Potential of Wild Relatives of Wheat: Ideal Genetic Resources for Future Breeding Programs



1
Seed and Plant Improvement Institute, Agricultural Research, Education and Extension Organization (AREEO), Karaj P.O. Box 3183964653, Iran
2
Department of Agronomy and Plant Breeding, Faculty of Agriculture, Bu-Ali Sina University, Hamedan 65178-38695, Iran
3
Botany Unit, Finnish Museum of Natural History, University of Helsinki, P.O. Box 7, FI-00014 Helsinki, Finland
4
Department of Agronomy and Plant Breeding, Agricultural College, University of Tehran, Karaj 31587-77871, Iran
*
Authors to whom correspondence should be addressed.
Agronomy 2021, 11(8), 1656; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081656
Submission received: 22 July 2021
/
Revised: 16 August 2021
/
Accepted: 18 August 2021
/
Published: 19 August 2021
Abstract
:Among cereal crops, wheat has been identified as a major source for human food consumption. Wheat breeders require access to new genetic diversity resources to satisfy the demands of a growing human population for more food with a high quality that can be produced in variable environmental conditions. The close relatives of domesticated wheats represent an ideal gene pool for the use of breeders. The genera Aegilops and Triticum are known as the main gene pool of domesticated wheat, including numerous species with different and interesting genomic constitutions. According to the literature, each wild relative harbors useful alleles which can induce resistance to various environmental stresses. Furthermore, progress in genetic and biotechnology sciences has provided accurate information regarding the phylogenetic relationships among species, which consequently opened avenues to reconsider the potential of each wild relative and to provide a context for how we can employ them in future breeding programs. In the present review, we have sought to represent the level of genetic diversity among the wild relatives of wheat, as well as the breeding potential of each wild species that can be used in wheat-breeding programs.
1. Introduction
Climate change and subsequent increasing abiotic and biotic stresses threaten food security globally, as they can hinder the potential yield performance, increase the number of pests and diseases generations, alter synchrony between plants and pests, increase risk of invasion by migratory pests, increase incidence of insect-transmitted plant diseases, and reduce the effectiveness of biological control, especially for natural enemies [1,2]. One worthwhile strategy for increasing crop productivity and stability that may be applied in a wide range of environments is ‘crop genetic improvement’ through the introgression of novel genes, QTLs, and even novel alleles from wild relatives to local or modern varieties [3]. As such, breeders must simultaneously both improve the genetic background and reduce the impact of environmental stresses on the grain production. Among cereal crops, wheat has a significant role in supplying the 20% of all calories consumed by people worldwide. Due to climate change, mainly caused by biotic and abiotic stresses, the demand for wheat bread wheat is predicted to increase dramatically in the future as the global human population increases. Hence, wheat production will have a vital bearing on food security in the coming decades [4].
After the green revolution, numerous bread wheat varieties were released by agronomists and breeders in different parts of the world. Although this task helped to increase the breeding population of wheat, the genetic basis of the wheat was narrowed through the shearing of the breeding lines and various breeding cycles, which in turn decreased species variability. Wild relatives of wheat offer great possibilities for breeders to develop new varieties with a more appropriate genetic background for various agricultural systems [5]. Hence, the use of an ancestors’ gene pool is an appropriate strategy for developing new superior bread wheat cultivars. Wheat wild relatives are closely related species that have a long history in wheat breeding, mostly for abiotic stress tolerance. These species are identified as the critical resources required to sustain global food supply [4]. The tribe Triticeae, which is part of the Pooideae subfamily of the grass family Poaceae, includes the genera Triticum and Aegilops L. These relatives provide important gene pools for wheat-breeding programs, since they are connected to the most important agricultural crop, Triticum aestivum L. [6]. In this review, we have summarized the phylogenetic relationships among wild wheats and their potential applications in wheat breeding.
2. The Trend of Bread-Wheat Evolution
Wheat was domesticated from its wild relatives during the pre-pottery Neolithic (PPN) period nearly 12,000 years ago, in the Middle East’s Fertile Crescent, which included several wide ranges of geographical zones from Lebanon, Israel, Jordan, and Syria via southeast Turkey, as well as into Iraq and western Iran via the Tigris and Euphrates rivers [7] (Figure 1). Research conducted by Weide et al. [8] reported that the first hybridization between wild wheats occurred in the west of Iran.
Triticum and Aegilops are two key genera which include various wild wheat with different genomic constitutions and which have played direct or indirect roles in wheat domestication. Table 1 shows some of the key articles that have denoted how wheat evolution occurred.
Although phylogenetic relationships among wild relatives of wheat have been extensively reviewed by many researchers e.g., [24], we report here an information flow diagram for the trend of wheat domestication (Figure 2). This diagram shows wheat’s evolution process and a general viewpoint of relationships among the close relatives of common wheat, which descended from a 3 million-year-old common ancestor and gave rise to the Aegilops and Triticum taxa [25]. Briefly, the T. urartu Tumanian ex Gandilyan (with A-genome) and A. speltoides Taush. (with B genome) have created a tetraploid form of wheat known as emmer wheat (T. turgidum ssp. dicoccoides (Körn) Aschers and Graebner Thell.), due to the natural hybridization processes occurring several hundred thousand years ago [13]. T. turgidum L. ssp. dicoccum Thell. and free-threshing T. turgidum L. spp. durum Desf. are the result of the domestication of wild emmer. A second hybridization event occurred between A. tauschii Coss. and T. turgidum, which resulted in the hexaploid bread wheat T. aestivum L. [13]. Consequently, the polyploidy genome of T. aestivum with an AABBDD genome contains three sub-genomes AA, BB, and DD from T. urartu, A. speltoides, and A. tauschii, respectively. Furthermore, Table 2 shows all wild wheat species with various alien genomic constitutions.
3. Levels of Genetic Diversity in Wheat Germplasm
3.1. Phenotypic Diversity
Wild wheat species are highly diverse and variable in terms of agronomic and morphological traits. The distribution of these species in different ecological zones has resulted in several species with unique traits. The first studies on phenotypic diversity in wild wheat species and landraces referred to a study conducted by Percival [28], which described diversity through plant height, spike length, number of spikelets, and straw quality among landraces of bread wheat from Iran. Jaradat [29] indicated significant genotypic variation for developmental and yield traits among Jordanian wheat landraces. Anker et al. [30] investigated 76 accessions of wild wheat species including T. boeoticum ssp. boeoticum, T. boeoticum ssp. thaoudar., T. urartu and T. monococcum in terms of 17 agronomic and morphological characteristics. The germplasm accessions showed significant variation for flowering data, length of spike, length of awns, and length of anthers. Arzani et al. [31] investigated a collection of wild relatives including 24 Aegilops and Triticum accessions belonging to Ae. crassa, Ae. umbellulata, Ae. caudata, Ae. cylindrica, Ae. glabra, Ae. squarrosa, Ae. triuncialis, T. monococcum, T. dicoccides, and T. compactum from different regions of Iran. These authors estimated genetic diversity using 25 agronomical and morphological characters, and finally reported that for qualitative qualities, there is a lot of variance across species, as well as inter- and intraspecific variation for quantitative traits. Later, a study involving 254 accessions of Ae. tauschii from several regions of Iran indicated a high diversity for spike-related characteristics such as rachis node length, spike width, number of seed per spike, rachis node width, spikelet glum length, seed length, and width [32]. The evaluation of 68 accessions of Ae. tauschii ssp. strangulata and Ae. tauschii ssp. tauschii sampled from different regions of Iran indicated significant variation within populations for most phenotypic traits, such as peduncle length, plant height, awn length, number of fertile tillers, spike length, and biological yield. Likewise, an experiment using 180 accessions of Aegilops and Triticum genera was performed by Pour-Aboughadareh et al. [33] to evaluate genetic diversity using different agronomic characters. In this study, the Shannon–Weaver (HSW) and Nei’s (HN) genetic diversity parameters revealed intermediate to high phenotypic diversity for most traits in Triticum and Aegilops species. These researchers recommended that the genetic diversity among studied Iranian Aegilops and Triticum species such as Ae. crassa, Ae. cylindrica and Ae. umbellulata can provide new insights for the rediscovery of valuable agronomic traits, which can be exploited for the improvement and adaptation of common wheat. In view of this fact, several studies have been performed to measure the extent and pattern of phenotypic diversity in the wheat germplasm collections using different traits. Table 3 shows that attempts have been made to characterize the estimate of genetic diversity levels in wild relatives of wheat using phenotypic data.
3.2. Plant Genetic Resources and Molecular Diversity
Over the last three decades, many studies have been performed to characterize the genetic diversity in ancestral and wild relatives of wheat based on different molecular-marker systems. A general pattern change in plant genetic resources (PGR) was triggered by the political system, as well as the advancement of molecular biology and electronic data processing, as seen here for wheat [53]:
- I.
- II.
- Inclusion of underutilized and neglected crop varieties [55]. Some plants are likely extinct in conventional farming zones, although landraces for others have only recently been discovered. Wild relatives have become increasingly important in wheat breeding, with Secale, Aegilops, Hordeum, and other genera being used in addition to wild Triticum varieties [53].
- III.
- Techniques for determining taxonomic diversity both inside and between taxonomic groups. Insights into population dynamics and evolution are being gained thanks to modern technology.
- IV.
- Genetic loss is an issue in genebanks as well [56].
- V.
- Landraces have a wide range of morphological variation. Breeders are less familiar with infraspecific classification schemes, which are helpful for characterization and handling.
- VI.
- Measurement techniques: Molecular markers are used to classify genetic variations on a basic basis, without taking into account ecological adaptation.
- VII.
- Traditional assessment programs can be expanded by genebanks. Pre-breeding will become more important.
- VIII.
- Under long-term storage and reproduction conditions, genebanks are accurate and cost-effective, but strategic reproduction principles are needed [57].
Genetic resources are critical for the current and future sustainability of world wheat production. They include a wide variety of genetic variation, which is essential for increasing and sustaining wheat production potential by providing novel sources of resistance and tolerance to biotic and abiotic stressors [58]. The germplasm preserved is particularly rich in wild crop relatives, traditional farmer cultivars, and ancient cultivars, all of which form a significant genetic diversity reserve. Ex situ or in situ conservation of material protects against genetic degradation and provides a source of resilience to biotic and abiotic stressors, and enhances quality and yield characteristics for future crop improvement. Breeders use well-adapted cultivars from particular regions to build modern high-yielding wheat cultivars, which are an assembly of genes or gene combinations. International agricultural research has greatly increased the availability of broadly adaptable, genetically varied germplasm [58]. Traditional approaches are often overlooked, whereas new possibilities are exaggerated. Landrace preservation in genebanks and on farms is difficult. The role of PGR is traditional. Orthodox approaches can be used to study landraces, but molecular methods can answer complex questions [53]. According to numerous studies, a high level of genetic diversity among wild relatives of wheat has been reported, which may refer to their natural distributions in a wide range of ecosystems and even in natural hybridization among various species. Hence, herein, we have only listed some studies that use at least one molecular marker system to investigate the level of genetic diversity, the genetic makeup of the population, as well as complementary genetic tests, such as association mapping or gene cloning. In accordance with previous reports, we found that, among wild wheats, Ae. tauschii Coss. has been the most subjected to genetic studies (Table 4). Indeed, this confirms the potential of this species for future wheat breeding programs.
4. Potential of Wild Relatives for Use in Wheat Breeding Programs
Farmers’ desired qualities (yield potential, large seed, high seed weight) and breeders’ preferred characteristics (high seed weight, large seed, yield potential) both benefit from genetic variety in wheat germplasm (e.g., biotic and abiotic resistance) [103]. It is clear that an increasing number of genetic diversity studies on wheat and their wild relatives are revealing the ideal potential of these natural resources and also suggesting ways of devising targeted methods to exploit the diverse existence of ex-situ germplasm collections [5]. To date, various attempts have been made to improve common wheat by using genetic diversity in wild relatives. Arguments for the greater use of wheat relatives’ include: the taxonomic relationship between wheat and its relatives, cross-compatibilities, F1 fertility, and subsequent progeny, exploration and utilization of this natural resource, and availability and regional financial support based on their geographic distribution [104]. In this section, we highlight some representative examples in which wild wheats showed specific ability versus different environmental stresses.
4.1. Drought
The extensive use of advanced lines has resulted in the loss of genetic basis of bred varieties of wheat that has, in some cases, led to increased susceptibility to various abiotic stresses [103]. Drought or a lack of water is one of the most significant environmental factors affecting global wheat production. Drought tolerance is a multi-dimensional quantitative characteristic that has a dramatic impact on plant development stages [105]. The impacts of drought stress have been intensely studied in wheat and its relatives. Among wild wheats, Aegilops species have been considered as an ideal source for improving the genetic background of bread wheat to tolerate drought stress.
Several Aegilops species, such as Ae. speltoides, Ae. tauschii, and Ae. geniculata, have indicated the ability to overcome drought stress [106,107]. Introgression of drought tolerance to bread wheat was achieved by hybridization between durum wheat and Ae. tauschii species. DNA fingerprinting of synthetic hexaploid (SHs), materials showed that they can respond well to water deficit due to their excellent features, such as a longer root system and higher soluble carbohydrates [108]. The use of carbon isotope discrimination (∆) has been proposed to estimate transpiration efficiency, water use efficiency, and drought tolerance in different wild wheat species [109]. This method was performed by Waines et al. [110] on several Aegilops species. Their results revealed a high intraspecific variation in most species, and, among the tested species, ∆ values were highest in Ae. speltoides. Wild emmer wheat (T. dicoccoides) can donate excellent drought tolerance compared to other wheat species. It is clear that this ancestral species can better overcome drought conditions compared to durum wheat [111]. Therefore, T. dicoccoides is one of the most important sources for drought tolerance and is highly proper as a donor for developing agronomical and physiological features related to tolerance in cultivated wheat species. In a comprehensive study conducted by Pour-Aboughadareh et al. [112], a set of 180 accessions belonging to 12 wild and domesticated species along with two commercial tolerant and sensitive control varieties were examined in terms of shoot dry mass and chlorophyll fluorescence parameters under two water regimes: optimum irrigation (FC = 100%) and drought stress (FC = 30%). A considerable number of wild wheats with alien genomes, such as Ae. crassa (DM genome), T. urartu (Au genome), Ae. cylindrica (DC genome), and Ae. caudata (C genome), indicated a higher tolerance to drought stress as compared with the domesticated genotypes and tolerant control, reflecting greater drought adaptations in these species.
Upon exposure to drought stress, plants undergo many physiological and molecular changes. One of these changes is the production of reactive oxygen species (ROS) in different plant tissues. The ROS family––including singlet oxygen (1O2), superoxide (O2–), hydroxyl radical (OH), and hydrogen peroxide (H2O2)––act as signal molecules in the response to drought stress. On the other hand, increasing the production of ROS may result in cellular damage and finally cell death [113]. Therefore, antioxidant gene overexpression could induce tolerance to this stress. Plants have antioxidant systems in place to scavenge excess ROS and protect themselves from the harmful consequences of oxidative stress by generating various antioxidants. A detailed description of antioxidant mechanisms can be found in the review by Bose et al. [114].
Some research has focused on the state of antioxidant activities and the ROS pathway in bread wheat germplasm. Recently, Ahmadi et al. [115] examined the antioxidant activities in several wild relatives of wheat in response to drought stress. The wild accessions used in their study revealed more activities in the expression of antioxidative enzymes—including catalase (CAT), guaiacol peroxidase (GPX), superoxide dismutase (SOD), and ascorbate peroxidase (APX)—than the cultivated wheats. Remarkably, four wild wheats—T. boeoticum, Ae. crassa, Ae. cylindrica, and Ae. Umbellulata—responded well to severe drought stress (FC = 25%) by elevating enzymatic antioxidants as the primary defense system, which contributes to the cell’s ability to maintain oxidative equilibrium. In another study, Pour-Aboughadareh et al. [116] showed a high rate of variability among Ae. cylindrical, Ae. tauschii, and Ae. crassa species, along with T. aestivum landraces in terms of several photosynthetic and physiological traits under severe water deficit stress. These authors stated that, among 200 samples related to these species, 19 accessions from Ae. crassa and 1 accession from Ae. tauschii indicated considerable tolerance to drought conditions through the maintenance of their biomass and other physiological capacities. When the top-ranked accessions were evaluated in terms of biochemical traits, Ae. crassa showed better potential for scavenging the ROS through higher activity of CAT, APX, GPX, and peroxidase (POD) antioxidant enzymes [117].
The root system, as an important organ for the uptake of water and nutrients, is the first part of a plant that senses water shortage [118]. Recently, many efforts have been made to indicate the relationships among root system features and drought tolerance adaptability in the wild relatives of wheat under drought conditions. Ahmadi et al. [119] focused on seedling root architectural traits in the whole collection of wheat germplasm containing 180 accessions along with two control bread wheat genotypes under two water regimes. They reported a high level of root variability in some wild wheat responding to drought stress. When the root system of the different species was compared under drought stress conditions, it was found that four species of wild wheat—Ae. speltoides (As a putative B genome), Ae. cylindrica (DC genome), Ae. neglecta (UM genome), and Ae. tauschii (D genome)—had a great ability to extend their root system. In another study, Djanaguiraman et al. [120] showed that Ae. speltoides was an ideal candidate for improving the root system architecture in wheat. These authors examined 48 Chinese spring wheat-alien chromosome lines belonging to Ae. longissima, Ae. geniculate, Ae. searsii, Ae. peregrine, Ae. speltoides, Th. intermedium, L. racemosus, and D. villosum in terms of several physiological and root features under drought stress. The results showed that the wheat-alien chromosome lines with chromosome segments from Ae. speltoides were identified as drought tolerant, so that their acceptable tolerance was associated with a deep and profuse root-system structure.
4.2. Salinity
Salinity stress is another environmental challenge that dramatically limits wheat production worldwide [121]. It has been reported that more than 800 million hectares of the world’s total land are affected by salinity [122]. As a result, attempts to improve wheat salt tolerance are critical for long-term agriculture and might considerably increase wheat output. The screening of wheat germplasm has a significant role in breeding programs aimed at improving salinity tolerance [123]. Although a number of salt-tolerance traits have been characterized by Colmer et al. [124], we mention several key studies that have introduced wild wheats with superior salt-tolerance. The effects of salinity on plant growth include osmotic pressure, oxidative stress, ionic toxicity, and nutritional imbalance, which in turn severely limits the development and function of all plants.
