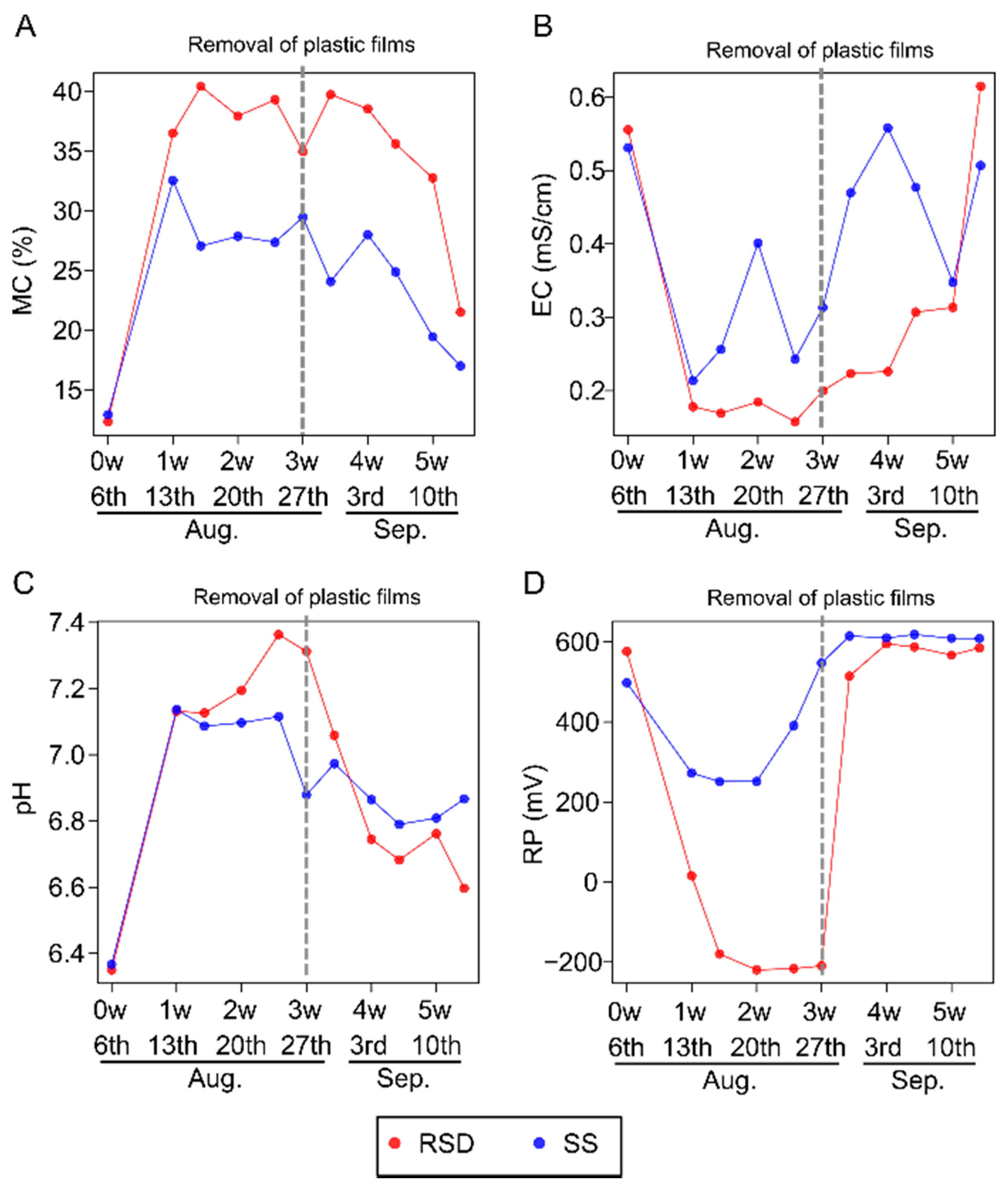
2.2. Environmental Factor Measurement
2.3. DNA Extraction and 16S rRNA Amplicon Sequencing
2.4. Microbiome Data Analysis
2.5. Quantification of F. oxysporum in Soils
2.6. Tomato Cultivation and Evaluation of the Growth and Yield
2.7. Statistical Analysis
3. Results
3.1. Fluctuation of Environmental Factors during Soil Disinfestations
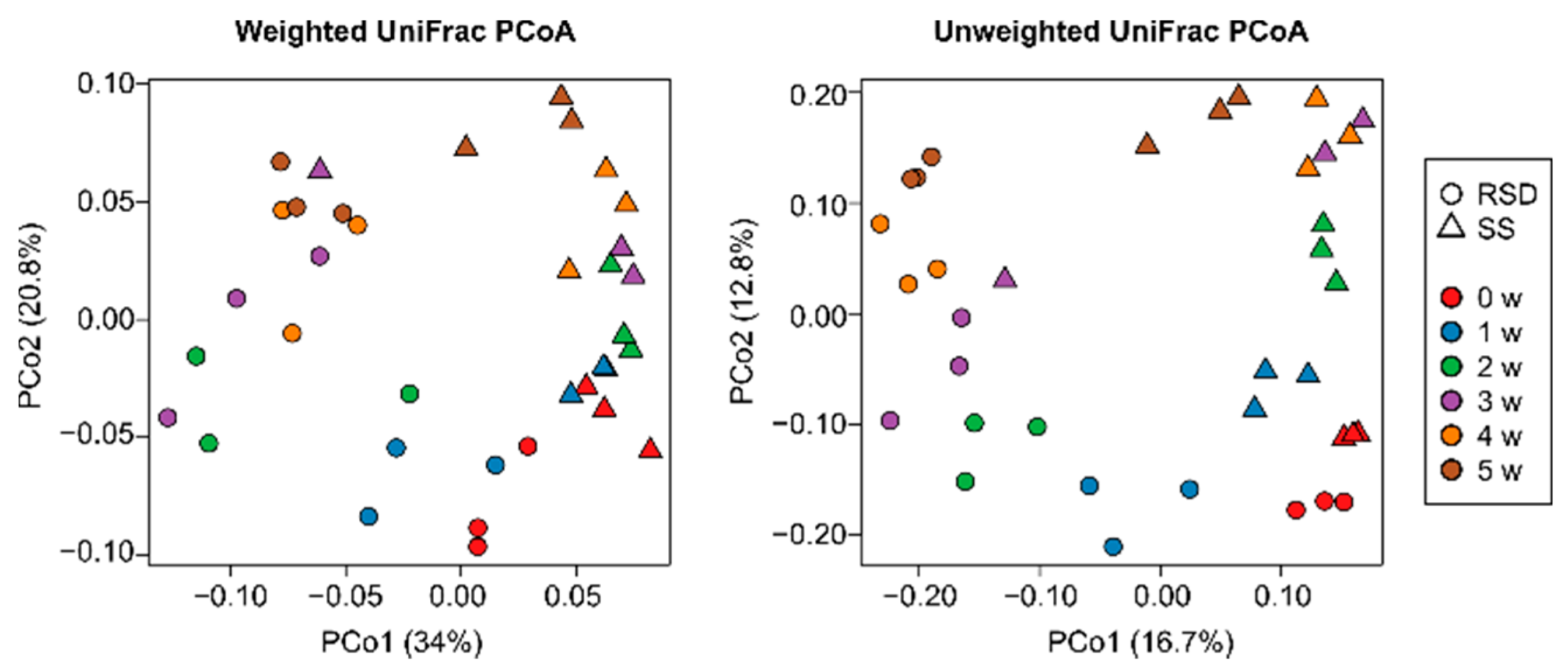
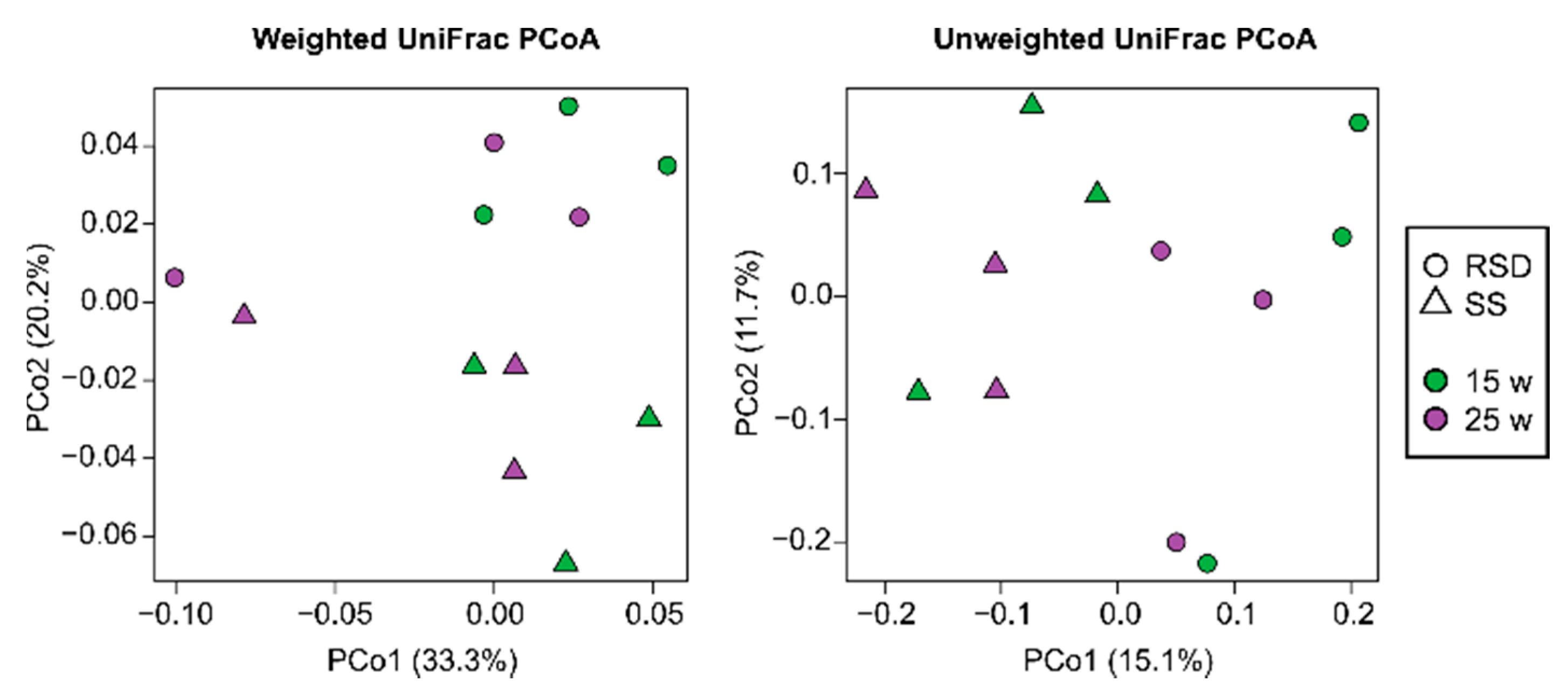
3.2. Changes in Bacterial Communities by Soil Disinfestations
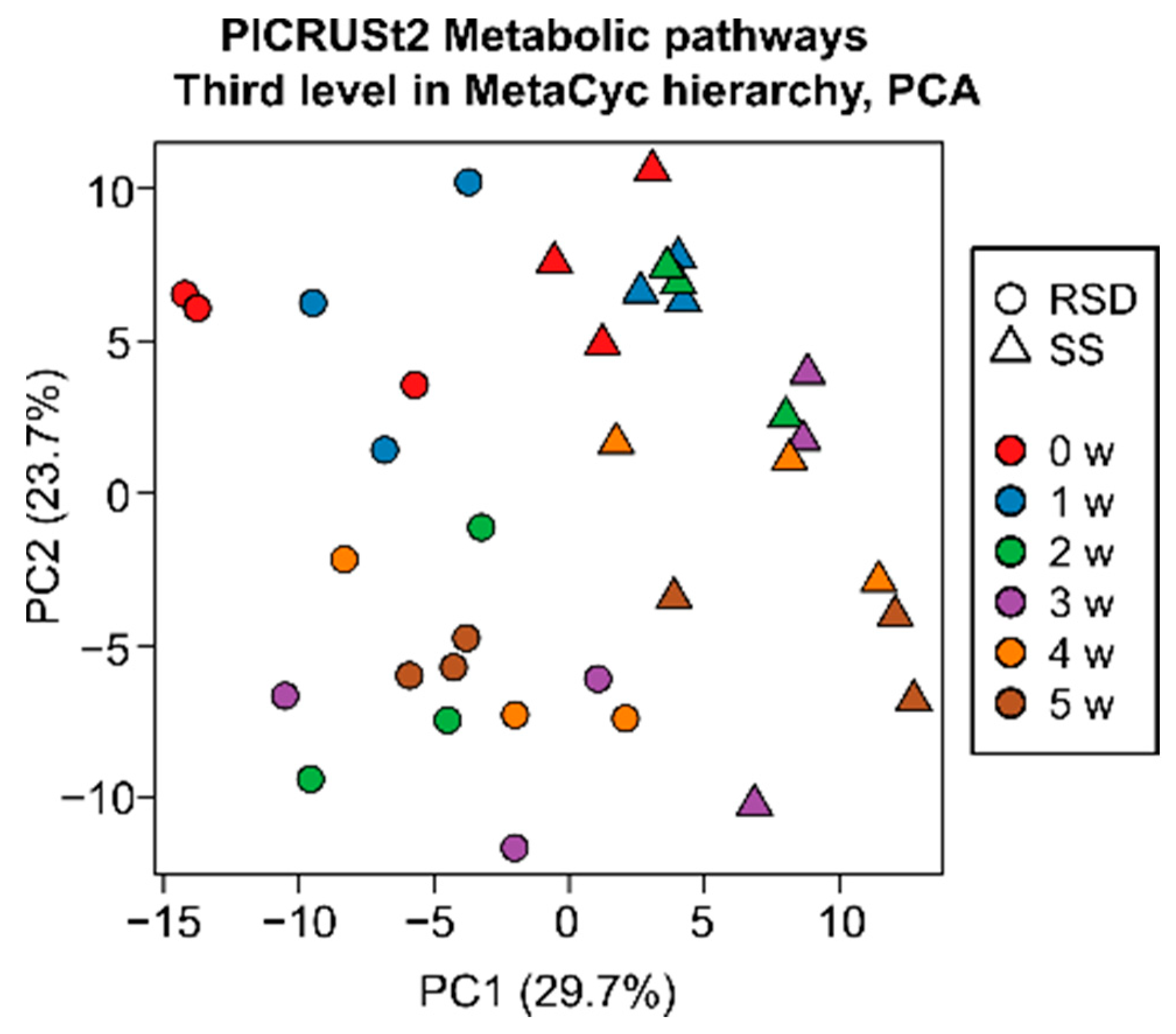
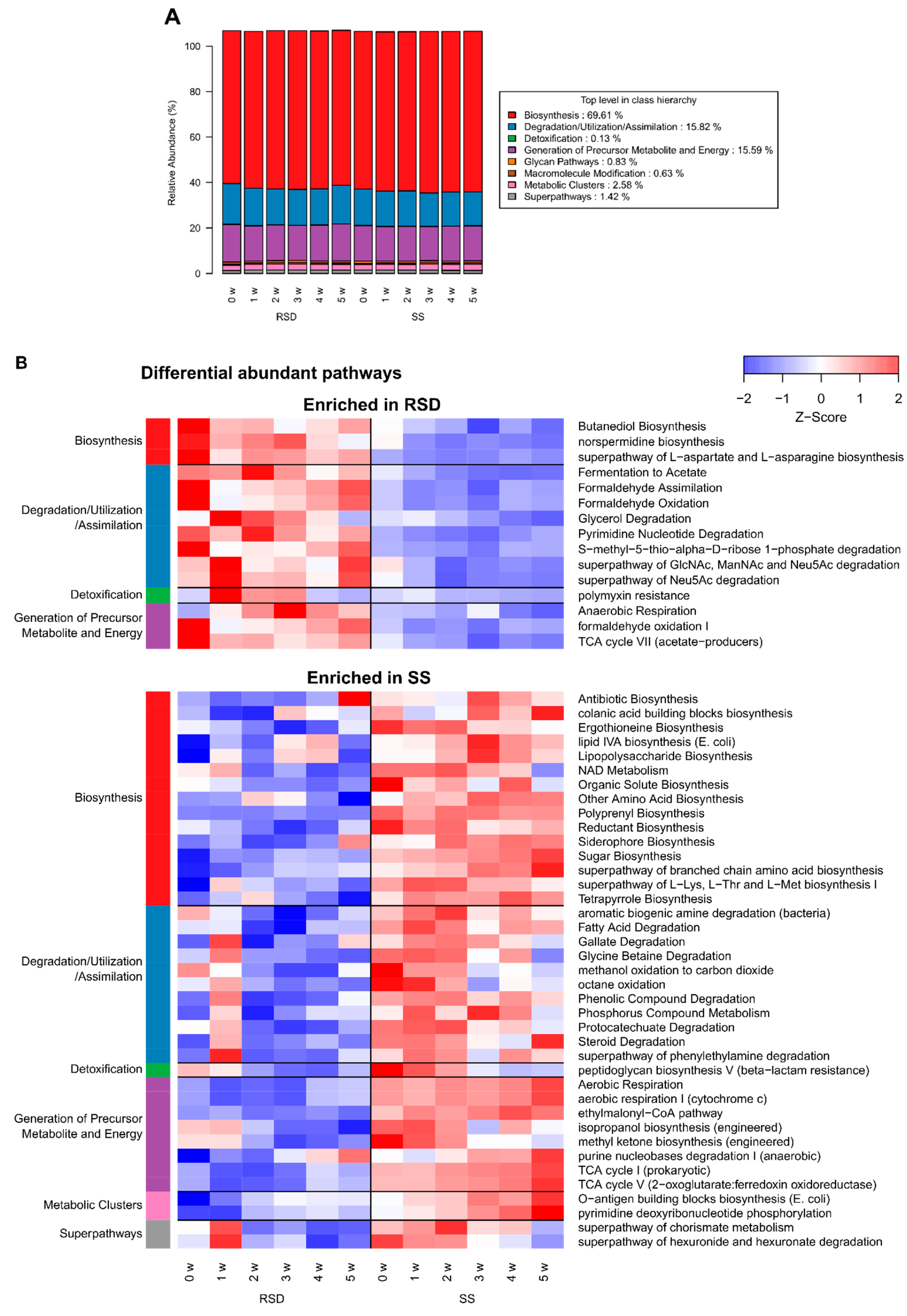
3.3. Changes in Bacterial Pathway Composition by Soil Disinfestations
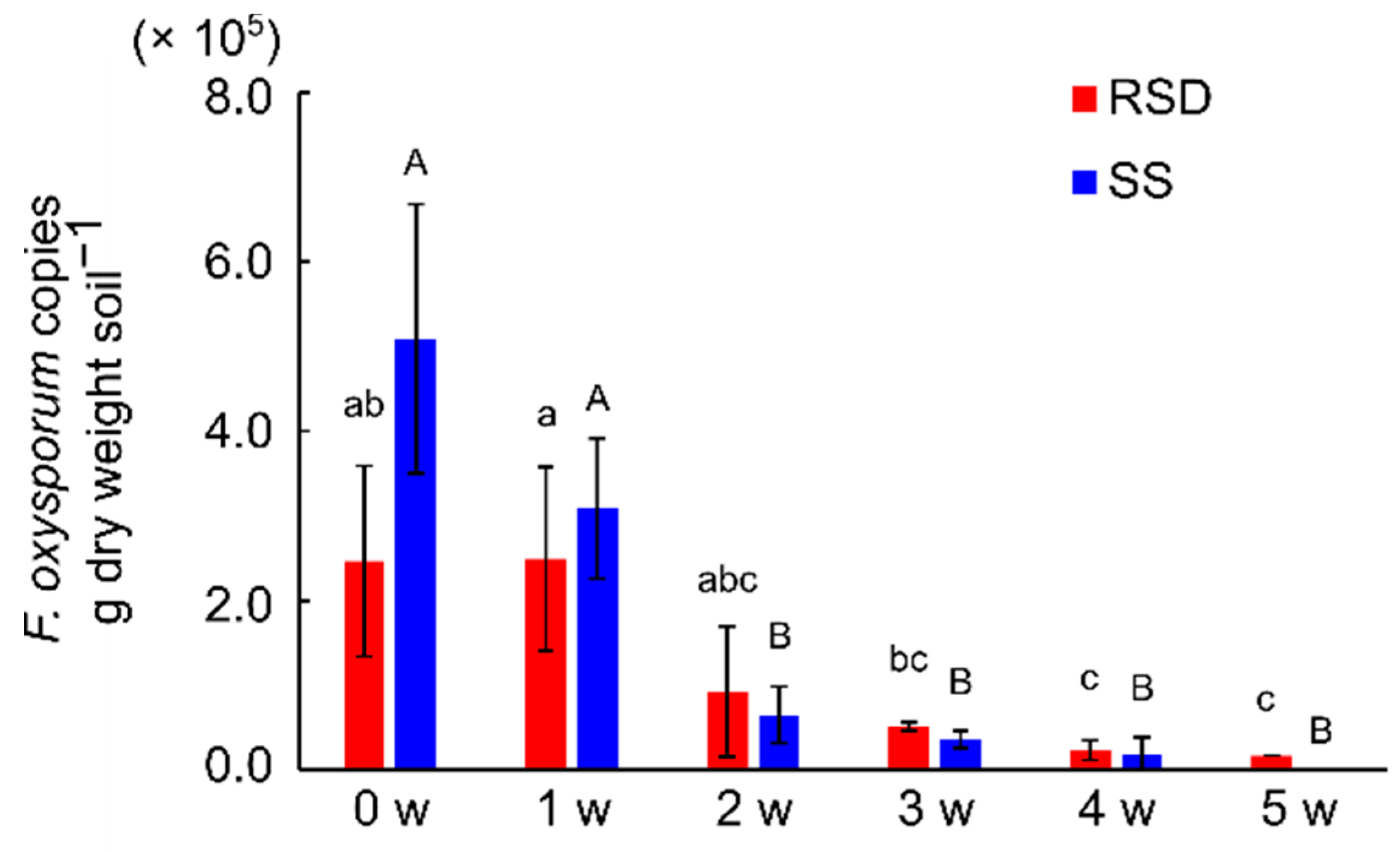
3.4. Difference of F. oxysporum Density between Two Soil Disinfestations
