
Trichoderma-Enriched Vermicompost Extracts Reduces Nematode Biotic Stress in Tomato and Bell Pepper Crops

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vermicompost Production and Trichoderma Strains Inoculation
2.2. Growth of Bell Pepper and Tomato Plants
2.3. M. Incognita Inoculation
2.4. Water-Soluble Fraction Obtained from the Vermicomposts
2.5. Gas Exchange Measurements
2.6. Nutrient Concentration on Plants Tissues
2.7. Statistical Analysis
3. Results
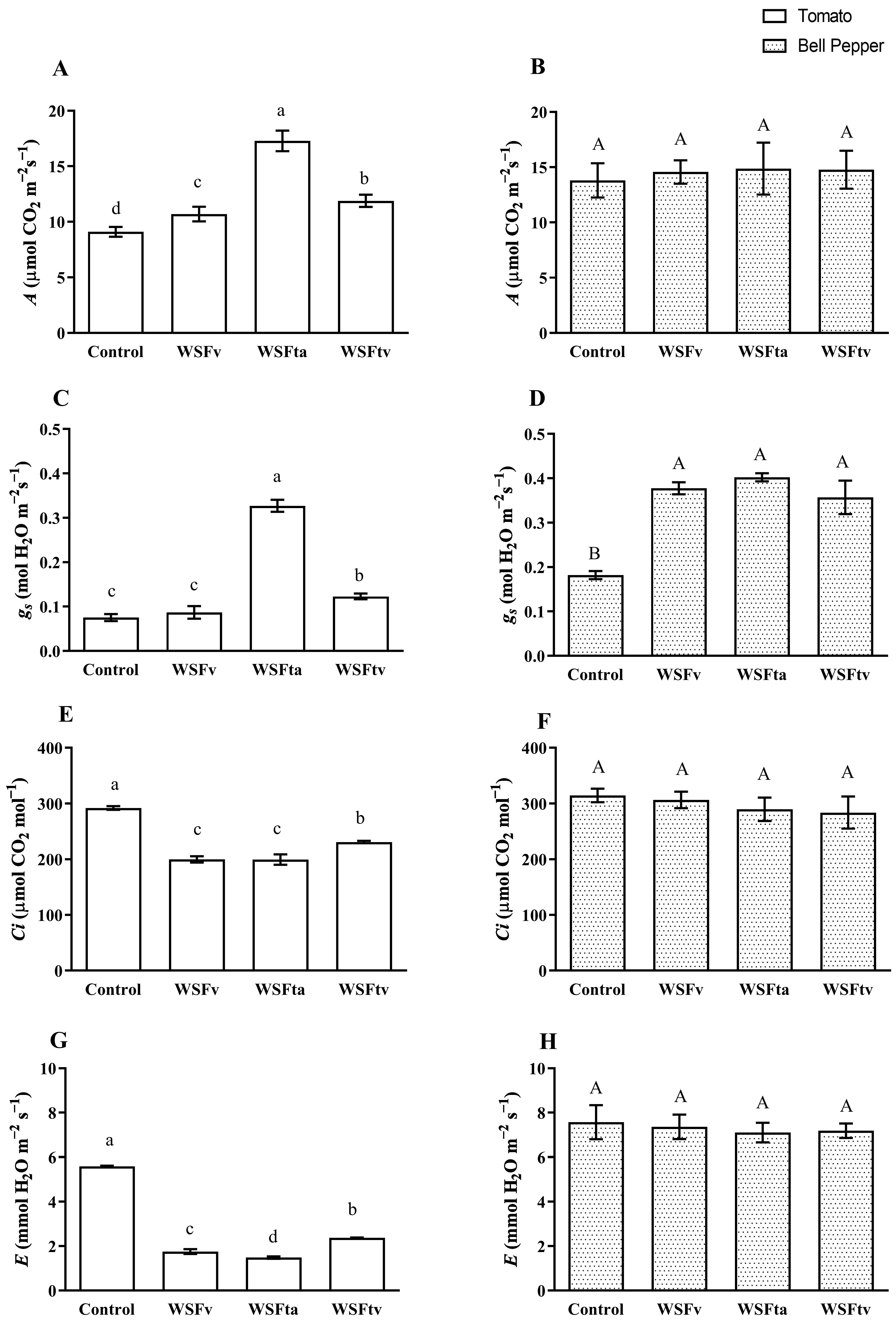
3.1. Gas Exchange Measurements
3.1.1. Photosynthetic Rate (A)
3.1.2. Stomatal Conductance (gs)
3.1.3. Internal CO2 Concentration (Ci) and Transpiration Rate (E)
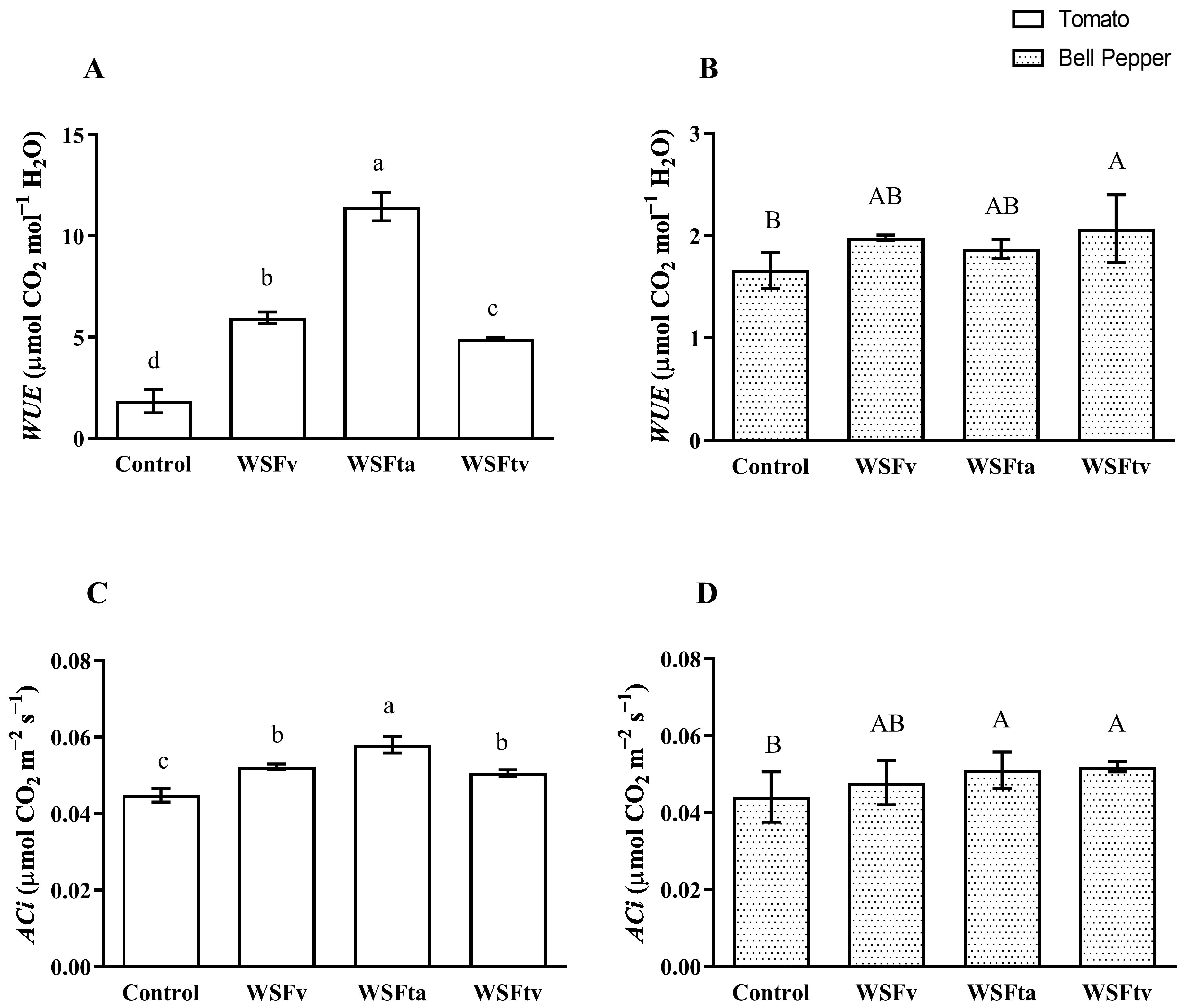
3.1.4. Water Use Efficiency (WUE) and Carboxylation Capacity (ACi)
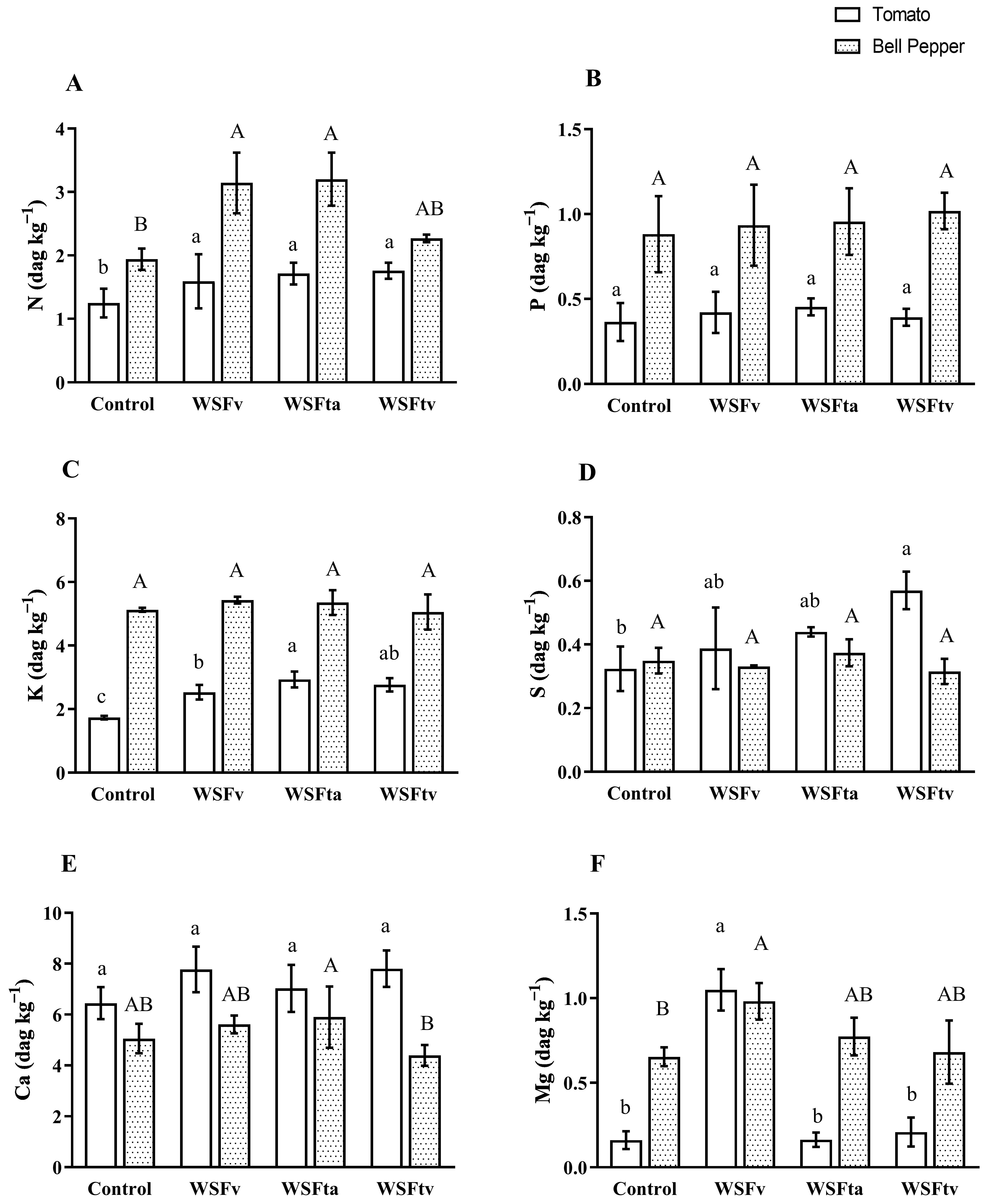
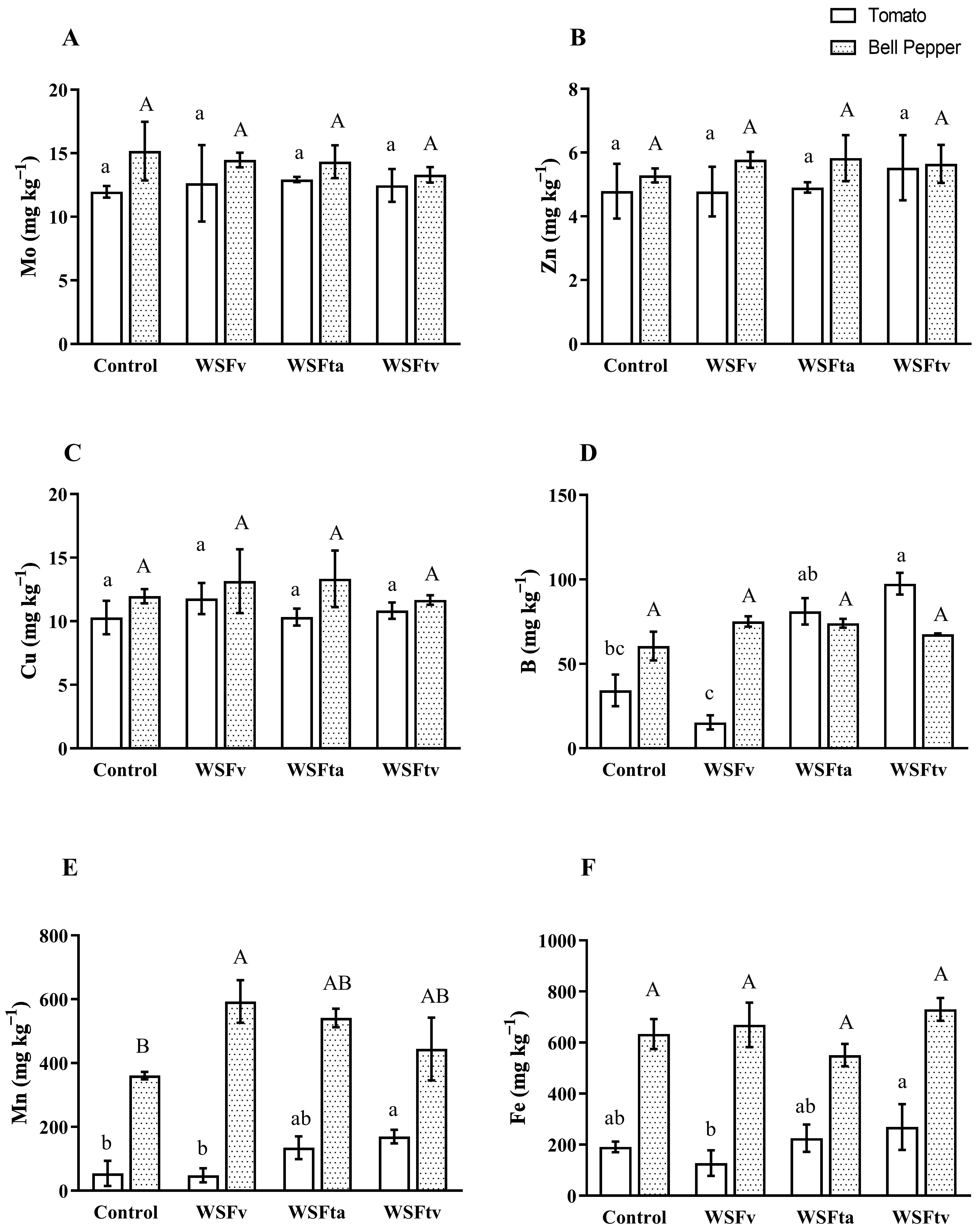
3.2. Nutrient Uptake
3.2.1. Macronutrients
3.2.2. Micronutrients
3.3. Effect of Adding the Different WSF
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coyne, D.L.; Cortada, L.; Dalzell, J.J.; Claudius-Cole, A.O.; Haukeland, S.; Luambano, N.; Talwana, H. Plant-Parasitic Nematodes and Food Security in Sub-Saharan Africa. Annu. Rev. Phytopathol. 2018, 56, 381–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asif, M.; Khan, A.; Siddiqui, M.A.; Tariq, M. Sustainable Management of Root Knot Nematode Meloidogyne incognita through Organic Amendment on Solanum lycopersicum L. Asian J. Biol. 2016, 1, 1–8. [Google Scholar] [CrossRef]
- Fuller, V.L.; Lilley, C.J.; Urwin, P.E. Nematode resistance. New Phytol. 2008, 180, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Kyndt, T.; Nahar, K.; Haegeman, A.; De Vleesschauwer, D.; Höfte, M.; Gheysen, G. Comparing systemic defence-related gene expression changes upon migratory and sedentary nematode attack in rice. Plant Biol. 2012, 14, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Bartlem, D.G.; Jones, M.; Hammes, U.Z. Vascularization and nutrient delivery at root-knot nematode feeding sites in host roots. J. Exp. Bot. 2014, 65, 1789–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strajnar, P.; Širca, S.; Urek, G.; Šircelj, H.; Železnik, P.; Vodnik, D. Effect of Meloidogyne ethiopica parasitism on water management and physiological stress in tomato. Eur. J. Plant Pathol. 2012, 132, 49–57. [Google Scholar] [CrossRef]