Plants often have a variety of tolerance mechanisms that enable them to reproduce in marine environments, including reducing root Na+ uptake and limiting salt concentration in the cytosol [125]. Effective removal of Na+ from roots and shoots and the isolation of specific organelles, intracellular compartments, or cells of Na+ are other important processes for surviving salinity stress [126]. Thus, the ability to maintain high K+ and low Na+ concentrations in leaves is associated with salt tolerance. Research on different wild wheats under salinity stress has indicated that some Aegilops species can withstand heavy salt stress and have a greater tolerance than other wheat species [127]. For example, Gorham et al. [128] revealed that Ae. tauschii has a higher Na+:K+ ratio in their leaves than other species. In fact, this result suggested that there is a close relationship between Na+ exclusion and D genome that enhanced the Na+:K+ ratio. In a study conducted by Ahmadi et al. [129], Ae. tauschii along with Ae. neglecta species responded well to severe salinity treatment at the seedling stage. Kiani et al. [130] found that Ae. cylindrica accessions differed significantly in terms of physiological characteristics, including their susceptibility to salt stress throughout the vegetative stage. The results from the last two reports have postulated the key impact of C genome on salt exclusion in this wild wheat. Furthermore, the observation of Ahmadi et al. [131] in the screening of wheat germplasm to explore the source of salinity tolerance revealed a high potential for some Aegilops and Triticum in coping with salinity stress. These researchers looked at root and shoot biomass, physiological parameters, and ROS scavenging enzymes in a core collection of 181 accessions that included landrace genotypes and various wild relatives of wheat. Based on their results, several wild species with alien genomes, such as Ae. cylindrica (DC), Ae. caudata (C), and T. boeoticum (Ab), revealed an appropriate response to salinity stress by increasing enzymatic antioxidants. These results support the fact that there is a good potential for salinity tolerance in wild relatives of wheat, which in turn reveals new insights for reconsidering the associations between wheat progenitors and salinity tolerance.
4.3. High Temperature
Crop growth rates are influenced by temperature conditions. The high temperature, as another environmental stress, has limited wheat production in the world. As a result of global warming, all crops may undergo many changes in their growth and development. Hence, an exploration of ideal sources of variability to high temperature tolerance for wheat and other field crops is required. High temperature fluctuations have a significant impact on plant development in a variety of ways. Chlorophyll concentration and the photosynthetic ability of leaves are reduced due to this stress [132]. Because thylakoid membranes are one of the most vulnerable cellular structures to this stress, there is a clear link between chlorophyll concentration and thylakoid membrane damage caused by high temperatures [133]. Thus, damaged thylakoid membranes leads to loss of chlorophyll content and reduced photosynthesis capacity [134]. In recent years, analyzing chlorophyll fluorescence has been one of the most popular approaches in plant physiology to measure photosynthetic activity. It plays a critical role in understanding key photosynthetic systems, plant responses to environmental change, and ecological diversity [135]. The analysis of chlorophyll fluorescence components, such as initial fluorescence (Fo), maximal quantum efficiency (Fv/Fm), and primary yield of photochemistry (Fv/Fo), is a critical approach for determining the integrity of the internal mechanisms within a leaf during photosynthetic activities. It also provides a precise method for assessing damage to photosystem II centers (PSII) and, as a result, identifying plants that are resistant to certain stressors, particularly high temperatures [112,136,137]. Fo is a measure of the stability of the light-harvesting complex among chlorophyll fluorescence components, and it is enhanced in the leaves of plants under high temperatures compared to optimal conditions [138]. In theory, an increase in this parameter might be read as a decrease in the rate constant of energy trapping by PSII, which could be the result of the light-harvesting complex becoming physically disconnected from the PSII core [139]. In genotypes of Ae. tauschii and Ae. speltoides, an increase in the Fo parameter has been recorded under high temperature stress [140]. Using this parameter, researchers have illustrated that Ae. tauschii had greater thermostability of the photosynthetic mechanism. High-temperature stress was also applied to wheat species at anthesis and maintained for 16 days, resulting in reductions in chlorophyll content, the number of grains per spike, grain weight, and grain yield per plant of 38, 40, 56, and 70%, respectively [141]. Pradhan et al. [141] indicated that among Aegilops species, Ae. geniculate and Ae. speltoides displayed a greater tolerance to high temperatures for grain yield (58–61% decline than optimum temperature), while Ae. longissima yields showed an 84% decline.
4.4. Low Temperature
Low temperature, as the fourth important abiotic stress, affects wheat growth and production in many regions of the world. Extreme cold stress causes a change in cell architecture by preventing exterior water mobility and converting internal water to a crystalline state. Low temperature stress-sensitive plants, as a rule, are unable to continue growing over extended periods of time and respond to cold by altering metabolite concentrations [142]. Generally, efforts to improve cold tolerance through existing plant genetic materials appear to have been unsuccessful because of limited genetic diversity for this characteristic. Wild relatives of wheat offer a potential source of additional genetic variability which could be utilized to improve the cold tolerance of wheat, and, up to now, numerous studies have focused on these species under different environmental conditions. Nonetheless, no work has been undertaken to investigate the whole set of wild relatives in response to cold stress. Only a few examples could be found. For instance, Limin and Fowler [143] found that Ae. tauschii responded well to freezing stress, while all species related to Sitopsis section were sensitive. However, it has been reported that Ae. speltoides and T. turdidum have a higher tolerance than other wild species [144,145]. In a study conducted by Stankova et al. [146], among different Aegilops species, Ae. cylindrica revealed the highest cold tolerance, whereas Ae. genioulata and Ae. biuncialis were identified as sensitive relatives. They also demonstrated that Ae. triuncialis and Ae. neglecta have intermediate tolerance. Moreover, Masoomi-Aladizgeh et al. [147] evaluated several Ae. tauschii accessions under controlled cold conditions, demonstrating the capability of this species in response to freezing stress.
4.5. Biotic Stresses
Unexpected biotic stressors similar to other environmental stresses continuously pose a danger to wheat production. Pathogen resurgences have occurred from the monoculture of contemporary wheat cultivars with limited genetic diversity, posing a danger to wheat supply [148]. Several living creatures, such as fungi, viruses, insects, nematodes, arachnids, and weeds, induce biotic stress in plants. Pathogenic fungi, among, other biotic stressors, pose a serious threat to wheat production across the world. Stripe rust, stem rust, leaf rust, powdery mildew, head blight, and other diseases negatively affect wheat production and grain quality worldwide. Yellow rust has historically caused and continues to cause substantial and severe losses in vulnerable wheat cultivars all throughout the world [149]. Aphid, hessian fly, green bug, and borers are among the most common insect pests that damage wheat.
Only a few of these biotic stress resistance genes in wheat have been identified and cloned thus far. Species from the primary gene pool (Triticum spp.), secondary gene pool (e.g., T. timopheevii), and tertiary gene pool (e.g., Aegilops), are among the R gene donors [150]. The wild relatives of wheat may be divided into main, secondary, and tertiary gene pools based on their genomic makeup [151,152]. These gene pools provide a plentiful supply of disease and pest resistance genes in wheat. Species with homologous genomes to farmed wheat make up the main gene pool. T. aestivum, T. turgidum, and species with the A and D genomes—T. monococcum, T. urartu, T. boeoticum, and Ae. tauschii––make up this group [152]. Many genes providing disease and insect pest resistance have been transmitted by hybridization and backcrossing procedures, and some of them are still used in cultivar improvement [153,154]. An active collection of 280 Ae. tauschii accessions is kept at the Punjab Agricultural University (PAU) in Ludhiana, India. Various biotic stressors, including as leaf rust, stripe rust, powdery mildew, and Karnal bunt (KB), have been shown to carry resistance genes in these accessions. The KB resistance of Ae. tauschii is quite strong. The polyploid Triticum and Aegilops species, which share at least one genome with wheat, make up the secondary gene pool of bread wheat. Many resistance genes have been given by these species, which have been utilized in cultivar development [151].
It has been reported that, in wheat or its wild relatives, more than 240 rust resistance genes have been identified and formally recognized, the majority of which are race-specific resistance genes [153]. The QTL regulating stripe rust resistance in T. monococcum was found on chromosome 2A (QYrtm.pau-2A), whereas the QTL controlling stripe rust resistance in T. boeoticum was found on chromosome 5A (QYrtm. pau-5A). Co-introgression of T. boeoticum sequences related to stripe rust-resistant QTL, QYrtb.pau-5A [155] revealed that one stripe rust-resistant gene from T. boeoticum acc. pau5088 was verified to be introgressed in cultivated wheat. T. durum was used as a bridge species to introduce leaf and stripe rust resistance genes from diploid species Ae. umbellulata and Ae. caudata [155,156]. In general, through the use of different approaches, a number of resistance genes have been transferred from wild relatives to bread wheat. Among the identified resistance genes, most numbers are related to leaf rust, followed by powdery mildew, and green bug [157]. In general, Table 5 indicates some identified or transferred resistance genes in various Aegilops species.
5. Transcriptome Analysis Uncovers Hidden Information about the Benefits of Wild Relative Potentials
Progress in biotechnological tools, such as the GeneChip® Wheat Genome array and RNA sequencing, have indicated that the transcripts associated with starch biosynthesis and defense proteins in hexaploid bread wheat are expressed 2–3 weeks after anthesis. The transcripts that are most numerous in growing plants, on the other hand, were associated with storage proteins that are expressed during the developmental phase [198,199,200,201]. While transcriptome analysis was done in hexaploid wheat during grain production [202,203], bread wheat and its wild progenitors have yet to be subjected to a thorough transcriptional characterization. Studying the expression of these key genomes at specific stages during grain development in bread wheat and its wild relatives can be very beneficial. This will be useful to understand the changes in wheat grain consistency that occurred during the transition from ancient bread wheat to modern bread wheat. Kaushik et al. [201] used RNAseq to study the development of bread wheat grains and their diploid progenitor cells to evaluate gene expression patterns and monitor differentially expressed genes. To analyze gene expression profiles and screen differentially expressed genes, they used transcriptomics methods to compare the expression of key genes in hexaploid bread wheat and its three diploid parents during the entire grain production process. It can help to better understand the molecular mechanisms of metabolic pathways involved in the production and regulation of key components of grain growth. In addition, transcriptome sequencing of bread wheat and its ancestors is used to study the genetics of wheat endosperm growth.
5.1. Grain Development Related Proteins
Global wheat security is under threat due to the need for wheat to feed the world’s growing population [203]. Wheat grain production is thought to be a significant determinant of yield and flour content. Understanding the process of wheat grain growth, as well as identifying key candidate genes that perform important functions during grain development, is crucial. Reduced expression of TabZIP60 has been reported to improve grain yield and nitrogen uptake through RNAi interference by upregulating TaNADH-GOGAT expression. Compared to the wild type, the grain yield of the overexpression line increased by 16.6–26.8% [204]. Previous studies have used genome-wide association analysis study (GWAS) to classify new genomic regions related to grain yield and characteristics related to grain yield in 123 synthetic hexaploid wheat (SHW) grown under drought stress. Among them, 35648 derived single nucleotide polymorphism (SNP) genotypes were sequenced [205]. They found that SHW has a significant genetic diversity in grain yield and yield-related traits. Under drought stress, GWAS in 123 synthetic hexaploid wheat established multiple new genomic regions or haplotype blocks related to grain yield and yield-related traits. Most marker-trait associations (MTAs) are found in the genome, and some are annotated with drought stress functions. This further demonstrates that the given MTA is reliable. MTAs are also linked to a variety of characteristics on other chromosomes, but only inside the genes that have the same annotation. This led to the identification of candidate genes from the same gene family that are considered important in grain yield and yield-related characteristics in drought-stressed SHWs [205]. Wild relatives of wheat may be useful for studying different facets of evolution and growth involved in the production of bread wheat. Grain production involves several biochemical and physiological processes that occur in a variety of tissues [206]. There are major stages including fertilization, multinucleate endosperm, cellularization, and early grain filling, full grain filling, and desiccation [207]. Grain production is critical for completing a crop’s life cycle because it accumulates various nutrient stocks as well as embryo development and maturation. The nutritional and economic value of wheat grain is determined by the accumulation of these nutrient stocks [201].
5.2. Nutrient Reservoir
Nutrient reserves (NRs), carbohydrate metabolism (CM), and defense proteins (DPs) are the most common genes involved in wheat grain production [206]. Carbohydrates are the most common, followed by storage proteins in NR. However, carbohydrates are metabolized by complex mechanisms affecting a variety of genes. The genes involved in anabolism and catabolism have been studied in CM, which has been designated as a separate class. Wheat genotypes contain different kinds of storage proteins in terms of consistency and quantity because the coding region of wheat storage protein is highly polymorphic [208]. Starch is the most prevalent nutrient in T. aestivum, followed by storage proteins and lipids [199]. Thus, the synthesis and aggregation of starch and storage proteins are crucial for the growth of wheat grain. Albumins, globulins, and gluten are the most common storage proteins. Glutenins and gliadins are two types of gluten. Glutenins are often made up of proteins with high and low molecular weights [201,208]. Prior research has revealed that the glutenin and gliadin subunits of wild wheat relatives varied greatly. Ahmadi et al. [209] used unique molecular markers to characterize the allelic variation of glutenin and gliadin in 180 Aegilops and Triticum accessions obtained from different parts of Iran. They revealed that the allelic status of Glu-3A and Gli-As.2 was related to genomic constitutions, resulting in a large variance in the gliadin and glutenin subunits in di- and tetraploid wild relatives of Ae. umbellulata, Ae. caudata, Ae. tauschii, and T. urartu genomes. As a result, previous studies may yield new avenues for improving dough consistency by reconsidering the connections between other progenitors and wild cousins. Researchers have been encouraged to hunt for unique and beneficial alleles to create new kinds that are better suited to new uses, given the discovery of this very varied gene pool.
5.3. Carbohydrate Metabolism
The most significant metabolic process in the formation of wheat grains is carbohydrate metabolism. According to research on the wheat grain proteome, 21% of differentially expressed proteins are involved in carbohydrate metabolism [210]. β-glucosidase is one of the most common enzymes studied in wheat. It is responsible for the hydrolysis of carbohydrates. Adenosine diphosphate ADP pyrophosphorylase, starch synthase [211], starch branching enzyme, and starch debranching enzyme [206] are the four major enzymes involved in the production of starch, the main ingredient of endosperm.
5.4. Defense Proteins during Grain Development
The preservation of developing grains is the next critical phase during grain production after the deposition of storage materials. To resist or postpone several biotic and abiotic pressures, plants can trigger a range of molecular and biochemical defense mechanisms through irreconcilable interactions between the host and biotic/abiotic stimuli, resulting in a variety of signaling pathways [212]. Several investigations have proven that amylase/trypsin inhibitors regulate wheat defense responses by inhibiting the enzyme activities of amylase and trypsin in pests [210,213,214]. Insect defensive responses are considered to be aided by the Bowman–Birk family of cysteine-rich proteases [215]. Considering the change in genome size, 55.62, 55.92, 68.13, and 103.33 million reads were generated for the genome species of T. urartu, Ae. speltoides, Ae. tauschii, and T. aestivum, respectively. There are significant differences in the genome size [216] and the number of genes [216,217,218,219] between wheat hexaploid and its diploid ancestors. The number of genes expressed in hexaploid wheat is not proportional to the size of its genome, although it is slightly higher than that in diploid wheat. It has also been suggested that the size of the genome in the polyploid genome is not proportional to gene expression [220,221,222]. In hexaploid wheat, however, a review of half the number of reads showed significantly fewer transcripts. Researchers discovered that Ae. speltoides (BB) shared the most transcripts of the three progenitors. When sub-genomic research was performed, it was discovered that the number of transcripts in hexaploid bread wheat’s B genome was somewhat greater [201].
5.5. Carbohydrate and Protein Related Transcripts
Kaushik et al. [201] also analyzed the gene expression of three main protein classes: nutrient reserves, defense protein, and carbohydrate metabolism. Albumin, globulin, prolamin, and glutenin are storage proteins formed primarily during grain growth. The gene ontology study of transcriptome data of hexaploid bread wheat and its diploid ancestors shows that nutrient pool behavior is remarkably rich. When ancient diploid wheat and modern bread wheat were compared, albumin was found to be highly upregulated in bread wheat. As a result, Ae. speltoides had a higher expression of this gene in diploid progenitors (BB) [201]. Prolamins are proline and glutamine-rich proteins that make up roughly half of the nitrogen in wheat grains [216,223,224]. Gliadins, which belong to the prolamin family, are the oldest wheat nutrient reservoirs [225]. When compared to T. monococcum, these proteins were upregulated in bread wheat. Bread wheat transcriptome comparisons demonstrated an upregulation of α/β gliadin and γ-gliadin B in Ae. tauschii (DD and) Ae. speltoides (BB). The diploid progenitor AA genome (T. urartu) genotype exhibited a lower gliadin gene expression in this example. In comparison to its diploid progenitors, high molecular weight glutenins expressed more transition in bread wheat. Gluten is formed when gliadin and glutenin combine, and it is much higher in bread wheat than in its diploid progenitors. α-β Gliadins were found to be actively expressed in wheat endosperm from 11 days to 4 weeks after anthesis [226]. High molecular weight glutenins were also found to be most abundant during early grain filling, a finding that had previously been recorded in Norin 61 bread wheat [227].
5.5.1. Carbohydrate Biosynthesis Genes
In bread wheat and its diploid progenitors, many glucose metabolism-related genes are differently expressed. Starch biosynthesis is carried out by two types of enzymes: granule associated starch synthase and starch synthases [228,229]. When hexaploid wheat was related to its diploid progenitors, granule bound starch synthase (GBSS-I) was down-regulated, and starch synthase genes were upregulated. This finding shows that hexaploid bread wheat produces less amylose and more amylopectin during grain production than its progenitors [201]. In bread wheat, major starch aggregation occurs during the early stages of grain production [230]. In T. aestivum and Ae. tauschii, stage-specific GBSS expression data is also related to this discovery. The starch in the endosperm is broken down to produce glucose, which is then used by the scutellum for the growth of the embryo [231]. Compared with hexaploid bread wheat, the gene expression of some important and abundant enzymes in starch metabolism such as AMY3 amylase and α-glucosidase is reduced in Ae. speltoides and T. monococcum [201,232]. In the late stage of grain filling, the gene expression of carbohydrate catabolism enzymes such as amylase and glucosidase was higher, but the gene expression of carbohydrate anabolism enzymes (such as starch synthase bound to carbohydrate granules) was larger at first. Therefore, although carbohydrate biosynthesis occurs early in the filling process, the consumption of carbohydrates for energy production (ATP) occurs later. This is due to the reduced amount of carbohydrates available from sources [233].
