1. Introduction
The pitaya (
Hylocereus spp.) is an emerging fruit crop of the
Hylocereus genus in the family Cactaceae. It is a climbing cactus that is native to the rainforests of northern South America, Central America, and Mexico [
1]. Pitaya has been cultivated economically for >30 years, and it is currently a global economic fruit crop primarily cultivated in Southeast Asian countries, including Vietnam, Taiwan, Indonesia, Malaysia, and China’s Guangxi and Hainan provinces [
2,
3]. In addition, it is also cultivated at a small scale in Israel, Nicaragua, Colombia, and Mexico. In recent years, with the northward shift of production locations, some subtropical regions with higher latitudes, including Guizhou in China and Kagoshima, Okinawa, and Fukuoka in Japan, have also started cultivating red-fleshed pitaya [
3,
4].
The optimal day/night reproductive temperature of pitaya (
H. undatus) is 30 °C/20 °C [
5]. Temperature stress is detrimental to shoot (cladode) development, flower bud formation, and fruit set, leading to reduced production [
5,
6,
7]. Although pitaya is widely distributed in its area of production, it is less tolerant to temperature stress than other cactus crops, potentially owing to its indigeneity to rainforest environments [
5,
8,
9]. In Guangxi and Guizhou in China, where winter temperatures reach 0 °C, chilling injury may occur in shoots; thus, only spring sprouts can be cultivated for autumn fruit production [
3]. Under times of prolonged daylight in winter, warm temperatures (32 °C/22 °C) promote flowering in the early spring or in the production period [
10], whereas high temperature (HT) reduces flowering batches and flower production [
5,
8]. In Israel, high summer temperatures of up to 38 °C result in the production of 3–4 waves of flowers (a plant flowers 3–4 times during the reproductive period), and in some areas of extreme heat (44–46 °C), only one wave of flowers can be produced, in September, resulting in significantly reduced production [
5].
“Da Hong” is a major variety of red-fleshed pitaya (
H. polyrhizus) cultivated in Taiwan and Southeast Asia with its self-compatibility, large fruit, high yield, and good flavor [
6,
11,
12,
13]. Although its flowering waves are unaffected by high summer temperatures [
7], an increase in the proportion of small fruits (<150 g) under high summer temperatures (>35 °C/25 °C) is often observed in open fields or netting facilities [
6,
7].
Chu and Chang [
6] confirmed that the fruit set of red-fleshed pitaya is affected by HT, resulting in the occurrence of small fruit size, by transferring plants into phytotrons and subjecting them to short-term HT treatment (40 °C/30 °C). Compared with the findings at 30 °C/20 °C, HT shortened the time to flower development, delayed flower opening and closing, reduced flower length, caused severe fruit drop 1 week after flowering, delayed the development of few fruit sets, and resulted in a seed weight of <1 g. In addition, although the shoots maintain a higher level of yellowing under high temperatures, the dry matter weight does not decrease, indicating that shoot yellowing under HT does not obstruct fruit set [
6]. The optimum temperature (OT) for in-vitro pollen germination in the red-fleshed pitaya “DR3” and white-flesh pitaya (
H. undatus) “Vietnam” is 25 °C [
14], and it is speculated that HT may have reduced pollen viability and caused poor fertilization, resulting in less fruiting because of poor fertilization [
6]. However, it remains unclear whether HT stress during flower development obstructs floral organ fertility.
During the peak of summer (June–August) in Taiwan, the day and night temperatures are close to 35 °C and 25 °C, respectively [
15]. Under this environment of high light intensity and elevated temperature, the sun-exposed shoots at the adaxial end often burn and exhibit yellowing, and the yellowed shoots naturally re-green in autumn (October) when the temperature decreases [
15]. During HT, a 50% sunshade net placed above the canopy can lower the temperature and reduce the yellowing of shoots [
15,
16,
17,
18]. However, compared with the findings in an open field, the yellowing of shoot in plants cultivated in a netting facility is not improved even though the light intensity is reduced, owing to the increase in temperature [
15]. The same response has been observed under the HT, the phytotorns in which the shoot at the adaxial end of the plants maintained a higher degree of yellowing after being moved from open-field (35 °C/25 °C) to HT (40 °C/30 °C) conditions in the phytotrons [
6]. Both a slightly elevated temperature (35 °C/25 °C) and HT (40 °C/30 °C) under bright light lead to shoot yellowing, which does not affect their dry weight [
6]; however, the effect of this yellowing on the photosynthetic efficiency of shoot needs to be further investigated.