- Lu, P.; Davis, R.F.; Kemerait, R.C.; Van Iersel, M.W.; Scherm, H. Physiological effects of Meloidogyne incognita infection on cotton genotypes with differing levels of resistance in the greenhouse. J. Nematol. 2014, 46, 352–359. [Google Scholar] [PubMed]
- Sharma, I.P.; Sharma, A.K. Mycorrhizal colonization and phosphorus uptake in presence of PGPRs along with nematode infection. Symbiosis 2019, 77, 185–187. [Google Scholar] [CrossRef]
- Khan, M.R.; Mohiddin, F.; Ahamad, F. Inoculant rhizobia suppressed root-knot disease, and enhanced plant productivity and nutrient uptake of some field-grown food legumes. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 68, 166–174. [Google Scholar] [CrossRef]
- Chen, P.; Tsay, T. Effect of crop rotation on Meloidogyne spp. and Pratylenchus spp. populations in strawberry fields in Taiwan. J. Nematol. 2006, 38, 339–344. [Google Scholar] [PubMed]
- Ferris, H.; Griffiths, B.S.; Porazinska, D.L.; Powers, T.O.; Wang, K.H.; Tenuta, M. Reflections on plant and soil nematode ecology: Past, present and future. J. Nematol. 2012, 44, 115–126. [Google Scholar] [PubMed]
- Nelson, S.; Locascio, S.; Allen, L.; Dickson, D.; Mitchell, D. Soil Flooding and Fumigant Alternatives to Methyl Bromide in Tomato and Eggplant Production. HortScience 2002, 37, 1057–1060. [Google Scholar] [CrossRef] [Green Version]
- Sikder, M.; Vestergård, M. Impacts of Root Metabolites on Soil Nematodes. Front. Plant Sci. 2020, 10, 1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albehadeli, Y.; Mamarabadi, M.; MahdiKhani, E. Possibility of the biocontrol of Meloidogyne javanica using the fungus Trichoderma harzianum under greenhouse condition. Plant Arch. 2019, 19, 47–51. [Google Scholar]
