
Modelling the Effect of Salt and PEG on Water Uptake in Wheat Seeds


1
Faculty of Life and Environmental Sciences, University of Tsukuba, Tsukuba 305-8577, Japan
2
Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130102, China
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(8), 1660; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081660
Submission received: 20 July 2021
/
Revised: 13 August 2021
/
Accepted: 17 August 2021
/
Published: 20 August 2021
(This article belongs to the Section Soil and Plant Nutrition)
Abstract
:Water uptake is a seminal process in seed germination. Salt and polyethylene glycol (PEG) are known to retard seed germination rates and percentages, which is often attributed to osmotic effects. Here, we quantified water uptake in wheat seeds killed with a hot needle, finding evidence of three distinct water uptake pools. The fast pool was unaffected by salt, and likely represents cell walls and other apoplastic material. Water uptake into the medium and slow pools was slowed by salt addition, with the medium pool thought to be cellular, while the slow pool is presumably related to endosperm hydration. Salt caused a minor decrease in the water uptake rates and maximum seed water content, while PEG strongly suppressed both parameters. Seeds transferred between water and salt solutions followed the water uptake trajectories of the solution into which they were transferred. Seeds transferred from PEG to water achieved final seed water contents similar to water control seeds, while seeds transferred from water to PEG achieved significantly higher final water contents than PEG controls. This work confirms that salt and PEG have distinct effects on water uptake by wheat seeds.
1. Introduction
The germination rate of non-dormant seeds is controlled by multiple factors, with water availability generally considered to be one of the most important. Reduced water availability, for example in experiments utilizing PEG as an osmoticum, or in dry soils, reduce water uptake and seed germination rates [1], with stronger solutions / drier soils repressing germination rate to a greater degree [2]. Despite a general understanding of the link between reductions in water uptake and germination, a detailed physiological explanation of the mechanism(s) remains elusive. Plant hormones including abscisic acid (ABA) [3], cytokinins, gibberellins and Indole-3-acetic acid (IAA) have been suggested to have a role [4], although it is unclear whether these are a cause of, or a response to, changes in seed water content.
As with PEG, salt treatment generally reduces germination percentages and rates, relative to water controls. Salt is reported to affect seed germination by osmotic effects, ion toxicity effects, or a combination of the two [4,5,6]. Differences in the germination rates and percentages between seeds incubated in salt and PEG are generally attributed to “ionic effects”, which can be either negative or positive. In some halophytic (salt-tolerant) species, incubation in high salt concentrations can arrest germination, although germination percentages can often recover when these seeds are transferred to pure water [7]. It is unclear whether arrested germination is directly due to osmotic effects, or to some non-toxic germination repression caused by the salt, although it has been noted that high concentrations of Na+ and Cl− ions can inhibit critical metabolic steps in cell division [8]. In some species, moderate salt levels have been shown to promote germination [4], and it has been suggested that salt can be compartmentalized and used as a cellular osmotica, allowing seeds to germinate under osmotic conditions which would otherwise preclude it [9,10]. Compartmentalization of salt into vacuoles has been shown to aid the growth and survival of vegetative plants [11,12].
The most widely used germination model, the hydrothermal time model, posits a base water potential designated Ψb (50), at which half of the viable seeds in a population cannot germinate. The physiological basis of Ψb remains unclear, yet is presumably related to the seeds ability to take up water. While both salt and PEG are able to exert osmotic effects, their effect are often not identical. As PEG is either unable to cross cell membranes, or can do so only slowly, it tends to act purely as an osmotica. Conversely, as mentioned above, at least in some species, salt can be taken up and compartmentalized, allowing seeds to avoid the negative consequences of low water potential environments. Few studies have attempted to directly quantify the kinetics of seed water uptake and relate it to germination mechanics. McDonald et al. [13] measured water uptake by maize seeds over a 48-h period, demonstrating the existence of two separate pathways of water uptake. The first pathway was rapid, and related to water uptake into the embryo, while the second involved a more gradual wetting of the endosperm, likely associated with starch breakdown. Other studies have used imaging techniques to investigate water uptake by seeds. Kikuchi et al. [14] noted rapid uptake of water into the testa and cotyledons of kidney beans following a lag period of a few hours. Munz et al. [15] revealed that water uptake followed a coordinated series of steps featuring multiple pools in oilseed rape seeds.
The main aim of this paper is to gain a better understanding of water uptake kinetics by seeds, particularly in relation to environmental water potential. Kinetic studies are frequently used to provide information about the regulation of complex systems, for example, enzymatic properties, biochemical pathways or sink / source relationships between plant organs. Here, we measured water uptake by wheat seeds and developed a mathematical model, which allowed us to compare the effects of salt and PEG on the rate of water uptake into different pools. Wheat (Triticum aesitvum L.) seeds were used due to their relatively large size, which allowed reliable estimates of water uptake rates based upon mass increase through time. Water uptake rates and contents were compared between seeds incubated in pure water, a −1MPa NaCl solution, and a −1 MPa PEG solution, both under constant conditions, and where the osmotic conditions were changed part-way through the experiment. Given the assumption that seeds can take up salt but not PEG, we hypothesized that water uptake rates would be lower in seeds incubated in PEG than in either the water or salt treatments, and that this would have a detectable effect on water uptake kinetics. We also hypothesized that when seeds were transferred between treatments, the initial treatment conditions would influence the final water content of the seeds.
2. Materials and Methods
All experiments described in this study used seeds from a single seed batch of a locally developed commercially available wheat variety, “Aoba”. Seeds between 40 and 50 mg were used, this representing approximately the middle 50th percentile, with larger and smaller seeds excluded.
Osmotica were made using either PEG 6000 or NaCl dissolved in milliQ grade water. PEG solutions were prepared according to Equation (5) from Michel [16] (Equation (1), below), and checked using a WP4 Dewpoint Osmometer (Decagon, Pullman, WA, USA).
where [PEG] is the concentration of PEG required (g g−1 H2O) to give a desired water potential (Ψ), at a given temperature (T).
Salt solutions were made according to Equation (2), which was derived from a dataset of measured values of water potential in relation to salt concentration at a range of temperatures. The dataset measurements were found to correspond well with published values [17].
where [NaCl] was the concentration of salt (g L−1) required to produce a solution of a desired water potential (Ψ) at 5 °C.
Seeds were placed equidistant from each other in a grid pattern on Whatmann 3 mm CHR chromotography paper wetted with the test solution of interest and sealed in a 15 cm diameter petri-dish. A small volume of extra solution was placed in each container, to reduce matric effects caused by the filter paper, and to buffer the system against water uptake by the seeds. Additional solution was added, or the chromatography paper/solution replaced on an “as necessary” basis. It was calculated that water uptake by seeds would be able to alter the concentration of the solution by less than 3%, and the petri dishes were sealed to minimize evaporative losses. Changing old solution for fresh solution had little impact on seed water content, confirming that the osmotic potentials of the solutions did not change greatly throughout the experimental period. Each experiment was conducted only once, but they were part of a larger series of trails, with water uptake patterns consistent across all experiments.