5.5.2. Defense Proteins
Wheat grains are mainly composed of carbohydrates and protein, which account for about 80% of grain weight. It is essential to safeguard these nutrient reservoirs in wheat grains against biotic and abiotic influences and to keep them alive before germination by defense proteins. Many differentially expressed genes linked to defense proteins are identified when different phases of bread wheat grain production are compared [206,234]. When opposed to T. monococcum, Ae. tauschii, and Ae. speltoides, a trypsin inhibitor, which is implicated in herbivorous pest resistance [235], was upregulated in hexaploid wheat. This suggested that both Aegilops species have an almost identical voice, which is better than T. monococcum. In diploid progenitors, a stage-specific study of this gene revealed a related pattern. In bread wheat, the defense protein thionin was found to be upregulated [236,237]. However, stage-specific expression showed that expression was higher in the early stages in hexaploids, while in diploids it was higher in the later stages. Some defense proteins, such as Bowman Birk type trypsin inhibitors (pathogen inactivator) [238,239], subtilisin chymotrypsin inhibitors (known to suppress insect larvae) [240], and wheat monomeric amylase inhibitors [241], were also downregulated in hexaploid wheat. The expression of both defense-related genes was shown to be higher in hexaploids at the early stages of grain filling and lower in diploids during the later stages.
6. Dynamic Wheat Transcriptome and Small RNA in Wild Relatives of Wheat under Abiotic Stresses
The genome’s substructure aids in mapping dynamically transcribed sections to biotic and abiotic stressors during plant growth and adaptation. Transcriptome profiling was carried out on the wheat genome to provide a better understanding of both gene expression levels and harvests [242]. In 2004, 35 individual cDNA libraries representing extremely detailed developmental stages of different grains and seedling tissues were used to create microarrays with high-density from the publicly accessible wheat EST resource, which included 26,382 sequences [242]. However, due to the immobile existence of probes and the reliance on genome annotation quality, the microarrays provided insufficient expression proof. Indirect measurement of gene expression levels was found to be illustrative of cDNA arrays through hybridization signs, and expression levels were detected to be illustrative of the tenacity of genuine transcript estimates in individual tissues or cell lines [243]. A systematic analysis of the entire transcriptional environment was carried out using next-generation RNA sequencing. Even though it demonstrated alternate splicing, it failed to display genome-wide gene function in terms of quantity [244,245].
Allohexaploid wheat is one example of the analysis of genetic associations between the three homologous genomes (A, B, and D) because it has recently suffered two allopolyploidization events. Several experiments have used microarrays and other methods to equate resynthesized or normal wheat allopolyploids to their progenitors [246,247,248]. In this survey, the non-additive gene expression in allohexaploid offspring was evaluated using the average parental gene expression level or number (i.e., the father’s average (MPV)) [249]. Nonetheless, it has been found that additivity is more extensive than non-additivity [248]. MicroRNA (miRNA) and small interference RNA (siRNA) are small RNAs that control gene expression through post-transcriptional processes and epigenetic modifications [250,251]. In interspecific hybrids and allopolyploidies of Arabidopsis, changes in miRNA expression induce the non-additive expression of target genes, hindering developmental adaptability and vitality [252]. Subsequent studies have shown that the cis- and trans-regulation of miRNA and other genes can affect the normal changes in the biochemical and metabolic pathways that drive growth vigor and stress response [253,254]. In addition, by directing DNA methylation, siRNA, especially those related to transposable factors (TE), can act as a genomic shock absorber and control gene expression.
After polyploidization of wheat, the number of siRNAs corresponding to ET is significantly reduced, and it is rich in repeating sequences originating from TE (>80%), indicating that they play important roles during allohexaploidization [255]. Small RNA-mediated genome alteration and gene regulation are possibly involved in allohexaploidization, according to new evidence [256]. However, analysis of the homologous expression and limited abundance of RNA from individuals throughout the genome requires information from the genome sequence [257,258,259]. When these tools are combined with next-generation sequencing technologies, they can answer questions regarding how allohexaploidization affects homeolog expression and changes molecular pathways that lead to the nascent allohexaploid of wheat’s growth vigor and adaptation, as well as whether small RNAs play a role in this process [242]. T. aestivum originated as a hybrid between Ae. tauschii and T. turgidum and outcompeted its parents in growth adaptability and vigor following chromosomal doubling. To further understand the molecular foundation for this achievement, Li et al. [244] used recently available A and D genome sequences to undertake mRNA and small RNA transcriptome studies in nascent allohexaploid wheat and subsequent generations, their progenitors, and the natural allohexaploid cultivar Chinese Spring [244]. Expressed protein-coding genes were found to be uncommon but essential for growth vigor. In addition, a considerable number of protein-coding genes showed an advantage in the expression level of the parent, and genes whose total homologue expression level in the offspring were the same as that of T. turgidum showed an advantage in the expression level of the parent, which is presumably involved in growth and those whose expression was similar to that in Ae. Tauschii possibly involved in adaptation. Furthermore, upon polyploidization, a large proportion of microRNAs exhibited nonadditive expression, theoretically resulting in differential expression of essential target genes. In addition, in heterozygous progeny, an increased density of small interfering RNAs was observed for transposable D homozygotes binding elements, which may explain the biased repression of D homoeologs. These findings shed light on small RNA-mediated dynamic homoeolog control pathways that can play a role in nascent hexaploid wheat heterosis [244]. In conclusion, Table 6 summarizes the important microRNAs which have been identified in various wild species of wheat under different growing conditions.
7. Next-Generation Sequencing in Bread Wheat
Encoded genomic sequence information is needed to fully leverage wheat’s ability to feed the world’s rising population. Since the bread wheat genome is so large, about 17 gigabases, conventional sequencing methods are challenging to use. Next-generation sequencing (NGS) can sequence a large genome in a limited amount of time. Refseq V1, as an important standard quality in hexaploid wheat reference genomes, was created by the International Wheat Genome Sequencing Consortium (IWGSC). It offered details on the locus and order of 107,891 genes from 21 sequenced chromosomes and has allowed for the discovery of more than 4 million molecular markers [242,287]. NGS could also help researchers obtain various reference-related genomes from bread wheat and review genome-wide relationships, epigenetic functionalities, and population genetics, among other things. This review compiles and discusses the existing knowledge about the use of NGS in wheat science, taking into account the relevance of wheat genomics and NGS. Exome capture combined with NGS technology is an important method for analyzing the wheat genome in depth. This approach may be used to sequence whole exon complements in the genome [288].
NGS technology is now commonly used to analyze transcripts. The sequence tags generated by these technologies represent genes that are expressed without prior knowledge of the gene sequence. Next-generation transcriptome sequencing can be used to analyze gene expression, the structure of genomic loci, and the sequence variations present in expressed loci [289]. This can be achieved by de novo assembly of the transcriptome sequence data or by aligning the reads with the genomic sequence. NGS technology revolutionized genome biology and has begun to provide important resources for the improvement of wheat crops. Compared to all of the previous sequencing technologies, these technologies provide a faster and cheaper method of generating wheat sequence data. Although the large size and complexity of the wheat genome make the use of NGS difficult, advances in sequencing technology and bioinformatics tools have made the application of NGS in wheat increasingly feasible. This type of application will eventually allow for a comprehensive package of wheat genome sequencing and annotations of the genome sequence to express genes and genomic variation across sub-genomes [290]. The application of NGS technology provides the opportunity to study and understand the structure and evolution of polyploid crop genomes. Discovery of numerous genome-wide molecular SNP markers has begun in wheat, which may have the largest and fastest impact on crop improvement. In the next few years, the challenge may shift from NGS analysis of the wheat genome to the association of sequence variation with heritable agronomic traits. As in rice, corn, and sorghum, the wheat genome sequence will advance our understanding of the genetic basis of agronomic traits, as well as aid the development of new technologies for improving wheat crops [290].
Genome-Wide Association Studies and SNP Microarrays
It is challenging to investigate wheat genetic variants. The most effective approach used to search for genetic mutations and their effects on phenotypes is to perform GWAS. Previously, genome-wide observations for various wheat genomes were obtained using single nucleotide polymorphism (SNP) microarrays. When compared to NGS, SNP microarray is less expensive and allows for a fast sequencing of the related genes. It aids in the comprehension of the marker traits connected in the creation of maps, as well as the ancestral relationships between populations. However, due to the sensitivity of recognition and the ability to recognize genetic variation, NGS outperforms SNP microarrays [291]. In addition, NGS-based genotyping will reveal new genetic variations and allow detailed analysis of population genetics.
Arora et al. [292] revealed that the variation of grain size in the germplasm of Ae. Tauschii and the genetic basis was analyzed using a GWAS. The grain length, width, and weight of 177 Ae.tauschii germplasm evaluations in 3 years showed an almost normal distribution, and the variation was 1.74, 1.75, and 2.82 times, respectively. These lines were genetically characterized using the Genotype Sequencing Protocol (GBS), which produced 11,489 SNP markers. Genetic diversity analysis revealed the existence of two distinct subgroups in Ae. tauschi. Based on GBS markers, the genetic similarity between germplasms was calculated, and GWAS was performed using 114 non-redundant germplasms and 5249 SNP markers. A total of 17 SNPs related to grain size traits distributed on seven chromosomes were revealed, of which 4444 had the most significant marker-trait associations with 6D, 5D, and 2D. For some related SNP markers, candidate genes related to cell division and differentiation were identified. Further efforts to validate these loci will help to understand their role in determining the grain size and allelic diversity in the current germplasm, as well as the effect on grain size when transferred to the bread wheat background [292].
In hexaploid wheat, GBS has recently become an important method for GWAS. GBS examines millions of SNP markers, which are used to discover genetic variation through the multiple analyses of samples from different genomes. Where the interested gene or locus genes necessitates map-based gene cloning, it is a popular method in forward genetics [293]. Outbreeding the mutant organism to create a population of mapping is part of map-based cloning in plants [294]. GWAS helps to promote crop improvement by raising awareness of Market Portrait Associations (MTAs). Lozada et al. [295] reported that, by using different panels of 239 soft red winter wheat (T. aestivum L.) genotypes, GWAS was performed on grain yield (GY), grain yield components, and agronomic characteristics over two growing seasons and eight site years. Analysis of variance showed the significant effects of the environment andgenotypes in the GY, and its components. Compared to other traits, including plant height and kernel weight, the narrow heritability of the GY is moderate. Using compressed linear mixed models and 5715 single nucleotide polymorphism markers to measure eight traits, 112 significant MTAs were detected. The MTA of GY and agronomic traits are consistent with the previously reported QTLs of winter and spring wheat. The highly significant MTA of the GY shows an overall negative allele effect on the minor alleles, indicating that the breeder has been selected for these alleles. The markers associated with multiple traits observed on chromosomes 1A, 2D, 3B, and 4B have slight positive effects and serve as potential targets for marker-assisted reproduction to select and enhance the GY and its related traits. After labeling verification, these multi-portrait sites have potential that can be used by MAS to enhance the GY and adaptability of soft red winter wheat [295].
GWAS was performed on 208 series durum wheat panels using 6211 DArTseq SNP. This panel has been phenotyped for 2 years under the conditions of yield potential (YP), drought stress (DT), and heat stress (HT). GWAS has identified the most important trait-marker linkages on chromosomes 2A and 2B, and the markers in the study explain variation in traits. Common markers were identified for stress tolerance indices: stress susceptibility index, stress tolerance, and stress tolerance index estimated for sub-DT and HT traits. The GWAS from the three environments of irrigation and stress and its comparison with the trait itself and the stress index determined the QTL hotspots on chromosomes 2A and 2B [296]. Thirteen traits were evaluated in 373 Ae. tauschii germplasm grown under normal drought stress conditions and simulated with polyethylene glycol, and performed a genome-wide association study using 7185 SNP markers [297]. They used a general linear model and a mixed linear model to determine 208 and 28 SNPs related to all traits, and both models detected 25 important SNPs distributed throughout the genome. Public database searches revealed several candidate/flanking genes related to drought resistance, which were divided into three categories based on the type of protein encoded. In another study, Mehrabi et al. [298] evaluated a set of T. durum landraces using SNP markers and root system features, as well as some agronomic characteristics, over three stages of plant growth and development. They reported that most significant MTAs were identified on chromosome B. Furthermore, 167 QTLs were detected for root system and agronomic traits, among which 16 QTLs for root-related traits overlapped with different measured agronomic traits.
8. Concluding Remarks
Plant genetic resources are valuable assets for humankind. Among crop wild relatives, wild wheat species have an important role in durum and bread wheat evolution. Hence, thousands of Aegilops and Triticum accessions have been collected and are conserved in various genebanks worldwide. These resources have donated an assortment of alleles needed for resistance/tolerance to various environmental stresses, however, they have yet to be used sufficiently in breeding programs. Our review has highlighted the potential of wild kinds of wheat to use in future breeding programs. We believe that this paper provides useful information and improves the understanding of natural diversity among plant genetic resources and biotechnological tools for graduate students and also for the practical applicability of the researchers.
Author Contributions
A.P.-A. conceived the idea; A.P.-A. and F.K. wrote the manuscript; A.P.-A. and H.M. prepared the figures; A.P.-A., F.K. and H.M. collected information and prepared the table. A.P.-A. and P.P. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Open access funding provided by University of Helsinki.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Skendzic, S.; Zovko, M.; Zivkovic, I.P.; Lesic, V.; Lemic, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef]
- Bakala, H.S.; Mandahal, K.S.; Sarao, L.K.; Srivastava, P. Breeding wheat for biotic stress resistance: Achievements, challenges and prospects. In Current Trends Wheat Research; IntechOpen: London, UK, 2021. [Google Scholar]