- de Medeiros, H.A.; De Araújo Filho, J.V.; De Freitas, L.G.; Castillo, P.; Rubio, M.B.; Hermosa, R.; Monte, E. Tomato progeny inherit resistance to the nematode Meloidogyne javanica linked to plant growth induced by the biocontrol fungus Trichoderma atroviride. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbss, L.B.; Dos Santos, T.C.; Pittarello, M.; De Souza, S.B.; Ramos, A.C.; Busato, J.G. Alleviation of iron toxicity in Schinus terebinthifolius Raddi (Anacardiaceae) by humic substances. Environ. Sci. Pollut. Res. 2018, 25, 9416–9425. [Google Scholar] [CrossRef] [PubMed]
- Zanin, L.; Tomasi, N.; Zamboni, A.; Sega, D.; Varanini, Z.; Pinton, R. Water-extractable humic substances speed up transcriptional response of maize roots to nitrate. Environ. Exp. Bot. 2018, 147, 167–178. [Google Scholar] [CrossRef]
- Jindo, K.; Olivares, F.L.; Malcher, D.J.D.P.; Sánchez-Monedero, M.A.; Kempenaar, C.; Canellas, L.P. From Lab to Field: Role of Humic Substances Under Open-Field and Greenhouse Conditions as Biostimulant and Biocontrol Agent. Front. Plant Sci. 2020, 11, 426. [Google Scholar] [CrossRef]
- Canellas, L.P.; Canellas, N.O.A.; Soares, T.S.; Olivares, F.L. Humic acids interfere with nutrient sensing in plants owing to the differential expression of TOR. J. Plant Growth Regul. 2019, 38, 216–224. [Google Scholar] [CrossRef]
- Radin, A.M.; Warman, P.R. Effect of Municipal Solid Waste Compost and Compost Tea as Fertility Amendments on Growth and Tissue Element Concentration in Container-Grown Tomato. Commun. Soil Sci. Plant Anal. 2011, 42, 1349–1362. [Google Scholar] [CrossRef]
- Zandonadi, D.; Santos, M.P.; Caixeta, L.S.; Marinho, E.B.; Peres, L.E.P.; Façanha, A.R. Plant proton pumps as markers of biostimulant action. Sci. Agric. 2016, 73, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Busato, J.G.; Zandonadi, D.B.; De Sousa, I.M.; Marinho, E.B.; Dobbss, L.B.; Mól, A.R. Efeito do extrato húmico solúvel em água e biofertilizante sobre o desenvolvimento de mudas de Callophyllum brasiliense. Pesqui. Florest. Bras. 2016, 36, 161. [Google Scholar] [CrossRef] [Green Version]
- Pane, C.; Palese, A.M.; Celano, G.; Zaccardelli, M. Effects of compost tea treatments on productivity of lettuce and kohlrabi systems under organic cropping management. Ital. J. Agron. 2014, 9, 153. [Google Scholar] [CrossRef] [Green Version]
