
Combined Influence of Grafting and Type of Protected Environment Structure on Agronomic and Physiological Traits of Single- and Cluster-Fruit-Bearing Cucumber Hybrids

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Plant Growth, Fruit Yield and Quality Parameters
2.3. Physio-Biochemcial Parameters
2.4. Leaf Mineral Analyses
2.5. Statistical Analyses
3. Results
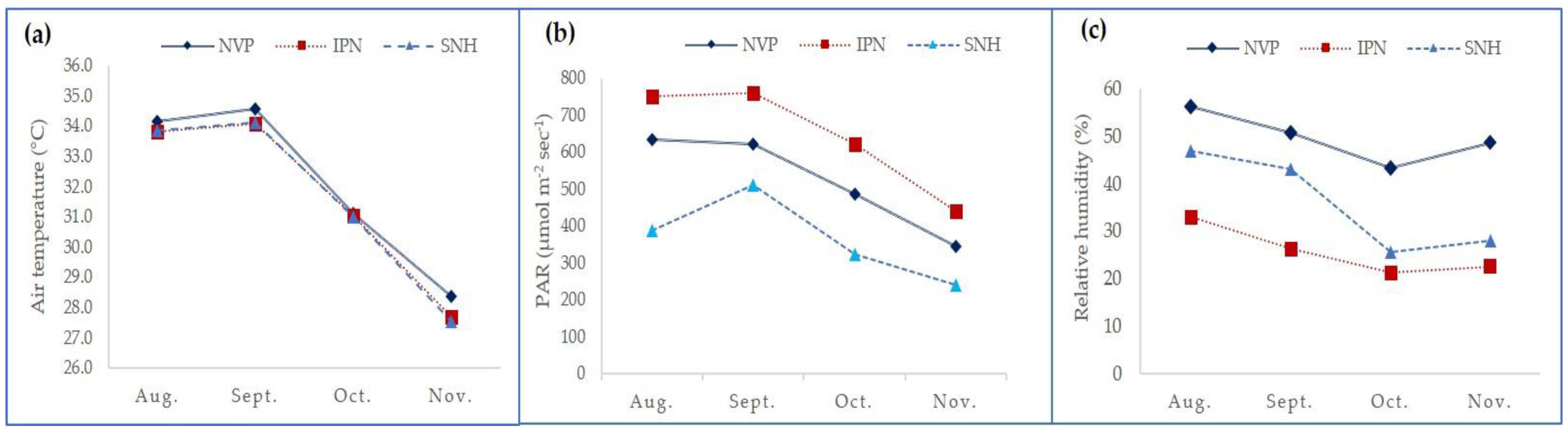
3.1. Microenvironment
3.2. Growth Parameters
3.3. Fruit Yield Parameters and Water Productivity
3.4. Fruit Quality
3.5. Physio-Biochemical Parameters
3.6. Mineral Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aznar-Sánchez, J.A.; Velasco-Muñoz, J.F.; López-Felices, B.; Román-Sánchez, I.M. An analysis of global research trends on greenhouse technology: Towards a sustainable agriculture. Int. J. Environ. Res. Public Health 2020, 17, 664. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Khapte, P.; Saxena, A.; Singh, A.; Parwar, N. Intergeneric grafting for enhanced growth, yield and nutrient acquisition in greenhouse cucumber during winter. J. Environ. Biol. 2019, 40, 295–301. [Google Scholar] [CrossRef]
- Ali, S.A. Modeling of solar radiation available at different orientations of greenhouses. Misr J. Agric. Eng. 2012, 29, 1181–1196. [Google Scholar] [CrossRef]
- Kumar, P.; Khapte, P.S.; Saxena, A.; Kumar, P. Evaluation of gynoecious cucumber (Cucumis sativus) hybrids for early-summer greenhouse production in western Indian arid plains. Indian J. Agric. Sci. 2019, 89, 545–550. [Google Scholar]
- Katsoulas, N.; Kittas, C. Impact of greenhouse microclimate on plant growth and development with special reference to the Solanaceae. Eur. J. Plant Sci. Biotechnol. 2008, 2, 31–44. [Google Scholar]
- Alsadon, A.; Al-Helal, I.; Ibrahim, A.; Abdel-Ghany, A.; Al-Zaharani, S.; Ashour, T. The effects of plastic greenhouse covering on cucumber (Cucumis sativus L.) growth. Ecol. Eng. 2016, 87, 305–312. [Google Scholar] [CrossRef]
- Li, T.; Yang, Q. Advantages of diffuse light for horticultural production and perspectives for further research. Front. Plant Sci. 2015, 6, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, F.A.; Medany, M.A.; Abd El-Moniem, E.M.; Abdallah, M.M.F. Influence of green-house cover on potential evapo-transpiration and cucumber water requirements. Ann. Agric. Sci. 2011, 56, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Baille, A.; Kittas, C.; Katsoulas, N. Influence of whitening on greenhouse microclimate and crop energy partitioning. Agric. For. Meteorol. 2001, 107, 293–306. [Google Scholar] [CrossRef]
- Cemek, B.; Demir, Y.; Uzun, S. Effects of Greenhouse Covers on Growth and Yield of Aubergine. Eur. J. Hortic. Sci. 2005, 70, 16–22. [Google Scholar]
- Taha, N.; Abdalla, N.; Bayoumi, Y.; El-Ramady, H. Management of greenhouse cucumber production under arid environments: A Review. Environ. Biodivers. Soil Secur. 2020, 4, 123–136. [Google Scholar] [CrossRef]
- Colla, G.; Kumar, P.; Cardarelli, M.; Rouphael, Y. Grafting an effective tool for abiotic stress alleviation in vegetables. In Horticulture for Food and Environment Security; Chadha, K.L., Singh, A.K., Singh, S.K., Dhillon, W.S., Eds.; Westville Publishing House: New Delhi, India, 2012; pp. 15–28. [Google Scholar]