- Nevo, E.; Chen, G. Drought and salt tolerances in wild relatives for wheat and barley improvement. Plant Cell Environ. 2010, 33, 670–685. [Google Scholar] [CrossRef]
- Redden, R. New Approaches for crop genetic adaptation to the abiotic stresses predicted with climate change. Agronomy 2013, 3, 419–432. [Google Scholar] [CrossRef]
- Glaszmann, J.C.; Kilian, B.; Upadhyaya, H.D.; Varshney, R.K. Accessing genetic diversity for crop improvement. Curr. Opin. Plant. Biol. 2010, 13, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Von Botmer, R.; Seberg, O.; Jacobsen, N. Genetic resources in the Triticeae. Hered. 1992, 116, 141–150. [Google Scholar] [CrossRef]
- Faris, J.D.; Zhang, Q.; Chao, S.; Zhang, Z.; Xu, S. Analysis of agronomic and domestication traits in a durum × cultivated emmer wheat population using a high-density single nucleotide polymorphism-based linkage map. Theor. Appl. Genet. 2014, 127, 2333–2348. [Google Scholar] [CrossRef] [PubMed]
- Weide, A.; Rieh, S.; Zeidi, M.; Conard, N.J. Using new morphological criteria to identify domesticated emmer wheat at the aceramic Neolithic site of Chogha Golan (Iran). J. Archaeol. Sci. 2013, 57, 109–118. [Google Scholar] [CrossRef]
- Kimber, G.; Sears, E.R. Evolution in the genus Triticum and the origin of cultivated wheat. In Wheat and Wheat Improvement; Heyne, E.G., Ed.; American Society of Agronomy: Madison, WI, USA, 1987. [Google Scholar]
- Kimber, G.; Tsunewaki, K. Genome symbols and plasma types in the wheat group. In Proceedings of the 7th International Wheat Genetic Symposium, Cambridge, UK, 13–19 July 1988. [Google Scholar]
- Gupta, P.K. Cytogenetics of wheat and its close wild relatives—Triticum and Aegilops. In Chromosome Engineering in Plants: Genetics, Breeding, Evolution; Gupta, P.K., Tsuchiya, T., Eds.; Elsevier: Amsterdam, The Netherlands, 1991. [Google Scholar]
- Wang, R.R.C.; van Bothmer, R.; Dvorak, R.; Fedak, G.; Linde-Laursen, I.; Muramatsu, M. Genome symbols in the Triticeae (Poaceae). In Proceedings of the 2nd International Triticeae Symposium, Logan, UT, USA, 20–24 June 1994. [Google Scholar]
- Petersen, G.; Seberg, O.; Yde, M.; Berthelsen, K. Phylogenetic relationships of Triticum and Aegilops and evidence for the origin of the A, B, and D genomes of common wheat (Triticum aestivum). Mol. Phylogenet. Evol. 2006, 39, 70–82. [Google Scholar] [CrossRef]
- Eckardt, N.A. Evolution of domesticated bread wheat. Plant Cell 2010, 22, 993. [Google Scholar] [CrossRef] [PubMed]
- Charmet, G. Wheat domestication: Lessons for the future. CR Biol. 2011, 334, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Tanno, K.; Willcox, G. Distinguishing wild and domestic wheat and barley spikelets from early Holocene sites in the Near East. Veget. Hist. Archaeobot. 2011, 21, 107–115. [Google Scholar] [CrossRef]
- Riehl, S.; Zeidi, M.; Conard, N.J. Emergence of agriculture in the foothills of the Zagros Mountains of Iran. Science 2013, 341, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Weide, A. On the identification of domesticated emmer wheat, Triticum turgidum subsp. dicoccum (Poaceae), in the aceramic neolithic of the Fertile Crescent. Arch. Inf. 2015, 38, 381–424. [Google Scholar]
- Edet, O.U.; Gorafi, Y.S.; Nasuda, S.; Tsujimoto, H. DArTseq-based analysis of genomic relationships among species of tribe Triticeae. Sci. Rep. 2018, 8, 16397. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.; Schreiber, M.; Mascher, M. Domestication and crop evolution of wheat and barley: Egnes, genomics, and future directions. J. Integr. Plant Biol. 2019, 61, 204–225. [Google Scholar] [CrossRef] [Green Version]
- Venske, E.; Santos, R.S.; Busanello, C.; Gustafson, P.; Oliveira, A.C. Bread wheat: A role model for plant domestication and breeding. Hereditas 2019, 156l, 16. [Google Scholar] [CrossRef] [Green Version]
- DeHaan, L.; Larson, S.; López-Marqués, R.L.; Wenkel, S.; Gao, C.; Palmgren, M. Roadmap for accelerated domestication of an emerging perennial grain crop. Trends Plant Sci. 2020, 25, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.; Islam, S.; Yu, Z.; She, M.; Nevo, E.; Ma, W. Current progress in understanding and recovering the wheat genes lost in evolution and domestication. Int. J. Mol. Sci. 2020, 21, 5836. [Google Scholar] [CrossRef]
- Kilian, B.; Ozkan, H.; Pozzi, C.; Salamini, F. Genetics and genomics of the Triticeae. In Plant Genetics and Genomics: Crops and Models 7; Springer: Berlin, Germany, 2009. [Google Scholar]
- Peng, J.H.; Sun, D.; Nevo, E. Domestication evolution, genetics and genomics in wheat. Mol. Breed. 2011, 28, 281–301. [Google Scholar] [CrossRef]
- Lilienfeld, F.A. Genome-analysis in Triticum and Aegilops. Concluding review. Cytologia 1951, 16, 101–123. [Google Scholar] [CrossRef] [Green Version]
- Kimber, G.; Sears, E.R. Assignment of genome symbols in the Triticeae. In Proceedings of the 6th International Wheat Genetics Symposium, Kyoto, Japan, 28 November–3 December 1951. [Google Scholar]
- Percival, J. The Wheat Plant; Duckworth: London, UK, 1921. [Google Scholar]
- Jaradat, A.A. Phenotypic divergence for morphological and yield-related traits among landrace genotypes of durum wheat from Jordan. Euphytica 1991, 52, 155–164. [Google Scholar] [CrossRef]
- Anker, C.C.; Niks, R.E. Prehaustorial resistance to the wheat leaf rust fungus, Puccinia triticina, in Triticum monococcum (s.s.). Euphytica 2001, 117, 209–215. [Google Scholar] [CrossRef]
- Arzani, A.; Khalighi, M.R.; Shiran, B.; Kharazian, N. Evaluation of diversity in wild relatives of wheat. Czech J. Genet. Plant Breed. 2005, 41, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Aghaei, M.J.; Mozafari, J.; Taleei, A.R.; Naghavi, M.R.; Omidi, M. Distribution and diversity of Aegilops tauschii in Iran. Genet. Resour. Crop Evol. 2008, 55, 341. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.A.; Moghaddam, M.; Etminan, A. Evaluation of agro-morphological diversity in wild relatives of wheat collected in Iran. J. Agric. Sci. Technol. 2017, 19, 943–956. [Google Scholar]
- Karagöz, A.; Pilanali, N.; Polat, T. Agro-morphological characterization of some wild wheat (Aegilops L. and Triticum L.) species. Turk. J. Agric. For. 2007, 30, 387–398. [Google Scholar]
- Nazari, M.; Moosavi, S.S.; Maleki, M. Morpho-physiological and proteomic responses of Aegilops tauschii to imposed moisture stress. Plant Physiol. Biochem. 2018, 132, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Moosavi, S.S.; Nazari, M.; Maleki, M. Responses of above and below-ground traits of wheat wild relative (Aegilops tauschii) and bread wheat (Triticum aestivum L.) to imposed moisture stress. Desert 2017, 22, 209–220. [Google Scholar]
- Suneja, Y.; Gupta, A.K.; Bains, N.S. Stress adaptive plasticity: Aegilops tauschii and Triticum dicoccoides as Potential donors of drought associated morpho-physiological traits in wheat. Front. Plant Sci. 2019, 10, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pour-Aboughadareh, A.; Mahmoudi, M.; Moghaddam, M.; Ahmadi, J.; Mehrabi, A.A.; Alavikia, S.S. Agro-morphological and molecular variability in Triticum boeoticum accessions from Zagros Mountains, Iran. Genet. Resour. Crop Evol. 2017, 64, 545–556. [Google Scholar] [CrossRef]
- Moosavi, S.S.; Abdi, F.; Abdollahi, M.R.; Tahmasebi-Enferadi, S.; Maleki, M. Phenological, morpho-physiological and proteomic responses of Triticum boeoticum to drought stress. Plant Physiol. Biochem. 2020, 156, 95–104. [Google Scholar] [CrossRef]
- Kianersi, F.; Mousavi, S.S.; Afiuni, D.; Abdollahi, M. Genetic diversity and heritability of some suitable morpho-physiological traits in bread wheat (Triticum aestivum L.) under terminal moisture stress. Adv. Crop Sci. 2013, 3, 376–388. [Google Scholar]
- Moosavi, S.S.; Kian Ersi, F.; Abdollahi, M.R. Application of multivariate statistical methods in detection of effective traits on bread wheat (Triticum aestivum L.) yield under drought stress condition. Cereal Res. 2013, 3, 119–130. [Google Scholar]
- Naghavi, M.R.; Toorchi, M.; Moghaddam, M.; Shakiba, M.R. Evaluation of diversity and traits correlation in spring wheat cultivars under drought stress. Not. Sci. Biol. 2015, 7, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Adel Ahmed, E.; Abd El-Aziz Afiah, S.; Abdulaziz Al-Doss, A.; Ibrahim, E.I. Morphological variability and genetic diversity of wheat genotypes grown on saline soil and identification of new promising molecular markers associated with salinity tolerance. J. Plant Interact. 2019, 14, 564–571. [Google Scholar]
- Tshikunde, N.M.; Mashilo, J.; Shimelis, H.; Odindo, A. Agronomic and physiological traits, and associated quantitative trait loci (QTL) affecting yield response in wheat (Triticum aestivum L.): A review. Front. Plant Sci. 2019, 10, 1428. [Google Scholar] [CrossRef] [Green Version]
- Rehman, H.U.; Tariq, A.; Ashraf, I.; Ahmed, M.; Muscolo, A.; Basra, S.M.A.; Reynolds, M. Evaluation of physiological and morphological traits for improving spring wheat adaptation to terminal heat stress. Plants 2021, 10, 455. [Google Scholar] [CrossRef]
- Peleg, Z.; Fahima, T.; Krugman, T.; Abbo, S.; Yakir, D.; Korol, A.B.; Saranga, Y. Genomic dissection of drought resistance in durum wheat x wild emmer wheat recombinant inbreed line population. Plant Cell Environ. 2009, 32, 758–779. [Google Scholar] [CrossRef] [PubMed]
- Peleg, Z.; Fahima, T.; Korol, A.B.; Abbo, S.; Saranga, Y. Genetic analysis of wheat domestication and evolution under domestication. J. Exp. Bot. 2011, 62, 5051–5061. [Google Scholar] [CrossRef] [Green Version]
- Fatiukha, A.; Deblieck, M.; Klymiuk, V.; Merchuk-Ovnat, L.; Peleg, Z.; Ordon, F.; Fahima, T.; Korol, A.; Saranga, Y.; Krugman, T. Genomic architecture of phenotypic plasticity in response to water stress in tetraploid wheat. Int. J. Mol. Sci. 2021, 22, 1723. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Mohammadi, R.; Etminan, A.; Shooshtari, L.; Maleki-Tabrizi, N.; Poczai, P. Effects of drought stress on some agronomic and morpho-physiological traits in durum wheat genotypes. Sustainability 2020, 12, 5610. [Google Scholar] [CrossRef]
- Huang, L.; Feng, L.; He, Y.; Tang, Z.; He, J.; Sela, H.; Krugman, T.; Fahima, T.; Liu, D.; Wu, B. Variation in stripe rust resistance and morphological traits in wild emmer wheat populations. Agronomy 2019, 9, 44. [Google Scholar] [CrossRef] [Green Version]
- Pour-Aboughadareh, A.; Alavikia, S.S.; Moghaddam, M.; Mehrabi, A.; Mazinani, M.A. Evaluation of morpho–physiological traits in some Triticum urartu populations from Iran under normal and water deficit stress conditions. J. Agric. 2013, 15, 135–148. [Google Scholar]
- Gurcan, K.; Demirel, F.; Tekin, M.; Demirel, S.; Akar, T. Molecular and agro-morphological characterization of ancient wheat landraces of turkey. BMC Plant Biol. 2017, 17, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, K.; Knüpffer, H. Genetic resources of Triticum. In Advances in Wheat Genetics: From Genome to Field; Springer: Tokyo, Japan, 2015. [Google Scholar]
- Brush, S.B. Genes in the Field: On-Farm Conservation of Crop Diversity; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Padulosi, S.; Bergamini, N.; Lawrence, T. On-farm conservation of neglected and underutilized species: Status, trends and novel approaches to cope with climate change. In Proceedings of the International Conference, Frankfurt, Germany, 14–16 June 2011. [Google Scholar]
- Teklu, Y.; Hammer, K. Farmers’ perception and genetic erosion of tetraploid wheat landraces in Ethiopia. Genet. Resour. Crop Evol. 2006, 53, 1099–1113. [Google Scholar] [CrossRef]
- Stubbe, H. Spontane und Strahleninduzierte Mutabilität; Georg Thieme Verlag: Stuttgart, Germany, 1937. [Google Scholar]
- Skovmand, B.; Reynolds, M.P.; DeLacy, I.H. Searching genetic resources for physiological traits with potential for increasing yield. In Application of Physiology in Wheat Breeding; CIMMYT: Mexico City, Mexico, 2001; pp. 17–28. [Google Scholar]
- Goriunova, S.V.; Chikida, N.N.; Kochieva, E.Z. RAPD analysis of the intraspecific and interspecific variation and phylogenetic relationships of Aegilops L. species with the U genome. Genetika 2010, 46, 945–959. [Google Scholar] [CrossRef]
- Okuno, K.; Ebana, K.; Noov, B.; Yoshida, H. Genetic diversity of Central Asian and north Caucasian Aegilops species as revealed by RAPD markers. Genet. Resour. Crop Evol. 1998, 45, 389–394. [Google Scholar] [CrossRef]
- Mguis, K.; Mahjoub, A.; Abassi, M.; Albouchi, A.; Ouerghi, Z.; Nadia, B.B.; Bejaoui, Z. Morphological and genetic variation in Aegilops geniculata Roth. from Tunisia. Int. J. Agric. Res. 2015, 6, 8–21. [Google Scholar]
- Mahjoub, A.; El, M.G.; Mguis, K.; El, M.G.; Brahim, N. Evaluation of genetic diversity in Aegilops geniculata Roth accessions using morphological and RAPD markers. Pak. J. Biol. Sci. 2009, 12, 994–1003. [Google Scholar] [CrossRef]
- Guadagnuolo, R.; Savova-Bianchi, D.; Felber, F. Gene flow from wheat (Triticum aestivum L.) to jointed goatgrass (Aegilops cylindrica Host.), as revealed by RAPD and microsatellite markers. Theor. Appl. Genet. 2001, 103, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mallabaeva, D.S.; Ignatov, A.N.; Sheiko, I.A.; Isikov, V.P.; Gelyuta, V.P.; Boiko, N.G.; Seryapin, A.A.; Dorokhov, D.B. Use of RAPD and ITE molecular markers in studying the genetic structure of the Crimean population of T. boeoticum Boiss. Cytol. Genet. 2007, 41, 181–189. [Google Scholar] [CrossRef]
- Zaharieva, M.; Santoni, S.; David, J. Use of RFLP markers to study genetic diversity and to build a core-collection of the wild wheat relative Ae. geniculata Roth (=Ae. ovata L.). Genet. Sel. Evol. 2001, 33, 269–288. [Google Scholar] [CrossRef] [Green Version]
- Naghavi, M.R.; Mardi, M.; Pirseyedi, S.M.; Kazemi, M.; Potki, P.; Gaffari, M.R. Comparison of genetic variation among accessions of Aegilops tauschii using AFLP and SSR markers. Genet. Resour. Crop Evol. 2007, 54, 237–240. [Google Scholar] [CrossRef]
- Monte, J.V.; De Nova, P.J.G.; Soler, C. AFLP-based analysis to study genetic variability and relationships in the Spanish species of the genus Aegilops. Hereditas 2001, 135, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Kaya, I.; Kirisozu, A.C.; Ersoy, F.Y.; Dere, S.; Akkaya, M.S. Genetic diversity and relationship analysis among accessions of Aegilops ssp. in Turkey using amplified fragment length polymorphism (AFLP) markers. Afr. J. Biotechnol. 2011, 72, 16167–16174. [Google Scholar]
- Goryunova, S.V.; Chikida, N.N.; Kochieva, E.Z. AFLP, RAPD, and ISSR analysis of intraspecific polymorphism and interspecific differences of allotetraploid species Aegilops kotschyi Boiss. and Aegilops variabilis Eig. Russ. J. Genet. 2017, 53, 568–575. [Google Scholar] [CrossRef]
- Medini, M.; Hamza, S.; Rebai, A.; Baum, M. Analysis of genetic diversity in Tunisian durum wheat cultivars and related wild species by SSR and AFLP markers. Genet. Resour. Crop Evol. 2005, 52, 21–31. [Google Scholar] [CrossRef]
- Eslami Farouji, A.; Khodayari, H.; Saeidi, H.; Rahiminejad, R. Genetic diversity of diploid Triticum species in Iran assessed using inter-retroelement amplified polymorphisms (IRAP) markers. Biologia 2015, 70, 52–60. [Google Scholar] [CrossRef]
- Taheri, M.T.; Alavi-Kia, S.S.; Mohammadi, S.A.; Moghaddam Vahed, M. Assessment of genetic diversity and relationships among Triticum urartu and Triticum boeoticum populations from Iran using IRAP and REMAP markers. Genet. Resour. Crop Evol. 2018, 65, 1867–1878. [Google Scholar] [CrossRef]
- Ahmadi, J.; Ourang, S.; Pour-Aboughadareh, A. Evaluation of genetic diversity in Aegilops populations possessing D genome using SCoT and TRAP markers. Mod. Genet. J. 2019, 14, 219–228. [Google Scholar]
- Baranduzi, A.; Sofalian, O.; Asghari Zakaria, R.; Asghari, A.; Skokroop, M. Assessment of genetic diversity in Aegilops species in North-West of Iran using ISSR marker. YYU J. Agric. Sci. 2013, 23, 66–75. [Google Scholar]
- Prazak, R.; Paczos, E. Genetic similarity among Aegilops Kotschyi Boiss. with Triticum aestivum L. hybrid lines. Rom. Agric. Res. 2017, 34, 15–23. [Google Scholar]
- Bouziani, M.; Bechkri, S.; Bellil, I.; Khelifi, D. Evaluation of genetic diversity of Algerian Aegilops ventricosa Tausch. using intersimple sequence repeat (ISSR) markers. World J. Environ. Biosci. 2014, 8, 1–6. [Google Scholar]
- Lelley, T.; Stachel, M.; Grausgruber, H.; Vollmann, J. Analysis of relationships between Aegilops tauschii and the D genome of wheat utilizing microsatellites. Genome 2000, 43, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fahima, T.; Peng, J.; Roder, M.S.; Kirzhner, V.M.; Korol, A.B.; Nevo, E. Edaphic microsatellite DNA divergence in wild emmer wheat, Triticum dicoccoides, at a microsite: Tabigha, Israel. Theor. Appl. Genet. 2000, 101, 1029–1038. [Google Scholar] [CrossRef]
- Aghaee-Sarbarzeh, M.; Singh, H.; Dhaliwal, H.S. A microsatellite marker linked to leaf rust resistance transferred from Aegilops triuncialis into hexaploid wheat. Plant Breed. 2001, 120, 259–261. [Google Scholar] [CrossRef]