When a healthy plant is exposed to stress, electron transport in the photosynthetic system is inhibited, causing chlorophyll fluorescence to rapidly decline. Therefore, changes in chlorophyll fluorescence reflect the photochemical efficiency, and these changes are often used to document the extent of damage to the photosystem caused by stress [
19]. The measurement of the maximum photochemical efficiency (Fv/Fm) of photosystem II (PSII) under dark-adapted conditions can be used as an indicator of temperature stress, light stress, and water stress [
19]. In several tropical fruit trees, such as pineapple, sugar apple, guava, and papaya trees, chlorophyll fluorescence can be used as an indicator of temperature stress [
20,
21,
22], but studies on red-fleshed pitaya are rare.
This study aims to extend the results of a previous study on HT-induced effect of suppressing fruiting in “Da Hong” red-fleshed pitaya [
6], which used potted plants with open-field cultivation as the study material and then transferred them to phytotrons simulating day/night temperatures of 40 °C/30 °C, 35 °C/25 °C, and 30 °C/20 °C, respectively. Through examinations of in-vitro pollen viability and self-pollination or mutual cross combinations under three temperature regimes, the effects of high-temperature stress on stamen and pistil fertility and fruit set in “Da Hong” red-fleshed pitaya have been investigated to understand the main causes of poor fruit set under HT. In addition, we also investigated the re-greening of shoots with different degrees of yellowing after treatments, particularly the sun-exposed side of triangular fleshy shoots, and assessed their PSII efficiency after treatment to evaluate the effect of short-term HT stress on the photochemical efficiency of photosynthesis to further understand the physiology of photosynthesis.
2. Materials and Methods
2.1. Plant Materials and Pre-Treatments
The red-fleshed pitaya “Da Hong,” which is sensitive to HT, was selected as the plant material, and the study was performed in the Kaohsiung District Agricultural Research and Extension Station (KDARES), Changzhi Township, Pingtung, southern Taiwan (22°41′ N, 120°31′ E) using 2–3-year-old plants, which were planted in cutting pairs in 15-L pots supported by bamboo poles. The plants were well irrigated, and a soluble fertilizer (N:P:K = 15:15:15) was added to the irrigation water once a week to maintain good plant development. Considering the space limitation of the phytotron (3.8 m × 2.4 m), five pots of plants were used in each treatment, and pots of uniform growth potential, including a total of 10–12 mature fruiting shoots in each pot, were selected.
To avoid overload in plants caused by fruiting, the flower buds that emerged before the beginning of the study were removed. The number of flowers left in each pot was adjusted to 5–6 flowers during the study, and the ratio of shoots to fruits was adjusted to 2:1. The study was conducted from July to September in 2017. Owing to the high average temperature (28.7 °C) and high light intensity from May to June, the sun-exposed shoots of the experimented plants displayed some yellowing before the experiment.
Temperature treatments were simulated as HT (40 °C/30 °C), moderately high temperature (MH, 35 °C/25 °C), and OT (served as control, 30 °C/20 °C) (temperature fluctuation of ±1 °C permitted for each condition). Potted plants initially grown in the field were transferred into the phytotrons at KDARES within 1–3 days after the emergence of the flower bud and subsequently transferred to OT on 13 July 2017 and to HT and MH on 21 July 2017.
2.2. Time to Flower Development and Flower Traits
Nine flowers per treatment were examined for the flower buds that emerged on 16 July or 20 July 2017, and the number of days between floral bud emergence, and flowering was investigated. The full length of the flower and diameter and circumference of the ovary were measured at 12:00 and 17:00 on the day of flowering.
2.3. In-Itro Pollen Viability Performance of Fresh Pollen
2.3.1. Preparation of Pollen Culture Medium
Pollen culture was performed using the modified Brewbacker and Kwack (BK) medium described by Weiss et al. [
23] and Cheng and Weng [
14]. The liquid medium contained 100 mg/L each of H
3BO
3, Ca(NO
3)
2·4H
2O, MgSO
4·7H
2O, and KNO
3. The sucrose concentration was 25%, and the pH was adjusted to 5.5 ± 0.1. The solid medium contained 100 mg/L each of H
3BO
3, Ca(NO
3)
2·4H
2O, MgSO
4·7H
2O, and KNO
3, as well as 1% agar and 20% sucrose. Its pH was adjusted to 5.5 ± 0.1, and the medium was sterilized at 121 °C and 1.2 kg/cm
2 for 20 min. It was then poured into Petri dishes and refrigerated at 4 °C for use.