2.1. Experiment 1
This experiment aimed to compare the water uptake kinetics of killed seeds, where the embryo had been destroyed with a hot needle, versus live seeds (Figure 1). Killed seeds are preferable for measuring water uptake as they do not germinate, allowing longer water uptake time courses to be obtained. Thirty-two live and 32 killed seeds were used in each trial, which was conducted at 14 °C under a range of PEG-generated water potentials (0, −0.23, −0.35, −0.47. −0.58, −0.77, −1.05, −1.42, −1.80, −2.18 and −2.46 MPa) to promote a range of water uptake rates in the seeds. At the start of the experiment, then 3, 6, 9, 12, 20, 27, 36, 45, 56, 69 and 84 h after the start of incubation, individual seed fresh weight was recorded to the nearest 0.1 mg. After 12, 27, 45, and 69 h, six replicate seeds were removed from each treatment, weighed, dried and re-weighed. After 84 h the remaining eight seeds for each treatment were weighed and the experiment terminated. Ten additional control seeds were weighed then dried at 80 °C for seven days and re-weighed, without being immersed in any test solution. The dry mass of the test seeds (approximately totalling 750) was compared with the dry mass of the control seeds, with no significant differences in mass noted between seed lots, suggesting no loss of mass due to respiration. In subsequent experiments it was therefore assumed that seed dry weight remained relatively constant throughout the experiments, and it was not further tested.
2.2. Experiment 2
This experiment had two goals. The first was to characterize water uptake kinetics in wheat seeds incubated in deionized water, −1 MPa NaCl and −1MPa PEG solutions. This water potential was selected based upon a pre-experiment, where we observed that a −1 MPa water potential was sufficient to cause a reduction in seed germination rate, and in the case of PEG, to cause a significant reduction in final seed water content. The second goal was to investigate the effects on water uptake of transferring the seeds between either of the osmotica and deionized water, after 8 or 96 h. These time points were chosen as they represent the seeds taking water up into pool 2 and pool 3, thought to be the embryo and endosperm respectively. Killed seeds were used in this experiment, with 12 seeds per treatment. Seeds were individually weighed prior to the start of treatments, then periodically throughout the treatment period in order to estimate water uptake. Measurement times were adjusted to try and capture detailed information about the curve. For example, being more frequent when water uptake was most rapid or immediately following transfer between treatments, and less frequently when water uptake was less rapid. The exact time points can be found in the associated dataset. Each seed was dried using tissue paper, weighed to the nearest 0.1 mg, and returned to the same solution. This experiment was conducted at 5 °C, with the controls (i.e., seeds maintained in water, NaCl or PEG for the whole experiment, without being transferred) incubated for 338 h, while the reciprocal transfer treatments were terminated after 314 h.
2.3. Data Analysis
All seed water contents (SWC) were expressed as mg of water per mg of the initial seed mass (SM). Experiment one showed that the initial water content of the seeds was very uniform, allowing us to ignore this without significantly affecting the results.
We used an exponential-rise-to maximum curve framework to model water uptake, following Finch-Savage et al. [1]. This model type, with a single exponential rise-to-max curve shown in Equation (3), can be easily understood in mechanistic terms, with water passing into a single pool with a max (m), over time (t), at a rate determined by the rate constant (K), proportional to the seed water content (SWC).
In terms of the underlying biology, this type of model often describes diffusional processes, particularly where a membrane is involved.
Equation (4) shows a double exponential model, with two pools, each with a maximum value (m1 and m2), and a rate constant (K1 and K2). Within our context, we may understand these as “fast” and “slow” pools, respectively.
Equation (5) shows a triple exponential model, representing three pools, each with maxima and rate constants.
More complex exponential model implies multi-phasic water uptake, with water being absorbed into two or more pools. The maximum seed water content is the sum of m-values for each pool, while the rate of water flux into each pool would be described by a linear relationship between the seed water content and time, with the rate constant being the gradient of this relationship.
Curve fitting was conducted in PRISM v9.1.2 (Graphpad software, San Diego, CA, USA), using a least squares regression method. Normality of a residuals was tested by Shapiro–Wilk test, while a Runs test was used to test for systemic lack of fit between the curve and the data. The Extra-sum-of-squares F-test was used to determine which curve to fit. Briefly, the extra-sum of squares test compares whether a more complex model fits the data better than can be explained purely from increased model complexity, compared to a simpler model.
Seed water uptake rate was estimated as the mass difference between two adjacent time points (or curve-fit predictions), divided by the time difference. The seed water content was estimated as the mean seed water content of any two adjacent time points.
Differences in seed water content for different treatments were compared using the GLM command, with a post-hoc REGWF test to identify group differences in IBM SPSS v.27 (IBM corporation, Armonk, NY, USA). Data was tested for heteroskedasticity and log-transformed where necessary.
