
Improved Conservation of Coffee (Coffea arabica L.) Germplasm via Micropropagation and Cryopreservation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Encapsulation–Vitrification
2.3. Droplet Vitrification
2.4. Shoot Multiplication, In Vitro Rooting, and Acclimatization
2.5. Differential Scanning Calorimetry (DSC) Analysis
2.6. Statistical Analysis
3. Results
3.1. Zygotic Embryo Culture
3.2. Cryopreservation Using Encapsulation–Vitrification
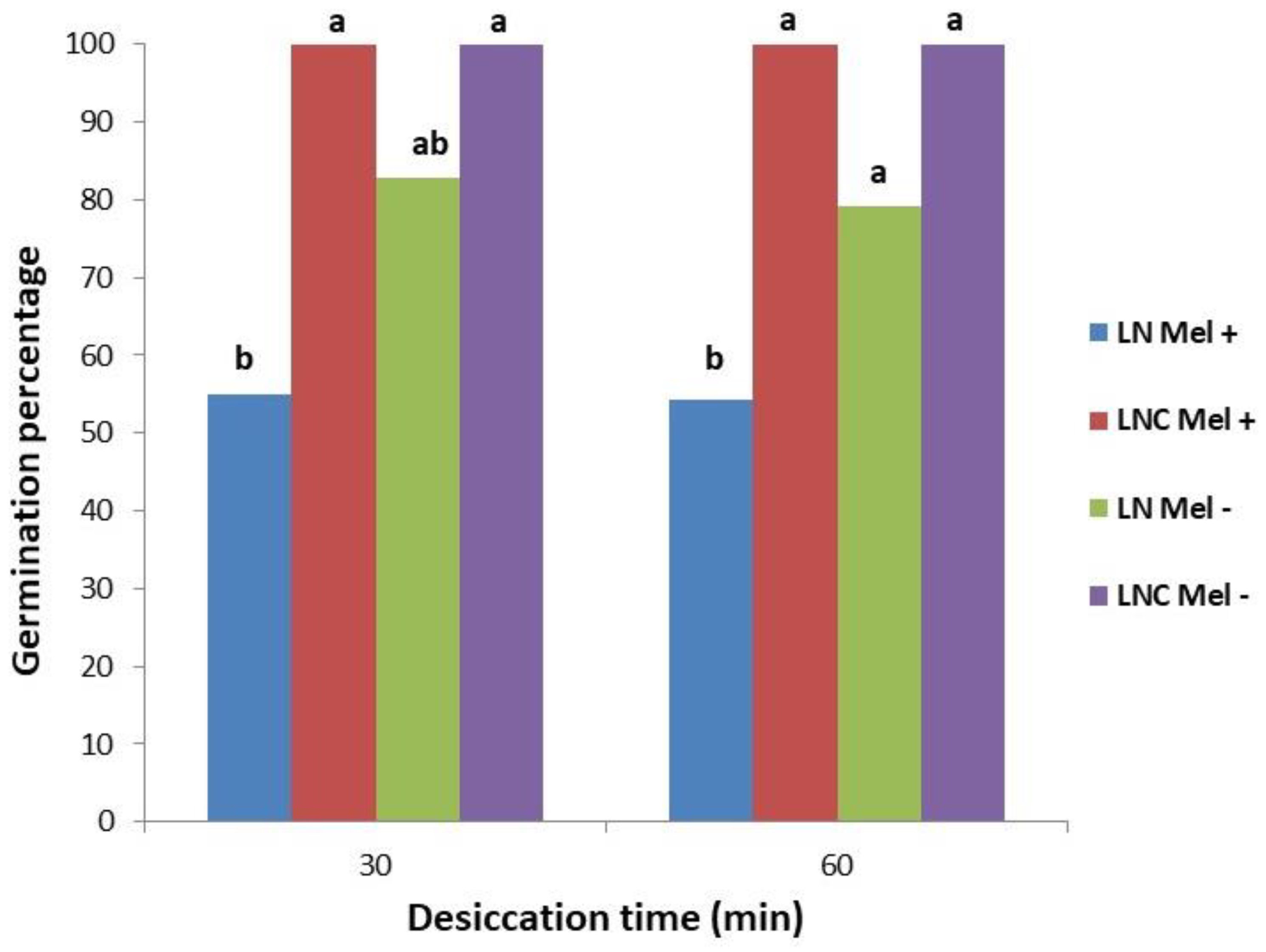
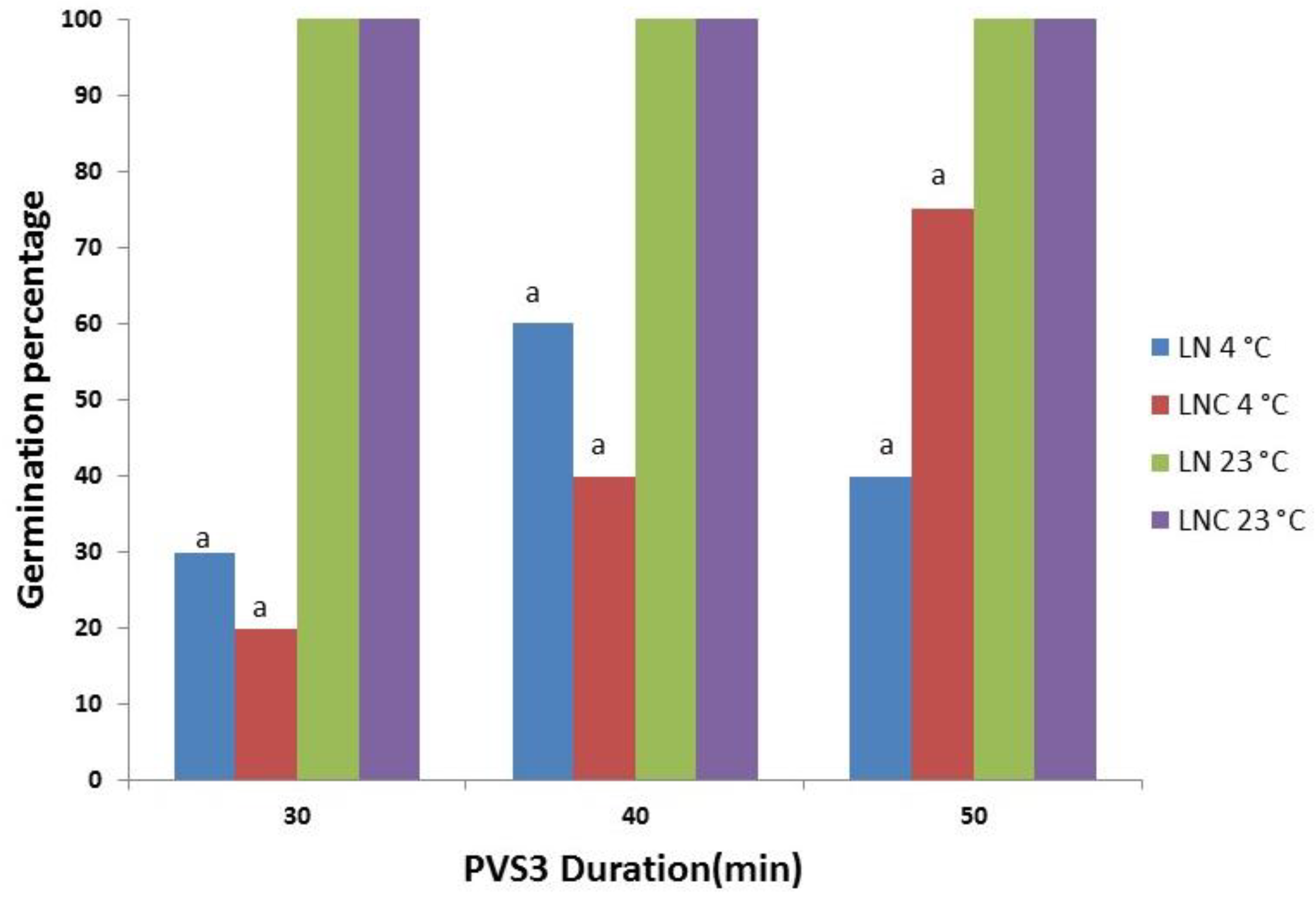
3.3. Cryopreservation Using Droplet Vitrification
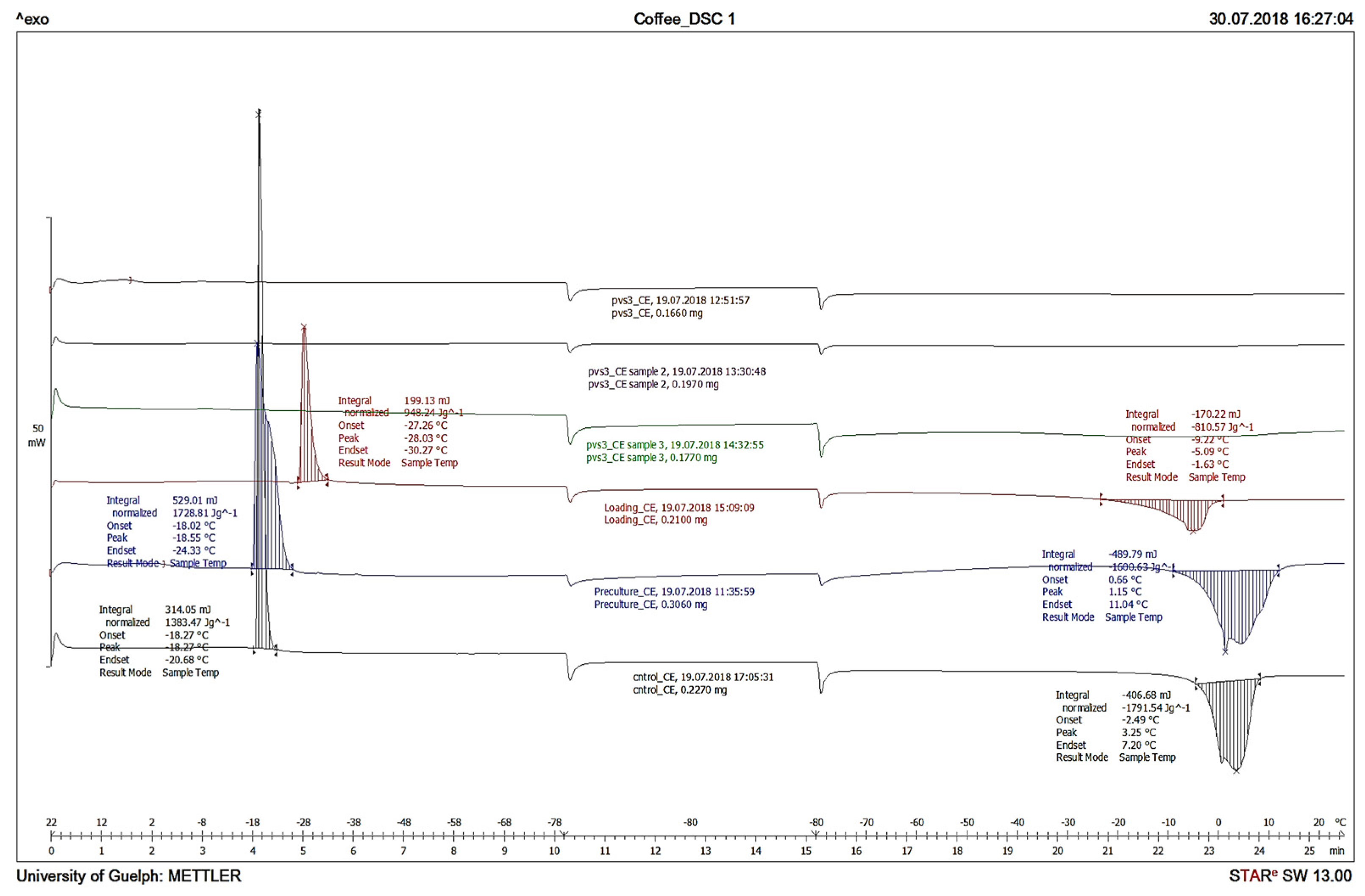
3.4. Differential Scanning Calorimetry
3.5. Shoot Multiplication and Acclimatization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bramel, P.; Krishnan, S.; Horna, D.; Lainoff, B.; Montagnon, C. Global Conservation Strategy for Coffee Genetic Resources; World Coffee Research-Crop Trust: Munich, Germany, 2017; pp. 5–7. Available online: https://worldcofferesearch.org/documents/42/Coffee_Strategy_Low_Res.pdf (accessed on 10 September 2021).