2.3.2. Pollen Culture and Observation
Fresh pollen was collected from the flowers that opened on the same day, and six flowers were collected for each treatment as replicates. The whole flowers were picked at 20:00 on the day of flowering. Ten anthers were randomly obtained from each flower and placed in a 1.5-mL micro centrifuge tube. After adding the liquid medium, the pollen and liquid medium were mixed well by vortexing the microcentrifuge tubes for 10 s on a touch basis. After the anthers were removed, 1 mL of liquid medium containing pollen was extracted with a dropper and placed on the solid medium. After 24 h of incubation, pollen germination was observed with an optical microscope (Eclipse 50i, Nikon, Japan). Three hundred pollen grains were examined in each replicate, and pollen tubes longer than twice the length of the pollen were considered as viable pollen.
2.4. Investigation of Artificial Pollination Combinations, Fruit Set and Fruit Characteristics
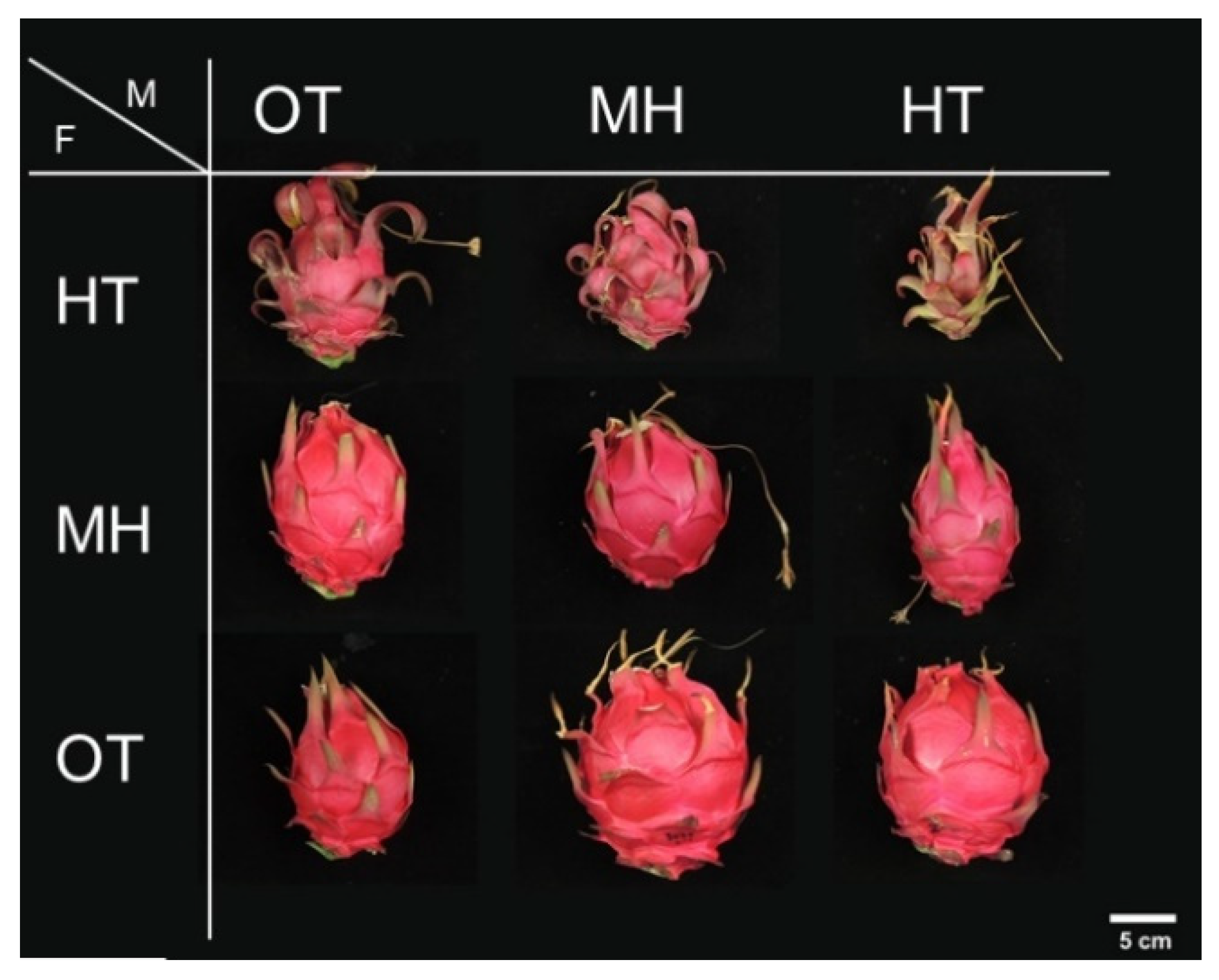
For the experimental plants that flowered from 30 July to 6 August 2017, the pollination of pollen and pistil under three temperature treatments was performed using self- or mutual cross pollinations by hand, including the following nine combinations: OT self-pollination, OT(♀) × MH(♂), OT(♀) × HT(♂), MH self-pollination, MH(♀) × OT(♂), MH(♀) × HT(♂), HT self-pollination, HT(♀) × OT(♂), and HT(♀) × MH(♂).