3. Results
3.1. Comparison of Water Uptake in Live vs. Killed Seeds
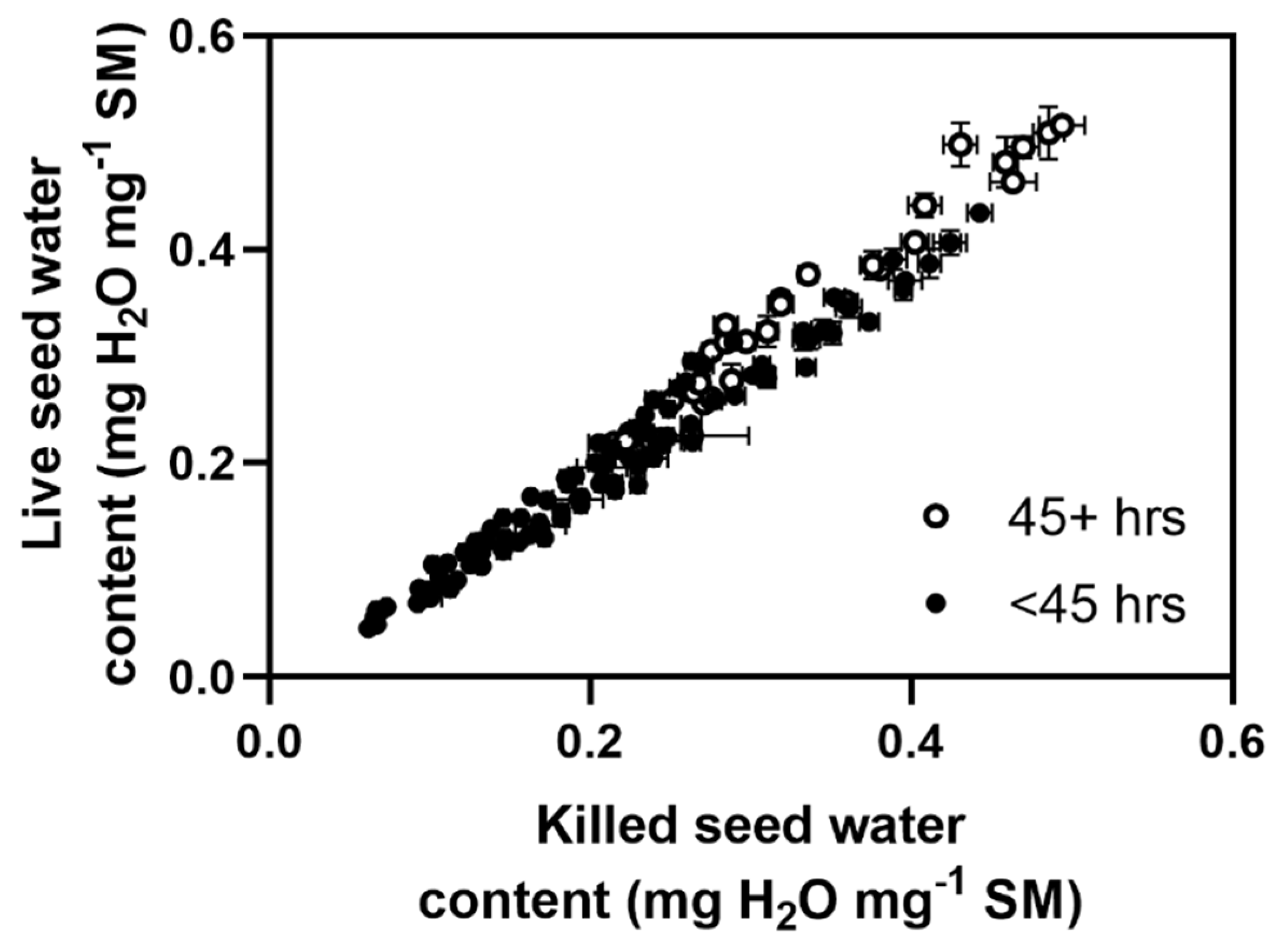
The slope of the relationship between killed and live seed water contents did not differ from 1 for periods less than 45 h (Figure 2; F(1,102) = 0.436, p = 0.511). Over 45 h, the gradient was 1.11, which was determined to be significantly greater than 1 (F(1,29) = 6.374, p = 0.017). It was assumed that this increase in gradient was due to water uptake to tissues in live seeds which had been damaged or destroyed in the killed seeds. Despite this, the relationship between SWC in live and killed seeds was very strong, and it was concluded that killed seeds would be a suitable model for understanding water uptake processes. We did not test the effect of salt on water uptake rates in this experiment, and it is an assumption that salt would not affect this relationship.
3.2. Determination of a Suitable Model
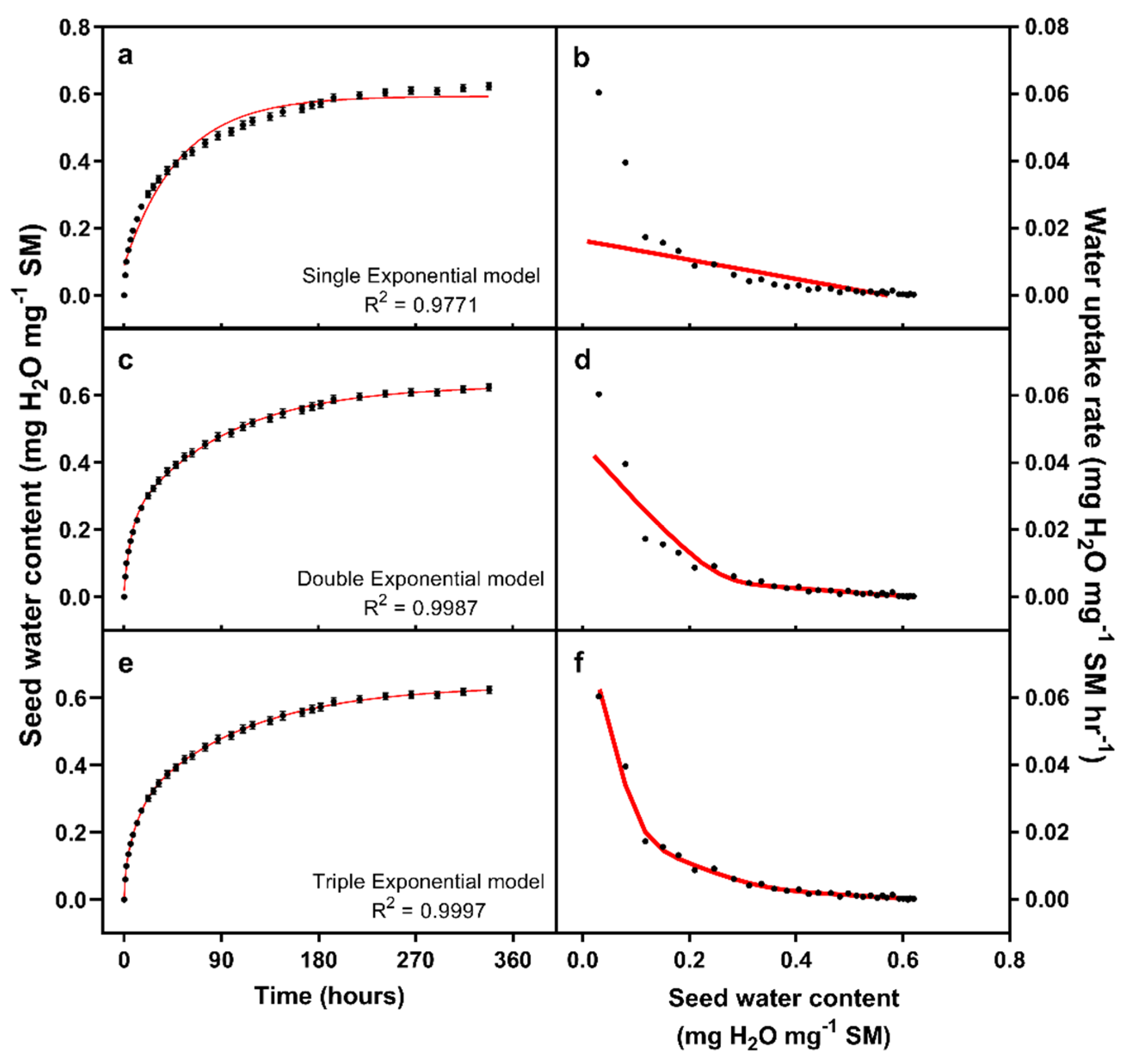
Our next goal was to fit a model to water uptake of the water control seeds of experiment two. Following Finch-Savage et al. [1], we initially tried to fit an exponential model rising to a maximum value, although significant lack of fit was evident, and the model failed a Runs test (Figure 3a; Table 1). This was particularly obvious when we plotted water uptake rate against seed water content (Figure 3b). The data suggested the existence of a second pool, with a comparison of model fits between the single and double exponential models favouring the more complex model (Figure 3c; p < 0.001). However, lack of fit could be found in the plot of water uptake against water content (Figure 3d), with the two-pool model continuing to fail a Runs test. We speculated the existence of a third pool, with the three-pool model fitting the data significantly better than would be expected purely as a result of increased model complexity (Figure 3e; p < 0.001). No lack of fit could be observed at this point (Figure 3f), and the model could pass both Shapiro–Wilk tests for the normality of the residuals and a Runs test. Given these conditions, we did not consider any more complex models.
