
Mitigation of High-Temperature Damage by Application of Kaolin and Pinolene on Young Olive Trees (Olea europaea L.): A Preliminary Experiment to Assess Biometric, Eco-Physiological and Nutraceutical Parameters

 , ,
, ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions, Experimental Design and Products Applications
2.2. Biometric and Physiological Analysis
2.3. Determination of Leaf Polyphenols
2.3.1. Chemicals
2.3.2. Ultrasound-Assisted Extraction of Polyphenolic Compounds
2.3.3. UHPLC-HRMS Analysis of Polyphenolic Compounds
2.4. Antioxidant Activity: ABTS Assay
2.5. Antioxidant Activity: DPPH Assay
2.6. Statistical Analysis
3. Results and Discussion
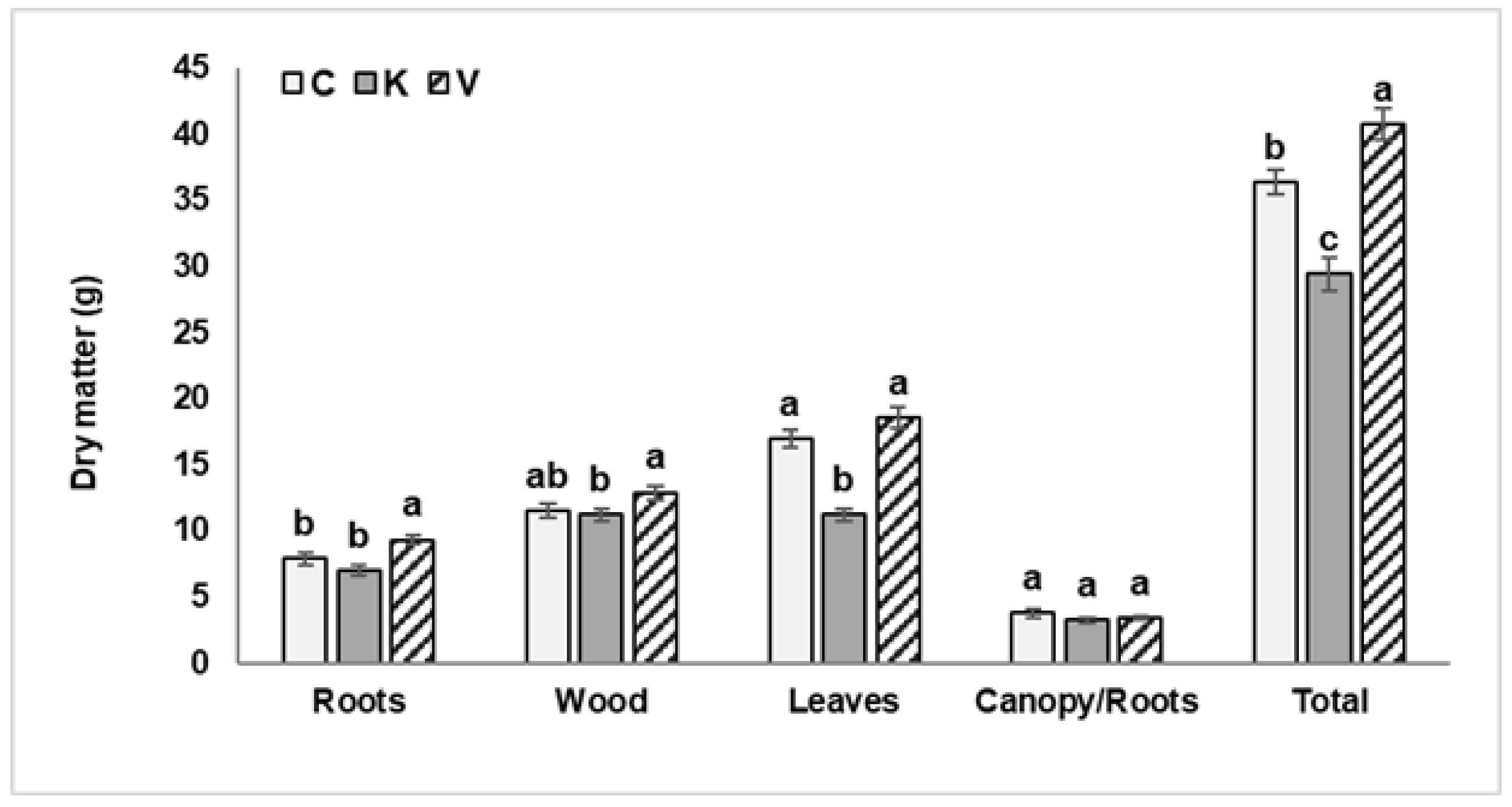
3.1. Implications of Kaolin and Pinolene for Biometric and Physiological Parameters
3.2. Analysis of Polyphenols by UHPLC-Q-Orbitrap HRMS
3.3. Antioxidant Activity of Polyphenolics Extracts
3.4. Principal Component Analysis (PCA)
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moriondo, M. Modelling Olive Trees and Grapevines in a Changing Climate. Environ. Model. 2015, 72, 387–401. [Google Scholar] [CrossRef]
- Orlandi, F.; Avolio, E.; Bonofiglio, T.; Federico, S.; Romano, B.; Fornaciari, M. Potential Shifts in Olive Flowering According to Climate Variations in Southern Italy. Meteorol. Appl. 2012, 20, 497–503. [Google Scholar] [CrossRef]
- Fraga, H.; Pinto, J.G.; Viola, F.; Santos, J.A. Climate Change Projections for Olive Yields in the Mediterranean Basin. Int. J. Climatol. 2020, 40, 769–781. [Google Scholar] [CrossRef] [Green Version]
- Mearns, L.O.; Rosenzweig, C.; Goldberg, R. Mean and variance change in climate scenarios: Methods, agricultural applications, and measures of uncertainty. Clim. Chang. 1997, 35, 367–396. [Google Scholar] [CrossRef]
- Orlandi, F.; Ruga, L.; Romano, B.; Fornaciari, M. Olive flowering as an indicator of local climatic changes. Theor. Appl. Climatol. 2005, 81, 169–176. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.; McGuire, M.A.; Teskey, R.O. Stomatal conductance increases with rising temperature. Plant Signal. Behav. 2017, 12, e1356534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, J.M.; Mitchell, J.F.B.; Brady, A.J. Summer Drought in Northern Midlatitudes in a Time-Dependent CO2 Climate Experiment. J. Clim. 1997, 10, 25. [Google Scholar] [CrossRef]
- Di Vaio, C.; Marallo, N.; Marino, G.; Caruso, T. Effect of Water Stress on Dry Matter Accumulation and Partitioning in Pot-Grown Olive Trees (Cv Leccino and Racioppella). Sci. Hortic. 2013, 164, 155–159. [Google Scholar] [CrossRef]