Each combination was pollinated using one-to-six flowers. Excluding the three self-pollination combinations, the maternal stigma was separated from the stamens before 13:00 on the day of flowering. The stigma was then covered with a paper bag to avoid self-pollination. Pollination was performed at 20:00–21:00 at night, and fresh pollen was collected on the same day of flowering. The stigma was brushed with an ink brush to make sure it was evenly covered with pollen for sufficient pollination before covering it with a paper bag, and the paper bags were removed 3 days after flowering. Fruit set was observed 1 week after flowering.
The fruit characteristics of each pollination combination were investigated at approximately 38 days after fruit development with a ripeness code of 819 [
13], including individual fruit weight, fruit transversal diameter, fruit longitudinal diameter, fruit flesh weight, fruit peel weight, edible rate, fruit peel thickness, total soluble solid content (TSSC) in the core of the fruit, and seed weight. The color difference between the sun-exposed and shaded sides of the fruit equator was investigated using a color spectrometer (ColorLite sph860; Katlenburg-Lindau, Germany), and the a* value was used to represent the degree of color change of the fruit.
2.5. Greening of Shoots and Chlorophyll Fluorescence
2.5.1. Labeling of the Investigated Parts
The plants were divided into the adaxial (the sun-exposed canopy) and abaxial ends (the parts where shoots hang down). As pitaya has a triangular fleshy shoot with the other side facing the sun, 1–3 parts of each treated plant with a similar degree of yellowing as the sun-exposed side were marked (
Figure 1) for investigation before and during the experiment. The data of the marked parts on the single plant were averaged as the data of the single plant. Therefore, five replicates of data were included for the statistical analysis of each treatment, and the degree of yellowing of the shoots before and after the experiment was recorded by capturing photographs.
2.5.2. Investigation of the Re-Greening of Yellowed Shoots
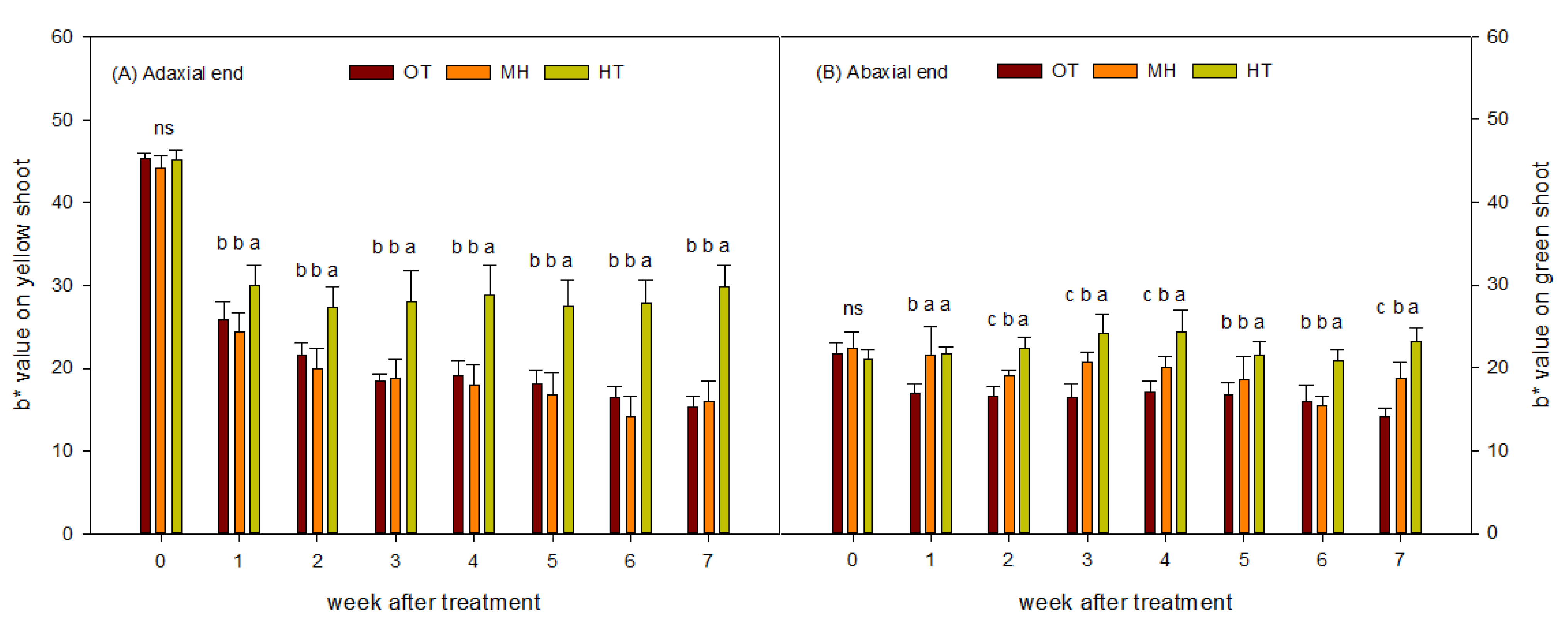
The shoot color at the adaxial and abaxial ends was measured using a color spectrometer (ColorLite sph860, Katlenburg-Lindau, Germany) weekly before and during the experiment period. The b* value was used to indicate the yellowing or re-greening of the shoots.
2.5.3. Chlorophyll Fluorescence Performance of Shoots
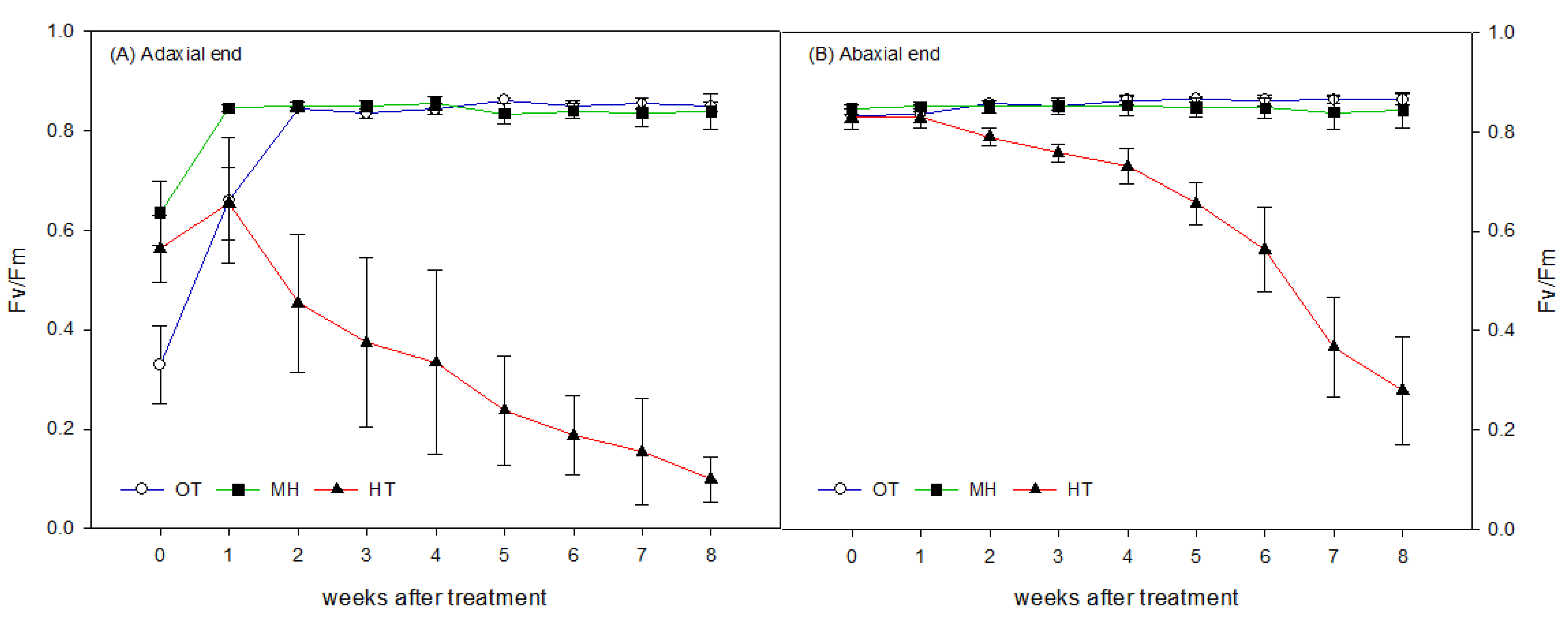
Fv/Fm of PSII was measured weekly using a pulse amplitude-modulated fluorimeter (Mini-Pam, WALZ. Effeltrich, Germany) before and after the experiment to estimate the degree of photoinhibition in the plants during the study. As it was difficult to dark-adapt the shoots of red-fleshed pitaya with leaf clips, measurements were performed at 20:00–21:00 instead of at dark. Fv/Fm of PSII was calculated by measuring Fo and Fm using measuring the light and saturating pulse [
24].