3.3. Water Uptake under Constant Conditions
Seed water content increased most rapidly in seeds incubated in water, and slowest in seeds incubated in a −1 MPa PEG solution (Figure 4a). SWC of the water and NaCl treated seeds could be described by a triple-exponential curve, while the PEG treatment was best described by a double-exponential curve (Table 2).
Curve fits for the water and NaCl seeds suggested the presence of a fast pool, contributing between 11 to 13% of total capacity, a medium pool, contributing 28 to 33% and a slow pool of approximately 54 to 60% of total capacity. In the PEG seeds, a fast pool of approximately 26% of seed water content and a slow pool of 74% were identified. The sizes of each individual pool could be calculated by multiplying the maximum SWC by the relative contribution of that pool to the total. The fast pools were approximately the same size in each case, yielding values of 0.071, 0.079 and 0.069 mg H2O mg−1 SM for water, salt and PEG seeds respectively. The medium pools gave values of 0.181 and 0.199 mg H2O mg−1 SM for water and salt. In PEG seeds, the slow pool had a value of 0.195 which was similar to the medium pool for the other two treatments, suggesting a similar mechanism.
The rate constants of the medium pool were an order of magnitude slower than the fast pool in the water and NaCl treatments, and the slow pool an order of magnitude slower than the medium pool. For the PEG seeds, water uptake into the fast pool was around 16 times faster than into the slow pool.
The confidence limits for the rate constants for the fast pool overlapped between the water and NaCl treatments, while the rate constants for the NaCl treatment were significantly lower in the medium and slow pools, suggesting a negative effect of salt on water uptake. For the PEG treated seeds, the rate constant of the fast pool was significantly lower than the water treated seeds, but not different from the NaCl treated seeds. The rate constant for the slow pool was of the same order of magnitude, but significantly lower than for the medium pool of either the water or NaCl treatments. Comparing the plot of water uptake rates versus seed water content, we can see a high degree of similarity between the water and salt treatments, while the PEG curve was markedly different from the other two (Figure 4b).
3.4. Water Uptake in Response to Changing Conditions
When the seeds were transferred between the water and salt solutions (Figure 5a,c), they tended to quickly adjust and follow the water uptake curve for the solution they were transferred to, both when transferred after 8 or 96 h. Seeds transferred from water to PEG after 8 h had a greatly reduced subsequent water uptake (Figure 5b), while those transferred after 96 h lost water to the environment (Figure 5d). Transfer from PEG to water after 8 h caused an increase in water uptake for around 10 h, after which the seeds followed the control water curve (Figure 5b, inset). Seeds transferred from PEG to after 96 h gained water rapidly, with this additional water uptake described by a double-exponential curve (Figure 6).
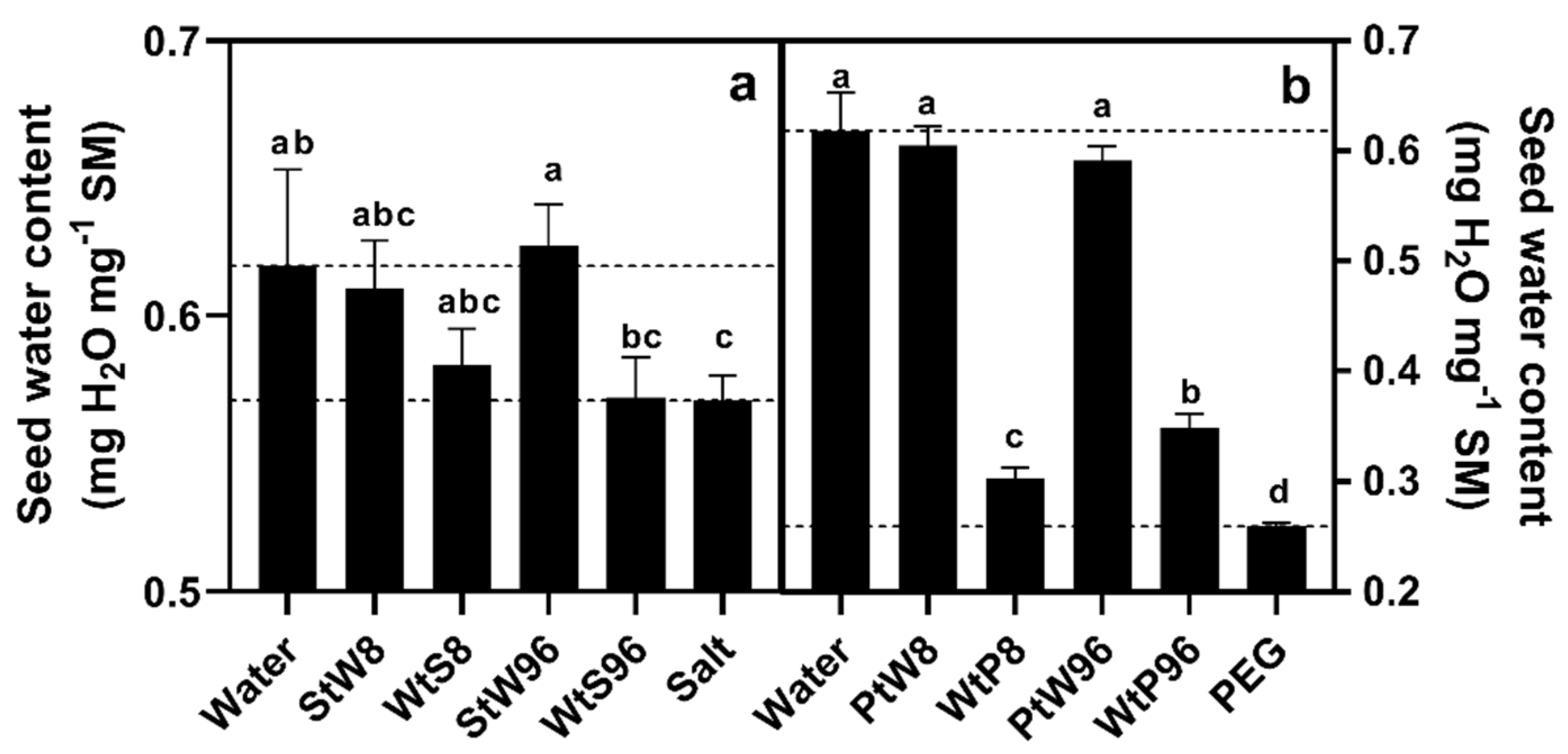
The SWC at 314 h was lower in salt controls than water controls (Figure 7a). Seeds switched between salt and water treatments had SWC values which did not differ from the controls of the solutions they were transferred into. This was clearer in the seeds transferred after 96 h, with the seeds transferred after 8 h being intermediate two both the salt and water controls. The SWC at 314 h was lower in PEG controls than water controls (Figure 7b). While the SWC of seeds transferred from PEG to water did not differ from water controls, the SWC of seeds transferred from water to PEG after 96 h was 134.4% of PEG controls, while those transferred after 8 h had values 116.9% of PEG controls.