- Arabbeigi, M.; Arzani, A.; Majidi, M.M.; Kiani, R.; Tabatabaei, B.E.S.; Habibi, F. Salinity tolerance of Aegilops cylindrica genotypes collected from hypersaline shores of Uremia Salt Lake using physiological traits and SSR markers. Acta Physiol. Plant. 2014, 36, 2243–2251. [Google Scholar] [CrossRef]
- Abbasov, M.; Brueggeman, R.; Raupp, J. Genetic diversity of Aegilops L. species from Azerbaijan and Georgia using SSR markers. Genet. Resour. Crop Evol. 2019, 66, 453–463. [Google Scholar] [CrossRef]
- Schneider, A.; Molnar, I.; Molnar-Lang, M. Production and FISH identification of wheat-Aegilops biuncialis addition lines and their use for the selection of U and M genome-specific molecular (SSR) markers. Acta Agron. Hung. 2010, 58, 151–158. [Google Scholar] [CrossRef]
- Saeidi, H.; Rahiminejad, M.; Vallian, S.; Heslop-Harrison, J.S. Biodiversity of diploid D-genome Aegilops tauschii Coss. in Iran measured using microsatellites. Genet. Resour. Crop Evol. 2006, 53, 1477–1484. [Google Scholar] [CrossRef]
- Dudnikov, A.J.; Kawahara, T. Aegilops tauschii: Genetic variation in Iran. Genet. Resour. Crop Evol. 2006, 53, 579–586. [Google Scholar] [CrossRef]
- Li, W.; Zhang, D.F.; Wei, Y.M.; Yan, Z.H.; Zheng, Y.L. Genetic diversity of Triticum turgidum L. based on microsatellite markers. Russ. J. Genet. 2006, 42, 311–316. [Google Scholar] [CrossRef]
- Moradkhani, H.; Mehrabi, A.A.; Etminan, A.; Pour-Aboughadareh, A. Molecular diversity and phylogeny of Triticum-Aegilops species possessing D genome revealed by SSR and ISSR markers. Plant Breed. Seed Sci. 2015, 71, 81–95. [Google Scholar] [CrossRef]
- Yamane, K.; Kawahara, T. Size homoplasy and mutational behavior of chloroplast simple sequence repeats (cpSSRs) inferred from intra- and interspecific variations in four chloroplast regions of diploid and polyploid Triticum and Aegilops species. Genet. Resour. Crop Evol. 2018, 65, 727–743. [Google Scholar] [CrossRef]
- Gandhi, H.T.; Vales, M.I.; Mallory-Smith, C.; Riera-Lizarazu, O. Genetic structure of Aegilops cylindrica Host in its native range and in the United States of America. Theor. Appl. Genet. 2009, 119, 1013–1025. [Google Scholar] [CrossRef]
- Gong, W.; Han, R.; Li, H. Agronomic traits and molecular marker identification of wheat-Aegilops caudata addition lines. Front. Plant Sci. 2017, 8, 1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urazaliev, R.; Yessimbekova, M.; Mukin, K.; Chirkin, A.; Ismagulova, G. Monitoring of Aegilops L local species genetic diversity of Kazakhstans flora Vavilovskii Zhurnal Genetikii Selektsii Vavilov. J. Genet. Breed. 2018, 22, 484–490. [Google Scholar]
- Yasui, Y.; Nasuda, S.; Matsuoka, Y.; Kawahara, T. The Au family, a novel short interspersed element (SINE) from Aegilops umbellulata. Theor. Appl. Genet. 2001, 102, 463–470. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.A.; Etminan, A.; Moghaddam, M. Assessment of genetic diversity among Iranian Triticum germplasm using agro-morphological traits and start codon targeted (SCoT) markers. Cereal Res. Commun. 2017, 45, 574–586. [Google Scholar] [CrossRef] [Green Version]
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.A.; Etminan, A.; Moghaddam, M. Insight into the genetic variability analysis and relationships among some Aegilops and Triticum species, as genome progenitors of bread wheat, using SCoT markers. Plant Biosyst. 2018, 152, 694–703. [Google Scholar] [CrossRef]
- Etminan, A.; Pour-Aboughadareh, A.; Mehrabi, A.A.; Shooshtari, L.; Ahmadi-Rad, A.; Moradkhani, H. Molecular characterization of the wild relatives of wheat using CAAT-box derived polymorphism. Plant Biosyst. 2019, 153, 398–405. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Etminan, A.; Shooshtari, L.; Maleki-Tabrizi, N. Comparative assessment of SCoT and CBDP markers for investigation of genetic diversity existing in different Aegilops Species. Agric. Biotechnol. J. 2019, 4, 153–174. [Google Scholar]
- Ivanizs, L.; Monostori, I.; Farkas, A. Unlocking the genetic diversity and population structure of a wild gene source of wheat, Aegilops biuncialis Vis., and its relationship with the heading time. Front. Plant Sci. 2019, 10, 1531. [Google Scholar] [CrossRef] [Green Version]
- Abbasov, M.; Sansaloni, C.P.; Burgueno, J. Genetic diversity analysis using DArTseq and SNP markers in populations of Aegilops species from Azerbaijan. Genet. Resour. Crop Evol. 2020, 67, 281–291. [Google Scholar] [CrossRef]
- Laido, G.; Mangini, G.; Taranto, F. Genetic diversity and population structure of tetraploid wheats (Triticum turgidum L.) estimated by SSR, DArT and pedigree data. PLoS ONE 2013, 8, e67280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, B.S. SNPing Aegilops tauschii genetic diversity and the birthplace of bread wheat. New Phytol. 2013, 198, 641–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, S.; Cheema, J.; Poland, J.; Uauy, C.; Chhuneja, P. Genome-wide association mapping of grain micronutrients concentration in Aegilops tauschii. Front. Plant Sci. 2019, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Zou, M.; Zhu, Y.; Han, X.; Li, Y.; Zhang, D.; Li, S. Analysis of population structure and origin in Aegilops tauschii Coss. from China through SNP markers. Genet. Resour. Crop Evol. 2020, 67, 923–934. [Google Scholar] [CrossRef]
- Szabo-Hever, A.; Zhang, Q.; Friesen, T.L.; Zhong, S.; Elias, E.M.; Cai, X.; Jin, Y.; Faris, J.D.; Chao, S.; Xu, S.S. Genetic diversity and resistance to Fusarium head blight in synthetic hexaploid wheat derived from Aegilops tauschii and diverse Triticum turgidum subspecies. Front. Plant Sci. 2018, 9, 1829. [Google Scholar] [CrossRef]
- Govindaraj, M.; Vetriventhan, M.; Srinivasan, M. Importance of genetic diversity assessment in crop plants and its recent advances: An overview of its analytical perspectives. Genet. Res. Int. 2015, 2015, 431487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Mittal, N.; Leamy, L.; Barazani, O.; Song, B.H. Back into the wild—Apply untapped genetic diversity of wild relatives for crop improvement. Evol. Appl. 2016, 10, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Budak, H.; Kantar, M.; Yucebilgili Kurtoglu, K. Drought tolerance in modern and wild wheat. Sci. World J. 2013, 2013, 548246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaharieva, M.; Gaulin, E.; Havaux, M.; Acevedo, E.; Monneveux, P. Drought and heat responses in the wild wheat relative Aegilops geniculata Roth: Potential interest for wheat improvement. Crop Sci. 2001, 41, 1321–1329. [Google Scholar] [CrossRef]
- Baalbaki, R.; Hajj-Hassan, N.; Zurayk, R. Aegilops species from semi-arid areas of Lebanon: Variation in quantitative attributes under water stress. Crop Sci. 2006, 46, 799–806. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Dreccer, F.; Trethowan, R. Drought adaptive traits derived from wheat wild relatives and landraces. J. Exp. Bot. 2007, 58, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Khazaei, H.; Monneveux, P.; Hongbo, S.; Mohammady, S. Variation for stomatal characteristics and water use efficiency among diploid, tetraploid and hexaploid Iranian wheat landraces. Genet. Resour. Crop Evol. 2010, 57, 307–314. [Google Scholar] [CrossRef]
- Waines, J.G.; Rafi, M.M.; Ehdaie, B. Yield Components and transpiration efficiency in wild wheats. In Biodiversity and Wheat Improvement; Damanian, A.B., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 1993. [Google Scholar]
- Budak, H.; Hussain, B.; Khan, Z.; Ozturk, N.Z.; Ullah, N. From genetics to functional genomics: Improvement in drought signaling and tolerance in wheat. Front. Plant Sci. 2015, 6, 1012. [Google Scholar] [CrossRef] [Green Version]
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.A.; Etminan, A.; Moghaddam, M.; Siddique, K.H.M. Physiological responses to drought stress in wild relatives of wheat: Implications for wheat improvement. Acta Physiol. Plant. 2017, 39, 106. [Google Scholar] [CrossRef]
- Hossain, M.S.; Dietz, K.-J. Tuning of redox regulatory mechanisms, reactive oxygen species and redox homeostasis under salinity stress. Front. Plant Sci. 2016, 7, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, J.; Rodrigo-Moreno, A.; Shabala, S. ROS homeostasis in halophytes in the context salinity stress tolerance. J. Exp. Bot. 2013, 65, 1241–1257. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, J.; Pour-Aboughadareh, A.; Ourang, S.F.; Mehrabi, A.A.; Siddique, K.H.M. Wild relatives of wheat: Aegilops–Triticum accessions disclose differential antioxidative and physiological responses to water stress. Acta Physiol. Plant. 2018, 40, 90. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Omidi, M.; Naghavi, M.R.; Etminan, A.; Mehrabi, A.A.; Poczai, P.; Bayat, H. Effect of water deficit stress on seedling biomass and physio-chemical characteristics in different species of wheat possessing the D genome. Agronomy 2019, 9, 522. [Google Scholar] [CrossRef] [Green Version]
- Pour-Aboughadareh, A.; Omidi, M.; Naghavi, M.R.; Etminan, A.; Mehrabi, A.A.; Poczai, P. Wild relatives of wheat respond well to water deficit stress: A comparative study of antioxidant enzyme activities and their encoding gene expression. Agriculture 2020, 10, 415. [Google Scholar] [CrossRef]
- Trachsel, S.; Stamp, P.; Hund, A. Growth of axile and lateral roots of maize: Response to desiccation stress induced by polyethylene glycol 8000. Maydica 2010, 55, 101–109. [Google Scholar]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki Ourang, S.; Mehrabi, A.A.; Siddique, K.H.M. Screening wheat germplasm for seedling root architectural traits under contrasting water regimes: Potential sources of variability for drought adaptation. Arch. Agron. Soil Sci. 2018, 64, 1351–1365. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Prasad, P.V.V.; Kumari, J.; Sehgal, S.K.; Friebe, B.; Djalovic, I.; Chen, Y.; Siddique, K.H.M.; Gill, B.S. Alien chromosome segment from Aegilops speltoides and Dasypyrum villosum increases drought tolerance in wheat via profuse and deep root system. BMC Plant Biol. 2019, 19, 242. [Google Scholar] [CrossRef] [Green Version]
- Pour-Aboughadareh, A.; Mehrvar, M.R.; Sanjani, S.; Amini, A.; Nikkhah-Chamanabad, H.; Asadi, A. Effects of salinity stress on seedling biomass, physiochemical properties, and grain yield in different breeding wheat genotypes. Acta Physiol. Plant. 2021, 43, 98. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Sanjani, S.; Nikkhah-Chamanabad, H.; Mehrvar, M.R.; Asadi, A.; Amini, A. Identification of salt-tolerant barley genotypes using multiple-traits index and yield performance at the early growth and maturity stages. Bull. Natl. Res. Cent. 2021, 45, 117. [Google Scholar] [CrossRef]
- Arzani, A. Improving salinity tolerance in crop plants: A biotechnological view. Cell. Dev. Biol. Plant 2008, 44, 373–383. [Google Scholar] [CrossRef]
- Colmer, T.D.; Flowers, T.J.; Munns, R. Use of wild relatives to improve salt tolerance in wheat. J. Exp. Bot. 2006, 57, 1059–1078. [Google Scholar] [CrossRef] [Green Version]
- Arzani, A.; Ashraf, A. Smart engineering of genetic resources for enhanced salinity tolerance in crop plants. Crit. Rev. Plant Sci. 2016, 35, 146–189. [Google Scholar] [CrossRef]
- Rezaei, M.; Arzani, A.; Saeidi, G.; Karami, M. Physiology of salinity tolerance in Bromus danthoniae genotypes originated from saline and non-saline areas of West Iran. Crop Pasture Sci. 2017, 68, 92–99. [Google Scholar] [CrossRef]
- Farooq, S.; Niazi, M.L.K.; Iqbal, N.; Shah, T.M. Salt tolerance potential of wild resources of the tribe Triticeae. II. Screening of species of genus Aegilops. Plant Soil. 1989, 119, 255–260. [Google Scholar] [CrossRef]
- Gorham, J.; Hardy, C.; Jones, R.G.W.; Joppa, L.R.; Law, C.N. Chromosomal location of a K+/Na+ discrimination character in the D genome of wheat. Theor. Appl. Genet. 1987, 74, 584–588. [Google Scholar] [CrossRef]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki Ourang, S.; Khalili, P.; Poczai, P. Unraveling salinity stress responses in ancestral and neglected wheat species at early growth stage: A baseline for utilization in future wheat improvement programs. Physiol. Mol. Biol. Plants 2020, 26, 537–549. [Google Scholar] [CrossRef]
- Kiani, R.; Arzani, A.; Habibi, F. Physiology of salinity tolerance in Aegilops ylindrica. Acta Physiol. Plant. 2015, 37, 135–145. [Google Scholar] [CrossRef]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki-Ourang, S.; Mehrabi, A.A.; Siddique, K.H.M. Screening wild progenitors of wheat for salinity stress at early stages of plant growth: Insight into potential sources of variability for salinity adaptation in wheat. Crop Pasture Sci. 2018, 69, 649–658. [Google Scholar] [CrossRef]
- Prasad, P.; Staggenborg, S.; Ristic, Z. Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. In Modeling Water Stress Effects on Plant Growth Processes; Ahuja, L., Saseendran, S., Eds.; CROP Science Society of America: Madison, WI, USA, 2008. [Google Scholar]
- Ristic, Z.; Bukovnik, U.; Prasad, P.V.V. Correlation between heat stability of thylakoid membranes and loss of chlorophyll in winter wheat under heat stress. Crop Sci. 2007, 47, 2067–2073. [Google Scholar] [CrossRef]
- Al-Khatib, K.; Paulsen, G.M. Mode of high temperature injury to wheat during grain development. Physiol. Plant. 1984, 61, 363–368. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.J.; Landolt, W.; Bucher, J.B.; Strasser, R.J. Beech (Fagus sylvatica) response to ozone exposure assessed with a chlorophyll fluorescence performance index. Environ. Pollut. 2000, 109, 501–507. [Google Scholar] [CrossRef]
- Percival, G.C.; Sheriffs, C.N. Identification of drought-tolerance woody perennials using chlorophyll fluorescence. Arboric. Urban For. 2002, 28, 215–223. [Google Scholar]
- Li, R.H.; Pei-guo, G.; Baum, M.; Grando, S.; Cecccarelli, S. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Li, Y.; Song, H.; Xu, Z.; Zhou, G. Tracking chlorophyll fluorescence as an indicator of drought and rewatering across the entire leaf lifespan in a maize field. Agric. Water Manag. 2019, 211, 190–201. [Google Scholar] [CrossRef]
- Hairat, S.; Khurana, P. Evaluation of Aegilops tauschii and Aegilops speltoides for acquired thermotolerance: Implications in wheat breeding programmes. Plant Physiol. Biochem. 2015, 95, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, G.P.; Prasad, P.V.V.; Fritz, A.K.; Kirkhan, M.B.; Gill, B.S. Response of Aegilops species to drought stress during reproductive stage of development. Funct. Plant Biol. 2012, 39, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limin, A.E.; Fowler, D.B. Cold hardiness of some wild relatives of hexaploid wheat. Botany 1981, 59, 572–573. [Google Scholar]
- Barashkova, E.A.; Vavilov, N. Physiological-genetic aspects of frost resistance in winter wheat. Relationship of frost hardiness with genome composition in wheat. In Proceedings of International Symposium: Wheat Breeding—Prospects and Future Approaches; Institute for Wheat and Sunflower: Albena, Bulgaria, 1991; pp. 379–384. [Google Scholar]
- Ehdaie, B.; Waines, J.G. Heat resistance in wild Triticum and Aegilops. J. Genet. Breed. 1992, 46, 221–228. [Google Scholar]
- Stankova, P.; Rekika, D.; Zaharieva, M.; Monneveux, P. Improvement of durum wheat for multiple stress tolerance: Potential interest of Aegilops sp. In Fibre and Cereal Crops Problems; Cotton and Durum Wheat Research Institute: Chirpan, Bulgaria, 1995. [Google Scholar]
- Masoomi-Aladizgeh, F.; Aalami, A.; Esfahani, M.; Aghaei, M.J.; Mozaffari, K. Identification of CBF14 and NAC2 genes in Aegilops tauschii associated with resistance to freezing stress. Appl. Biochem. Biotechnol. 2015, 176, 1059–1070. [Google Scholar] [CrossRef]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of plant diseases—A field perspective. Mol. Plant Path. 2017, 19, 1523–1536. [Google Scholar] [CrossRef]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Bertholdsson, N.O. Early vigor and allelopathy-Two useful traits for enhancing barley and wheat competitiveness against weeds. Weed Res. 2005, 45, 94–102. [Google Scholar] [CrossRef]
- Jiang, J.; Friebe, B.; Gill, B.S. Recent advances in alien gene transfer in wheat. Euphytica 1994, 73, 199–212. [Google Scholar] [CrossRef]
- Friebe, B.; Jiang, J.; Raupp, W.J.; McIntosh, R.A.; Gill, B.S. Characterization of wheat-alien translocations conferring resistance to diseases and pests: Current status. Euphytica 1996, 91, 59–87. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Wellings, C.R.; Park, R.F. Wheat Rusts: An Atlas of Resistance Genes; CSIRO Publishing: Melbourne, Australia, 1995. [Google Scholar]
- McIntosh, R.A.; Yamazaki, Y.; Dubcovsky, J.; Rogers, J.; Morris, C.; Appels, R.; Xia, X.C. Catalogue of gene symbols for wheat. In Proceedings of the 12th International Wheat Genetics Symposium, Yokohama, Japan, 8–10 April 2013. [Google Scholar]