- Zaccardelli, M.; Pane, C.; Villecco, D.; Palese, A.M.; Celano, G. Compost tea spraying increases yield performance of pepper (Capsicum annuum L.) grown in greenhouse under organic farming system. Ital. J. Agron. 2018, 13, 229–234. [Google Scholar] [CrossRef]
- Zandonadi, D.B.; Matos, C.R.R.; Castro, R.N.; Spaccini, R.; Olivares, F.L.; Canellas, L.P. Alkamides: A new class of plant growth regulators linked to humic acid bioactivity. Chem. Biol. Technol. Agric. 2019, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Vujinović, T.; Zanin, L.; Venuti, S.; Contin, M.; Ceccon, P.; Tomasi, N.; Pinton, R.; Cesco, S.; De Nobili, M. Biostimulant Action of Dissolved Humic Substances from a Conventionally and an Organically Managed Soil on Nitrate Acquisition in Maize Plants. Front. Plant Sci. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C.A.; Arancon, N.Q.; Sherman, R.L. (Eds.) Vermiculture Technology; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar] [CrossRef]
- Arancon, N.; Van Cleave, J.; Hamasaki, R.; Nagata, K.; Felts, J. The influence of vermicompost water extracts on growth of plants propagated by cuttings. J. Plant Nutr. 2020, 43, 176–185. [Google Scholar] [CrossRef]
- dos Santos Pereira, T.; Gomes Macêdo, A.; da Silva, J.; Borges Pinheiro, J.; Monteiro de Paula, A.; Biscaia, D.; Busato, J.G. Water-extractable fraction of vermicomposts enriched with Trichoderma enhances the growth of bell pepper and tomato as well as their tolerance against Meloidogyne incognita. Sci. Hortic. 2020, 272, 109536. [Google Scholar] [CrossRef]
- Carneiro, R.M.D.G.; Almeida, M.R.A. Técnica de Eletroforese Usada no Estudo de Enzimas de Nematóides de Galhas Para Identificação de Espécies. Nematol. Bras. 2001. Available online: https://nematologia.com.br/files/revnb/25_1.pdf (accessed on 18 August 2021).
- Eisenback, J.D.; Triantaphyllou, H.H. Root-knot Nematodes: Meloidogyne species and races. In Manual of Agricultural Nematology; CRC Press: Boca Raton, FL, USA, 1991; pp. 281–286. [Google Scholar]