4. Discussion
Our data suggests the existence of three distinct pools into which water is absorbed by wheat seeds. The existence of multiple pools and multiphasic water uptake is consistent with previous studies in wild oat [18], maize [13] and soybean [19]. The first pool is the smallest-comprising around 11–13% of the total capacity. It has a very high-rate constant, filling in just a few hours. This first pool is relatively unaffected by salt, but absorption of water was slowed by the PEG solution. It seems likely that this pool represents apoplastic components of the seed, such as cell walls [20]. It has been estimated that the water potential of seeds can be as low as −400 MPa [21], which is mainly considered to be attributable to matric effects.
The second pool comprises around 30% of the total capacity. The rate constant is around one-tenth that of the first pool, with a half-life of around eight hours in the water control treatment. Under optimal conditions, wheat and barley seeds can germinate in as little as 24 h, at which time we would expect the first pool to be completely saturated, and this second pool at 80–90% of its maximum capacity. This pool is seemingly weakly affected by salt, with the half-life increasing by around 50%, but no change in its maximum size. Given the influence of salt on water uptake to this pool, this suggests a more biological than physical explanation, with water crossing a membrane over which salt can slowly cross. We assume this pool to be cellular. Although the embryo is relatively small, accounting for only around 3% of the mass of the seed [20], it has been shown to take up several times its own mass when incubated in water [22].
The third pool is the largest, and slowest to fill. Its rate constant is an order of magnitude lower than for the second pool, taking 70 h to half-fill in the water controls, and nearly 100 h in the salt treatment. The rate constant is affected by salt, suggesting this to be a biological phenomenon, which we assume to be starch breakdown. Previous investigators have noted a reduction in the rate of α-amylase-mediated starch degradation to soluble sugars when seeds are incubated in saline solutions [23,24,25], with these sugars acting as osmotica.
The water uptake curve of the PEG incubated seeds was best described by a double-exponential model. The maximum water content achieved by the PEG treatment was 41.4% that of the water control seeds. The rate constants were the same order of magnitude as those of the fast and medium pools identified in water control seeds, yet the values were lower than those for salt or water controls.
Switching seeds between the water and salt treatments led to the seeds following the water uptake curve of the solution they were exposed to. This was particularly clear in the seeds switched after 96 h, with the water to salt seeds achieving a maximum water content lower than the water controls and not significantly different with the salt controls. The salt to water seeds similarly achieved a maximum water content not different from the water controls, and significantly higher than the salt controls. In studies where germination is suppressed by salt, germination percentages often recover after the seeds are transferred to pure water [7,26]. This is often used to conclude that the retardation of germination is caused by osmotic effects. For this conclusion to be valid, we would expect to see similar reduction in germination percentage where PEG solutions were used. Zhang et al. [10] showed in two barley genotypes that germination percentages were more strongly suppressed by PEG than salt, concluding that both salt and water were being taken up by the seeds. This corresponds strongly with our study, where NaCl incubated seeds achieved final water contents only 4.8% lower than water control seeds, suggesting that any osmotic effects are very mild. An alternate plausible explanation for salt suppression of germination may be that osmotic factors are relatively unimportant, with reductions in germination being a direct result of salt entering the embryo, which has been shown to inhibit seed germination at low concentrations and kill the seed at high concentrations [27]. Transfer to pure water would presumably allow salt export from the seed.
Switching between water and PEG had a stronger effect. Transfer from water to PEG resulted in final water contents significantly higher than PEG controls, with this increase significantly greater in the 96-h transfer than the 8-h transfer seeds. This suggests that some changes occurred in the seeds during the period they were incubated in water, which could not be undone by the PEG solution. Particularly in the 96-h seeds, it seems evident that the third water pool is involved in this change, suggesting that starch breakdown accounts for this phenomenon. Previous studies have reported that seed priming-soaking seeds in water for between 10 and 24 h prior to planting leads to improvements in germination percentages and plant establishment, as well as the subsequent vigour, disease resistance and ultimately yield of crop plants in semi-arid tropical areas [1,28]. It seems plausible that improved water and carbohydrate availability during germination and seedling establishment may at least partially explain these observations [29].