- Colla, G.; Rouphael, Y.; Jawad, R.; Kumar, P.; Rea, E.; Cardarelli, M. The effectiveness of grafting to improve NaCl and CaCl2 tolerance in cucumber. Sci. Hortic. 2013, 164, 380–391. [Google Scholar] [CrossRef]
- Guan, W.; Haseman, D.; Nowaskie, D. Rootstock Evaluation for Grafted Cucumbers Grown in High Tunnels: Yield and Plant Growth. HortScience 2020, 55, 914–919. [Google Scholar] [CrossRef]
- Li, Y.; Tian, X.; Wei, M.; Shi, Q.; Yang, F.; Wang, X. Mechanisms of tolerance differences in cucumber seedlings grafted on rootstocks with different tolerance to low temperature and weak light stresses. Turk. J. Bot. 2015, 39, 606–614. [Google Scholar] [CrossRef]
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Demirtas, C.; Ayas, S. Deficit irrigation effects on pepper (Capsicum annuum L. Demre) yield in unheated greenhouse condition. J. Food Agric. Environ. 2009, 7, 989–993. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. polyphenoloxidase in beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1995. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Printice-Hall of India Pvt. Ltd.: New Delhi, India, 1973; p. 498. [Google Scholar]
- Chapman, D.H.; Pratt, P.F. Methods of Analysis for Soils, Plants and Water; University of California: Berkeley, CA, USA, 1961. [Google Scholar]
- Derderian, M.D. Determination of Calcium and Magnesium in Plant Material with EDTA. Anal. Chem. 1961, 33, 1796–1798. [Google Scholar] [CrossRef]
- Palaskar, M.S.; Babrekar, P.G.; Ghosh, A.B. A rapid analytical technique to estimate sulphur in soil and plant extracts. J. Indian Soc. Soil Sci. 1981, 29, 249–256. [Google Scholar]
- Hemming, S.; Dueck, T.; Janse, J.; Van Noort, F. The effect of diffuse light on crops. Acta Hortic. 2008, 801, 1293–1300. [Google Scholar] [CrossRef] [Green Version]
- Sudhakar, K.; Srivastava, T.; Satpathy, G.; Premalatha, M. Modelling and estimation of photosynthetically active incident radiation based on global irradiance in Indian latitudes. Int. J. Energy Environ. Eng. 2013, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, A.; Hu, Y.; Tanny, J.; Asante, E.A. Effects of shading and insect-proof screens on crop microclimate and pro-duction: A review of recent advances. Sci. Hortic. 2018, 241, 241–251. [Google Scholar] [CrossRef]
- Okeyo, D.O.; Fry, J.D.; Bremer, D.J.; Chandra, A.; Genovesi, A.D.; Engelke, M.C. Stolon growth and tillering of experimental zoysia grasses in shade. HortScience 2011, 46, 1418–1422. [Google Scholar] [CrossRef] [Green Version]
- Van Holsteijn, G.P.A. Small differences in temperature, great differences in yield. Groenten Fruit 1987, 43, 45–48. [Google Scholar]
- Savvas, D.; Ntatsi, G.; Barouchas, P. Impact of grafting and rootstock genotype on cation uptake by cucumber (Cucumis sativus L.) exposed to Cd or Ni stress. Sci. Hortic. 2013, 149, 86–96. [Google Scholar] [CrossRef]
- Omar, G.F.; Elhamahmy, M.A.M. Effect of rootstocks on vegetative growth, yield and fruit quality of cucumber. Hortsci. J. Suez Canal Univ. 2019, 8, 1–10. [Google Scholar]