- Taylor, A.L.; Sasser, J.N. Biology, Identification and Control of Root-Knot Nematodes (Meloidogyne Species); Department of Plant Pathology, North Carolina State University, United States Agency for International Development: Raleigh, NC, USA, 1978. [Google Scholar]
- Silva, F.M.D.O.; Lichtenstein, G.; Alseekh, S.; Rosado-Souza, L.; Conte, M.; Suguiyama, V.F.; Lira, B.S.; Fanourakis, D.; Usadel, B.; Bhering, L.L.; et al. The genetic architecture of photosynthesis and plant growth-related traits in tomato. Plant Cell Environ. 2018, 41, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Blumwald, E. The Roles of ROS and ABA in Systemic Acquired Acclimation. Plant Cell 2015, 27, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zandonadi, D.B.; Busato, J.G. Vermicompost humic substances: Technology for converting pollution into plant growth regulators. Int. J. Environ. Sci. Eng. Res. 2012, 3, 73–84. [Google Scholar]
- Mishra, S.; Wang, K.-H.; Sipes, B.S.; Tian, M. Suppression of Root-Knot Nematode by Vermicompost Tea Prepared From Different Curing Ages of Vermicompost. Plant Dis. 2017, 101, 734–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, Y.; Meon, S.; Ismail, M.R.; Ali, A. Trichoderma-fortified compost extracts for the control of choanephora wet rot in okra production. Crop. Prot. 2008, 27, 385–390. [Google Scholar] [CrossRef]
- Frydrych, J. Factors affecting photosynthetic productivity of sweet pepper and tomatoes grown in CO2-enriched atmosphere. Acta Hortic. 1984, 271–278. [Google Scholar] [CrossRef]
- Onkendi, E.M.; Kariuki, G.M.; Marais, M.; Moleleki, L.N. The threat of root-knot nematodes (Meloidogyne spp.) in Africa: A review. Plant Pathol. 2014, 63, 727–737. [Google Scholar] [CrossRef] [Green Version]
- Olaetxea, M.; Mora, V.; Baigorri, R.; Zamarreño, A.M.; García-Mina, J.M. The Singular Molecular Conformation of Humic Acids in Solution Influences Their Ability to Enhance Root Hydraulic Conductivity and Plant Growth. Molecules 2020, 26, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tan, S.N.; Wong, W.S.; Ng, C.Y.L.; Teo, C.H.; Ge, L.; Chen, X.; Yong, J.W.H. Mass spectrometric evidence for the occurrence of plant growth promoting cytokinins in vermicompost tea. Biol. Fertil. Soils 2014, 50, 401–403. [Google Scholar] [CrossRef]