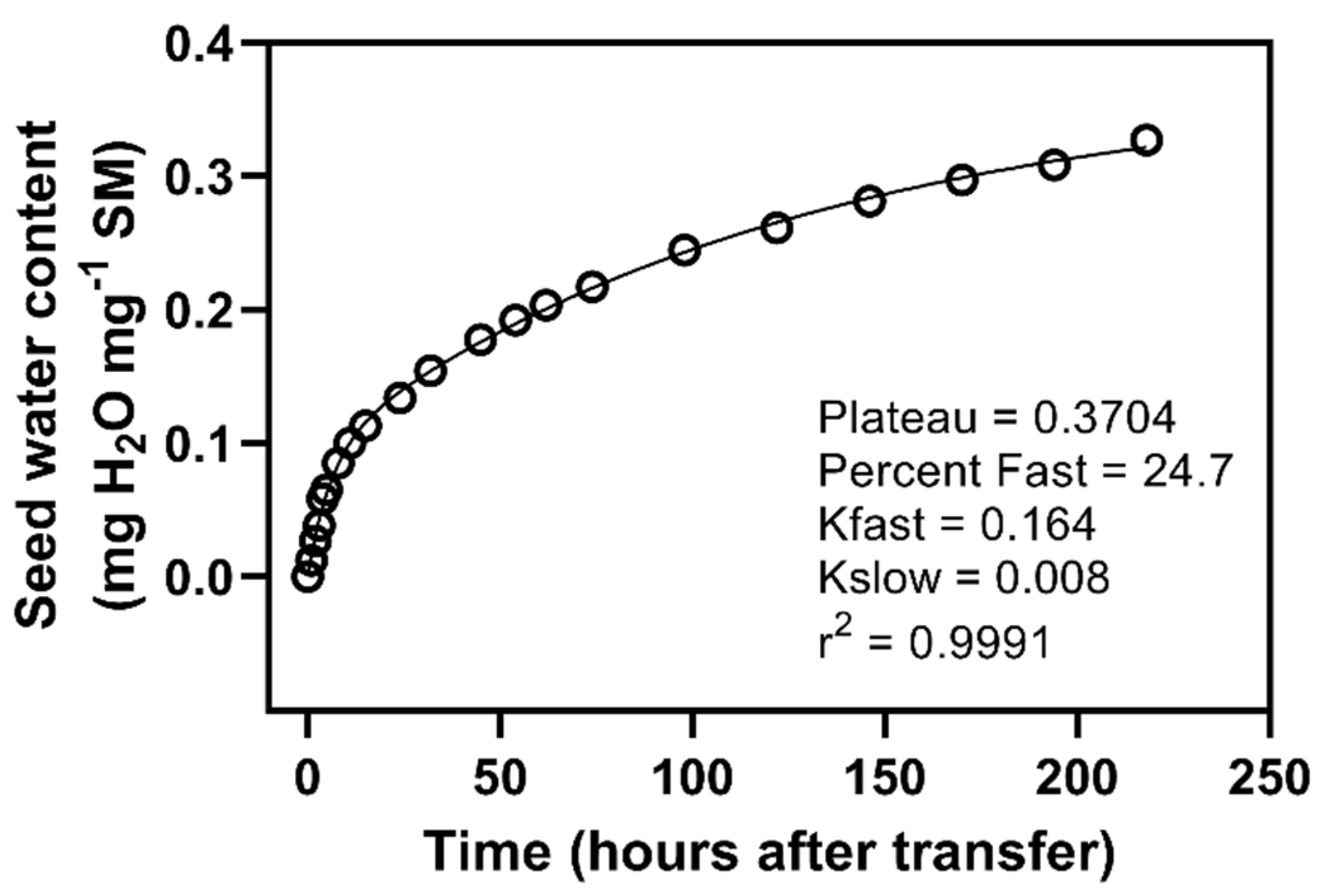
Transferring from PEG to water led to the seeds achieving maximum water contents not different from water control seeds. Isolating post-transfer water uptake data in the 96-h seeds allowed curve fitting, and an investigation of water uptake kinetics (Figure 6). A double-exponential curve fitted the data well, with half-lives of around 4 and 86 h for the two pools. The faster pool represented around 25% of the total water uptake. The use of a double-exponential fit to this data suggests the existence of two pools. The larger of these pools has a rate constant (8.01 × 10−3) consistent with the slow pool of water control seeds, which we hypothesize to be related to starch breakdown. The smaller pool has a rate constant (1.64 × 10−1) intermediate between pools one and two, although more strongly influenced by the second. Our data appears to suggest that some amount of starch breakdown occurs even under the PEG control conditions, although water uptake was suppressed.
Within the context of our experiment, water uptake rates may be described by changes in the rate constants and sizes of the three pools. PEG appears to have a strong ability to alter both water uptake rate and the maximum water content, while salt caused declines in the rate constants of the second and third pool, with the maximum water content less affected. This may explain the greater germination percentages achieved in iso-osmotic salt solutions compared with PEG solutions [10]. Our data furthermore suggests the existence of a minimum water content, under which the seeds cannot germinate. Speculatively, seed to seed variation may be explained by differences in factors which control water uptake, such as seed size, membrane permeability, or the activities of key enzymes.
In conclusion, using killed seeds, we characterised water uptake into at least three pools, comprising approximately 11%, 28%, and 60% of the total water uptake capacity, with half-lives of 0.7, 8, and 70 h in water, respectively. Incubating seeds in a −1 MPa salt solution slowed the rate of water uptake into the second and third pools but caused only a small decline in maximum water content. When the seeds were incubated in −1 MPa PEG, water uptake rates and the maximum water content were strongly suppressed. Transferring seeds between water and salt solutions had only a small effect on water uptake, while transferring between water and PEG had strong effects on water uptakes. Notably, seeds incubated in water then transferred to PEG exhibited maximum seed water contents higher than PEG controls, which may explain the benefits of pre-soaking seeds before planting.
Author Contributions
Conceptualization, L.J.I. and H.Z.; methodology, L.J.I.; software, L.J.I.; validation, L.J.I.; formal analysis, L.J.I.; investigation, L.J.I.; resources, L.J.I.; data curation, L.J.I.; writing—original draft preparation, L.J.I.; writing—review and editing, H.Z.; visualization, L.J.I.; project administration, L.J.I.; funding acquisition, L.J.I. All authors have read and agreed to the published version of the manuscript.
Funding
The data collection for this research was funded by a JSPS post-doctoral fellowship to L.J.I., grant number PO8097.
Data Availability Statement
The dataset for this study is publicly available at https://0-doi-org.brum.beds.ac.uk/10.5281/zenodo.5115478, accessed on 15 July 2021.