- Aguilar, M.E.; Ortiz, J.L.; Mesen, F.; Jimenez, L.D.; Altmann, F. Cafe Arabica Coffea arabica L. In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants; Forestry Sciences; Jain, S.M., Gupta, P., Eds.; Springer International Publishing AG: Basel, Switzerland, 2018; Volume 85, pp. 39–62. [Google Scholar] [CrossRef]
- ICO. Coffee Production Worldwide from 2003/04 to 2019/20 (in Million 60 Kilogram Bags)*. Statista, Statista Inc. Available online: https://0-www-statista-com.brum.beds.ac.uk/statistics/263311/worldwide-production-of-coffee/ (accessed on 31 January 2021).
- González, M.E.; Rosales, P.R.; Castilla, Y.; Lacerra, J.A.; Ferrer, M. Efecto del Bioenraiz® como estimulante de la germinación y el desarrollo de plántulas de cafeto (Coffea arabica L.). Cultiv. Trop. 2015, 36, 73–79. [Google Scholar]
- Wintgens, J.N.; Zamarripa, C.A. Coffee propagation. In Coffee: Growing, Processing, Sustainable Production: A Guidebook for Growers, Processors, Traders and Researchers; WILEY-VCH Verlag GMBH & Co. KGaA: Weinheim, Germany, 2004; pp. 91–140. [Google Scholar]
- Roubik, D. The value of bees to the coffee harvest. Nature 2002, 417, 708. [Google Scholar] [CrossRef]
- Ebrahim, N.; Shibli, R.; Makhadmeh, I.; Shatnawi, M.; Abu-Ein, A. In vitro propagation and in vivo acclimatization of three coffee cultivars (Coffea arabica L.) from Yemen. World Appl. Sci. J. 2007, 2, 142–150. [Google Scholar]
- Eira, M.T.S.; Amaral da Silva, E.A.; de Castro, R.D.; Dussert, S.; Walters, C.; Bewley, J.D.; Hilhorst, H.W.M. Coffee seed physiology. Braz. J. Plant Physiol. 2006, 18, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Isaac, E.; Hernández, C.; González, J.L.; González, M.E.; Fung, Y.; Ferrer, A.E. Effect of electromagnetic field on mineral content and chemical group during in vitro establishment and multiplication phases of coffee seedlings. Rev. Cubana Quím. 2016, 28, 692–702. [Google Scholar]
- Dussert, S.; Chabrillange, N.; Engelmann, F.; Anthony, F.; Hamon, S. Cryopreservation of coffee (Coffea arabica L.) seeds: Importance of the precooling temperature. CryoLetters 1997, 18, 269–276. [Google Scholar]
- Benson, E.E. Cryopreservation of phytodiversity: A critical appraisal of theory & practice. Crit. Rev. Plant Sci. 2008, 27, 141–219. [Google Scholar] [CrossRef]
- Engelmann, F. Use of biotechnologies for the conservation of plant biodiversity. Cell. Dev. Biol. Plant 2011, 47, 5–16. [Google Scholar] [CrossRef]
- Wang, M.R.; Chen, L.; Teixeira da Silva, J.A.; Volk, G.M.; Wang, Q.C. Cryobiotechnology of apple (Malus spp.): Development, progress and future prospects. Plant Cell Rep. 2018, 37, 689–709. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Bonnart, R.; Volk, G.M. Challenges in implementing plant shoot tip cryopreservation technologies. Plant Cell Tissue Organ Cult. 2021, 144, 21–34. [Google Scholar] [CrossRef]
- Popova, E.; Shukla, M.; Hoon, H.; Saxena, P.K. Plant Cryopreservation for Biotechnology and Breeding. In Advances in Plant Breeding Strategies: Breeding, Biotechnology and Molecular Tools; Al-Khayri, J.M., Jain, M., Johnson, D.V., Eds.; Springer International Publishing: Basel, Switzerland, 2015; pp. 63–93. [Google Scholar] [CrossRef]
