1. Introduction
The concept of high-density plantings (HDP) in apple orchards began with the launching of the dwarfing rootstocks from the East Malling Research Station’s (UK) rootstock development program. The most popular rootstock from the East Malling M-series was the dwarfing, ‘M9’ [
1]. A joint breeding program between the East Malling Research Station and the John Innes Institute in Merton, England, focused on producing woolly apple aphid-resistant rootstocks. As a result of this collaboration, two series of rootstocks were formed: the Merton–Immune (MI) series and the Malling–Merton (MM) series (MM101–115) [
2].
The most evident effect of dwarfing rootstocks is the reduction in tree height. For example, the ‘M27’ rootstock is one of the most dwarfing apple rootstocks, and it can reduce the height of a tree by up to 80% compared to a seedling rootstock. Conversely, ‘MM111’ only reduces tree size by approximately 20% compared with a seedling rootstock [
3]. Beyond vigor control, rootstocks also provide other agronomical advantages, including variation in different aspects such as precocity [
4,
5], floral induction [
4,
6,
7,
8], fruit set [
3], photosynthetic activity [
9,
10], disease susceptibility [
11], cold tolerance, and resistance to unfavorable soil conditions [
12].
High-density plantings depend on many rootstock traits, especially the dwarfing and precocity attributes, allowing an early return of investment for the initial costs of orchard planting and improving profitability. In young trees, production occurs simultaneously with canopy development. This is possible due to more efficient light utilization, as HDP can increase light interception and distribution within the canopy [
13,
14,
15,
16,
17,
18].
Dwarfing rootstocks have played a central role in training system evolution, leading to the introduction of the spindle bush and the slender spindle in the 1950s and 1960s [
19]. The slender spindle was demonstrated to optimize light interception and canopy light availability, especially in growing sites characterized by low vigor, such as Northern Europe or Alpine regions [
20]. Alternatively, the slender spindle training and pruning methods were not recommended for high-vigor sites because the top part of the tree may become overgrown [
21].
Studies found that many training systems could not guarantee high illumination across the whole canopy, especially early on in the orchard’s lifespan [
22,
23]. For this reason, increasingly planar and 2-D canopies have been developed and evaluated, such as the Geneva Y-trellis [
24], Tatura trellis leading to the Washington Vee trellis [
23], the bi-axis, or other experimental multi-leader trees [
25,
26,
27,
28]. Planar canopy systems are obtained generally, with two (bi-axis) or three to six (multi-leader) main vertical axes oriented in the parallel direction of the row, with numerous short lateral limbs [
25,
26,
27,
28], conferring to the tree a narrow 2-D shape. A more recent concept, the Planar Cordon [
29,
30] or Guyot [
31], uses up to 10 non-branching vertical fruiting stems on a bi-cordon tree. The renewed interest in planar divided-canopy training systems has increased with the availability of preformed, split-branch trees from the nursery, eliminating the need for orchard topping (i.e., heading back the tree) and delayed shoot system formation [
25,
26,
32,
33].
In mature apple trees, it has been shown that dwarfing rootstocks influence the length of the growing period, inducing an early set of terminal buds and reducing the number of nodes present on each shoot [
5,
34,
35,
36]. Rootstocks can affect shoot length by affecting the number of nodes, branching density, position, and other branch characteristics [
36]. In fact, in two different studies [
37,
38], the percentage of bud break in axillary buds on a given axis was unaffected by the selected rootstock. Thus, the number of developing axillary structures per branch seems to depend on the number of nodes developed during the previous year [
39,
40]. For example, Seleznyova et al. [
36] demonstrated that dwarfing rootstocks produced fewer nodes and axillary buds when compared to vigorous rootstocks, which contributed to diminished shoot growth in the following year. The same authors demonstrated that these shoots with reduced nodes also have smaller internode lengths [
36]. Many trials have reported architectural modifications of the scion in the first year after planting due to the dwarfing rootstock’s effects, which include a reduction in shoot length [
41,
42], number of nodes [
39], and number of second-order shoots (i.e., lateral feathers) [
43]. This reduction in scion shoot growth in a growing season is primarily due to the earlier shoot extension termination conferred by dwarfing rootstocks [
44]. Ultimately, it was shown that dwarfing rootstocks affect the type of growth units produced throughout an annual growth cycle, reducing the number of extension growth units, thus influencing the architecture and vigor of the tree [
5]. These dwarfing effects are further enhanced by the dwarfing rootstocks’ precocious transition to flowering to help reduce vegetative vigor through early fruiting induction [
5], causing a greater number of subsequent sympodial shoots, which typically have fewer nodes. Overall, these results suggest that the effect of the rootstock on scion growth is cumulative and superimposed year after year and may begin as early as the first or second year after planting by modifying tree architecture [
4,
5].