Acknowledgments
L.J.I would like to thank Yuu Hariya and Izumi Takagi (Mathematical Institute, Tohoku University) for advice on mathematical modelling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Finch-Savage, W.E.; Rowse, H.R.; Dent, K.C. Development of combined imbibition and hydrothermal threshold models to simulate maize (Zea mays) and chickpea (Cicer arietinum) seed germination in variable environments. New Phytol. 2005, 165, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, V.; Bradford, K.J. A hydrothermal time model explains the cardinal temperatures for seed germination. Plant Cell Environ. 2002, 25, 1061–1069. [Google Scholar] [CrossRef]
- Gendreau, E.; Romaniello, S.; Barad, S.; Leymarie, J.; Benech-Arnold, R.; Corbineau, F. Regulation of cell cycle activity in the embryo of barley seeds during germination as related to grain hydration. J. Exp. Bot. 2008, 59, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungar, I.A. Halophyte seed germination. Bot. Rev. 1978, 44, 233–264. [Google Scholar] [CrossRef]
- Sosa, L.; Llanes, A.; Reinoso, H.; Reginato, M.; Luna, V. Osmotic and specific ion effects on the germination of Prosopis strombulifera. Ann. Bot. 2005, 96, 261–267. [Google Scholar] [CrossRef]
- Welbaum, G.E.; Tissaoui, T.; Bradford, K.J. Water relations of seed development and germination in muskmelon (Cucumis melo L.): III. Sensitivity of germination to water potential and abscisic acid during development. Plant Physiol. 1990, 92, 1029–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Feng, G.; Zhang, F. Salinity and temperature effects on germination for three salt-resistant euhalophytes, Halostachys caspica, Kalidium foliatum and Halocnemum strobilaceum. Plant Soil 2006, 279, 201–207. [Google Scholar] [CrossRef]
- Neumann, P. Salinity resistance and plant growth revisited. Plant Cell Environ. 1997, 20, 1193–1198. [Google Scholar] [CrossRef]
- Pujol, J.A.; Calvo, J.F.; Ramírez-Díaz, L. Seed germination, growth, and osmotic adjustment in response to NaCl in a rare succulent halophyte from southeastern Spain. Wetlands 2001, 21, 256–264. [Google Scholar] [CrossRef]
- Zhang, H.; Irving, L.J.; McGill, C.; Matthew, C.; Zhou, D.; Kemp, P. The effects of salinity and osmotic stress on barley germination rate: Sodium as an osmotic regulator. Ann. Bot. 2010, 106, 1027–1035. [Google Scholar] [CrossRef]
- Fukuda, A.; Nakamura, A.; Tagiri, A.; Tanaka, H.; Miyao, A.; Hirochika, H.; Tanaka, Y. Function, intracellular localization and the importance in salt tolerance of a vacuolar Na+/H+ antiporter from rice. Plant Cell Physiol. 2004, 45, 146–159. [Google Scholar] [CrossRef] [Green Version]
- Garthwaite, A.J.; von Bothmer, R.; Colmer, T.D. Salt tolerance in wild Hordeum species is associated with restricted entry of Na+ and Cl− into the shoots. J. Exp. Bot. 2005, 56, 2365–2378. [Google Scholar] [CrossRef]
- McDonald, M.; Sullivan, J.; Lauer, M.J. The pathway of water uptake in maize seeds. Seed Sci. Technol. 1994, 22, 79–90. [Google Scholar]
- Kikuchi, K.; Koizumi, M.; Ishida, N.; Kano, H. Water uptake by dry beans observed by micro-magnetic resonance imaging. Ann. Bot. 2006, 98, 545–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munz, E.; Rolletschek, H.; Oeltze-Jafra, S.; Fuchs, J.; Guendel, A.; Neuberger, T.; Ortleb, S.; Jakob, P.M.; Borisjuk, L. A functional imaging study of germinating oilseed rape seed. New Phytol. 2017, 216, 1181–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, B.E. Evaluation of the water potentials of solutions of polyethylene glycol 8000 both in the absence and presence of other solutes. Plant Physiol. 1983, 72, 66–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochrane, T.T.; Cochrane, T.A. Osmotic potential properties of solutes common in the soil-plant solution continuum. Soil Sci. 2005, 170, 433–444. [Google Scholar] [CrossRef]
- Hou, J.Q.; Kendall, E.J.; Simpson, G.M. Water uptake and distribution in non-dormant and dormant wild oat (Avena fatua L.) caryopses. J. Exp. Bot. 1997, 48, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Koizumi, M.; Kikuchi, K.; Isobe, S.; Ishida, N.; Naito, S.; Kano, H. Role of seed coat in imbibing soybean seeds observed by micro-magnetic resonance imaging. Ann. Bot. 2008, 102, 343–352. [Google Scholar] [CrossRef]
- Laddomada, B.; Caretto, S.; Mita, G. Wheat bran phenolic acids: Bioavailability and stability in whole wheat-based foods. Molecules 2015, 20, 15666–15685. [Google Scholar] [CrossRef]
- Shaykewich, C.F. Proposed method for measuring swelling pressure of seeds prior to germination. J. Exp. Bot. 1973, 24, 1056–1061. [Google Scholar] [CrossRef]
- Dell’aquila, A. Water uptake and protein synthesis in germinating wheat embryos under the osmotic stress of polyethylene glycol. Ann. Bot. 1992, 69, 167–171. [Google Scholar] [CrossRef]
- Almansouri, M.; Kinet, J.M.; Lutts, S. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant Soil 2001, 231, 243–254. [Google Scholar] [CrossRef]
- Bouaziz, A.; Hicks, D.R. Consumption of wheat seed reserves during germination and early growth as affected by soil water potential. Plant Soil 1990, 128, 161–165. [Google Scholar] [CrossRef]
- Lin, C.C.; Kao, C.H. NaCl stress in rice seedlings: Starch mobilization and the influence of gibberellic acid on seedling growth. Bot. Bull. Acad. Sin. 1995, 36, 169–173. [Google Scholar]
- Zhang, H.; Zhang, G.; Lü, X.; Zhou, D.; Han, X. Salt tolerance during seed germination and early seedling stages of 12 halophytes. Plant Soil 2015, 388, 229–241. [Google Scholar] [CrossRef]
- Zhang, H.-X.; Tian, Y.; Zhou, D.-W.; Zheng, W.; Wang, M.-L. Research on modeling germination response to salinity of barley seeds. Chin. J. Plant Ecol. 2013, 36, 849–858. [Google Scholar] [CrossRef]
- Harris, D.; Joshi, A.; Khan, P.A.; Gothkar, P.; Sodhi, P.S. On-farm seed priming in semi-arid agriculture: Development and evaluation in maize, rice and chickpea in India using participatory methods. Exp. Agric. 1999, 35, 15–29. [Google Scholar] [CrossRef]
- Carrillo-Reche, J.; Newton, A.C.; Quilliam, R.S. Using seed respiration as a tool for calculating optimal soaking times for ‘on-farm’ seed priming of barley (Hordeum vulgare). Seed Sci. Res. 2021, 31, 116–124. [Google Scholar] [CrossRef]
Figure 1.
Killed (left) and live (right) wheat seeds. In the killed seed, the embryo was killed using a soldering iron.
Figure 1.
Killed (left) and live (right) wheat seeds. In the killed seed, the embryo was killed using a soldering iron.

Figure 2.
Water content of live vs seeds killed with a hot needle. Filled circles represent measurements of seeds within the first 45 h of incubation, while opened circles represent measurements made after 45 h. Error bars represent standard errors.
Figure 2.
Water content of live vs seeds killed with a hot needle. Filled circles represent measurements of seeds within the first 45 h of incubation, while opened circles represent measurements made after 45 h. Error bars represent standard errors.

Figure 3.
Seed water uptake kinetics fitted with a single (a), double (c), or triple (e) exponential curve. Seed water uptake rates were calculated and plotted against seed water content, for the data (points), and the regression models (red lines, Figures (b,d,f)).
Figure 3.
Seed water uptake kinetics fitted with a single (a), double (c), or triple (e) exponential curve. Seed water uptake rates were calculated and plotted against seed water content, for the data (points), and the regression models (red lines, Figures (b,d,f)).

Figure 4.
(a) Water uptake curves for seeds incubated in water (red curve), −1 MPa NaCl (blue), or −1MPa PEG (green) solutions. (b) rate of seed water uptake plotted against seed water content.
Figure 4.
(a) Water uptake curves for seeds incubated in water (red curve), −1 MPa NaCl (blue), or −1MPa PEG (green) solutions. (b) rate of seed water uptake plotted against seed water content.

Figure 5.