- Kumar, P.; Rouphael, Y.; Cardarelli, M.; Colla, G. Vegetable Grafting as a Tool to Improve Drought Resistance and Water Use Efficiency. Front. Plant Sci. 2017, 8, 1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryder, P.; McKeown, P.C.; Fort, A.; Spillane, C. Epigenetics and heterosis in crop plants. In Epigenetics in Plants of Agronomic Importance: Fundamentals and Applications: Transcriptional Regulation and Chromatin Remodelling in Plants; Alvarez-Venegas, R., de la Pena, C., Casas-Mollano, J.A., Eds.; Springer: New York, NY, USA, 2014; pp. 13–31. [Google Scholar]
- Khah, E.M.; Katsoulas, N.; Tchamitchian, M.; Kittas, C. Effect of grafting on eggplant leaf gas exchanges under Mediterranean greenhouse conditions. Intern. J. Plant Prod. 2011, 5, 121–134. [Google Scholar]
- Uysal, N.; Tüzel, Y.; Oztekin, G.; Tuzel, I. Effects of different rootstocks on greenhouse cucumber production. Acta Hortic. 2012, 927, 281–289. [Google Scholar] [CrossRef]
- Davis, A.R.; Perkins-Veazie, P.; Hassell, R.; Levi, A.; King, S.R.; Zhang, X. Grafting Effects on Vegetable Quality. HortScience 2008, 43, 1670–1672. [Google Scholar] [CrossRef] [Green Version]
- Pathare, P.; Opara, U.L.; Al-Said, F.A.-J. Colour Measurement and Analysis in Fresh and Processed Foods: A Review. Food Bioprocess Technol. 2012, 6, 36–60. [Google Scholar] [CrossRef]
- Renna, M.; D’Imperio, M.; Gonnella, M.; Parente, A.; Santamaria, P.; Serio, F. Barattiere: An Italian local variety of Cucumis melo L. with quality traits between melon and cucumber. Plants 2020, 9, 578. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.S.; Tiwari, K.N.; Jha, M.K. Design and technology for greenhouse cooling in tropical and sub-tropical regions: A review. Energy Build. 2009, 41, 1269–1275. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Rea, E.; Colla, G. Improving melon and cucumber photosynthetic activity, mineral composition, and growth performance under salinity stress by grafting onto Cucurbita hybrid rootstocks. Photosynthetica 2012, 50, 180–188. [Google Scholar] [CrossRef]
- Liu, S.; Li, H.; Lv, X.; Ahammed, G.J.; Xia, X.; Zhou, J.; Shi, K.; Asami, T.; Yu, J.; Zhou, Y. Grafting cucumber onto luffa improves drought tolerance by increasing ABA biosynthesis and sensitivity. Sci. Rep. 2016, 6, 20212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savvas, D.; Ntatsi, G.; Rodopoulou, M.; Goumenaki, F. Nutrient uptake concentrations in a cucumber crop grown in a closed hydroponic system under mediterranean climatic conditions as influenced by irrigation schedule. Acta Hortic. 2014, 1034, 545–552. [Google Scholar] [CrossRef]
- Castilla, N.; Montero, J. Environmental control and crop production in mediterranean greenhouses. Acta Hortic. 2008, 797, 25–36. [Google Scholar] [CrossRef]
- Yan, Q.; Duan, Z.; Mao, J.; Li, X.; Dong, F. Effects of root-zone temperature and N, P, and K supplies on nutrient uptake of cucumber (Cucumis sativus L.) seedlings in hydroponics. Soil Sci. Plant Nutr. 2012, 58, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Bie, Z.; He, S.; Hua, B.; Zhen, A.; Liu, Z. Improving cucumber tolerance to major nutrients induced salinity by grafting onto Cucurbita ficifolia. Environ. Exp. Bot. 2010, 69, 32–38. [Google Scholar] [CrossRef]