- Chhuneja, P.; Kaur, S.; Garg, T.; Ghai, M.; Kaur, S.; Prashar, M.; Bains, N.S.; Goel, R.K.; Keller, B.; Dhaliwal, H.S.; et al. Mapping of adult plant stripe rust resistance genes in diploid a genome wheat species and their transfer to bread wheat. Theor. Appl. Genet. 2008, 116, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Riar, A.K.; Kaur, S.; Dhaliwal, H.S.; Singh, K.; Chhuneja, P. Introgression of a leaf rust resistance gene from Aegilops caudata to bread wheat. J. Genet. 2012, 91, 155–161. [Google Scholar] [CrossRef]
- Kishii, M. An update of recent use of Aegilops species in wheat breeding. Front. Plant Sci. 2019, 10, 585. [Google Scholar] [CrossRef] [Green Version]
- Doussinault, G.; Delibes, A.; Sanchez-Monge, R.; Garcia-Olmedo, F. Transfer of a dominant gene for resistance to eyespot disease from a wild grass to hexaploid wheat. Nature 1983, 303, 698–700. [Google Scholar] [CrossRef]
- McIntosh, R.A. Catalogue of gene symbols for wheat. In Proceedings of the 7th international wheat Genetics Symposium, Cambridge, UK, 13–19 July 1988. [Google Scholar]
- Hsam, S.L.K.; Huang, X.Q.; Ernst, F.; Hartl, L.; Zeller, F.J. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell.). Alleles at the Pm1 locus. Theor. Appl. Genet. 1998, 96, 1129–1134. [Google Scholar] [CrossRef]
- Klindworth, D.L.; Niu, Z.; Chao, S.; Friesen, T.L.; Jin, Y.; Faris, J.D.; Cai, X.; Xu, S. Introgression and characterization of a goatgrass gene for a high level of resistance to ug99 stem rust in tetraploid wheat. G3 2012, 2, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Hsam, S.L.K.; Lapochkina, I.F.; Zeller, F.J. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell.). gene Pm32 in a wheat-Aegilops speltoides translocation line. Euphytica 2003, 133, 367–370. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Miller, T.E.; Chapman, V. Cytogenetical studies in wheat XII. Lr28 for resistance to Puccinia recondita and Sr34 for resistance to P. graminis tritici. Z. Pflanzenzucht. Can. Sci. Publ. 1982, 89, 295–306. [Google Scholar]
- Petersen, S.; Lyerly, J.H.; Worthington, M.L.; Parks, W.R.; Cowger, C.; Marshall, D.S.; Brown-Guedira, G.; Murphy, J.P. Mapping of powdery mildew resistance gene Pm53 introgressed from Aegilops speltoides into soft red winter wheat. Theor. Appl. Genet. 2015, 128, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Bariana, H.S.; McIntosh, R.A. Cytogenetic studies in wheat. XV. location of rust resistance genes in VPM1 and their genetic linkage with other disease resistance genes in chromosome 2A. Genome 1993, 36, 476–482. [Google Scholar] [CrossRef]
- Donini, P.; Koebner, R.M.; Ceoloni, C. Cytogenetic and molecular mapping of the wheat-Aegilops longissima chromatin breakpoints in powdery mildew-resistant introgression lines. Theor. Appl. Genet. 1995, 91, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Jin, Y.; Rouse, M.; Friebe, B.; Gill, B.S.; Pumphrey, M.O. Development and characterization of wheat-Ae. searsii robertsonian translocations and a recombinant chromosome conferring resistance to stem rust. Theor. Appl. Genet. 2011, 122, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Zeller, F.J.; Kong, L.; Hartl, L.; Mohler, V.; Hsam, S.L.K. Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell.). Gene Pm29 in line Pova. Euphytica 2002, 123, 187–194. [Google Scholar] [CrossRef]
- Liu, W.; Rouse, M.; Friebe, B.; Jin, Y.; Gill, B.S.; Pumphrey, M.O. Discovery and molecular mapping of a new gene conferring resistance to stem rust, Sr53, derived from Aegilops geniculata and characterization of spontaneous translocation stocks with reduced alien chromatin. Chromosome Res. 2011, 19, 669–682. [Google Scholar] [CrossRef]
- Liu, W.; Koo, D.H.; Xia, Q.; Li, C.; Bai, F.; Song, Y.; Friebe, B.; Gill, B.S. Homoeologous recombination-based transfer and molecular cytogenetic mapping of powdery mildew-resistant gene Pm57 from Aegilops searsii into wheat. Theor. Appl. Genet. 2017, 130, 841–848. [Google Scholar] [CrossRef]
- Sears, E.R. The transfer of leaf rust resistance from Aegilops umbellulata to wheat. In Brook-Haven Symposia in Biology; Brookhaven National Laboratory, Associated Universities: Champaign, IL, USA, 1956. [Google Scholar]
- Lutz, J.; Hsam, S.L.K.; Limpert, E.; Zeller, F.J. Chromosomal location of powdery mildew resistance genes in Triticum aestivum L. (common wheat) genes Pm2 and Pm19 from Aegilops squarrosa L. Heredity 1995, 74, 152–156. [Google Scholar] [CrossRef]
- Bansal, M.; Kaur, S.; Dhaliwal, H.S.; Baines, N.S.; Bariana, H.S.; Chhuneja, P.; Bansal, U.K. Mapping of Aegilops umbellulata-derived leaf rust and stripe rust loci in wheat. Plant Pathol. 2017, 66, 38–44. [Google Scholar] [CrossRef]
- Miranda, L.M.; Murphy, J.P.; Marshall, D.; Leath, S. Pm34: A new powdery mildew resistance gene transferred from Aegilops tauschii Coss. to common wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 113, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Delibes, A.; Romero, D.; Aguaded, S.; Duce, A.; Mena, M.; Lopez-Brana, I.; Andres, M.F.; Martin-Sanchez, J.A.; Garcia-Olmedo, F. Resistance to the cereal cyst nematode (Heterodera avenae Woll.) transferred from the wild grass Aegilops ventricosa to hexaploid wheat by a “stepping-stone” procedure. Theor. Appl. Genet. 1993, 87, 402–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahier, J.; Tanguy, A.M.; Abelard, P.; Rivoal, R. Utilization of deletions to localize a gene for resistance to the cereal cyst nematode, Heterodera avenae, on an Aegilops ventricosa chromosome. Plant Breed. 1996, 115, 282–284. [Google Scholar] [CrossRef]
- Helguera, M.; Khan, I.A.; Dubcovsky, J. Development of PCR markers for the wheat leaf rust gene Lr47. Theor. Appl. Genet. 2000, 101, 625–631. [Google Scholar] [CrossRef] [Green Version]
- Ogbonnaya, F.C.; Seah, S.; Delibes, A.; Jahier, J.; Lopez-Brana, I.; Eastwood, R.F.; Lagudah, E.S. Molecular-genetic characterisation of a new nematode resistance gene in wheat. Theor. Appl. Genet. 2001, 102, 623–629. [Google Scholar] [CrossRef]
- Helguera, M.; Vanzetti, L.; Soria, M.; Khan, I.A.; Kolmer, J.; Dubcovsky, J. PCR markers for Triticum speltoides leaf rust resistance gene Lr51 and their use to develop isogenic hard red spring wheat lines. Crop Sci. 2005, 45, 728–734. [Google Scholar] [CrossRef] [Green Version]
- Romero, M.D.; Montes, M.J.; Sin, E.; López-Braña, I.; Duce, A.; Martn-Sanchez, J.A. A cereal cyst nematode (Heterodera avenae Woll.) resistance gene transferred from Aegilops triuncialis to hexaploid wheat. Theor. Appl. Genet. 1998, 96, 1135–1140. [Google Scholar] [CrossRef]
- Marais, G.F.; Badenhorst, P.E.; Eksteen, A.; Pretorius, Z.A. Reduction of Aegilops sharonensis chromatin associated with resistance genes Lr56 and Yr38 in wheat. Euphytica 2010, 171, 15–22. [Google Scholar] [CrossRef]
- Barloy, D.; Lemoine, J.; Abelard, P.; Tanguy, A.M.; Rivoal, R.; Jahier, J. Marker-assisted pyramiding of two cereal cyst nematode resistance genes from Aegilops variabilis in wheat. Mol. Breed. 2007, 20, 31–40. [Google Scholar] [CrossRef]
- Marais, G.F.; McCallum, B.; Snyman, J.E.; Pretorius, Z.A.; Marais, A.S. Leaf rust and stripe rust resistance genes Lr54 and Yr37 transferred to wheat from Aegilops kotschyi. Plant Breed. 2005, 124, 538–541. [Google Scholar] [CrossRef]
- Yu, M.Q.; Person-Dedrywer, F.; Jahier, J. Resistance to root knot nematode, Meloidogyne naasi (Franklin) transferred from Aegilops variabilis Eig to bread wheat. Agronomie 1990, 6, 451–456. [Google Scholar] [CrossRef] [Green Version]
- Kuraparthy, V.; Chhuneja, P.; Dhaliwal, H.S.; Kaur, S.; Bowden, R.L.; Gill, B.S. Characterization and mapping of cryptic alien introgression from Aegilops geniculata with new leaf rust and stripe rust resistance genes Lr57 and Yr40 in wheat. Theor. Appl. Genet. 2007, 114, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Williamson, V.M.; Thomas, V.; Ferris, H.; Dubcovsky, J. An Aegilops ventricosa translocation confers resistance against root-knot nematodes to common wheat. Crop Sci. 2013, 53, 1412–1418. [Google Scholar] [CrossRef] [Green Version]
- Kuraparthy, V.; Sood, S.; Guedira, G.B.; Gill, B.S. Development of a PCR assay and marker-assisted transfer of leaf rust resistance gene Lr58 into adapted winter wheats. Euphytica 2011, 180, 227–234. [Google Scholar] [CrossRef]
- Delibes, A.; Del Morala, J.; Martin-Sanchez, J.A.; Mejias, A.; Gallego, M.; Casado, D. Hessian fly-resistance gene transferred from chromosome 4Mv of Aegilops ventricosa to Triticum aestivum. Theor. Appl. Genet. 1997, 94, 858–864. [Google Scholar] [CrossRef]
- Marais, G.F.; Marais, A.S.; McCallum, B.; Pretorius, Z. Transfer of leaf rust and stripe rust resistance genes Lr62 and and Yr42 from Aegilops neglecta Req. ex Bertol. to common wheat. Crop Sci. 2009, 49, 871–879. [Google Scholar] [CrossRef]
- Martin-Sanchez, J.A.; Gomez-Colmenarejo, M.; Del Morel, J.; Sin, E.; Montes, M.J.; Gonzalez-Belinchon, C. A new Hessian fly resistance gene (H30) transferred from wild grass Aegilops triuncialis to hexaploid wheat. Theor. Appl. Genet. 2003, 106, 1248–1255. [Google Scholar] [CrossRef]
- Marais, G.F.; Wessels, W.G.; Horn, M.; du Toit, F. Association of a stem rust resistance gene (Sr45) and two Russian wheat aphid resistance genes (Dn5 and Dn7) with mapped structural loci in common wheat. S. Afr. J. Plant Soil 1988, 15, 67–71. [Google Scholar] [CrossRef]
- Raupp, W.J.; Amri, A.; Hatchett, J.H.; Gill, B.S.; Wilson, D.L.; Cox, T.S. Chromosomal location of hessian fly-resistance genes H22, H23, and H24 derived from Tricum tauschii in the D Genome of wheat. J. Hered. 1993, 84, 142–145. [Google Scholar] [CrossRef]
- Riley, R.; Chapman, V.; Johnson, R. The incorporation of alien disease resistance to wheat by genetic interference with regulation of meiotic chromosome synapsis. Genet. Res. Camb. 1968, 12, 199–219. [Google Scholar] [CrossRef]
- Friebe, B.; Mukai, Y.; Dhaliwal, H.S.; Martin, T.J.; Gill, B.S. Identification of alien chromatin specifying resistance to wheat streak mosaic and greenbug in wheat germplasm by C-banding and in situ hybridization. Theor. Appl. Genet. 1991, 81, 381–389. [Google Scholar]
- Hollenhorst, M.M.; Joppa, L.R. Chromosomal location of genes for resistance to greenbug in ‘Largo’ and ‘Amigo’ wheats. Crop Sci. 1993, 23, 91–93. [Google Scholar] [CrossRef]
- Thomas, J.B.; Conner, R.I. Resistance to colonization by the wheat curl mite in Aegilops squarrosa and its inheritance after transfer to common wheat. Crop Sci. 1986, 26, 527–530. [Google Scholar] [CrossRef]
- Malik, R.; Brown-Guerdira, G.L.; Smith, C.M.; Harvey, T.L.; Gill, B.S. Genetic mapping of wheat curl mite resistance genes Cmc3 and Cmc4 in common wheat. Crop Sci. 2003, 43, 644–650. [Google Scholar] [CrossRef]
- Wan, Y.; Poole, R.L.; Huttly, A.K.; Toscano-Underwood, C.; Feeney, K.; Welham, S.; Mitchell, R.A. Transcriptome analysis of grain development in hexaploid wheat. BMC Genom. 2008, 99, 121. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Zhen, S.; Wang, S.; Wang, Y.; Cao, H.; Zhang, Y.; Yan, Y. Comparative transcriptome analysis of wheat embryo and endosperm responses to ABA and H2O2 stresses during seed germination. BMC Genom. 2016, 17, 97. [Google Scholar] [CrossRef] [Green Version]
- Rangan, P.; Furtado, A.; Henry, R.J. The transcriptome of the developing grain: A resource for understanding seed development and the molecular control of the functional and nutritional properties of wheat. BMC Genom. 2017, 18, 766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, M.; Rai, S.; Venkadesan, S.; Sinha, S.K.; Mohan, S.; Mandal, P.K. Transcriptome analysis reveals important candidate genes related to nutrient reservoir, carbohydrate metabolism, and defence proteins during grain development of hexaploid bread wheat and its diploid progenitors. Genes 2020, 11, 509. [Google Scholar] [CrossRef]
- Brenchley, R.; Spannagl, M.; Pfeifer, M.; Barker, G.L.; D’Amore, R.; Allen, A.M.; Kay, S. Analysis of the bread wheat genome using whole-genome shotgun sequencing. Nature 2012, 491, 705–710. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Li, G.; Chu, Z.; Ru, Z.; Jiang, X.; Wen, Z.; Wei, W. Transcriptome analysis reveals important candidate genes involved in grain-size formation at the stage of grain enlargement in common wheat cultivar “Bainong 4199”. PLoS ONE 2019, 14, e0214149. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, M.; Li, W. Reducing expression of a nitrate—Responsive bZIP transcription factor increases grain yield and N use in wheat. Plant Biotechnol. J. 2019, 17, 1823–1833. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, M.; Morgounov, A.; Belamkar, V.; Baenziger, P.S. Genome-wide association study reveals novel genomic regions for grain yield and yield-related traits in drought-stressed synthetic hexaploid wheat. Int. J. Mol. Sci. 2018, 19, 3011. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.E.I.; Zhihui, W.U.; Yufeng Zhang, D.G. Transcriptome analysis of wheat grain using RNA-Seq. Front. Agric. Sci. Eng. 2015, 11, 214–222. [Google Scholar]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef]
- Kucek, L.K.; Veenstra, L.D.; Amnuaycheewa, P.; Sorrells, M.E. A grounded guide to gluten: How modern genotypes and processing impact wheat sensitivity. Compr. Rev. Food Sci. Food Saf. 2015, 14, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki-Ourang, S.; Mehrabi, A.A. Molecular detection of glutenin and gliadin genes in the domesticated and wild relatives of wheat using allele-specific markers. Cereal Res. Commun. 2018, 46, 510–520. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Chen, F.; Cui, D. Proteomic analysis of middle and late stages of bread wheat (Triticum aestivum L.) grain development. Front. Plant Sci. 2015, 6, 735. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Mouille, G.; Kosar-Hashemi, B.; Rahman, S.; Clarke, B.; Gale, K.R.; Morell, M.K. The structure and expression of the wheat starch synthase III gene. Motifs in the expressed gene define the lineage of the starch synthase III gene family. Plant Physiol. 2000, 123, 613–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowles, D.J. Defense-related proteins in higher plants. Annu. Rev. Biochem. 1990, 59, 873–907. [Google Scholar] [CrossRef]
- Junker, Y.; Zeissig, S.; Kim, S.J.; Barisani, D.; Wieser, H.; Leffer, D.A.; Kelly, C.P. Wheat amylase trypsin inhibitors drive intestinal inflammation via activation of toll-like receptor 4. J. Exp. Med. 2012, 209, 2395–2408. [Google Scholar] [CrossRef]
- Ziegler, K.; Neumann, J.; Liu, F.; Fröhlich-Nowoisky, J.; Cremer, C.; Saloga, J.; Lucas, K. Nitration of wheat amylase trypsin inhibitors increases their innate and adaptive immunostimulatory potential in vitro. Front. Immunol. 2018, 9, 3174. [Google Scholar] [CrossRef] [Green Version]
- Wielkopolan, B.; Krawczyk, K.; Obrepalska-Steplowska, A. Gene expression of serine and cysteine proteinase inhibitors during cereal leaf beetle larvae feeding on wheat: The role of insect-associated microorganisms. Arthropod Plant Interact. 2018, 12, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Borrill, P.; Adamski, N.; Uauy, C. Genomics as the key to unlocking the polyploid potential of wheat. New Phytol. 2015, 208, 1008–1022. [Google Scholar] [CrossRef]
- Yu, Y.; Zhu, D.; Ma, C.; Cao, H.; Wang, Y.; Xu, Y.; Yan, Y. Transcriptome analysis reveals key differentially expressed genes involved in wheat grain development. Crop J. 2016, 44, 92–106. [Google Scholar] [CrossRef] [Green Version]
- El Baidouri, M.; Murat, F.; Veyssiere, M.; Molinier, M.; Flores, R.; Burlot, L.; Salse, J. Reconciling the evolutionary origin of bread wheat (Triticum aestivum). New Phytol. 2017, 213, 1477–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scossa, F.; Laudencia-Chingcuanco, D.; Anderson, O.D.; Vensel, W.H.; Lafiandra, D.; D’Ovidio, R.; Masci, S. Comparative proteomic and transcriptional profiling of a bread wheat cultivar and its derived transgenic line overexpressing a low molecular weight glutenin subunit gene in the endosperm. Proteomics 2008, 88, 2948–2966. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.F.; Geniza, M.; Hanumappa, M.; Naithani, S.; Sullivan, C.; Preece, J.; Tiwari, V.K.; Elser, J.; Leonard, J.M.; Sage, A. De Novo transcriptome assembly and analyses of gene expression during photomorphogenesis in diploid wheat Triticum monococcum. PLoS ONE 2014, 9, e95855. [Google Scholar] [CrossRef]
- Appels, R.; Eversole, K.; Feuillet, C.; Keller, B.; Rogers, J.; Stein, N.; Ronen, G. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [PubMed] [Green Version]
- Miki, Y.; Yoshida, K.; Mizuno, N.; Nasuda, S.; Sato, K.; Takumi, S. Origin of wheat B-genome chromosomes inferred from RNA sequencing analysis of leaf transcripts from section Sitopsis species of Aegilops. DNA Res. 2019, 26, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Payne, P.I. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu. Rev. Plant Physiol. 1987, 38, 141–153. [Google Scholar] [CrossRef]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 77, 945. [Google Scholar]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [Green Version]
