
Industrial Hemp (Cannabis sativa L.) Varieties and Seed Pre-Treatments Affect Seed Germination and Early Growth of Seedlings

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Petri Dish Germination Assay (Experiment 1)
2.3. Glasshouse Germination Assay (Experiment 2)
2.4. Seed Pre-Treatments (Experiment 3)
2.5. Data Collection
2.6. Statistical Analysis
3. Results
3.1. Germination Response in Petri Dish Assay (Experiment 1)
3.2. Germination Response in the Glasshouse (Experiment 2)
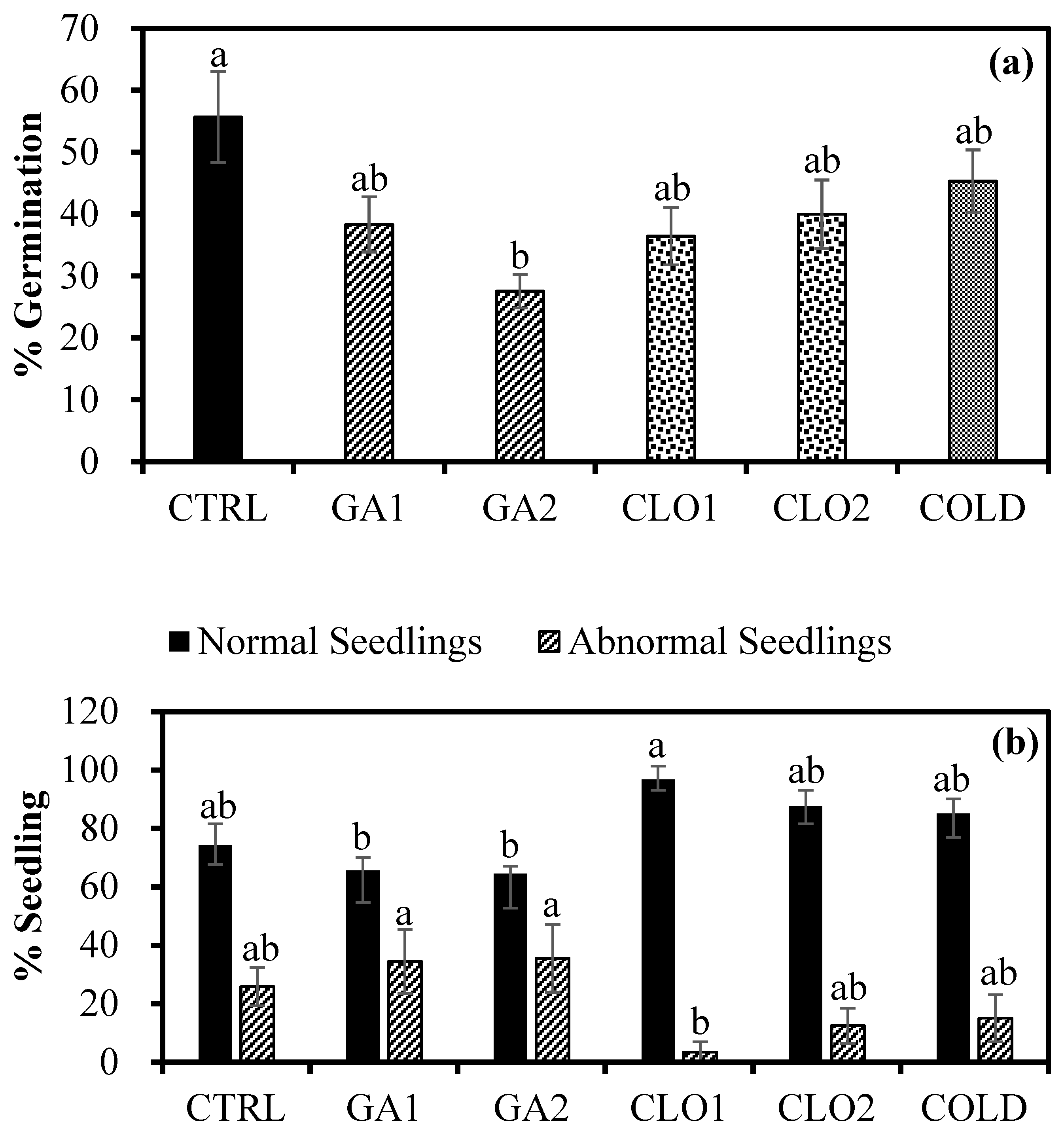
3.3. Germination Response after Seed Pre-Treatments (Experiment 3)
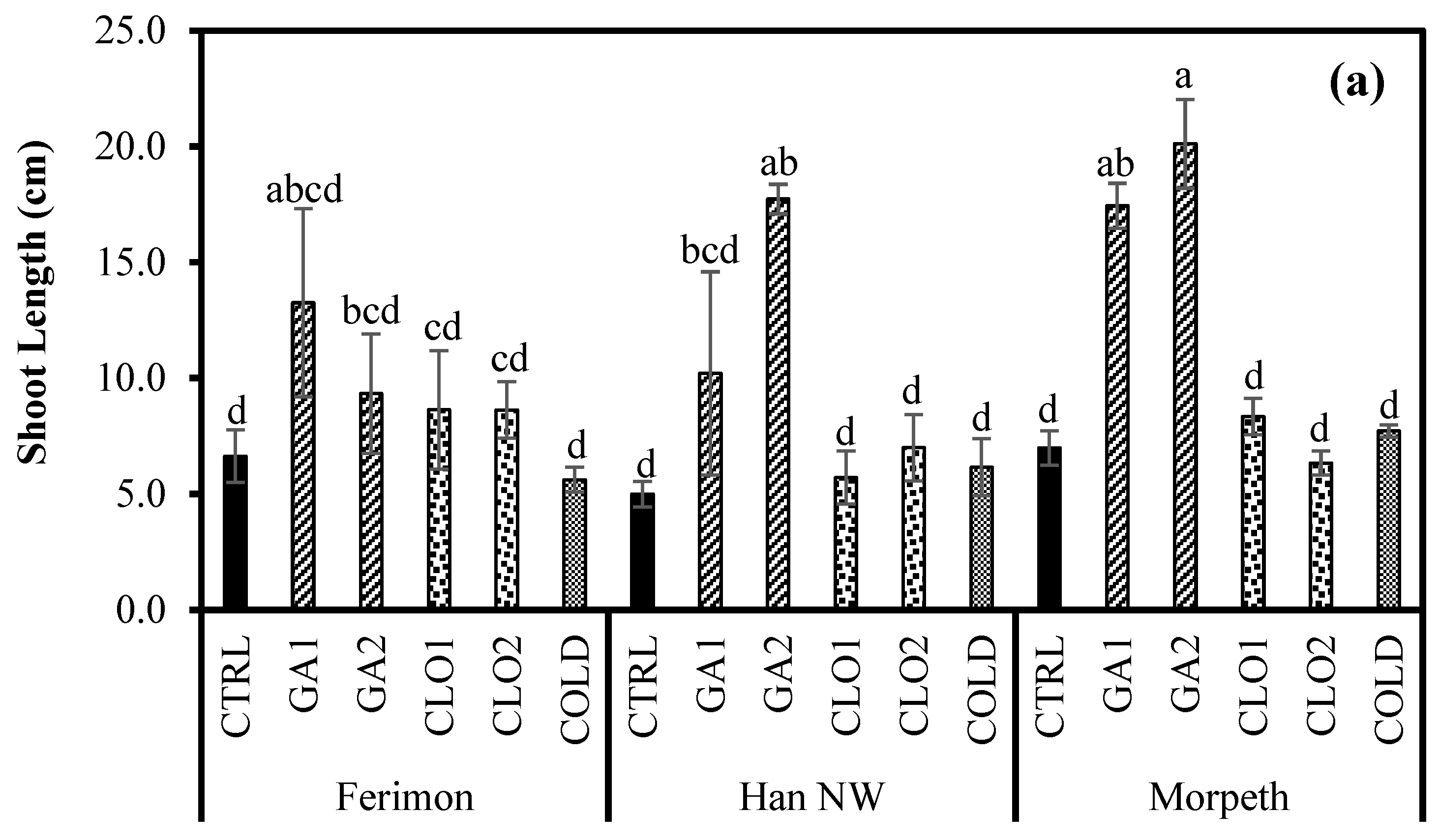
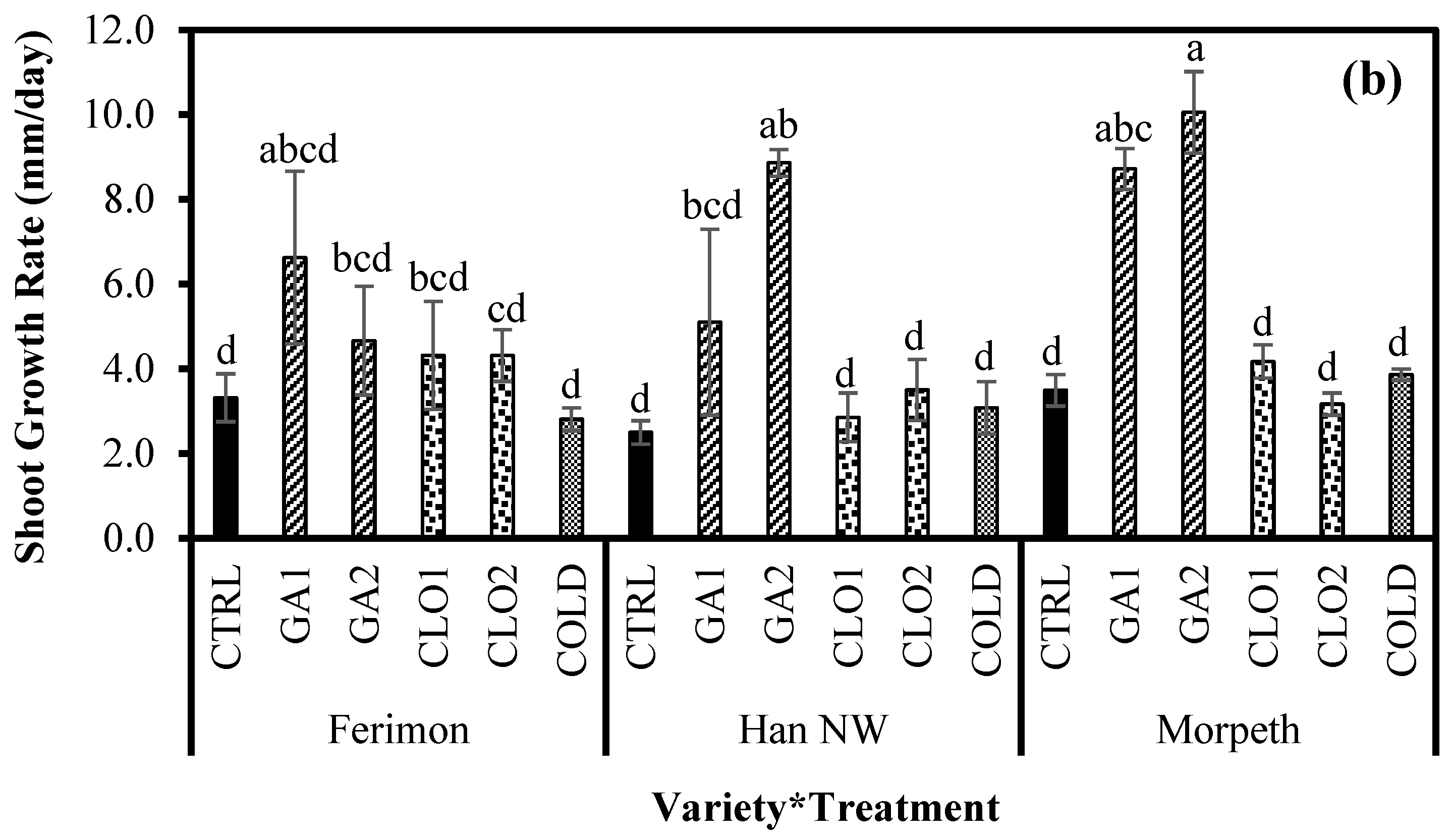