- Angelopoulos, K.; Dichio, B.; Xiloyannis, C. Inhibition of photosynthesis in olive trees (Olea europaea L.) during water stress and rewatering. Exp. Bot. J. 1996, 47, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Fernández, J.E.; Moreno, F.; Girón, I.F.; Blázquez, O.M. Stomatal control of water use in olive tree leaves. Plant Soil 1997, 190, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M.; Goncalves, B.C.; Ferreira, H.F.; Correia, C.M. Immediate Responses and Adaptative Strategies of Three Olive Cultivars under Contrasting Water Availability Regimes: Changes on Structure and Chemical Composition of Foliage and Oxidative Damage. Plant Sci. 2006, 170, 596–605. [Google Scholar] [CrossRef]
- Bosabalidis, A.M.; Kofidis, G. Comparative Effects of Drought Stress on Leaf Anatomy of Two Olive Cultivars. Plant Sci. 2002, 163, 375–379. [Google Scholar] [CrossRef]
- Boughalleb, F.; Hajlaoui, H. Physiological and Anatomical Changes Induced by Drought in Two Olive Cultivars (Cv Zalmati and Chemlali). Acta Physiol. Plant. 2011, 33, 53–65. [Google Scholar] [CrossRef]
- Denaxa, N.-K. Comparative Effects of Exogenous Glycine Betaine, Kaolin Clay Particles and Ambiol on Photosynthesis, Leaf Sclerophylly Indexes and Heat Load of Olive Cv. Chondrolia Chalkidikis under Drought. Sci. Hortic. 2012, 137, 87–94. [Google Scholar] [CrossRef]
- Wang, S. Genotypic Variation in Growth and Physiological Response to Drought Stress and Re-Watering Reveals the Critical Role of Recovery in Drought Adaptation in Maize Seedlings. Front. Plant Sci. 2016, 6, 15. [Google Scholar]
- Perez-Martin, A.; Torres-Ruiz, J.M.; Fernández, J.E.; Diaz-Espejo, A.; Flexas, J.; Michelazzo, C.; Sebastiani, L. Physiological and genetic response of olive leaves to water stress and recovery: Implications of mesophyll conductance and genetic expression of aquaporins and carbonic anhydrase. In Proceedings of the XXVIII International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on CLIMWATER 2010: Horticultural Use of Water in a Changing Climate, Lisbon, Portugal, 22–27 August 2010; Fernández, J.E., Ferreira, M.I., Eds.; ISHS Acta Horticulturae: Leuven, Belgium, 2010; Volume 922, pp. 99–105. [Google Scholar]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M. Physiological Behaviour, Oxidative Damage and Antioxidative Protection of Olive Trees Grown under Different Irrigation Regimes. Plant Soil. 2007, 292, 1. [Google Scholar] [CrossRef]
- Petridis, A. Effect of Water Deficit on Leaf Phenolic Composition, Gas Exchange, Oxidative Damage and Antioxidant Activity of Four Greek Olive (Olea europaea L.) Cultivars. Plant Physiol. Biochem. 2012, 60, 1–11. [Google Scholar] [CrossRef]