The overall purpose of this study was to investigate, at both the nursery level and the first year after planting, how multi-leader trees modify the initial scion architecture with respect to shoot node production and length, along with how these traits affect precocity (e.g., the initial occurrence of flowering). Different rootstocks were also used to observe modifications in vigor, and two propagation methods were tested to assess which was the easiest technique to successfully develop mature trees. In addition, we studied the influence of growing conditions in three different environments: New Zealand and two sites in Italy (Bologna and Trento).
2. Materials and Methods
2.1. Experimental Sites and Design
In 2009, three sites were selected for this trial: the first location was in New Zealand, at the Plant & Food Research Institute Ltd.—Hawke’s Bay Research Centre (HB), (39°39′12.1″ S, 176°52′43.7″ E), and the other two were in Italy (Bologna and Trento). In Italy, the experiment was carried out at the University of Bologna (BO)—Cadriano Experimental farm (44°32′55.1″ N, 11°23′10.0″ E), and at the Fondazione Edmund Mach—Istituto Agrario di San Michele all’Adige (SM) Experimental Farm (46°11′19.1″ N, 11°06′17.3″ E).
The Hawke’s Bay soil was a silt clay loam, fully cultivated, then fumigated with chloropicrin, and aerated before planting in September 2008. The soil in the Bologna site was not fumigated and had no apple or pear planting history before the present trial. The soil was a deep silt clay loam, with optimal levels of potassium and available magnesium, and was plowed in April 2009. The San Michele all’Adige soil was fertile, with an upper profile of 150 cm of sandy clay loam and sand in the deeper strata, rich in organic matter and potassium. The soil was fumigated with metam potassium (Tamifum Eastman Milano, Italia S.r.l.) at the rate of 340 L/ha of commercial product in early March 2009. A study of nursery tree growth was performed to explore the influences of the site of cultivation, the propagation method, the rootstock, and the number of leaders per tree on the early development of tree architecture for apple cultivar ‘Fuji’.
2.2. Fertilization and Irrigation at Experimental Sites
A different fertilization strategy was established for each site in order to optimize the growth according to soil type. In the Hawke’s Bay site, calcium ammonium nitrate (CAN, 27% N) was applied to the soil at a rate of 14 g CAN/m
2 four times monthly, starting from the second week of November 2008 and completing the schedule of fertigation in March 2009 (
Table S1). This resulted in an application of 1.512 g of N per tree per application, for a total of approx. 6 g over the season. In Bologna and San Michele all’Adige, a complex NPK mineral fertilizer (21-7-14) supplemented with calcium nitrate (15.5% N) was distributed over the season (
Table S1). Total amounts of nitrogen per tree were 12.5 and 38.5 g of N per tree in each location, respectively. Trickle irrigation was provided at each site to maintain soil moisture content close to field capacity.
2.3. Tree Production and Planting
For the Hawke’s Bay experiment, three apple rootstocks were grafted with ‘Fuji’ (Nagafu 6). ‘Fuji’ was selected due to the higher vigor of this cultivar compared to other accessions [
45,
46]. One-year-old rooted rootstocks were produced in stool beds in a local nursery, and scions were collected from orchards set up at the Hawke’s Bay Research Centre. Three different rootstocks were used, providing an increasing vigor scale: ‘M27’ (very dwarf), ‘M9’ EMLA (dwarf), ‘MM106’ (semi-vigorous). Two grafting methods were adopted to establish the multi-leader treatments: 50% of the rootstocks were chip-budded with one, two, or three buds [
40,
47], and 50% were bench-grafted with four bud scions (one-year-old wood with four buds to increase opportunities for multi-leader development). During the growing season, we selected in the nursery one, two, or three active shoots as they developed. This design provided 18 treatments in a 3 × 3 × 2 factorial design: three rootstocks, three leader systems (single, double, or triple), and two propagation methods (chip budding and bench grafting). Subsequently, trees were planted in the nursery at a spacing of 3.0 m × 0.5 m (6667 trees/ha) in a completely randomized design with 12 replications (trees) per treatment. Trees are commonly planted at 90–100 cm × 10–15 cm, independently of the rootstock vigor in a nursery. To minimize shading and growth interference from the trees, we quintupled the standard inter-tree within-row spacing to 50 cm, while inter-row spacing was tripled (3.0 m) to reduce light limitations and minimize any border effects. The design was a completely randomized block with 12 blocks containing 1 tree per each of the 18 treatments. The total number of trees in the trial in this specific location was 216 (3 rootstocks × 3 levels of leader × 2 propagation methods × 12 trees per treatment). Grafting height was standardized at 30 cm above the root system for all the trees. Graft unions were protected with tape, and the top of the rootstock/graft was sealed with a pruning wax paste.