- Ramos, A.C.; Dobbss, L.B.; Santos, L.A.; Fernandes, M.S.; Olivares, F.L.; Aguiar, N.O.; Canellas, L.P. Humic matter elicits proton and calcium fluxes and signaling dependent on Ca2+-dependent protein kinase (CDPK) at early stages of lateral plant root development. Chem. Biol. Technol. Agric. 2015, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Jannin, L.; Arkoun, M.; Ourry, A.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F.; et al. Microarray analysis of humic acid effects on Brassica napus growth: Involvement of N, C and S metabolisms. Plant Soil 2012, 359, 297–319. [Google Scholar] [CrossRef]
- García, A.C.; Santos, L.A.; de Souza, L.G.A.; Tavares, O.C.H.; Zonta, E.; Gomes, E.T.M.; García-Mina, J.M.; Berbara, R.L.L. Vermicompost humic acids modulate the accumulation and metabolism of ROS in rice plants. J. Plant Physiol. 2016, 192, 56–63. [Google Scholar] [CrossRef]
- Mora, V.; Bacaicoa, E.; Zamarreño, A.-M.; Aguirre, E.; Garnica, M.; Fuentes, M.; García-Mina, J.-M. Action of humic acid on promotion of cucumber shoot growth involves nitrate-related changes associated with the root-to-shoot distribution of cytokinins, polyamines and mineral nutrients. J. Plant Physiol. 2010, 167, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Meon, S.; Wallace, H.; Fisher, J. Water relations of tomato (Lycopersicon esculentum Mill. cv. Early Dwarf Red) infected with Meloidogyne javanica (Treub), Chitwood. Physiol. Plant Pathol. 1978, 13, 275–281. [Google Scholar] [CrossRef]
- Dorhout, R. Water Transport Through Tomato Roots Infected with Meloidogyne incognita. Phytopathology 1991, 81. [Google Scholar] [CrossRef]
- Briar, S.S.; Fonte, S.J.; Park, I.; Six, J.; Scow, K.; Ferris, H. The distribution of nematodes and soil microbial communities across soil aggregate fractions and farm management systems. Soil Biol. Biochem. 2011, 43, 905–914. [Google Scholar] [CrossRef]
- Nardi, S.; Muscolo, A.; Vaccaro, S.; Baiano, S.; Spaccini, R.; Piccolo, A. Relationship between molecular characteristics of soil humic fractions and glycolytic pathway and krebs cycle in maize seedlings. Soil Biol. Biochem. 2007, 39, 3138–3146. [Google Scholar] [CrossRef]