- Abdelnour-Esquivel, A.; Villalobos, V.; Engelmann, F. Cryopreservation of zygotic embryos of Coffea spp. CryoLetters 1992, 13, 297–302. [Google Scholar]
- Pinto, M.S.; Paiva, R.; Correa, D.P.; Almeida, P.A.; Theresan, R.; Coutinho, L. Cryopreservation of coffee zygotic embryos: Dehydration and osmotic rehydration. Ciência e Agrotecnologia 2016, 40, 380–389. [Google Scholar] [CrossRef] [Green Version]
- Martínez, M.; González-Arnao, M.T.; Urra, C.; Rojas, R.; Cuba, M.; García, D. Estudios preliminares para la crioconservación de embriones cigóticos de Coffea arabica variedad 9722. Cultiv. Trop. 1996, 17, 79–81. [Google Scholar]
- Castilla, Y.; Dussert, S.; González, M.E.; Engelmann, F. Crioconservación de embriones cigóticos de Coffea canephora mediante vitrificación: Primeras aproximaciones. In Proceedings of the 8th International Congress on Plant Biotechnology and Agriculture Bioveg 2011, Ciego de Ávila, Cuba, 2–6 May 2011; ISBN 978-959-16-1286-1. [Google Scholar]
- Castilla, Y.; Martínez-Montero, M.E.; González, M.E.; Engelmann, F. Nuevos resultados en la crioconservación de dos especies de cafeto (Coffea spp) mediante el método de vitrificación. In Proceedings of the 9th International Congress on Plant Biotechnology and Agriculture Bioveg 2013, Ciego de Ávila, Cuba, 7–10 May 2013; ISBN 978-959-16-2045-3. [Google Scholar]
- De Freitas, R.T.; Paiva, R.; Silva, T.; Pedrosa, D.; Valquiria, M.; de Souza, A.C.; Veiga, S.D. Cryopreservation of Coffea arabica L. zygotic embryos by vitrification. Not. Bot. Horti Agrobot. 2016, 44, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Niino, T. The development of plant vitrification solution 2 and recent PVS2-based vitrification protocols. Acta Hortic. 2014, 1039, 21–28. [Google Scholar] [CrossRef]
- Matsumoto, T. Cryopreservation of plant genetic resources: Conventional and new methods. Rev. Agric. Sci. 2017, 5, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Carra, A.; Carimi, F.; Bettoni, J.C.; Pathirana, R. Progress and Challenges in the Application of Synthetic Seed Technology for Ex Situ Germplasm Conservation in Grapevine (Vitis spp.). In Synthetic Seeds; Faisal, A., Alatar, A.A., Eds.; Springer Nature: Basel, Switzerland, 2019; pp. 439–467. [Google Scholar] [CrossRef]
- Sakai, A.; Engelmann, F. Vitrification, encapsulation-vitrification and droplet-vitrification: A review. CryoLetters 2007, 28, 151–172. [Google Scholar] [PubMed]
- Martínez, M.E.; Harding, K. Cryobionomics: Evaluating the concept in plant cryopreservation. In Plant Omics: The Omics of Plant Science; Barh, D., Khan, M.S., Davies, E., Eds.; Springer: New Delhi, India, 2015; pp. 655–678. [Google Scholar] [CrossRef]
- Lazo-Javelera, M.F.; Tiznado-Hernández, M.E.; Vargas-Arispuro, I.; Valenzuela-Soto, E.; Rocha-Granados, M.C.; Mar-tínez-Montero, M.E.; Rivera-Domínguez, M. Data on antioxidant activity in grapevine (Vitis vinifera L.) following cryopreservtion by vitrification. Data Brief 2015, 5, 549–555. [Google Scholar] [CrossRef] [Green Version]