Seed water content curves for seeds transferred between water and salt (a,c) or water and PEG (b,d) solutions after 8 (a,b) or 96 (c,d) h. Red, blue, and green lines represent the time course for water, salt, and PEG controls respectively. Filled symbols represent seeds transferred from water to NaCl or PEG solutions, while open symbols represent seeds transferred from NaCl or PEG to water. Insets show details of water contents immediately around the time of transfer.
Figure 5.
Seed water content curves for seeds transferred between water and salt (a,c) or water and PEG (b,d) solutions after 8 (a,b) or 96 (c,d) h. Red, blue, and green lines represent the time course for water, salt, and PEG controls respectively. Filled symbols represent seeds transferred from water to NaCl or PEG solutions, while open symbols represent seeds transferred from NaCl or PEG to water. Insets show details of water contents immediately around the time of transfer.

Figure 6.
Isolated seed water uptake curve for seeds transferred from PEG to water after 96 h. A double-exponential curve has been fitted with the parameters reported.
Figure 6.
Isolated seed water uptake curve for seeds transferred from PEG to water after 96 h. A double-exponential curve has been fitted with the parameters reported.

Figure 7.
Seed water content after 314 h, for control seeds (Water, NaCl (a), PEG (b)) and seed transferred from water to salt (WtS), salt to water (StW), water to PEG (WtP) or PEG to water (PtW) after 8 or 96 h.
Figure 7.
Seed water content after 314 h, for control seeds (Water, NaCl (a), PEG (b)) and seed transferred from water to salt (WtS), salt to water (StW), water to PEG (WtP) or PEG to water (PtW) after 8 or 96 h.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Curve-fit parameters of single, double, and triple exponential models fitted to water uptake data for water control seeds in Figure 3. Numbers in parentheses represent the 95% confidence intervals of the estimates. An extra-sum-of-squares F-test was used to determine whether a more complex model fit the data significantly better than the immediately simpler model.
Table 1.
Curve-fit parameters of single, double, and triple exponential models fitted to water uptake data for water control seeds in Figure 3. Numbers in parentheses represent the 95% confidence intervals of the estimates. An extra-sum-of-squares F-test was used to determine whether a more complex model fit the data significantly better than the immediately simpler model.
| Single | Double | Triple | |
|---|---|---|---|
| Plateau | 0.5714 (0.5446–0.5998) | 0.6264 (0.6158–0.6384) | 0.6367 (0.6309–0.6431) |
| Percent fast | 33.35 (30.31–36.45) | 11.09 (8.82–13.59) | |
| Percent medium | 66.65 | 28.48 (26.13–30.61) | |
| Percent slow | 60.43 | ||
| Kfast (×10−1) | 2.86 (2.31–3.58) | 1.97 (1.59–2.49) | 9.64 (6.86–14.83) |
| Kmedium (×10−2) | 1.16 (1.03–1.30) | 8.67 (7.17–10.38) | |
| Kslow (×10−3) | 9.90 (9.19–10.57) | ||
| Fast half-life (hours) | 24.26 (19.37–30.05) | 3.53 (2.78–4.36) | 0.719 (0.467–1.01) |
| Medium half-life (hours) | 59.61 (53.53–67.05) | 8.00 (6.68–9.74) | |
| Slow half-life (hours) | 70.0 (65.6–75.4) | ||
| F-value (d.f.) | 417.4 (2, 28) | 108.5 (1, 26) | |
| F-test p-value | <0.001 | <0.001 | |
| Better model | Double-exponential | Triple-exponential | |
| R2 value | 0.9452 | 0.9982 | 0.9997 |
| Shapiro-Wilk p-value | 0.0528 (passed) | 0.1812 (passed) | 0.3504 (passed) |
| Runs test p-value | <0.0001 (failed) | 0.0005 (failed) | 0.6424 (passed) |
Table 2.
Curve-fit parameters for water uptake time course data for seeds incubated in water, NaCl or PEG solutions in Figure 4. Numbers in parentheses represent the 95% confidence intervals of the estimates. An extra-sum-of-squares F-test was used to determine whether a more complex model fit the data significantly better than the immediately simpler model.
Table 2.
Curve-fit parameters for water uptake time course data for seeds incubated in water, NaCl or PEG solutions in Figure 4. Numbers in parentheses represent the 95% confidence intervals of the estimates. An extra-sum-of-squares F-test was used to determine whether a more complex model fit the data significantly better than the immediately simpler model.
| Water | −1 MPa NaCl | −1 MPa PEG | |
|---|---|---|---|
| Plateau | 0.6367 (0.6309–0.6431) | 0.6063 (0.5946–0.6235) | 0.2640 (0.2612–0.2669) |
| Percent fast | 11.09 (8.82–13.56) | 13.02 (10.28–16.00) | 26.21 (22.29–30.83) |
| Percent medium | 28.48 (26.13–30.61) | 32.83 (30.12–35.39) | 73.79 |
| Percent slow | 60.43 | 54.15 | |
| Kfast (×10−1) | 9.64 (6.86–14.83) | 6.02 (4.26–9.20) | 4.02 (2.76–6.12) |
| Kmedium (×10−2) | 8.67 (7.17–10.38) | 5.58 (4.39–6.99) | 2.48 (2.24–2.72) |
| Kslow (×10−3) | 9.90 (9.19–10.57) | 7.00 (5.80–8.01) | |
| Fast half-life (hours) | 0.719 (0.47–1.01) | 1.152 (0.75–1.63) | 1.752 (1.13–2.51) |
| Medium half-life (hours) | 8.00 (6.7–9.7) | 12.42 (9.9–15.8) | 27.97 (25.5–30.9) |
| Slow half-life (hours) | 70.0 (65.6–75.4) | 98.9 (86.5–119.4) | |
| F-value (2, 26) | 78.26 | 83.5 | 2.01 |
| F-test p-value | <0.001 | <0.001 | 0.154 |
| Better model | Triple-exponential | Triple-exponential | Double-exponential |
| R2 value | 0.9997 | 0.9997 | 0.9974 |
| Shapiro-Wilk p-value | 0.3504 (passed) | 0.0905 (passed) | 0.2413 (passed) |
| Runs test p-value | 0.6424 (passed) | 0.7548 (passed) | 0.1085 (passed) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Irving, L.J.; Zhang, H. Modelling the Effect of Salt and PEG on Water Uptake in Wheat Seeds. Agronomy 2021, 11, 1660. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081660
AMA Style
Irving LJ, Zhang H. Modelling the Effect of Salt and PEG on Water Uptake in Wheat Seeds. Agronomy. 2021; 11(8):1660. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081660
Chicago/Turabian StyleIrving, Louis John, and Hongxiang Zhang. 2021. "Modelling the Effect of Salt and PEG on Water Uptake in Wheat Seeds" Agronomy 11, no. 8: 1660. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081660
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.