- Van Der Werf, A.; Kooijman, A.; Welschen, R.; Lambers, H. Respiratory energy costs for the maintenance of biomass, for growth and for ion uptake in roots of Carex diandra and Carex acutiformis. Physiol. Plant. 1988, 72, 483–491. [Google Scholar] [CrossRef]
- Bakshi, A.; Moin, M.; Madhav, M.S.; Kirti, P.B. Target of rapamycin, a master regulator of multiple signalling pathways and a potential candidate gene for crop improvement. Plant Biol. 2018, 21, 190–205. [Google Scholar] [CrossRef] [PubMed]
- Canellas, N.O.A.; Olivares, F.L.; Canellas, L.P. Metabolite fingerprints of maize and sugarcane seedlings: Searching for markers after inoculation with plant growth-promoting bacteria in humic acids. Chem. Biol. Technol. Agric. 2019, 6. [Google Scholar] [CrossRef]
- Fu, L.; Wang, P.; Xiong, Y. Target of Rapamycin Signaling in Plant Stress Responses. Plant Physiol. 2020, 182, 1613–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.; Singh, A.; Srivastava, M.; Shanker, K.; Pandey, R. Plant-microbe interactions endorse growth by uplifting microbial community structure of Bacopa monnieri rhizosphere under nematode stress. Microbiol. Res. 2019, 218, 87–96. [Google Scholar] [CrossRef]
- Singh, M.; Awasthi, A.; Soni, S.K.; Singh, R.; Verma, R.K.; Kalra, A. Complementarity among plant growth promoting traits in rhizospheric bacterial communities promotes plant growth. Sci. Rep. 2015, 5, 15500. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop | A | Gs | Ci | E | WUE | Aci | |
|---|---|---|---|---|---|---|---|
| µmol CO2 m−2 s−1 | mol H2O m−2 s−1 | µmol CO2 m−2 s−1 | mol H2O m−2 s−1 | µmol CO2 m−2 s−1 | µmol CO2 m−2 s−1 | ||
| Tomato | Control | 9.10 | 0.08 | 292.23 | 5.59 | 1.83 | 0.044 |
| WSF | 13.29 | 0.18 | 210.11 | 1.87 | 7.44 | 0.053 | |
| Variation (%) | 46 (↑) | 125 (↑) | 28 (↓) | 67 (↓) | 307 (↑) | 20 (↑) | |
| Bell pepper | Control | 13.80 | 0.18 | 314.64 | 7.57 | 1.66 | 0.044 |
| WSF | 14.74 | 0.38 | 293.45 | 7.22 | 1.92 | 0.050 | |
| Variation (%) | 7 (↑) | 111 (↑) | 7 (↓) | 6 (↓) | 16 (↑) | 14 (↑) |
| Crop | N | P | K | Ca | Mg | S | |