- Salentijn, E.M.; Esselink, D.G.; Goryunova, S.V.; van der Meer, I.M.; Gilissen, L.J.; Smulders, M.J. Quantitative and qualitative differences in celiac disease epitopes among durum wheat varieties identified through deep RNA-amplicon sequencing. BMC Genom. 2013, 14, 905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katagiri, M.; Masuda, T.; Tani, F.; Kitabatake, N. Expression and development of wheat proteins during maturation of wheat kernel and the rheological properties of dough prepared from the flour of mature and immature wheat. Food Sci. Technol. Res. 2011, 17, 111–120. [Google Scholar] [CrossRef] [Green Version]
- McMaugh, S.J.; Thistleton, J.L.; Anschaw, E.; Luo, J.; Konik-Rose, C.; Wang, H.; Morell, M.K. Suppression of starch synthase I expression affects the granule morphology and granule size and fine structure of starch in wheat endosperm. J. Exp. Bot. 2014, 65, 2189–2201. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Cabrera, A.; Hoffstetter, A.; Griffey, C.; Van Sanford, D.; Costa, J.; Sneller, C. Genomic selection for wheat traits and trait stability. Theor. Appl. Genet. 2016, 129, 1697–1710. [Google Scholar] [CrossRef]
- Tetlow, I.; Emes, M. Starch biosynthesis in the developing endosperms of grasses and cereals. Agronomy 2017, 7, 81. [Google Scholar] [CrossRef] [Green Version]
- Stanley, D.; Rejzek, M.; Naested, H.; Smedley, M.; Otero, S.; Fahy, B.; Denyer, K. The role of α-glucosidase in germinating barley grains. Plant Physiol. 2011, 155, 932–943. [Google Scholar] [CrossRef] [Green Version]
- Whan, A.; Dielen, A.S.; Mieog, J.; Bowerman, A.F.; Robinson, H.M.; Byrne, K.; Ral, J.P. Engineering—Amylase levels in wheat grain suggests a highly sophisticated level of carbohydrate regulation during development. J. Exp. Bot. 2014, 65, 5443–5457. [Google Scholar] [CrossRef] [Green Version]
- Blackig, M.; Corbineau, F.; Grzesikit, M.; Guyi, P.; Come, D. Carbohydrate metabolism in the developing and maturing wheat embryo in relation to its desiccation tolerance. J. Exp. Bot. 1996, 47, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Malik, A.H. Nutrient Uptake, Transport and Translocation in Cereals: Influences of Environmental and Farming Conditions; Swedish University of Agricultural Sciences: Alnarp, Sweden, 2009. [Google Scholar]
- Altpeter, F.; Diaz, I.; McAuslane, H.; Gaddour, K.; Carbonero, P.; Vasil, I.K. Increased insect resistance in transgenic wheat stably expressing trypsin inhibitor CMe. Mol. Breed. 1999, 5, 53–63. [Google Scholar] [CrossRef]
- Florack, D.E.A.; Stiekema, W.J. Thionins: Properties, possible biological roles and mechanisms of action. Plant Mol. Biol. 1994, 26, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Plattner, S.; Gruber, C.; Stadlmann, J.; Widmann, S.; Gruber, C.W.; Altmann, F.; Bohlmann, H. Isolation and characterization of a thionin proprotein-processing enzyme from barley. J. Biol. Chem. 2015, 290, 18056–18067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, L.J.; Chen, J.; Liu, M.; Pan, N.; Okamoto, H.; Lin, Z.; Zhao, X. Molecular cloning and functional analysis of a novel type of Bowman-Birk inhibitor gene family in rice. Plant Physiol. 2003, 133, 560–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chilosi, G.; Caruso, C.; Caporale, C.; Leonardi, L.; Bertini, L.; Buzi, A.; Buonocore, V. Antifungal activity of a bowman–birk-type trypsin inhibitor from wheat kernel. J. Phytopathol. 2000, 148, 477–481. [Google Scholar] [CrossRef]
- Tedeschi, F.; Di Maro, A.; Facchiano, A.; Costantini, S.; Chambery, A.; Bruni, N.; Capuzzi, V.; Ficca, A.G.; Poerio, E. Wheat Subtilisin/Chymotrypsin Inhibitor (WSCI) as a scaffold for novel serine protease inhibitors with a given specificity. Mol. Biosyst. 2012, 8, 3335–3343. [Google Scholar] [CrossRef]
- Tundo, S.; Lupi, R.; Lafond, M.; Giardina, T.; Larré, C.; Denery-Papini, S.; Masci, S. Wheat ATI CM3, CM16 and 0.28 allergens produced in Pichia Pastoris display a different eliciting potential in food allergy to wheat. Plants 2018, 7, 101. [Google Scholar] [CrossRef] [Green Version]
- Rauf, K.; Rahman, R.; Saeed, A.; Ali, M.; Noureen, F.; Amir, R.; Gul, A. Next-generation sequencing in bread wheat. In Climate Change and Food Security with Emphasis on Wheat; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Wang, B.; Sun, Y.F.; Song, N.; Wang, X.J.; Feng, H.; Huang, L.L.; Kang, Z.S. Identification of UV-B-induced microRNAs in wheat. Genet. Mol. Res. 2013, 12, 4213–4221. [Google Scholar] [CrossRef]
- Li, A.; Liu, D.; Wu, J. mRNA and small RNA transcriptomes reveal insights into dynamic homoeolog regulation of allopolyploid heterosis in nascent hexaploid wheat. Plant Cell 2014, 26, 1878–1900. [Google Scholar] [CrossRef] [Green Version]
- Feng, N.; Song, G.; Guan, J. Transcriptome profiling of wheat inflorescence development from spikelet initiation to floral patterning identified stage-specific regulatory genes. Plant Physiol. 2017, 174, 1779–1794. [Google Scholar] [CrossRef]
- Akhunova, A.R.; Matniyazov, R.T.; Liang, H.; Akhunov, E.D. Homoeolog-specific transcriptional bias in allopolyploid wheat. BMC Genom. 2010, 11, 505. [Google Scholar] [CrossRef] [Green Version]
- Qi, B.; Huang, W.; Zhu, B. Global transgenerational gene expression dynamics in two newly synthesized allohexaploid wheat (Triticum aestivum) lines. BMC Biol. 2012, 10, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelaifa, H.; Chague, V.; Chalabi, S. Prevalence of gene expression additivity in genetically stable wheat allohexaploids. New Phytol. 2013, 197, 730–736. [Google Scholar] [CrossRef]
- Wang, J.; Tian, L.; Lee, H.S. Genomewide nonadditive gene regulation in Arabidopsis allotetraploids. Genetics 2006, 172, 507–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaucheret, H. Post-transcriptional small RNA pathways in plants: Mechanisms and regulations. Genes Dev. 2006, 20, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Zhang, C.; Baulcombe, D.C.; Chen, Z.J. Maternal siRNAs as regulators of parental genome imbalance and gene expression in endosperm of Arabidopsis seeds. Proc. Natl. Acad. Sci. USA 2012, 109, 5529–5534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.; Kim, E.D.; Chen, Z.J. Duplicate genes increase expression diversity in closely related species and allopolyploids. Proc. Natl. Acad. Sci. USA 2009, 106, 2295–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, D.W.; Lu, J.; Chen, Z.J. Big roles for small RNAs in polyploidy, hybrid vigor, and hybrid incompatibility. Curr. Opin. Plant Biol. 2012, 15, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Ng, D.W.; Zhang, C.; Miller, M.; Palmer, G.; Whiteley, M.; Tholl, D.; Chen, Z.J. Cis- and trans-regulation of miR163 and target genes confers natural variation of secondary metabolites in two Arabidopsis species and their allopolyploids. Plant Cell 2011, 23, 1729–1740. [Google Scholar] [CrossRef] [Green Version]
- Kenan-Eichler, M.; Leshkowitz, D.; Tal, L.; Noor, E.; Melamed-Bessudo, C.; Feldman, M.; Levy, A.A. Wheat hybridization and polyploidization results in deregulation of small RNAs. Genetics 2011, 188, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.J. Genomic and epigenetic insights into the molecular basesofheterosis. Nat. Rev. Genet. 2013, 14, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Lu, J.; Tian, L. Small RNAs serve as a genetic buffer against genomic shock in Arabidopsis interspecific hybrids and allopolyploids. Proc. Natl. Acad. Sci. USA 2009, 106, 17835–17840. [Google Scholar] [CrossRef] [Green Version]
- Grover, C.E.; Gallagher, J.P.; Szadkowski, E.P.; Yoo, M.J.; Flagel, L.E.; Wendel, J.F. Homoeolog expression bias and expression level dominance in allopolyploids. New Phytol. 2012, 196, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, D.; Chelaifa, H.; Jahier, J.; Chalhoub, B. Re-programming of gene expression in the genetically stable bread allo-hexaploid wheat. In Polyploid and Hybrid Genomics; Chen, Z.J., Birchler, J.A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Han, R.; Jian, C.; Lv, J. Identification and characterization of microRNAs in the flag leaf and developing seed of wheat (Triticum aestivum L.). BMC Genom. 2014, 15, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, S.K.; Rani, M.; Bansal, N.; Gayatri, V.K.; Mandal, P.K. Nitrate starvation induced changes in root system architecture, carbon:nitrogen metabolism, and miRNA expression in nitrogen-responsive wheat genotypes. Appl. Biochem. Biotechnol. 2015, 177, 1299–1312. [Google Scholar] [CrossRef]
- Li, Y.F.; Wei, K.; Wang, M. Identification and temporal expression analysis of conserved and novel MicroRNAs in the leaves of winter wheat grown in the field. Front. Genet. 2019, 10, 779. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Wang, Y.; Yao, Y.; Xie, C.; Peng, H.; Ni, Z.; Sun, Q. Diverse set of microRNAs are responsive to powdery mildew infection and heat stress in wheat (Triticum aestivum L.). BMC Plant Biol. 2010, 10, 123. [Google Scholar] [CrossRef] [Green Version]
- Akdogan, G.; Tufekci, E.D.; Uranbey, S.; Unver, T. miRNA-based drought regulation in wheat. Funct. Integr. Genom. 2016, 16, 221–233. [Google Scholar] [CrossRef]
- Chen, X.Y.; Yang, Y.; Ran, L.P.; Dong, Z.D.; Zhang, E.J.; Yu, X.R.; Xiong, F. Novel insights into miRNA regulation of storage protein biosynthesis during wheat caryopsis development under drought stress. Front. Plant Sci. 2017, 8, 1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Wang, Q.; Wei, L.; Liang, Y.; Dai, J.; Xia, G.; Liu, S.H. Small RNA and degradome sequencing used to elucidate the basis of tolerance to salinity and alkalinity in wheat. BMC Plant Biol. 2018, 18, 195. [Google Scholar] [CrossRef]
- Ma, X.; Xin, Z.; Wang, Z.; Yang, Q.; Guo, S.; Guo, X.; Cao, L.; Lin, T. Identification and comparative analysis of differentially expressed miRNAs in leaves of two wheat (Triticum aestivum L.) genotypes during dehydration stress. BMC Plant Biol. 2015, 15, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.; Zhang, L.; Xu, C.; Yuan, S.; Zhang, F.; Zheng, Y.; Zhao, C. Uncovering small RNA-mediated responses to cold stress in a wheat thermosensitive genic male-sterile line by deep sequencing. Plant Physiol. 2012, 159, 721–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alptekin, B.; Budak, H. Wheat miRNA ancestors: Evident by transcriptome analysis of A, B, and D genome donors. Funct. Integr. Genom. 2016, 17, 171–187. [Google Scholar] [CrossRef]
- Gupta, O.P.; Meena, N.L.; Sharma, I.; Sharma, P. Differential regulation of microRNAs in response to osmotic, salt and cold stresses in wheat. Mol. Biol. Rep. 2014, 41, 4623–4629. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.; Chen, J.; Xu, H.; Dong, Z.; Chen, F.; Cui, D. Identification and comparative analysis of microRNA in wheat (Triticum aestivum L.) callus derived from mature and immature embryos during in vitro culture. Front. Plant Sci. 2016, 7, 1302. [Google Scholar] [CrossRef] [Green Version]
- Hou, G.; Du, C.; Gao, H. Identification of microRNAs in developing wheat grain that are potentially involved in regulating grain characteristics and the response to nitrogen levels. BMC Plant Biol. 2020, 20, 87. [Google Scholar] [CrossRef]
- Pandey, R.; Joshi, G.; Bhardwaj, A.R.; Agarwal, M.; Katiyar-Agarwal, S. A comprehensive genome-wide study on tissue-specific and abiotic stress-specific miRNAs in Triticum aestivum. PLoS ONE 2014, 9, e95800. [Google Scholar] [CrossRef]
- Kantar, M.; Akpinar, B.A.; Valarik, M.; Lucas, S.J.; Dolezel, J.; Hernandez, P.; Budak, H. Subgenomic analysis of microRNAs in polyploidy wheat. Funct. Integr. Genom. 2012, 12, 465–479. [Google Scholar] [CrossRef]
- Song, G.; Zhang, R.; Zhang, S. Response of microRNAs to cold treatment in the young spikes of common wheat. BMC Genom. 2017, 18, 212. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Guo, G.; Du, J.; Guo, W.; Peng, H.; Ni, Z.; Sun, Q.; Yao, Y. Whole-genome discovery of miRNAs and their targets in wheat (Triticum aestivum L.). BMC Plant Biol. 2014, 14, 142. [Google Scholar] [CrossRef] [Green Version]
- Zuluaga, D.L.; De Paola, D.; Janni, M.; Curci, P.L.; Sonnante, G. Durum wheat miRNAs in response to nitrogen starvation at the grain filling stage. PLoS ONE 2017, 12, e0183253. [Google Scholar] [CrossRef]
- Liu, H.; Searle, I.R.; Watson-Haigh, N.S.; Baumann, U.; Mather, D.E.; Able, A.J.; Able, J.A. Genome-wide identification of microRNAs in leaves and the developing head of four durum genotypes during water deficit stress. PLoS ONE 2015, 10, e0142799. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Genotypic water-deficit stress responses in durum wheat: Association between physiological traits, microRNA regulatory modules and yield components. Funct. Plant. Biol. 2017, 44, 538–551. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Able, A.J.; Able, J.A. Water-deficit responsive microRNAs and their targets in four durum wheat genotypes. Funct. Integr. Genom. 2017, 17, 237–251. [Google Scholar] [CrossRef]
- Fileccia, V.; Bertolini, E.; Ruisi, P. Identification and characterization of durum wheat microRNAs in leaf and root tissues. Funct. Integr. Genom. 2017, 17, 583–598. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Small RNAs and their targets are associated with the transgenerational effects of water-deficit stress in durum wheat. Sci. Rep. 2021, 11, 3613. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Nitrogen starvation-responsive MicroRNAs are affected by transgenerational stress in durum wheat seedlings. Plants 2021, 10, 826. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Nie, X.; Cui, L.; Deng, P.; Wang, M.; Song, W. Genome-wide identification and characterization of salinity stress-responsive miRNAs in wild emmer wheat (Triticum turgidum ssp. dicoccoides). Genes 2017, 8, 156. [Google Scholar] [CrossRef] [Green Version]
- Akpinar, B.A.; Kantar, M.; Budak, H. Root precursors of microRNAs in wild emmer and modern wheats show major differences in response to drought stress. Funct. Integr. Genom. 2015, 15, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Akpinar, B.A.; Budak, H. Dissecting miRNAs in wheat D genome progenitor, Aegilops tauschii. Front. Plant. Sci. 2016, 4, 606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appels, R.; Eversole, K.; Stein, N. Shifting the limits in wheat research and breeding using a fully an-notated reference genome by the international wheat genome sequencing consortium. Science 2018, 361, 7191. [Google Scholar]
- Hodges, E.; Xuan, Z.; Balija, V. Genome-wide in situ exon capture for selective resequencing. Nature 2007, 39, 1522. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.; Marra, M.A. Applications of next-generation sequencing technologies in functional genomics. Genomics 2008, 82, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Berkman, P.J.; Lai, K.; Lorenc, M.T.; Edwards, D. Next-generation sequencing applications for wheat crop improvement. Am. J. Bot. 2012, 99, 365–371. [Google Scholar] [CrossRef]
- Wang, S.; Wong, D.; Forrest, K. Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Arora, S.; Singh, N.; Kaur, S.; Bains, N.S.; Uauy, C.; Poland, J.; Chhuneja, P. Genome-wide association study of grain architecture in wild wheat Aegilops tauschii. Front. Plant Sci. 2017, 8, 886. [Google Scholar] [CrossRef] [Green Version]
- Clavijo, B.J.; Venturini, L.; Schudoma, C. An improved assembly and annotation of the allohexaploid wheat genome identifies complete families of agronomic genes and provides genomic evidence for chromosomal translocations. Genome Res. 2017, 27, 885–896. [Google Scholar] [CrossRef] [Green Version]
- Mooney, E.H.; Mcgraw, J.B. Effects of self-pollination and outcrossing with cultivated plants in small natural populations of American ginseng, Panax quinquefolius. Am. J. Bot. 2007, 94, 1677–1687. [Google Scholar] [CrossRef] [Green Version]
- Lozada, D.N.; Mason, R.E.; Babar, M.A. Association mapping reveals loci associated with multiple traits that affect grain yield and adaptation in soft winter wheat. Euphytica 2017, 213, 1–15. [Google Scholar] [CrossRef]
- Sukumaran, S.; Reynolds, M.P.; Sansaloni, C. Genome-wide association analyses identify QTL hotspots for yield and component traits in durum wheat grown under yield potential, drought, and heat stress environments. Front. Plant Sci. 2018, 9, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, P.; Lin, Y.; Hu, Y. Genome-wide association study of drought-related resistance traits in Aegilops tauschii. Genet. Mol. Biol. 2016, 39, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, A.A.; Pour-Aboughadareh, A.; Mansouri, S.; Hosseini, A. Genome-wide association analysis of root system architecture features and agronomic traits in durum wheat. Mol. Breed. 2020, 40, 55. [Google Scholar] [CrossRef]
Figure 1.
The position of the Fertile Crescent (FC) in the Middle East and world. This region embraces a wide geographical range between the Persian Gulf and the eastern coast of the Mediterranean Sea (highlighted in green).
Figure 1.
The position of the Fertile Crescent (FC) in the Middle East and world. This region embraces a wide geographical range between the Persian Gulf and the eastern coast of the Mediterranean Sea (highlighted in green).

Figure 2.
Information flow diagram for the domestication trend of wheat.