2.6. Experimental Design and Statistical Analysis
The data were analyzed using SigmaPlot 14 software (Systat Software, Inc., San Jose, CA, USA), and graphs were plotted. One-way analysis of variance was used for the test data, and the least significant difference (LSD) was used to analyze whether the difference in a pairwise comparison of two treatments were significant. p < 0.05 indicated a statistically significant difference.
4. Discussion
The effects of HT stress on flowering and fruit set included delayed or early flowering, dichogamy of pistil and androecium development, defects in floral tissues (e.g., developmental failure of the ovule and ovary, obstructed anther dehiscence), and gamete defects (e.g., reduced pollen viability, poor pollen tube growth), all of which directly affect crop yield and quality [
25]. In general, C3 or C4 plants, such as cotton, have increased stomatal conductance at HT, which helps them dissipate heat and reduce leaf temperature [
26]. However, the pitaya is a crassulacean acid metabolism (CAM) plant in which the stomata of shoots are closed the during daytime [
5], resulting in the inability to dissipate heat during HT stress. When the temperature at noon is over 35 °C, the surface temperature of red-fleshed pitaya shoots can exceed above 50 °C [
7]; thus, HT stress can cause damage in red-fleshed pitaya during the summer season.
In this study, HT treatment (40 °C/30 °C) shortened the time to flower development, and the number of days required for flower development after flower emergence decreased significantly with increasing temperature. This trend was similar to the results of a previous study [
6], validating our results. A variety of
Oryza sativa subsp.
indica, IR64, also exhibits similar performance under HT, in which the proportion of early flowering spikelets is increased when the temperature reaches 36.2 °C compared with that at 29.2 °C [
27], suggesting that the timing of “Da Hong” flowering can be accelerated when the accumulated temperature is increased by HT treatment.
The flowers of the “Da Hong” continue to develop on the day of flowering. HT reduced flower length, which is consistent with the results of a previous study [
6]. Although the flowers under MH treatment opened 5 days earlier, the flower length, ovary diameter, and ovary perimeter on the day of flowering were not significantly different from those under OT treatment, indicating that MH accelerated flower development without affecting flower characteristics.
The in-vitro pollen germination rate was significantly lower under MH or HT than under OT, confirming that pollen viability during flower development is significantly reduced if the temperature exceeds 35 °C/25 °C even though MH did not affect flower characteristics. Wu [
28] preliminarily reported that the collection of “Da Hong” pollen on the day of flowering, thus, incubation at 25 °C resulted in the highest in-vitro germination rate of 61%. When the incubation temperature was 30–40 °C, the in-vitro germination rate decreased, but the pollen viability was approximately 10–22%, which was higher than that under MH and HT at current study, indicating that continuous HT treatment during flower development has a greater impact on the floral organ fertility of the red-fleshed pitaya than HT only on the day of flowering, which is similar to the observation in cherimoya [
29]. Six days after floral bud emergence in the pitaya, the mother cells of pollen entered meiosis to form microspores and then underwent mitosis, and by 2–3 days before flowering, three-celled pollen grains developed [
30]. Müller and Rieu [
31] noted that the meiotic and mitotic phases of pollen development are sensitive to HT, which can lead to abnormal chromosome segregation. The tapetum, which supports pollen development, is also affected by HT. Specifically, the tapetum becomes defective under HT, making pollen less fertile [
31]. No studies to date have explored the effects of HT on pollen development and fertility in red-fleshed pitaya. However, it can be tentatively concluded that HT treatment during flower development may affect the meiosis of microspore mother cells during the formation of pollen grains. Therefore, this is more likely to reduce pollen fertility than HT only on the day of flowering.