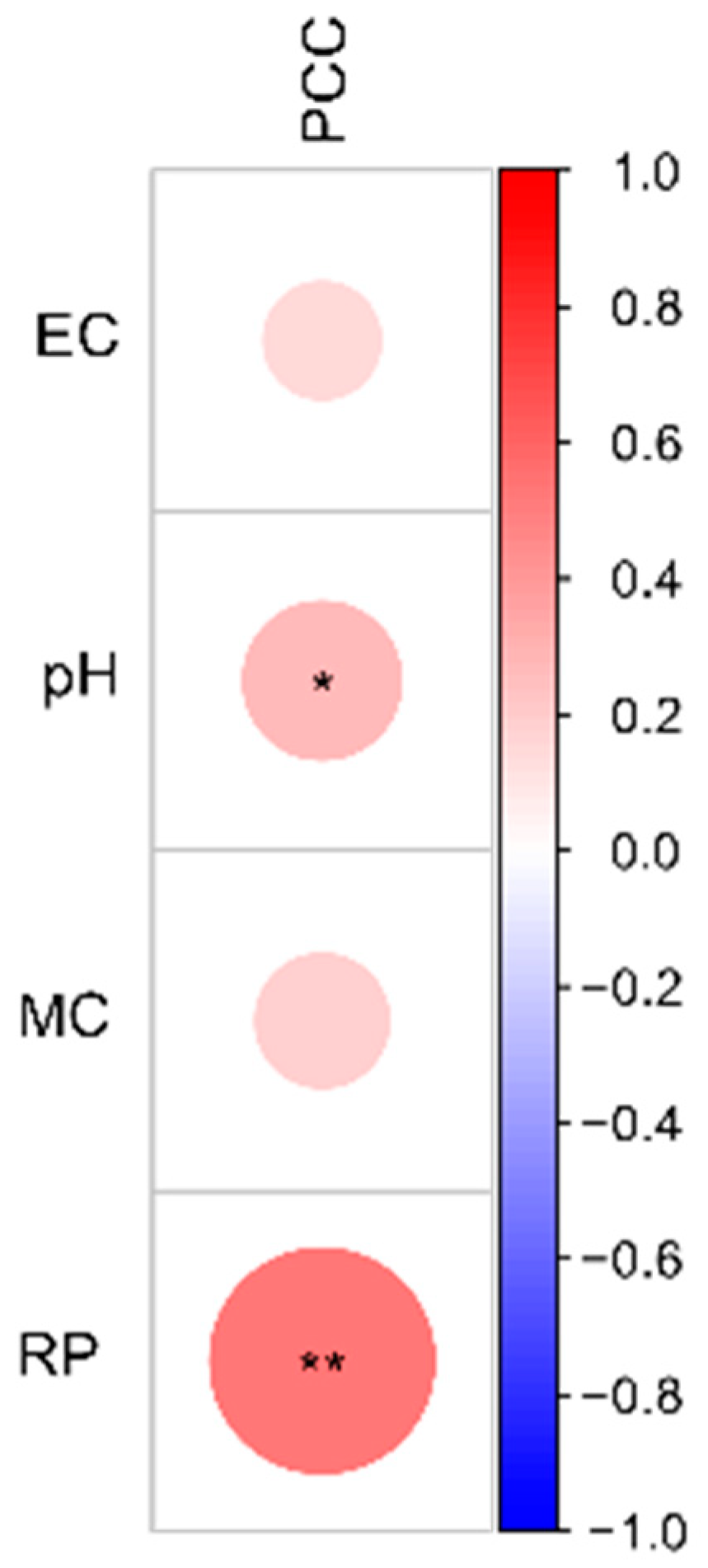
3.5. Effects of Soil Disinfestation on Soil Bacterial Communities and Tomato Growth during Cultivation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Barnard, E.L.; Gilly, S.; Ash, E. An evaluation of dazomet and metam-sodium soil fumigants for control of Macrophomina phaseolina in a Florida forest nursery. Tree Plant. Notes 1994, 45, 91–95. [Google Scholar]
- Carey, W.A. Fumigation with chloropicrin, metham sodium, and EPTC as replacements for methyl bromide in southern pine nurseries. South. J. Appl. For. 2000, 24, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, C.M.; McGiffen, M.E.; Sims, J.J.; Becker, J.O. Fumigant combinations for Cyperus esculentum L control. Pest. Manag. Sci. 2004, 60, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuomisto, H.L.; Hodge, I.D.; Riordan, P.; Macdonald, D.W. Does organic farming reduce environmental impacts?—A meta-analysis of European research. J. Environ. Manag. 2012, 112, 309–320. [Google Scholar] [CrossRef]
- Cavigelli, M.A.; Mirsky, S.B.; Teasdale, J.R.; Spargo, J.T.; Doran, J. Organic grain cropping systems to enhance ecosystem services. Renew. Agric. Food Syst. 2013, 28, 145–159. [Google Scholar] [CrossRef]
- Seufert, V.; Ramankutty, N.; Foley, J.A. Comparing the yields of organic and conventional agriculture. Nature 2012, 485, 229–232. [Google Scholar] [CrossRef]
- Muller, A.; Schader, C.; Scialabba, N.E.-H.; Bruggemann, J.; Isensee, A.; Erb, K.-H.; Smith, P.; Klocke, P.; Leiber, F.; Stolze, M.; et al. Strategies for feeding the world more sustainably with organic agriculture. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Erb, K.-H.; Lauk, C.; Kastner, T.; Mayer, A.; Theurl, M.C.; Haberl, H. Exploring the biophysical option space for feeding the world without deforestation. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, P.; Pellerin, S.; Seufert, V.; Smith, L.; Ramankutty, N.; Nesme, T. Global option space for organic agriculture is delimited by nitrogen availability. Nat. Food 2021, 2, 363–372. [Google Scholar] [CrossRef]
- van Bruggen, A.H.C.; Finckh, M.R. Plant diseases and management approaches in organic farming systems. Annu. Rev. Phytopathol. 2016, 54, 25–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willer, H.; Lernoud, J. The World of Organic Agriculture. Statistics and Emerging Trends 2019; Research Institute of Organic Agriculture (FiBL): Frick, Switzerland; IFOAM—Organics International: Bonn, Germany, 2019. [Google Scholar]
- Katan, J. Diseases caused by soilborne pathogens: Biology, management and challenges. J. Plant. Pathol. 2017, 99, 305–315. [Google Scholar]
- Blok, W.J.; Lamers, J.G.; Termorshuizen, A.J.; Bollen, G.J. Control of soilborne plant pathogens by incorporating fresh organic amendments followed by tarping. Phytopathology 2000, 90, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Shinmura, A. Causal agent and control of root rot of welsh onion. In Proceedings of the PSJ Soil-Borne Disease Workshop Report; 2000; pp. 133–143. Available online: https://ci.nii.ac.jp/naid/10029720077/ (accessed on 5 July 2021).