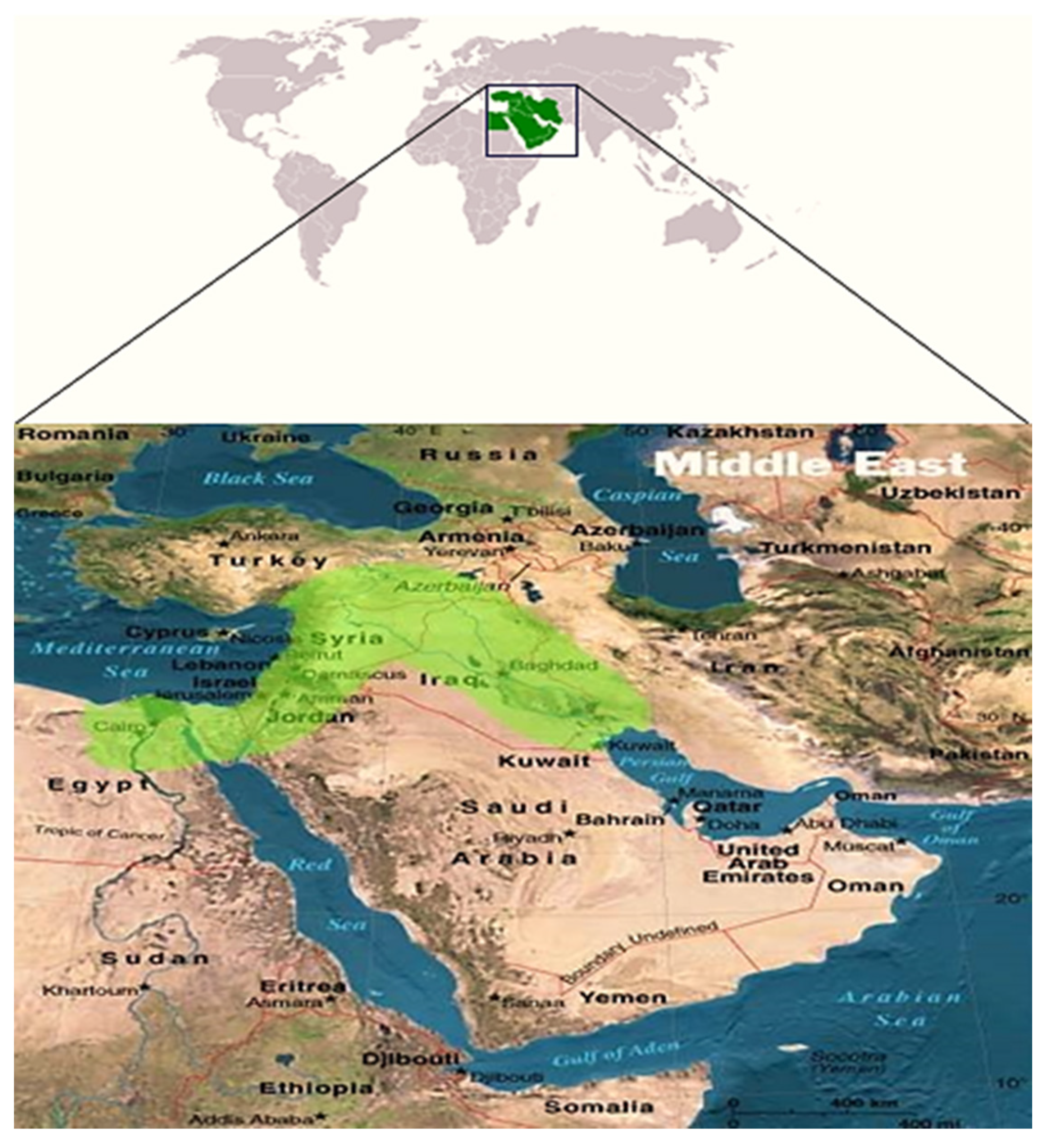{kind=link}
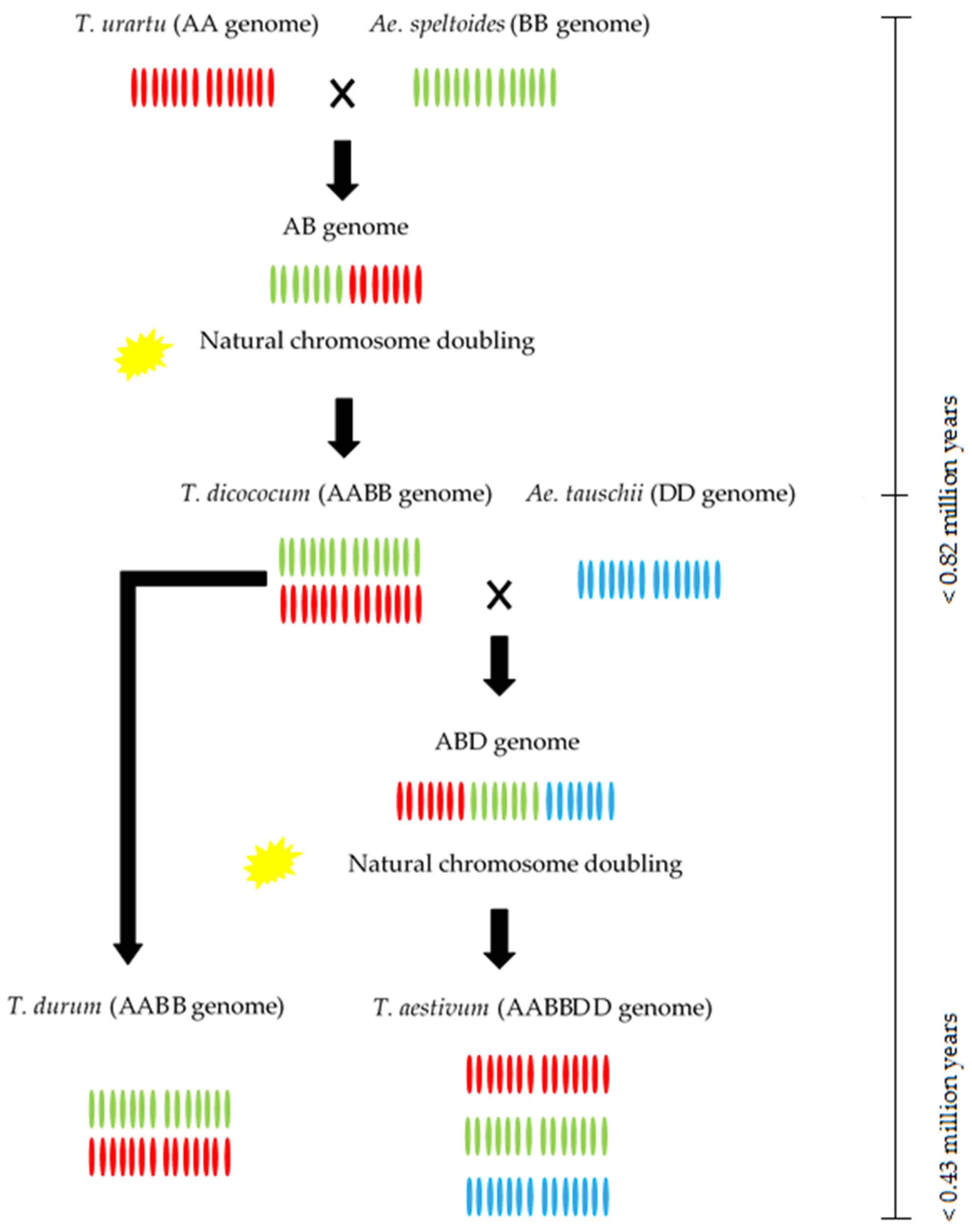{kind=link}
Table 1.
Some key articles regarding the trend of wheat evolution.
| Article | Reference |
|---|---|
| Evolution in the genus Triticum and the origin of cultivated wheat | [9] |
| Genome symbols and plasma types in the wheat group | [10] |
| Cytogenetics of wheat and its close wild relatives-Triticum and Aegilops | [11] |
| Genome symbols in the Triticeae (Poaceae) | [12] |
| Phylogenetic relationships of Triticum and Aegilops and evidence for the origin of the A, B, and D genomes of common wheat (Triticum aestivum) | [13] |
| Evolution of domesticated bread wheat | [14] |
| Wheat domestication: Lessons for the future | [15] |
| Distinguishing wild and domestic wheat and barley spikelets from early Holocene sites in the Near East | [16] |
| Emergence of agriculture in the foothills of the Zagros mountains of Iran | [17] |
| On the Identification of domesticated Emmer wheat, Triticum turgidum subsp. dicoccum (Poaceae), in the Aceramic Neolithic of the Fertile Crescent | [18] |
| DArTseq-based analysis of genomic relationships among species of tribe Triticeae | [19] |
| Domestication and crop evolution of wheat and barley: Genes, genomics, and future directions | [20] |
| Bread wheat: a role model for plant domestication and breeding | [21] |
| Roadmap for accelerated domestication of an emerging perennial grain crop | [22] |
| Current progress in understanding and recovering the wheat genes lost in evolution and domestication | [23] |
Table 2.
List of different Aegilops and Triticum species along with their genomic constitutions.
| Species | Genome | ||
|---|---|---|---|
| References | |||
| [26] | [27] | [10] | |
| T. monococcum L. | A | ||
| T. urartu Tumanian ex Gandilyan | A | ||
| Ae. speltoides Tausch | S | ||
| Ae. bicorne (Forsk.) Jaub. & Sp. | Sb | ||
| Ae. longissimum Schweinf. & Muschli in Muschli | Sl | ||
| Ae. sharonensis Eig | Sl | ||
| Ae. searsii Feldman & Kislev | Ss | ||
| Ae. mutica Bioss. | Mt | T | |
| Ae. tauschii Coss. | D | ||
| Ae. comosa Sibth. & Sm. | M | ||
| Ae. uniaristata Vis. | Mt | Un | N |
| Ae. caudata L. | C | ||
| Ae. umbellulata Zhuk. | Cu | U | |
| T. dicoccoides Korn | AB | ||
| T. dicoccum Schrank | AB | ||
| T. durum Desf. | AB | ||
| T. turgidum L. | AB | ||
| T. persicum (Percival) Vavilov ex Zhukovsky | AB | ||
| T. aestivum L. em. Thell. | ABD | ||
| T. spelta L. | ABD | ||
| T. compactum Host | ABD | ||
| T. sphaerococcum Perc. | ABD | ||
| T. macha Dek. et Men. | ABD | ||
| T. timopheevi Zhuk. | AG | M | |
| T. zhukovskyi Men. et Er. | AAG | AAUG | |
| Ae. ovate L. | CUMO | UM | UM |
| Ae. biuncialis Vis. | CUMb | UM | |
| Ae. columnaris Zhuk. | CUMc | UM | |
| Ae. triaristata Wild. | CUMt | UM | UM |
| Ae. recta (Zhuk.) Chen. | CUMtX | UMUn | UMX |
| Ae. variabilis L. | CUSV | US | USl |
| Ae. triuncialis L. | CUC | UC | |
| Ae. cylindrica Host | CD | ||
| Ae. crassa (4x) Boiss. | DJ | DM | DCX |
| Ae. crassa (6x) Boiss. | DJX | DDM | DCXSS |
| Ae. vavilovi (Zhuk.) Chen. | DMS | CCXSS | |
| Ae. ventricosa Tausch | DMV | DUn | DN |
| Ae. juvenile (Thell.) Eig | DMU | ||
Table 3.
List of several studies that reveal high levels of phenotypic diversity in wild relatives of wheat.
Table 3.
List of several studies that reveal high levels of phenotypic diversity in wild relatives of wheat.
| Target Species | Traits | References |
|---|---|---|
| A. tauschii | Plant height, peduncle length, number of tillers per plant, number of spikes per plant, number of spikelets per spike, spike length, leaf length, number of grains per spike, length and width of rachis node and spike, seed color, glume color, glume hairiness | [32,33,34,35,36,37] |
| T. boeoticum | Plant height, peduncle length, spike length, number of spikes, number of spikelet per spike, number of grains per spike, leaves number, glume shape, glume color, branched spike, anther color, grain color, awn color, plant height, glume length, flag leaf length, leaf length | [31,33,35,38,39] |
| T. aestivum | Plant height, peduncle length, number of tillers per plant, number of spikes per plant, number of spikelets per spike, spike length, leaf length, number of grains per spike, glume shape, glume color, branched spike, anther color, grain color, awn color, plant height, glume length | [31,33,36,39,40,41,42,43,44,45] |
| T. durum | Leaf length, number of seeds per spike, spike dry matter, spike length, number of spikes, number of spikelets per spike, thousand seeds weight, harvest index, grain yield, various physiological traits | [31,33,46,47,48,49] |
| T. dicoccoides | Plant height, peduncle length, number of tillers per plant, number of spikes per plant, number of spikelets per spike, spike length, heading date, flag leaf length, flag leaf width, harvest index, glume shape, glume color, branched spike, anther color, grain color, awn color, glume length | [31,33,37,46,47,48,50] |
| T. urartu | Plant height, peduncle length, leaf length, number of spikes, spike length, number of spikelets per spike, seed yield | [33,34,51] |
| Ae. caudate, Ae. cylindrical, Ae. crassa, Ae. speltoide, Ae. umbellulata, Ae. neglecta, Ae. triuncialis, Ae. strangulata, Ae. ligustica, Ae. biuncialis, Ae. columnaris, Ae. vavilovii | Glume shape, glume color, branched spike, anther color, grain color, awn color, plant height, glume length, flag leaf length, spike length, number of stems, plant height, peduncle length, number of spikes, number of spikelets per spike, length and width of rachis node and spike, number seeds per spike, seed color, glume hairiness | [31,32,33] |
| T. monoccocum | Plant height, grain per spike, heading time, maturity time, hairiness, waxiness, growth habitus and grain hull | [34,52] |
| T. dicoccum | Plant height, grain per spike, heading time, maturity time, hairiness, waxiness, growth habitus and grain hull | [52] |
| T. aegilopoides | Plant height, spike length, number of stems | [34] |
Table 4.
List of several studies that reveal a high level of molecular diversity in wild relatives of wheat.
Table 4.
List of several studies that reveal a high level of molecular diversity in wild relatives of wheat.
| Marker | Species | References |
|---|---|---|
| RAPD | Ae. kotschyi, Ae. variabilis, Ae. tauschii, Ae. cylinddrica, Ae. crassa, Ae. biuncialis, Ae. triuncialis, Ae. geniculata, T. boeoticum | [59,60,61,62,63,64] |
| RFLP | Ae. geniculata | [65] |
| AFLP | Ae. kotschyi, Ae. variabilis, T. turgidum, T. dicoccon, T. dicoccoides, T. araraticum, T. monococcum, Ae. geniculata, Ae. ventricosa | [66,67,68,69,70] |
| IRAP | T. monococcum, T. boeoticum subsp. boeoticum, T. boeoticum subsp. Thaoudar., T. urartu | [71] |
| REMAP | T. boeoticum | [72] |
| TRAP | Ae. tauschii, Ae. cylindrica, and Ae. crassa | [73] |
| ISSR | Ae. ventricosa, Ae. taucshii, Ae. cylindrica, Ae. umbellulata, Ae. triuncialis, Ae. biuncialis, Ae. crassa, Ae. kotschyi, Ae. speltoides | [59,74,75,76] |
| SSR | T. turgidum, T. dicoccon, T. dicoccoides, T. araraticum, T. monococcum, Ae. geniculata, Ae. ventricosa, Ae. crassa, Ae. cylindrica, Ae. biuncialis, Ae. triuncialis, Ae. tauschii | [63,66,70,77,78,79,80,81,82,83,84,85,86] |
| cpSSR | Ae. cylinddrica, Ae. kotschyi, Ae. peregrina, Ae. triuncialis, Ae. uniaristata, Ae. speltoides, Ae. longissima, Ae. sharonensis, Ae. searsii, Ae. bicornis, Ae. tauschii, Ae. crassa, Ae. vavilovii, Ae. ventricosa, Ae. juvenalis, T. urartu, T. boeoticum, T. araraticum, T. dicoccoides | [87,88] |
| EST | Ae. caudata | [89] |
| EST-SSR | Ae. cylindrica, Ae. tauschii, Ae. triuncialis, Ae. crassa | [90] |
| SINE | Ae. umbellulata | [91] |
| ITE | T. boeoticum | [64] |
| SCoT | T. turgidum, T. boeoticum, T. urartu, Ae. tauschii, Ae. caudata, Ae. crassa, Ae. neglecta, Ae. triuncialis, Ae. speltoides | [73,92,93] |
| CBDP | T. turgidum, T. boeoticum, T. urartu, Ae. tauschii, Ae. caudata, Ae. crassa, Ae. neglecta, Ae. triuncialis, Ae. speltoides | [94,95] |
| DArT | T. turgidum, Ae. Kotschyi, Ae. cylindrica, Ae. neglecta, Ae. colummaris, Ae. biuncialis, Ae. triuncialis, Ae. juvenalis, Ae. tauschii | [96,97,98] |
| SNP | T. turgidum, Ae. Kotschyi, Ae. cylindrica, Ae. neglecta, Ae. colummaris, Ae. biuncialis, Ae. triuncialis, Ae. juvenalis, Ae. tauschii | [97,99,100,101,102,103] |
Table 5.
Identified or transferred resistance genes from various Aegilops species into bread wheat.
| Pest | Species | Gene | References | Disease | Species | Gene | References |
|---|---|---|---|---|---|---|---|
| Eyespot | Ae. ventricosa | Pch1 | [158] | Stem rust | Ae. speltoides | Sr32 | [159] |
| Powdery mildew | Ae. speltoides | Pm1d | [160] | Sr47 | [161] | ||
| Pm32 | [162] | Ae. comosa | Sr34 | [163] | |||
| Pm53 | [164] | Ae. ventricosa | Sr38 | [165] | |||
| Ae. longissima | Pm13 | [166] | Ae. searsii | Sr51 | [167] | ||
| Ae. geniculate | Pm29 | [168] | Ae. geniculate | Sr53 | [169] | ||
| Ae. umbellulata | Pm57 | [170] | Leaf rust | Ae. umbellulata | Lr9 | [171] | |
| Ae. tauschii | Pm19 | [172] | Lr76 | [173] | |||
| Pm34 | [174] | Ae. speltoides | Lr28 | [163] | |||
| Cyst nematode | Ae. ventricosa | Cre2 | [175] | Lr37 | [165] | ||
| Cre5 | [176] | Lr47 | [177] | ||||
| Cre6 | [178] | Lr51 | [179] | ||||
| Ae. triuncialis | Cre7 | [180] | Lr66 | [181] | |||
| Ae. peregrine | CreX | [182] | Ae. kotschyi | Lr54 | [183] | ||
| CreY | [182] | Ae. sharonensis | Lr56 | [181] | |||
| Root knot nematode | Ae. peregrine | Rkn2 | [184] | Ae. geniculate | Lr57 | [185] | |
| Ae. ventricosa | Rkn3 | [186] | Ae. triuncialis | Lr58 | [187] | ||
| Hessian fly | Ae. ventricosa | H27 | [188] | Ae. peregrine | Lr59 | [189] | |
| Ae. triuncialis | H30 | [190] | Ae. neglecta | Lr62 | [191] | ||
| Ae. tauschii | H22 | [192] | Strip rust | Ae. comosa | Yr8 | [193] | |
| H23 | [192] | Ae. ventricosa | Yr17 | [165] | |||
| Green bug | Ae. speltoides | Gb5 | [194] | Ae. sharonensis | Yr38 | [181] | |
| Ae. tauschii | Gb3 | [195] | Ae. geniculate | Yr40 | [185] | ||
| Russian wheat aphid | Ae. tauschii | Dn3 | [196] | Ae. neglecta | Yr42 | [189] | |
| Wheat curl mite | Ae. tauschii | Cmc4 | [197] | Ae. umbellulata | Yr70 | [173] |
Table 6.
The brief of identified microRNAs in wheat and some of its relatives.
| Species | Tissue | Conditions | References |
|---|---|---|---|
| T. aestivum | seedlings, flag leaves, seeds, root, leaf, spike, immature and mature embryos, grain, shoot, spikelet, dry grain, embryo of germinating seed, shoot, spike, grains, flag leaves | N-stress, cold stress, heat stress, drought stress, dehydration stress, UV-stress, high temperature stress, salinity stress, water-deficit stress | [260,261,262,263,264,265,266,267,268,269,270,271,272,273,274,275,276] [277,278,279,280,281,282,283] |
| T. durum | roots, leaves, flag leaf, spikes, heads, seedling, young seedlings | N-stress, water deficit, stress, early water stress, heat stress | [277,283] |
| T. dicoccoides | leaves, plant | drought and stresses | [274,284,285] |
| Ae. sharonensis, Ae. speltoides, Ae. tauschii, T. monococcum, and T. urartu | leaf, root | drought stress | [286] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pour-Aboughadareh, A.; Kianersi, F.; Poczai, P.; Moradkhani, H. Potential of Wild Relatives of Wheat: Ideal Genetic Resources for Future Breeding Programs. Agronomy 2021, 11, 1656. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081656
AMA Style
Pour-Aboughadareh A, Kianersi F, Poczai P, Moradkhani H. Potential of Wild Relatives of Wheat: Ideal Genetic Resources for Future Breeding Programs. Agronomy. 2021; 11(8):1656. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081656
Chicago/Turabian StylePour-Aboughadareh, Alireza, Farzad Kianersi, Peter Poczai, and Hoda Moradkhani. 2021. "Potential of Wild Relatives of Wheat: Ideal Genetic Resources for Future Breeding Programs" Agronomy 11, no. 8: 1656. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081656
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.