- Wungrampha, S.; Joshi, R.; Singla-Pareek, S.L.; Pareek, A. Photosynthesis and salinity: Are these mutually exclusive? Photosynthetica 2018, 56, 366–381. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops—What Is the Cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Roháček, K.; Soukupová, J.; Barták, M. Chlorophyll fluorescence: A wonderful tool to study plant physiology and plant stress. Plant Cell Compart.-Sel. Top. Res. Signpost Kerala India 2008, 37, 41–104. [Google Scholar]
- Del Amor, F.M. Growth, Photosynthesis and Chlorophyll Fluorescence of Sweet Pepper Plants as Affected by the Cultivation Method. Ann. Appl. Biol. 2006, 148, 133–139. [Google Scholar] [CrossRef]
- Cantore, V.; Pace, B.; Albrizio, R. Kaolin-Based Particle Film Technology Affects Tomato Physiology, Yield and Quality. Environ. Exp. Bot. 2009, 66, 279–288. [Google Scholar] [CrossRef]
- Lombardini, L.; Harris, M.K.; Glenn, D.M. Effects of particle film application on leaf gas exchange, water relations, nut yield, and insect populations in mature pecan trees. HortScience 2005, 40, 1376–1380. [Google Scholar] [CrossRef] [Green Version]
- Rosati, A.; Metcalf, S.G.; Buchner, R.P.; Fulton, A.E.; Lampinen, B.D. Effects of kaolin application on light absorption and distribution, radiation use efficiency and photosynthesis of almond and walnut canopies. Ann. Bot. 2007, 99, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Palliotti, A.; Poni, S.; Berrios, J.G.; Bernizzoni, F. Vine performance and grape composition as affected by early-season source limitation induced with anti-transpirants in two red Vitis vinifera L. cultivars. Aust. J. Grape Wine Res. 2010, 16, 426–433. [Google Scholar] [CrossRef]
- Gu, S.; Fuchigami, L.H.; Cheng, L.; Guak, S.H.; Shin, C. Effects of antitranspirant and leaching on medium solution osmotic potential, leaf stomatal status, transpiration, abscisic acid content and plant growth in’Early Girl’tomato plants (Lycopersicon esculentum). J. Hortic. Sci. 1998, 73, 473–477. [Google Scholar] [CrossRef]
- Mola, I.D.; Conti, S.; Cozzolino, E.; Melchionna, G.; Ottaiano, L.; Testa, A.; Sabatino, A.; Rouphae, Y.; Mori, M. Plant-Based Protein Hydrolysate Improves Salinity Tolerance in Hemp: Agronomical and Physiological Aspects. Agronomy 2021, 11, 342. [Google Scholar] [CrossRef]
- Di Vaio, C.; Graziani, G.; Gaspari, A.; De Luca, L.; Aiello, A.; Cirillo, A.; Bruno, A.; Romano, R.; Ritieni, A. Drupe Characters, Fatty Acids, Polyphenolic and Aromatic Profile of Olive Oil Obtained from “Oliva Bianca”, Minor Autochthonous Cultivar of Campania. Plants 2021, 10, 1119. [Google Scholar] [CrossRef] [PubMed]
- Talhaoui, N.; Gómez-Caravaca, A.M.; León, L.; De la Rosa, R.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Determination of Phenolic Compounds of ‘Sikitita’ Olive Leaves by HPLC-DAD-TOF-MS. Comparison with Its Parents ‘Arbequina’ and ‘Picual’ Olive Leaves. LWT 2014, 58, 28–34. [Google Scholar] [CrossRef]