- Zhu, J.; Bie, Z.; Huang, Y.; Han, X. Effect of grafting on the growth and ion concentrations of cucumber seedlings under NaCl stress. Soil Sci. Plant Nutr. 2008, 54, 895–902. [Google Scholar] [CrossRef]
- Lee, J.M. Cultivation of grafted vegetables: Current status, grafting methods, and benefits. HortScience 1994, 29, 235–239. [Google Scholar] [CrossRef]
- Aslam, W.; Noor, R.S.; Hussain, F.; Ameen, M.; Ullah, S.; Chen, H. Evaluating morphological growth, yield, and post-harvest fruit quality of cucumber (Cucumis sativus L.) grafted on cucurbitaceous rootstocks. Agriculture 2020, 10, 101. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ballesta, C.M.; Alcaraz-Lópeza, C.; Muriesa, B.; Mota-Cadenasa, C.; Carvajal, M. Physiological aspects of root-stock-scion interactions. Sci. Hortic. 2010, 127, 112–118. [Google Scholar] [CrossRef]


{kind=link}
| Particulars | Grafted Seedling Cost (INR Plant−1) | |
|---|---|---|
| ‘Nefer’ (Single Fruiting) | ‘Terminator’ (Multiple Fruiting) | |
| F1 hybrid scion | 1.5 | 6.0 |
| Rootstock cost | 2.0 | 2.0 |
| Seedling production charges * | 4.5 | 5.5 |
| Final grafted seedling cost on site | 8.0 | 13.5 |
| Treatments | Dry Mass (g Plant−1) | Leaf Area (m2 Plant−1) | Vine Length (cm) | Node Number | Stem Girth (cm) | |
|---|---|---|---|---|---|---|
| Leaf | Stem | |||||
| Structures (S) | ||||||
| NVP | 74.68a | 28.50a | 1.21a | 3.76 | 51.00a | 1.29a |
| IPN | 72.25ab | 25.45b | 1.20a | 3.68 | 50.08a | 1.20b |
| SNH | 66.85b | 21.74c | 1.03b | 3.55 | 41.16b | 1.03c |
| Grafting (G) | ||||||
| Nefer | 56.24c | 19.32c | 1.00c | 3.12c | 41.37b | 0.87d |
| Nefer/NS 55 | 73.88b | 22.36b | 1.10b | 3.53bc | 45.14b | 1.13c |
| Terminator | 61.91c | 23.65b | 1.16b | 3.81ab | 50.07a | 1.30b |
| Terminator/NS 55 | 93.00a | 35.56a | 1.35a | 4.11a | 53.07a | 1.38a |
| Significance | ||||||
| S | NS | *** | *** | NS | *** | *** |
| G | *** | *** | *** | ** | *** | *** |
| S × G | NS | * | NS | NS | NS | NS |
| Treatments | Fruit Number (Plant−1) | Mean Fruit Weight (g) | Fruit Yield (kg Plant−1) | WP (kg m−3) |
|---|---|---|---|---|
| Structures (S) | ||||
| NVP | 26.33a | 145.64a | 3.85a | 31.11a |
| IPN | 23.81b | 141.75a | 3.37b | 27.24a |
| SNH | 16.04c | 128.00b | 2.06c | 18.17b |
| Grafting (G) | ||||
| Nefer | 17.88c | 132.86 | 2.40c | 19.79c |
| Nefer/NS 55 | 21.88b | 140.02 | 3.07b | 25.36b |
| Terminator | 22.39b | 137.78 | 3.15b | 25.93ab |
| Terminator/NS 55 | 26.08a | 143.20 | 3.75a | 30.94a |
| Significance | ||||
| S | *** | ** | *** | *** |
| G | *** | NS | *** | *** |
| S × G | NS | NS | NS | NS |
| Treatments | Fruit Length (cm) | Fruit Girth (cm) | Fruit Firmness (kg cm−2) | TSS (%) | Fruit Dry Matter (%) |
|---|---|---|---|---|---|
| Structures (S) | |||||
| NVP | 14.30a | 4.07a | 3.83 | 3.28 | 3.63a |
| IPN | 13.95b | 3.95b | 3.76 | 3.21 | 3.20b |
| SNH | 13.63c | 3.88c | 3.84 | 3.20 | 2.88c |
| Grafting (G) | |||||
| Nefer | 13.32c | 3.86b | 3.89 | 3.39a | 3.52a |
| Nefer/NS 55 | 13.98b | 3.93b | 3.83 | 2.99b | 3.12b |
| Terminator | 13.96b | 4.09a | 3.77 | 3.50a | 3.10b |
| Terminator/NS 55 | 14.57a | 3.97ab | 3.75 | 3.03b | 3.12b |
| Significance | |||||
| S | ** | ** | NS | NS | *** |
| G | ** | ** | NS | *** | ** |
| S × G | NS | NS | NS | * | NS |