Bench-grafted trees were stored in a controlled environment (7–10 °C) with humidity and placed in a wooden bin containing moist untreated soil for at least two weeks to improve the callus tissue formation [
33,
48]. Planting height was the same for all treatments, with the graft union at 10 cm above the ground.
A single nursery supplied all the tree material used in the two Italian sites (Griba Baumschule, Niederthorstraße 9, I-39018 Terlan BZ, Italy). The ‘M27’ rootstocks were two years old because it was not possible to obtain one-year-old stools with appropriate caliper quality due to the low vigor of this rootstock.
Trees were bench-grafted by the Italian nursery during spring 2009 (6 April 2009) and planted in the nursery a few days later (15 April 2009 in San Michele all’Adige and 16 April 2009 in Bologna). A different budding strategy was adopted for chip-budded trees compared to the Hawke’s Bay experiment in Italy. Three scion buds were budded on the rootstocks for all of the multi-leader treatments to standardize the total area of rootstock bark viable from the budded area among the treatments (i.e., there was a lot of damaged bark because of the budding depending on the number of buds). In this way, there was a similar amount of functional bark on the rootstock in the early stage of development, irrespective of budding treatment. Moreover, the rootstock was not headed at budding but approximately a month after planting to improve survival. Trees at the Italian sites were planted with a spacing of 3.0 m × 0.5 m (6667 trees/ha), the same utilized in NZ. A completely randomized block design with 20 replications (trees) in Bologna (BO) and 18 replications (trees) in San Michele all’Adige (SM) per each treatment was used. In more detail, the design in BO was a completely randomized block with 20 blocks, while in SM, there were 18 blocks. In both cases, a block consisted of 1 tree for each of the 18 treatments under investigation.
The total number of trees in BO was 360 (3 rootstocks × 3 levels of leader × 2 propagation methods × 20 trees per treatment), while that in SM was 324 (3 rootstocks × 3 levels of leader × 2 propagation methods × 18 trees per treatment). Unrequired scion growing shoots were removed from respective axis number treatments when they reached the average length of 15 cm, leaving only the required number per rootstock (single, double, or triple leader) for the experiment. Suckers from the rootstocks were removed throughout the growing season.
2.4. Scion Development Measurements
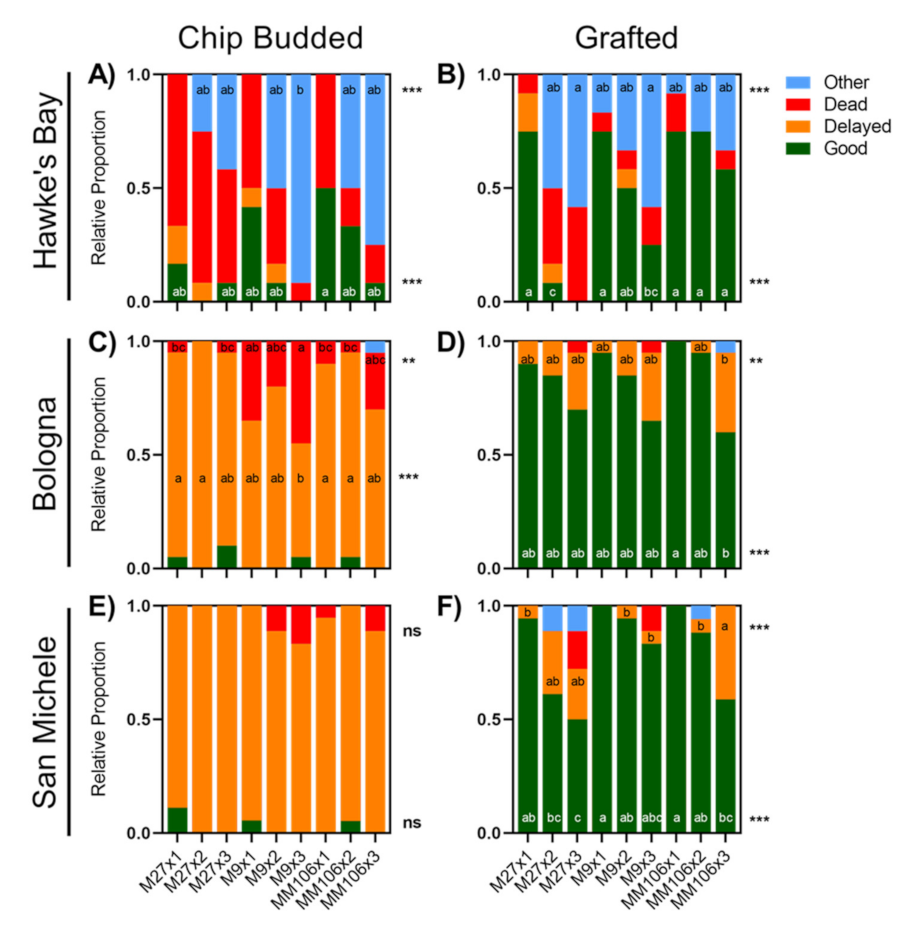
At the end of the growing season, the trees were evaluated for their shoot growth and graft survival. The proportion of trees presenting different levels of development was determined. We classified these trees into four qualitative categories: good (large), delayed (small), dead, and other (wrong number of axes). The proportion of each tree classification was determined for each treatment combination and averaged for treatment comparisons. In the Italian sites, leaves were removed from the scion before leaf abscission to determine the number of leaves and leaf area (cm2) per tree using a leaf area scanner (Li-3100, LI-COR Biotechnology, Lincoln, NE, USA). No leaf measurements were carried out at the Hawke’s Bay site.