- Dini, I.; Graziani, G.; Fedele, F.L.; Sicari, A.; Vinale, F.; Castaldo, L.; Ritieni, A. Effects of Trichoderma Biostimulation on the Phenolic Profile of Extra-Virgin Olive Oil and Olive Oil By-Products. Antioxidants 2020, 9, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Roussos, P.A.; Denaxa, N.-K.; Damvakaris, T.; Stournaras, V.; Argyrokastritis, I. Effect of Alleviating Products with Different Mode of Action on Physiology and Yield of Olive under Drought. Sci. Hortic. 2010, 125, 700–711. [Google Scholar] [CrossRef]
- MI, A.D.; Abd El-Rhman, I.E.; Sahar, A.F. Effect of Some Antitranspirants and Supplementery Irrigation on Growth, Yield and Fruit Quality of Sultani Fig (Ficus carica) Grown in the Egyption Western Coastal Zone under Rainfed Conditions. AGBR 2009, 5, 899–908. [Google Scholar]
- Davenport, D.C.; Fisher, M.A.; Hagan, R.M. Some counteractive effects of antitranspirants. Plant Physiol. 1972, 49, 722–724. [Google Scholar] [CrossRef] [Green Version]
- Di Vaio, C.; Marallo, N.; Di Lorenzo, R.; Pisciotta, A. Anti-transpirant effects on vine physiology, berry and wine composition of cv. Aglianico (Vitis vinifera L.) grown in south Italy. Agronomy 2019, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Di Vaio, C.; Villano, C.; Lisanti, M.T.; Marallo, N.; Cirillo, A.; Di Lorenzo, R.; Pisciotta, A. Application of Anti-Transpirant to Control Sugar Accumulation in Grape Berries and Alcohol Degree in Wines Obtained from Thinned and Unthinned Vines of cv. Falanghina (Vitis vinifera L.). Agronomy 2020, 10, 345. [Google Scholar] [CrossRef] [Green Version]
- Nielson, R.E.; Jarvis, P.G. Photosynthesis in Sitka Spruce (Picea sitchensis (Bong.) Carr.): VI. Response of Stomata to Temperature. J. Appl. Ecol. 1975, 12, 879–891. [Google Scholar] [CrossRef]
- Turner, N.C. Concurrent comparisons of stomatal behavior, water status, and evaporation of maize in soil at high or low water potential. Plant Physiol. 1975, 55, 932–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Wang, W.; Zhang, C.; Ma, Q.; Li, Y. Postharvest physiochemical responses of cut rose (Rosa hybrida L.) to antitranspirant and vacuum cooling. Philipp. Agric. Sci. 2011, 94, 368–374. [Google Scholar]
- Misra, A.K.; Das, B.K.; Datta, J.K.; De, G.C. Effect of antitranspirants on water status and growth pattern of Mulberry (Morus alba L.) under two levels of irrigation. Indian J. Agric. Res. 2009, 43, 307–310. [Google Scholar]
- Glenn, D.M. The mechanisms of plant stress mitigation by kaolin-based particle films and applications in horticultural and agricultural crops. HortScience 2012, 47, 710–711. [Google Scholar] [CrossRef] [Green Version]