- Shrestha, U.; Auge, R.M.; Butler, D.M. A meta-analysis of the impact of anaerobic soil disinfestation on pest suppression and yield of horticultural crops. Front. Plant. Sci. 2016, 7, 1254. [Google Scholar] [CrossRef]
- Momma, N.; Kobara, Y.; Uematsu, S.; Kita, N.; Shinmura, A. Development of biological soil disinfestations in Japan. Appl. Microbiol. Biotechnol. 2013, 97, 3801–3809. [Google Scholar] [CrossRef]
- Ueki, A.; Kaku, N.; Ueki, K. Role of anaerobic bacteria in biological soil disinfestation for elimination of soil-borne plant pathogens in agriculture. Appl. Microbiol. Biotechnol. 2018, 102, 6309–6318. [Google Scholar] [CrossRef] [PubMed]
- Momma, N.; Momma, M.; Kobara, Y. Biological soil disinfestation using ethanol: Effect on Fusarium oxysporum f. sp lycopersici and soil microorganisms. J. Gen. Plant. Pathol. 2010, 76, 336–344. [Google Scholar] [CrossRef]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. organic amendments: Microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Chen, L.; Zhang, J.B.; Yin, J.; Huang, S.M. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lori, M.; Symnaczik, S.; Mader, P.; De Deyn, G.; Gattinger, A. Organic farming enhances soil microbial abundance and activity-A meta-analysis and meta-regression. PLoS ONE 2017, 12, e0180442. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P. The rhizosphere microbiome and plant health. Trends Plant. Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Mowlick, S.; Takehara, T.; Kaku, N.; Ueki, K.; Ueki, A. Proliferation of diversified clostridial species during biological soil disinfestation incorporated with plant biomass under various conditions. Appl. Microbiol. Biotechnol. 2013, 97, 8365–8379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowlick, S.; Yasukawa, H.; Inoue, T.; Takehara, T.; Kaku, N.; Ueki, K.; Ueki, A. Suppression of spinach wilt disease by biological soil disinfestation incorporated with Brassica juncea plants in association with changes in soil bacterial communities. Crop. Prot. 2013, 54, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Mowlick, S.; Inoue, T.; Takehara, T.; Tonouchi, A.; Kaku, N.; Ueki, K.; Ueki, A. Usefulness of Japanese-radish residue in biological soil disinfestation to suppress spinach wilt disease accompanying with proliferation of soil bacteria in the Firmicutes. Crop. Prot. 2014, 61, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Rosskopf, E.N.; Burelle, N.; Hong, J.; Butler, D.M.; Noling, J.W.; He, Z.; Booker, B.; Sances, F. Comparison of anaerobic soil disinfestation and drip-applied organic acids for raised-bed specialty crop production in Florida. In Proceedings of the VIII International Symposium on Chemical and Non-Chemical Soil and Substrate Disinfestation, Torino, Italy, 13–17 July 2014; pp. 221–228. [Google Scholar]
- Strauss, S.L.; Kluepfel, D.A. Anaerobic soil disinfestation: A chemical-independent approach to pre-plant control of plant pathogens. J. Integr. Agric. 2015, 14, 2309–2318. [Google Scholar] [CrossRef]
- Ueki, A.; Takehara, T.; Ishioka, G.; Kaku, N.; Ueki, K. Degradation of the fungal cell wall by clostridial strains isolated from soil subjected to biological soil disinfestation and biocontrol of Fusarium wilt disease of spinach. Appl. Microbiol. Biotechnol. 2017, 101, 8267–8277. [Google Scholar] [CrossRef]
- Ueki, A.; Takehara, T.; Ishioka, G.; Kaku, N.; Ueki, K. Production of beta-1,3-glucanase and chitosanase from clostridial strains isolated from the soil subjected to biological disinfestation. AMB Express 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.G.; Kunitomo, E.; Iida, T.; Nakaho, K.; Ohkuma, M. Soil prokaryotes are associated with decreasing Fusarium oxysporum density during anaerobic soil disinfestation in the tomato field. Appl. Soil Ecol. 2020, 155, 103632. [Google Scholar] [CrossRef]
- Katan, J. Solar Heating (Solarization) of Soil for Control of Soilborne Pests. Annu. Rev. Phytopathol. 1981, 19, 211–236. [Google Scholar] [CrossRef]
- Ichihashi, Y.; Date, Y.; Shino, A.; Shimizu, T.; Shibata, A.; Kumaishi, K.; Funahashi, F.; Wakayama, K.; Yamazaki, K.; Umezawa, A.; et al. Multi-omics analysis on an agroecosystem reveals the significant role of organic nitrogen to increase agricultural crop yield. Proc. Natl. Acad. Sci. USA 2020, 117, 14552–14560. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.; Defago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Hoitink, H.A.J.; Boehm, M.J. Biocontrol within the context of soil microbial communities: A substrate-dependent phenomenon. Annu. Rev. Phytopathol. 1999, 37, 427–446. [Google Scholar] [CrossRef] [PubMed]
- Weller, D.M.; Raaijmakers, J.M.; Gardener, B.B.M.; Thomashow, L.S. Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu. Rev. Phytopathol. 2002, 40, 309–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 17. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2-approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, 10. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc database of metabolic pathways and enzymes—A 2019 update. Nucleic Acids Res. 2020, 48, D445–D453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mule, G.; Susca, A.; Stea, G.; Moretti, A. Specific detection of the toxigenic species Fusarium proliferatum and F. oxysporum from asparagus plants using primers based on calmodulin gene sequences. Fems Microbiol. Lett. 2004, 230, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. vegan: Community Ecology Package. Available online: https://CRAN.R-project.org/package=vegan (accessed on 5 July 2021).