|---|---|---|---|---|---|---|---|
| dag kg−1 | |||||||
| Tomato | Control | 1.25 | 0.36 | 1.73 | 6.45 | 0.16 | 0.32 |
| WSF | 1.69 | 0.42 | 2.74 | 7.54 | 0.47 | 0.47 | |
| Variation (%) | 35 (↑) | 17 (↑) | 58 (↑) | 17 (↑) | 194 (↑) | 47 (↑) | |
| Bell pepper | Control | 1.94 | 0.99 | 5.13 | 5.06 | 0.65 | 0.34 |
| WSF | 2.87 | 0.99 | 5.28 | 5.30 | 0.81 | 0.34 | |
| Variation (%) | 48 (↑) | 0 (-) | 3 (↑) | 5 (↑) | 25 (↑) | 0 (-) | |
| Crop | Mo | Zn | Cu | B | Mn | Fe | |
|---|---|---|---|---|---|---|---|
| dag kg−1 | |||||||
| Tomato | Control | 11.97 | 4.79 | 10.29 | 34.30 | 54.30 | 191.00 |
| WSF | 12.68 | 5.07 | 10.98 | 64.60 | 117.71 | 207.63 | |
| Variation (%) | 6 (↑) | 6 (↑) | 7 (↑) | 88 (↑) | 111 (↑) | 9 (↑) | |
| Bell pepper | Control | 15.17 | 5.28 | 11.97 | 60.53 | 360.67 | 633.33 |
| WSF | 14.93 | 5.75 | 12.72 | 72.24 | 526.11 | 650.00 | |
| Variation (%) | 2 (↓) | 9 (↑) | 6 (↑) | 19 (↑) | 46 (↑) | 3 (↑) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos Pereira, T.; Monteiro de Paula, A.; Ferrari, L.H.; da Silva, J.; Borges Pinheiro, J.; Navas Cajamarca, S.M.; Jindo, K.; Pupo Santos, M.; Zandonadi, D.B.; Busato, J.G. Trichoderma-Enriched Vermicompost Extracts Reduces Nematode Biotic Stress in Tomato and Bell Pepper Crops. Agronomy 2021, 11, 1655. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081655
dos Santos Pereira T, Monteiro de Paula A, Ferrari LH, da Silva J, Borges Pinheiro J, Navas Cajamarca SM, Jindo K, Pupo Santos M, Zandonadi DB, Busato JG. Trichoderma-Enriched Vermicompost Extracts Reduces Nematode Biotic Stress in Tomato and Bell Pepper Crops. Agronomy. 2021; 11(8):1655. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081655
Chicago/Turabian Styledos Santos Pereira, Tiago, Alessandra Monteiro de Paula, Luis Henrique Ferrari, Juscimar da Silva, Jadir Borges Pinheiro, Sabrina Magaly Navas Cajamarca, Keiji Jindo, Mirella Pupo Santos, Daniel Basílio Zandonadi, and Jader Galba Busato. 2021. "Trichoderma-Enriched Vermicompost Extracts Reduces Nematode Biotic Stress in Tomato and Bell Pepper Crops" Agronomy 11, no. 8: 1655. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081655