| Treatments | L* | a* | b* |
|---|---|---|---|
| Structures (S) | |||
| NVP | 88.86a | −53.06 | 9.81 |
| IPN | 88.83a | −53.03 | 8.93 |
| SNH | 87.85b | −53.22 | 9.32 |
| Grafting (G) | |||
| Nefer | 85.58c | −48.71d | 3.74a |
| Nefer/NS 55 | 85.79c | −49.84c | 5.23b |
| Terminator | 92.51a | −58.16a | 1.52c |
| Terminator/NS 55 | 90.17b | −55.69b | 1.31d |
| Significance | |||
| S | * | NS | NS |
| G | *** | *** | *** |
| S × G | NS | *** | *** |
| Treatments | Total Chlorophyll (µg mL−1) | WP (-bars) | PSII (Fv/Fm) |
|---|---|---|---|
| Structures (S) | |||
| NVP | 15.08a | 5.87 | 0.827a |
| IPN | 12.46b | 5.75 | 0.822a |
| SNH | 14.27a | 5.75 | 0.784b |
| Grafting (G) | |||
| Nefer | 11.66c | 6.16b | 0.802b |
| Nefer/NS 55 | 15.08a | 5.50a | 0.816a |
| Terminator | 13.35b | 6.15b | 0.802b |
| Terminator/NS 55 | 15.67a | 5.33a | 0.823a |
| Significance | |||
| S | *** | NS | *** |
| G | *** | *** | ** |
| S × G | NS | NS | NS |
| Treatments | N | P | K | Ca | Mg | S |
|---|---|---|---|---|---|---|
| Structures (S) | ||||||
| NVP | 60.7a | 2.1b | 16.8c | 47.0b | 4.0 | 2.0a |
| IPN | 46.3b | 2.4a | 20.2b | 52.5a | 4.0 | 2.0a |
| SNH | 41.1c | 2.5a | 30.4a | 52.2a | 4.2 | 1.7b |
| Grafting (G) | ||||||
| Nefer | 50.6 | 2.1c | 20.1b | 48.7b | 7.2a | 1.6c |
| Nefer/NS 55 | 50.5 | 2.4ab | 21.2b | 49.8b | 4.2b | 2.0b |
| Terminator | 48.1 | 2.2bc | 24.4a | 50.2b | 3.1c | 1.7c |
| Terminator/NS 55 | 48.3 | 2.6a | 24.1a | 53.6a | 1.4d | 2.3a |
| Significance | ||||||
| S | *** | ** | *** | *** | NS | *** |
| G | NS | *** | *** | ** | *** | *** |
| S × G | *** | NS | *** | * | *** | *** |
| Treatments | Cu | Mn | Zn | Fe |
|---|---|---|---|---|
| Structures (S) | ||||
| NVP | 3.05a | 68.13a | 13.92b | 73.95b |
| IPN | 1.99b | 48.64c | 8.51c | 92.21a |
| SNH | 1.90b | 61.95b | 16.10a | 92.88a |
| Grafting (G) | ||||
| Nefer | 2.59a | 58.67 | 13.53 | 88.55 |
| Nefer/NS 55 | 2.26bc | 62.38 | 13.66 | 86.84 |
| Terminator | 2.07c | 57.74 | 12.65 | 84.76 |
| Terminator/NS 55 | 2.34b | 59.52 | 11.54 | 85.23 |
| Significance | ||||
| S | *** | *** | *** | *** |
| G | ** | NS | NS | NS |
| S × G | * | NS | NS | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khapte, P.S.; Kumar, P.; Panwar, N.R.; Burman, U.; Rouphael, Y.; Kumar, P. Combined Influence of Grafting and Type of Protected Environment Structure on Agronomic and Physiological Traits of Single- and Cluster-Fruit-Bearing Cucumber Hybrids. Agronomy 2021, 11, 1604. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081604
Khapte PS, Kumar P, Panwar NR, Burman U, Rouphael Y, Kumar P. Combined Influence of Grafting and Type of Protected Environment Structure on Agronomic and Physiological Traits of Single- and Cluster-Fruit-Bearing Cucumber Hybrids. Agronomy. 2021; 11(8):1604. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081604
Chicago/Turabian StyleKhapte, Pratapsingh Suresh, Pradeep Kumar, Nav Raten Panwar, Uday Burman, Youssef Rouphael, and Praveen Kumar. 2021. "Combined Influence of Grafting and Type of Protected Environment Structure on Agronomic and Physiological Traits of Single- and Cluster-Fruit-Bearing Cucumber Hybrids" Agronomy 11, no. 8: 1604. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081604