- Del Amor, F.M.; Cuadra-Crespo, P.; Walker, D.J.; Cámara, J.M.; Madrid, R. Effect of foliar application of antitranspirant on photosynthesis and water relations of pepper plants under different levels of CO2 and water stress. J. Plant Physiol. 2010, 167, 1232–1238. [Google Scholar] [CrossRef] [PubMed]
- Mikiciuk, G.; Mikiciuk, M.; Ptak, P. The effects of anitranspirant di-1-p-menthene on some physiological traits of strawberry. J. Ecol. Eng. 2015, 16, 161–167. [Google Scholar] [CrossRef]
- Abdel-Fattah, G.H. Response of water-stressed rose of China (Hibiscus rosa-sinensis L.) plant to treatment with calcium carbonate and vapor gard antitranspirants. J. Appl. Sci. Res. 2013, 9, 3566–3572. [Google Scholar]
- Björkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef]
- Segura-Monroy, S.; Uribe-Vallejo, A.; Ramirez-Godoy, A.; Restrepo-Diaz, H. Effect of kaolin application on growth, water use efficiency, and leaf epidermis characteristics of Physallis peruviana seedlings under two irrigation regimes. J. Agric. Sci. Technol. 2015, 17, 1585–1596. [Google Scholar]
- Latocha, P.; Ska, M.C.; Pietkiewicz, S.; Kalaji, M.H. Preliminary Assessment of Antitranspirant Vapor Gard® Influence on Actinidia Arguta Growing under Drought Stress Conditions. Ann. Wars. Univ. Life Sci. 2009, 30, 149–150. [Google Scholar]
- Lawlor, D.W. Limitation to Photosynthesis in Water-Stressed Leaves: Stomata vs. Metabolism and the Role of ATP. Ann. Bot. 2002, 89, 871–885. [Google Scholar] [CrossRef]
- Percival, G.C.; Keary, I.P.; Marshall, K. The use of film-forming polymers to control Guignardia leaf blotch and powdery mildew on Aesculus hippocastanum L. and Quercus robur L. Arboric. Urban For. 2006, 32, 100. [Google Scholar]
- Anderson, J.M. Photoregulation of the composition, function, and structure of thylakoid membranes. Annu. Rev. Plant Physiol. 1986, 37, 93–136. [Google Scholar] [CrossRef]
- Shekour, G.M.; Brathwaite, R.A.I.; McDavid, C.R. Dry Season Sweet Corn Response to Mulching and Antitranspirants1. J. Agron. 1987, 79, 629–631. [Google Scholar] [CrossRef]
- Denaxa, N.-K.; Damvakaris, T.; Roussos, P.A. Antioxidant Defense System in Young Olive Plants against Drought Stress and Mitigation of Adverse Effects through External Application of Alleviating Products. Sci. Hortic. 2020, 259, 108812. [Google Scholar] [CrossRef]
- Brillante, L.; Belfiore, N.; Gaiotti, F.; Lovat, L.; Sansone, L.; Poni, S.; Tomasi, D. Comparing Kaolin and Pinolene to Improve Sustainable Grapevine Production during Drought. PLoS ONE 2016, 11, e0156631. [Google Scholar] [CrossRef] [PubMed]
- Valentini, G.; Pastore, C.; Allegro, G.; Muzzi, E.; Seghetti, L.; Filippetti, I. Application of Kaolin and Italian Natural Chabasite-Rich Zeolitite to Mitigate the Effect of Global Warming in Vitis vinifera L. cv. Sangiovese. Agronomy 2021, 11, 1035. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Shoots Growth (cm) | TCSA (cm2) | Leaf Area per Plant (cm2) |