- Reed, B.M. Antioxidants and cryopreservation, the new normal? Acta Hort. 2014, 1039, 41–48. [Google Scholar] [CrossRef]
- Bettoni, J.; Kretzschmar, A.A.; Bonnart, R.; Shepherd, A.; Volk, G.M. Cryopreservation of 12 Vitis species using apical shoot tips derived from plants grown in vitro. HortScience 2019, 54, 976–981. [Google Scholar] [CrossRef] [Green Version]
- Uchendu, E.E.; Muminova, M.; Gupta, S.; Reed, B.M. Antioxidant and antistress compounds improve regrowth of cryopreserved Rubus shoot tips. Cell. Dev. Biol. Plant 2010, 46, 386–393. [Google Scholar] [CrossRef]
- Uchendu, E.E.; Keller, E.R.J. Melatonin-loaded alginate beads improve cryopreservation of yam (Dioscorea alata and D. cayenensis). Cryoletters 2016, 37, 77–87. [Google Scholar]
- Ha, J.S.; Lee, J.W.; Seo, S.H.; Ahn, C.Y.; Rho, G.J.; Lee, H.G.; Oh, H.M. Optimized cryopreservation of Ettlia sp. using short cold acclimation and controlled freezing procedure. J. Appl. Phycol. 2019, 31, 2277–2287. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dereuddre, J.; Scottez, C.; Arnaud, Y.; Duron, M. Resistance of alginate-coated axillary shoot tips of pear tree (Pyrus communis L. cv. Beurre Hardy) in vitro plantlets to dehydration and subsequent freezing in liquid nitrogen. C. R. Acad. Sci. Paris 1990, 310, 317–322. [Google Scholar]
- Fabre, J.; Dereuddre, J. Encapsulation-dehydration: A new approach to cryopreservation of Solanum shoot tips. CryoLetters 1990, 11, 413–426. [Google Scholar]
- Nishizawa, S.; Sakai, A.; Amano, Y.; Matsuzawa, T. Cryopreservation of asparagus (Asparagus officinalis L.) embryogenic suspension cells and subsequent plant regeneration by vitrification. Plant Sci. 1993, 91, 67–73. [Google Scholar] [CrossRef]
- Nicholson, J.; Shukla, M.R.; Saxena, P.K. In vitro rooting of Hybrid hazelnuts (Corylus avellana X Corylus americana) in a temporary immersion system. Botany 2020, 98, 343–352. [Google Scholar] [CrossRef]
- Shukla, M.R.; Singh, A.S.; Piunno, K.; Saxena, P.K.; Jones, A.M.P. Application of 3D printing to prototype and develop novel plant tissue culture systems. Plant Methods 2017, 13, 6. [Google Scholar] [CrossRef] [Green Version]
- Tran, H.T.M.; Lee, L.S.; Furtado, A.; Smyth, H.; Henry, R.J. Advances in genomics for the improvement of quality in coffee. Sci. Food Agric. 2016, 96, 3300–3312. [Google Scholar] [CrossRef] [Green Version]
- Rathwell, R.; Popova, E.; Shukla, M.R.; Saxena, P.K. Development of cryopreservation methods for cherry birch (Betula lenta L.), an endangered tree species in Canada. Can. J. For. Res. 2016, 46, 1284–1292. [Google Scholar] [CrossRef] [Green Version]
- Hatanaka, T.; Yasuda, T.; Yamaguchi, T.; Sakai, A. Direct regrowth of encapsulated somatic embryos of coffee (Coffea canephora) after cooling in liquid nitrogen. CryoLetters 1994, 15, 47–52. [Google Scholar]
- Mari, S.; Engelmann, F.; Chabrillange, N.; Huet, C.; Michaux-Ferriere, N. Histo-cytological studies of apices of coffee (Coffea racemosa and C. sessiliflora) in vitro plantlets during their cryopreservation using the encapsulation-dehydration technique. CryoLetters 1995, 16, 289–298. [Google Scholar]