The function of the pistil could be initially assessed by post-pollination fruit set, individual fruit weight, and seed weight. In this study, the results indicated that under HT, the pistil maintains relatively good functioning (
Table 2). After pollination with more viable pollen, fertilization success was enhanced, and seed weight was increased to stimulate fruit development. Under MH, even when HT pollen with extremely low viability was pollinated, fruit set remained high, but individual fruit/seed weight was not substantially improved. If self-pollination or pollination with OT pollen was performed, the individual fruit weight could be increased to >390 g. Additionally, there were no significant differences in fruit set, individual fruit weight, and seed weight between the pollination of HT pollen with MH and OT stigma; thus, it can be assumed that MH treatment does not affect the function of the pistil.
The pollen viability of the papaya variety “Wonder Flare” fell to 0% when the average temperature reached 28.2–30.5 °C from July to September, but for plants from open-field cultivation and those with more viable pollen (35%), the fruit set, fruit weight, and number of seeds were increased [
32], suggesting that papaya and “Da Hong” pitaya behave similarly under HT, with the function of the pistil being less affected than that of the stamen.
The results of in-vitro pollen germination and pollination combinations at various temperature treatments confirmed the previous hypothesis that HT reduces pollen viability, resulting in poor fertilization and small fruit size in “Da Hong” red-fleshed pitaya [
6]. Metz et al. [
33] revealed that the in-vitro germination rate of “clone C” red-fleshed pitaya decreased to <11% when pollen was stored for 3 months, but when the pollen intensity (pollen amount) was sufficient, the pollen remained able to promote fruit set by artificial pollination. Therefore, although pollen viability was significantly reduced under MH treatment, the function of the pistil was less affected by HT treatment. In this study, even if pollens with lower viability were used for artificial pollination; however, pollen fully covering the stigma would be sufficient to stimulate normal fruit set and development. Therefore, when pollen viability is reduced by HT or storage conditions, fruit set can be promoted if the sufficient pollination, is provided.
The seed weight of all fruit under the HT condition was similar to that reported by Chu and Chang [
6]. However, fruit set and seed weight could be restored for HT pistils pollinated with MH or OT, indicating that the increase in seed weight (number) both increased the fruit weight and ensured fruit set. In papaya, 70% of flowers can set fruit and form seedless fruits even without artificial pollination at HT [
32]. Tomatoes can activate parthenocarpy at HT [
34]; however, “Da Hong” pitaya does not undergo parthenocarpy in its natural state; thus, it can be inferred that a certain number of seeds must be produced after pollination for fruit set and normal development to proceed, similar to stone fruit [
35].
Our previous study confirmed that HT delays fruit development, results in poor color changes in fruit appearance, and induces the development of sharply downward-curving bracts [
6]. Similar characteristics were observed in fruit developed under HT in the present study. The flesh of “Da Hong” pitaya continues to develop after fruit set until harvest, whereas the development of the peel is less significant. As the fruit matures, chlorophyll degradation and betalain formation will cause the peel to gradually change from green to red in color and to become thin because of the reduced water content; thus, the flesh formation rate continues to increase [
17,
36,
37]. The coloring of the fruit peel of red-fleshed pitaya is related to the degree of bag shading, in which the a* value and coloration improve with increases in the degree of shading, whereas HT increases the thickness of the fruit peel and bracts [
38]. During the study, the fruit of all treatments were not bagged. However, the poor color change in the fruit peel in the HT self-pollination group was more significant than that for the cross-pollination of HT pistils under MH and OT pollen treatments, the peel thickness on the sun-exposed side was significantly higher than that of the other treatments, and the edibility rate was lower than that of the other treatments, indicating that the poor fertilization caused by HT resulted in both delayed fruit development and extremely low flesh growth, leading to higher peel thickness and changes in coloration.
According to Zinn et al. [
25] and Liu [
39], HT affects the flowering and fruiting of plants and damages photosynthetic tissues, thereby inhibiting photosynthesis. HT stress decreases the activity of the oxygen-evolving complex of PSII, which disrupts the structure of the thylakoid membrane, resulting in blocked electron transport. By measuring chlorophyll fluorescence, we could assess Fv/Fm, through which the photosynthetic efficiency of plants subjected to different temperature stresses could be assessed.
Short-term HT stress (45 °C/35 °C) for 7–10 days in white-fleshed pitaya resulted in reduced PSII activity and net CO
2 uptake [
24]. After 10 weeks under HT treatment, the daily variation of net CO
2 uptake was significantly lower than those under OT and MH, and the yellowing and gangrene of shoots occurred. Prolonged HT stress for 19 weeks resulted in a lower dry plant weight, and gangrene of the shoots was not reversed after lowering the temperature [
5]. However, Chu and Chang [
6] reported that when entire red-fleshed pitaya plants in pots were placed under HT for 8 weeks, only the sun-exposed shoots at the adaxial end maintained a higher degree of yellowing without a significant reduction in dry matter weight, indicating that short-term HT stress does not affect the carbon supply of shoots.