|---|---|---|---|
| C | 14.44 ± 1.55 b | 17.05 ± 1.78 b | 682.02 ± 16.00 b |
| K | 18.20 ± 1.08 ab | 16.21 ± 1.51 b | 759.03 ± 25.94 a |
| V | 21.17 ± 1.84 a | 22.97 ± 2.09 a | 746.30 ± 23.88 a |
| Significance | *** | * | * |
| Theorical Mass | Experimental Mass | Error | ||
|---|---|---|---|---|
| Compound | Formula | [M-H]− | Δ ppm | |
| Ligstroside | C25H32O12 | 523.18210 | 523.18079 | −2.50 |
| Oleuropein aglycone | C19H22O8 | 377.12419 | 377.12442 | 0.61 |
| Verbascoside | C29H36O15 | 623.19814 | 623.19952 | 2.21 |
| Oleuropein | C25H32O13 | 539.17701 | 539.17792 | 1.69 |
| OH-tyrosol-glucoside | C13H18O8 | 301.09289 | 301.09329 | −1.85 |
| Luteolin | C15H10O6 | 285.04062 | 285.04083 | 0.74 |
| Polyphenols | Treatments | Significance | ||
|---|---|---|---|---|
| C | K | V | T | |
| Ligstroside | 54.01 ± 1.12 b | 266.16 ± 1.54 a | 67.79 ± 1.54 b | *** |
| Oleuropein aglycone | 54.67 ± 2.02 b | 230.89 ± 0.45 a | 45.9 0± 1.23 c | *** |
| Verbascoside | 156.9 1± 4.91 b | 381.39 ± 5.71 a | 31.51 ± 0.34 c | *** |
| Oleuropein | 629.34 ± 5.81 b | 1868.74 ± 6.54 a | 575.93 ± 3.58 b | *** |
| OH-tyrosol glucoside | 2185.11 ± 0.34 b | 8215.27 ± 8.43 a | 980.91 ± 7.34 c | *** |
| Luteolin | 205.00 ± 3.43 ab | 217.59 ± 3.94 a | 191.02 ± 2.58 b | ns |
| Total polyphenols | 3285.04 b | 11,180.05 a | 1893.07 c | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cirillo, A.; Conti, S.; Graziani, G.; El-Nakhel, C.; Rouphael, Y.; Ritieni, A.; Di Vaio, C. Mitigation of High-Temperature Damage by Application of Kaolin and Pinolene on Young Olive Trees (Olea europaea L.): A Preliminary Experiment to Assess Biometric, Eco-Physiological and Nutraceutical Parameters. Agronomy 2021, 11, 1884. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091884
Cirillo A, Conti S, Graziani G, El-Nakhel C, Rouphael Y, Ritieni A, Di Vaio C. Mitigation of High-Temperature Damage by Application of Kaolin and Pinolene on Young Olive Trees (Olea europaea L.): A Preliminary Experiment to Assess Biometric, Eco-Physiological and Nutraceutical Parameters. Agronomy. 2021; 11(9):1884. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091884
Chicago/Turabian StyleCirillo, Aurora, Stefano Conti, Giulia Graziani, Christophe El-Nakhel, Youssef Rouphael, Alberto Ritieni, and Claudio Di Vaio. 2021. "Mitigation of High-Temperature Damage by Application of Kaolin and Pinolene on Young Olive Trees (Olea europaea L.): A Preliminary Experiment to Assess Biometric, Eco-Physiological and Nutraceutical Parameters" Agronomy 11, no. 9: 1884. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091884