- Takagi, H. Recent developments in cryopreservation of shoot apices of tropical species. In Cryopreservation of Tropical Plant Germplasm: Current Research Progress and Applications; Engelmann, F., Takagi, H., Eds.; Japan International Research Center for Agricultural Sciences (JIRCAS): Tsukuba, Japan; International Plant Genetic Resources Institute (IPGRI): Rome, Italy, 2000; pp. 178–193. [Google Scholar]
- Ciringer, T.; Martín, C.; Sajna, N.; Kaligaric, N.; Ambrozik-Dolinsek, J. Cryopreservation of an endangered Hladniki apastinacifolia Rchb. by shoot-tip encapsulation-dehydration and encapsulation-vitrification. Cell. Dev. Biol. Plant 2018, 54, 565–575. [Google Scholar] [CrossRef]
- Malik, S.K.; Chaudhury, R. Cryopreservation techniques for conservation of tropical horticultural species using various explants. In Conservation and Utilization of Horticultural Genetic Resources; Rajasekharan, P., Rao, V., Eds.; Springer Nature: New York, NY, USA, 2019; pp. 579–594. [Google Scholar] [CrossRef]
- Jouve, L.; Engelmann, F.; Noirot, M.; Charrier, A. Evaluation of biochemical markers (sugar, proline, malonedialdehyde and ethylene) for cold sensitivity in microcuttings of two coffee species. Plant Sci. 1993, 91, 109–116. [Google Scholar] [CrossRef]
- Volk, G.M.; Harris, J.L.; Rotindo, K.E. Survival of mint shoot tips after exposure to cryoprotectant solution components. Cryobiology 2006, 52, 305–308. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Souza, J.A.; Volk, G.M.; Dalla Costa, M.; da Silva, F.N.; Kretzschmar, A.A. Eradication of latent viruses from apple cultivar ’Monalisa’ shoot tips using droplet-vitrification cryotherapy. Sci. Hortic. 2019, 250, 12–18. [Google Scholar] [CrossRef]
- Kim, H.H.; Popova, E.; Shin, D.J.; Yi, J.Y.; Kim, C.H.; Lee, J.S.; Yoon, M.K.; Engelmann, F. Cryobanking of Korean Allium Germplasm Collections: Results from a 10 Year Experience. Cryoletters 2012, 33, 45–57. [Google Scholar]
- Uchendu, E.E.; Shukla, M.R.; Reed, B.M.; Saxena, P.K. Melatonin enhances the recovery of cryopreserved shoot tips of American elm (Ulmus americana L.). J. Pineal Res. 2013, 55, 435–442. [Google Scholar] [CrossRef]
- Uchendu, E.E.; Shukla, M.R.; Reed, B.M.; Saxena, P.K. An efficient method for cryopreservation of St Jhon’s Wort and tobacco: Role of melatonin. ISHS Acta Hortic. 2013, 1039, 233–241. [Google Scholar]
- Erland, L.A.E.; Saxena, P.K. Melatonin in plant morphogenesis. Cell. Dev. Biol. Plant 2018, 54, 3–24. [Google Scholar] [CrossRef]
- Campos, C.M.; Ávila, R.G.; Dázio de Souza, K.R.; Azevedo, L.M.; Alves, J.D. Melatonin reduces oxidative stress and promotes drought tolerance in young Coffea arabica L. plants. Agric. Water Manag. 2019, 211, 37–47. [Google Scholar] [CrossRef]
- Ramakrishna, A.; Giridhar, P.; Sankar, K.U.; Ravishankar, G.A. Melatonin and serotonin profiles in beans of Coffea species. J. Pineal Res. 2012, 52, 470–476. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández, J. Growth conditions determine different melatonin levels in Lupinus albus L. J. Pineal Res. 2013, 55, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Latawa, J.; Shukla, M.R.; Saxena, P.K. An efficient temporary immersion system for micropropagation of hybrid hazelnut. Botany 2016, 94, 1–8. [Google Scholar] [CrossRef]