Before plants were transferred into phytotrons in this study, the adaxial end of potted red-fleshed pitaya plants displayed greater yellowing than the abaxial end. After 1 week of treatment, the b* value of the adaxial end was significantly decreased, indicating that the shoots started to re-green, and the effect was more significant under OT and MH than under HT. These findings are identical to the results of a previous study [
6]. After treatment of the abaxial end, only the b* values under OT and MH decreased, and re-greening was observed; however, the degree of re-greening under OT was higher. The average temperature of the HT environment, average low temperature, and average temperature of the open field before the experiment were 34 °C, 24.9 °C, and 28.7 °C, respectively, which were close to the temperatures of MH treatment. The light penetration rate in the phytotrons was reduced by approximately 20–30%. After the plants were moved to the phytotrons, the re-greening performance of the adaxial and abaxial ends of the MH-treated plants suggested that high light intensity is the main cause of shoot yellowing in summer. Under HT, the higher light intensity received at the adaxial end of shoot maintained a higher level of yellowing, indicating that light and HT have a synergistic effect on the yellowing of shoots.
The PSII efficiency of the adaxial end of the plant before treatment was lower than that of the abaxial end, whereas the PSII efficiency of the abaxial end was close to the constant of 0.83 for healthy plants. The adaxial end displayed an upward trend 1 week after treatment, indicating that the lower PSII efficiency of the adaxial end before treatment was influenced by high light intensity. In comparison with the re-greening of the shoots, it was assumed that the yellowing of the adaxial end is a temporary phenomenon under high light intensity that does not damage the cells of the fleshy stem.
In this study, we obtained the same results as Chu and Chang [
6], who reported the inhibition of shoot yellowing and re-greening on the sun-exposed shoots under HT accompanied by decreasing PSII efficiency, and the response of PSII to HT stress became negative approximately 2 weeks after treatment. The PSII efficiency of the HT sun-exposed adaxial end increased because of shading after a week of treatment but then began to decline in the second week before continuing to decline until the end of the study. The HT abaxial end also displayed a similar trend. Although the yellowing rate of the MT abaxial end shoots was slightly higher than that of the OT shoots, this did not affect the PSII performance. The PSII efficiency under HT can be understood from the fact that the efficiency gradually decreased after 2 weeks under HT and dropped to <0.6 after 4 weeks and <0.4 after 8 weeks of treatment. However, the plants still did not exhibit gangrene. As the shoots of red-fleshed pitaya are fleshy stems with three sides, the photosynthetic tissues were damaged to different degrees under HT, and only the sun-exposed side exhibited yellowing and photoinhibition [
6]. This may explain the reason why short-term HT did not reduce the dry matter weight of shoots as reported by Chu and Chang [
6].
The HT-treated plants flowered and completed pollination and fertilization in approximately 2 weeks after treatment, and then the PSII efficiency began to decrease. This indicated that the development and fertility of floral organs are first inhibited under HT, and then the photosynthetic capacity of sun-exposed shoots decreases with increasing HT stress time, confirming that the occurrence of small fruit is mainly caused by HT-induced reductions of pollen fertility. This leads to poor fertilization and fewer seeds, and providing sufficient and viable pollen can increase the number of seeds and fruit weight under HT treatment. Therefore, the short-term reduction of PSII efficiency under HT does not affect fruit development. However, if the duration of HT treatment increases, the shoots would develop continuous and prolonged photoinhibition or even gangrene [
15], which would not exclude the reduction of assimilate accumulation and could even affect the subsequent flower bud emergence and fruit set. The effects of long-term HT stress on red-fleshed pitaya flowering and fruit set to be further investigated.
The effects of MH and HT groups on floral development, floral organ fertility, shoot yellowing, and PSII efficiency can be understood from the differences in HT tolerance among organs, such as the reduction of pollen fertility under MH. The PSII efficiency of shoots decreased only when the temperature is increased to 40 °C/30 °C for 2 weeks. Moreover, the extent to which the function of the pistil or stigma–pollen interaction is affected by HT may be somewhere in between the two, but follow-up study is required for validation.








{kind=link}
{kind=link}
{kind=link}
{kind=link}