- Fernandes, A.; Macklaim, J.; Linn, T.; Reid, G.; Gloor, G. ANOVA-like differential gene expression analysis of single-organism and meta-RNA-seq. PLoS ONE 2013, 8, e67019. [Google Scholar]
- Fernandes, A.D.; Reid, J.N.; Macklaim, J.M.; McMurrough, T.A.; Edgell, D.R.; Gloor, G.B. Unifying the analysis of high-throughput sequencing datasets: Characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome 2014, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.R.; DeLaune, R.D. Biogeochemistry of Wetlands: Science and Applications; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Larkin, R.P.; Fravel, D.R. Efficacy of various fungal and bacterial biocontrol organisms for control of Fusarium wilt of tomato. Plant. Dis. 1998, 82, 1022–1028. [Google Scholar] [CrossRef] [Green Version]
- Browne, G.; Ott, N.; Poret-Peterson, A.; Gouran, H.; Lampinen, B. Efficacy of anaerobic soil disinfestation for control of prunus replant disease. Plant. Dis. 2018, 102, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Hewavitharana, S.S.; Ruddell, D.; Mazzola, M. Carbon source-dependent antifungal and nematicidal volatiles derived during anaerobic soil disinfestation. Eur. J. Plant. Pathol. 2014, 140, 39–52. [Google Scholar] [CrossRef]
- Messiha, N.A.S.; van Diepeningen, A.D.; Wenneker, M.; van Beuningen, A.R.; Janse, J.D.; Coenen, T.G.C.; Termorshuizen, A.J.; van Bruggen, A.H.C.; Blok, W.J. Biological Soil Disinfestation (BSD), a new control method for potato brown rot, caused by Ralstonia solanacearum race 3 biovar 2. Eur. J. Plant. Pathol. 2007, 117, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Momma, N.; Yamamoto, K.; Simandi, P.; Shishido, M. Role of organic acids in the mechanisms of biological soil disinfestation (BSD). J. Gen. Plant. Pathol. 2006, 72, 247–252. [Google Scholar] [CrossRef]
- Shennan, C.; Muramoto, J.; Koike, S.; Baird, G.; Fennimore, S.; Samtani, J.; Bolda, M.; Dara, S.; Daugovish, O.; Lazarovits, G.; et al. Anaerobic soil disinfestation is an alternative to soil fumigation for control of some soilborne pathogens in strawberry production. Plant. Pathol. 2018, 67, 51–66. [Google Scholar] [CrossRef]
- Strauss, S.L.; Greenhut, R.F.; McClean, A.E.; Kluepfel, D.A. Effect of anaerobic soil disinfestation on the bacterial community and key soilborne phytopathogenic agents under walnut tree-crop nursery conditions. Plant. Soil 2017, 415, 493–506. [Google Scholar] [CrossRef]
- Huang, X.Q.; Liu, L.L.; Wen, T.; Zhu, R.; Zhang, J.B.; Cai, Z.C. Illumina MiSeq investigations on the changes of microbial community in the Fusarium oxysporum f.sp cubense infected soil during and after reductive soil disinfestation. Microbiol. Res. 2015, 181, 33–42. [Google Scholar] [CrossRef]
- Liu, L.L.; Huang, X.Q.; Zhao, J.; Zhang, J.B.; Cai, Z.C. Characterizing the key agents in a disease-suppressed soil managed by reductive soil disinfestation. Appl. Environ. Microbiol. 2019, 85, e02992-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, T.Z.; Ren, G.D.; Wang, G.F.; Ma, Y. Impacts on soil microbial characteristics and their restorability with different soil disinfestation approaches in intensively cropped greenhouse soils. Appl. Microbiol. Biotechnol. 2019, 103, 6369–6383. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.Y.; Liao, H.K.; Shu, L.Z.; Yao, H.Y. Effect of different substrates on soil microbial community structure and the mechanisms of reductive soil disinfestation. Front. Microbiol. 2019, 10, 2851. [Google Scholar] [CrossRef]
- Testen, A.L.; Miller, S.A. Anaerobic Soil Disinfestation to Manage Soilborne Diseases in Muck Soil Vegetable Production Systems. Plant. Disease 2019, 103, 1757–1762. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhou, X.; Jiang, A.Q.; Fan, J.Z.; Lan, T.; Zhang, J.B.; Cai, Z.C. Distinct impacts of reductive soil disinfestation and chemical soil disinfestation on soil fungal communities and memberships. Appl. Microbiol. Biotechnol. 2018, 102, 7623–7634. [Google Scholar] [CrossRef]