- Etienne-Barry, D.; Bertrand, B.; Vasquez, N.; Etienne, H. Direct sowing of Coffea arabica somatic embryos mass-produced in a bioreactor and regeneration of plants. Plant Cell Rep. 1999, 19, 111–117. [Google Scholar] [CrossRef]
- Albarrán, J.; Bertrand, B.; Lartaud, M.; Etienne, H. Cycle characteristics in a temporary immersion bioreactor affect regeneration, morphology, water and mineral status of coffee (Coffea arabica) somatic embryos. Plant Cell Tissue Organ Cult. 2005, 81, 27–36. [Google Scholar] [CrossRef]
- Etienne, H.; Dechamp, E.; Barry-Etienne, D.; Bertrand, B. Bioreactors in coffee micropropagation. Braz. J. Plant Physiol. 2006, 18, 45–54. [Google Scholar] [CrossRef]
- Gatica-Arias, A.M.; Arrieta-Espinoza, G.; Espinoza Esquivel, A.M. Plant regeneration via indirect somatic embryogenesis and optimization of genetic transformation in Coffea arabica L. cvs. Caturra and Catuaí. Electron. J. Biotechnol. 2008, 11, 101–112. [Google Scholar]
- Shukla, M.R.; Jones, A.M.P.; Sullivan, J.A.; Liu, C.; Gosling, S.; Saxena, P.K. In vitro conservation of American elm (Ulmus americana): Potential role of auxin metabolism in sustained plant proliferation. Can. J. For. Res. 2012, 42, 686–697. [Google Scholar] [CrossRef]
- Shukla, M.R.; Kibler, A.; Turi, C.E.; Erland, L.A.E.; Sullivan, J.A.; Murch, S.J.; Saxena, P.K. Selection and Micropropagation of an Elite Melatonin Rich Tulsi (Ocimum sanctum L.) Germplasm Line. Agronomy 2021, 11, 207–231. [Google Scholar] [CrossRef]
- Naylor-Adelberg, J.A.; Adelberg, J.W.; Rapaka, V.K. A rooting matrix and tissue culture vessel system yielded larger, faster growing Echeveria plants. Acta Hortic. 2016, 1113, 53–58. [Google Scholar] [CrossRef]
- Adelberg, J. Bioreactors and “smart vessels” for large-scale propagation. Acta Hortic. 2017, 1187, 123–138. [Google Scholar] [CrossRef]
- Economou, A.S. From microcutting rooting to microplant establishment: Key points to consider for maximum success in woody plants. Acta Hortic. 2013, 988, 43–56. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valdés, Y.C.; Shukla, M.R.; González Vega, M.E.; Saxena, P.K. Improved Conservation of Coffee (Coffea arabica L.) Germplasm via Micropropagation and Cryopreservation. Agronomy 2021, 11, 1861. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091861
Valdés YC, Shukla MR, González Vega ME, Saxena PK. Improved Conservation of Coffee (Coffea arabica L.) Germplasm via Micropropagation and Cryopreservation. Agronomy. 2021; 11(9):1861. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091861
Chicago/Turabian StyleValdés, Yanelis Castilla, Mukund R. Shukla, María Esther González Vega, and Praveen K. Saxena. 2021. "Improved Conservation of Coffee (Coffea arabica L.) Germplasm via Micropropagation and Cryopreservation" Agronomy 11, no. 9: 1861. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091861