In each site, tree development was measured using a coding method described by Costes et al. [
49], which provides the indications to describe different orders of shoot axes, the node they originated from, and their position. Shoot node number, length (cm), and cross-sectional area (cm
2) were also measured. Shoot length (cm) and node numbers were summed across single and multi-leader trees to evaluate total tree growth. Axis cross-sectional area (ACSA) (cm
2) of each primary axis and shoot cross-sectional area (SCSA) (cm
2) of secondary axes (feathers) were calculated. ACSA was recorded by measuring the axis diameter two cm above the graft, while the SCSA measured the diameters of shoots two cm above their insertion. The average internode length (cm) of shoots was calculated by dividing the length of shoots by the number of nodes. Axillary bud break proportion (feather production) was calculated by dividing the total number of secondary shoots by the total number of bud nodes developed over the first vegetative season.
Within each leader, it was possible to discriminate single growth units (GU) [
50]—which are sections of shoot developed during a single growth flush—and multiple growth units. The latter case occurred when the shoot apical meristem temporarily stopped growing or slowed its activity and resumed later. To identify GUs, morphological markers as rings of bud-scale scars or compressed internodes were used [
36]. Floral cluster number/tree was counted at full bloom, which occurred on 12 October 2009 in HB, 16 April 2010 in BO, and 29 April 2010 in SM.
2.5. Statistical Analysis
Data were analyzed using the statistical package Statistica (version 9, StatSoft, Tulsa, OK, USA). Data were first checked for homogeneity of variance and a normal distribution. ANOVAs were run to evaluate statistical significance at p < 0.05. Data for each site and for each rootstock × axis number combination were first analyzed separately using a general linear model; all the data were then analyzed to study the effect of each variable of the factorial design (three sites, three rootstocks, three-axis numbers, two propagation methods). Relative proportions of tree status were evaluated amongst treatments, with only the first two dominant classes (e.g., good, other, delayed, dead) from each site × propagation method being analyzed by HSD Tukey’s mean comparisons. For significant effects or interactions, comparisons of treatment means were made using HSD Tukey’s test, and different letters were assigned for differing means based on this test.
5. Conclusions
The wide range of training systems proposed for apple cultivation in the last few decades share some common features: the rapid achievement of high light interception and early cropping, the preservation of optimal light distribution within the canopy over the season and orchard lifespan, and more efficient harvesting due to canopy size control and planar architectures.
The data presented in this research at the nursery level indicate that multi-leader trees seem to be very suitable for guaranteeing rapid canopy development and subsequent good light interception. With three rootstocks of differing vigor, we determined larger total canopy development on newly grafted ‘Fuji’ trees with multiple leaders compared to single-leader trees. This finding highlights the need for specific rootstock trials to optimize the multi-leader systems.
Further, the cultivation site played an important role in modulating early tree performance. The production of second-order shoots was effective in the Italian sites, while no growth of feathers was observed in the New Zealand site. Multi-leader trees reduced their average shoot length in the Italian sites in the first year after grafting. The number of shoots developed on multi-leader trees was higher than in single-axis trees at the same planting density. This could be of considerable benefit to growers, who can use canopy architecture manipulation (e.g., increased number of axes) as a tool for vigor control when their site of cultivation and/or rootstock selection is too vigorous to easily establish HDP orchards, as well as the newer concepts for 2-D planar fruiting walls.

 ,
, 


{kind=link}