- Janvier, C.; Villeneuve, F.; Alabouvette, C.; Edel-Hermann, V.; Mateille, T.; Steinberg, C. Soil health through soil disease suppression: Which strategy from descriptors to indicators? Soil Biol. Biochem. 2007, 39, 1–23. [Google Scholar] [CrossRef]
- Zhou, X.; Li, C.Y.; Liu, L.L.; Zhao, J.; Zhang, J.B.; Cai, Z.C.; Huang, X.Q. Control of Fusarium wilt of lisianthus by reassembling the microbial community in infested soil through reductive soil disinfestation. Microbiol. Res. 2019, 220, 1–11. [Google Scholar] [CrossRef]
- Mowlick, S.; Hirota, K.; Takehara, T.; Kaku, N.; Ueki, K.; Ueki, A. Development of anaerobic bacterial community consisted of diverse clostridial species during biological soil disinfestation amended with plant biomass. Soil Sci. Plant. Nutr. 2012, 58, 273–287. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Q.; Liu, L.L.; Wen, T.; Zhang, J.B.; Wang, F.H.; Cai, Z.C. Changes in the soil microbial community after reductive soil disinfestation and cucumber seedling cultivation. Appl. Microbiol. Biotechnol. 2016, 100, 5581–5593. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Q.; Liu, L.L.; Zhao, J.; Zhang, J.B.; Cai, Z.C. The families Ruminococcaceae, Lachnospiraceae, and Clostridiaceae are the dominant bacterial groups during reductive soil disinfestation with incorporated plant residues. Appl. Soil Ecol. 2019, 135, 65–72. [Google Scholar] [CrossRef]
- Huang, X.Q.; Zhao, J.; Zhou, X.; Han, Y.S.; Zhang, J.B.; Cai, Z.C. How green alternatives to chemical pesticides are environmentally friendly and more efficient. Eur. J. Soil Sci. 2019, 70, 518–529. [Google Scholar] [CrossRef]
- Hewavitharana, S.S.; Klarer, E.; Reed, A.J.; Leisso, R.; Poirier, B.; Honaas, L.; Rudell, D.R.; Mazzola, M. Temporal dynamics of the soil metabolome and microbiome during simulated anaerobic soil disinfestation. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Poret-Peterson, A.T.; Sayed, N.; Glyzewski, N.; Forbes, H.; Gonzalez-Orta, E.T.; Kluepfel, D.A. Temporal responses of microbial communities to anaerobic soil disinfestation. Microb. Ecol. 2020, 80, 191–201. [Google Scholar] [CrossRef] [Green Version]
- van Agtmaal, M.; van Os, G.J.; Hol, W.H.G.; Hundscheid, M.P.J.; Runia, W.T.; Hordijk, C.A.; de Boer, W. Legacy effects of anaerobic soil disinfestation on soil bacterial community composition and production of pathogen-suppressing volatiles. Front. Microbiol. 2015, 6, 701. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Stem (cm) | Internode (cm) | Leaf Area (cm2) | Perimeter of Stem (cm) | ||
|---|---|---|---|---|---|
| October 16 | RSD | 94.4 ± 7.42 | 24.7 ± 1.89 | 1.41×103 ± 12.7 | 3.3 ± 0.053 |
| SS | 95.4 ± 4.21 | 24.1 ± 1.10 | 1.45×103 ± 229 | 3.4 ± 0.081 | |
| October 30 | RSD | 151 ± 7.18 * | 26.7 ± 0.700 * | 1.90×103 ± 41.8 | 2.9 ± 0.11 |
| SS | 139 ± 9.64 | 22.7 ± 1.94 | 1.79×103 ± 313 | 3.3 ± 0.22 * | |
| November 13 | RSD | 188 ± 7.51 | 18.8 ± 2.79 | 1.99×103 ± 89.2 | 3.8 ± 0.061 |
| SS | 178 ± 14.7 | 19.5 ± 3.31 | 2.04×103 ± 431 | 4.0 ± 0.42 | |
| November 27 | RSD | 226 ± 2.60 * | 18.0 ± 1.37 | 1.63×103 ± 56.0 | 3.1 ± 0.14 |
| SS | 209 ± 20.8 | 15.6 ± 0.133 | 1.80×103 ± 497 | 3.2 ± 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakayasu, M.; Ikeda, K.; Yamazaki, S.; Aoki, Y.; Yazaki, K.; Washida, H.; Sugiyama, A. Two Distinct Soil Disinfestations Differently Modify the Bacterial Communities in a Tomato Field. Agronomy 2021, 11, 1375. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071375
Nakayasu M, Ikeda K, Yamazaki S, Aoki Y, Yazaki K, Washida H, Sugiyama A. Two Distinct Soil Disinfestations Differently Modify the Bacterial Communities in a Tomato Field. Agronomy. 2021; 11(7):1375. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071375
Chicago/Turabian StyleNakayasu, Masaru, Kyoko Ikeda, Shinichi Yamazaki, Yuichi Aoki, Kazufumi Yazaki, Haruhiko Washida, and Akifumi Sugiyama. 2021. "Two Distinct Soil Disinfestations Differently Modify the Bacterial Communities in a Tomato Field" Agronomy 11, no. 7: 1375. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11071375