3.4. Seedling Growth Response in the Glasshouse (Experiment 2)
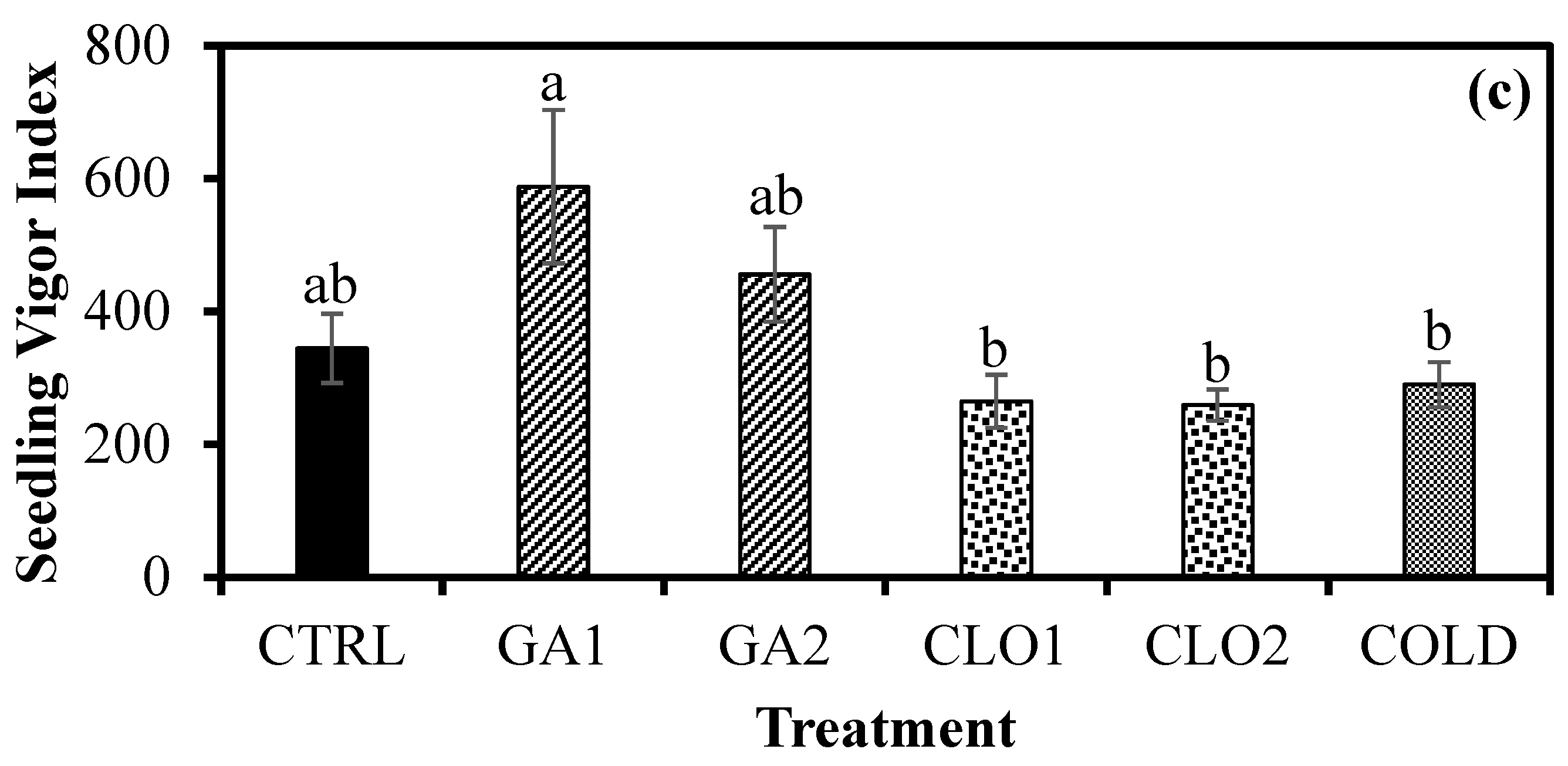
3.5. Seedling Growth Response after Seed Pre-Treatments (Experiment 3)
3.6. Root Growth Response in the Glasshouse (Experiment 2)
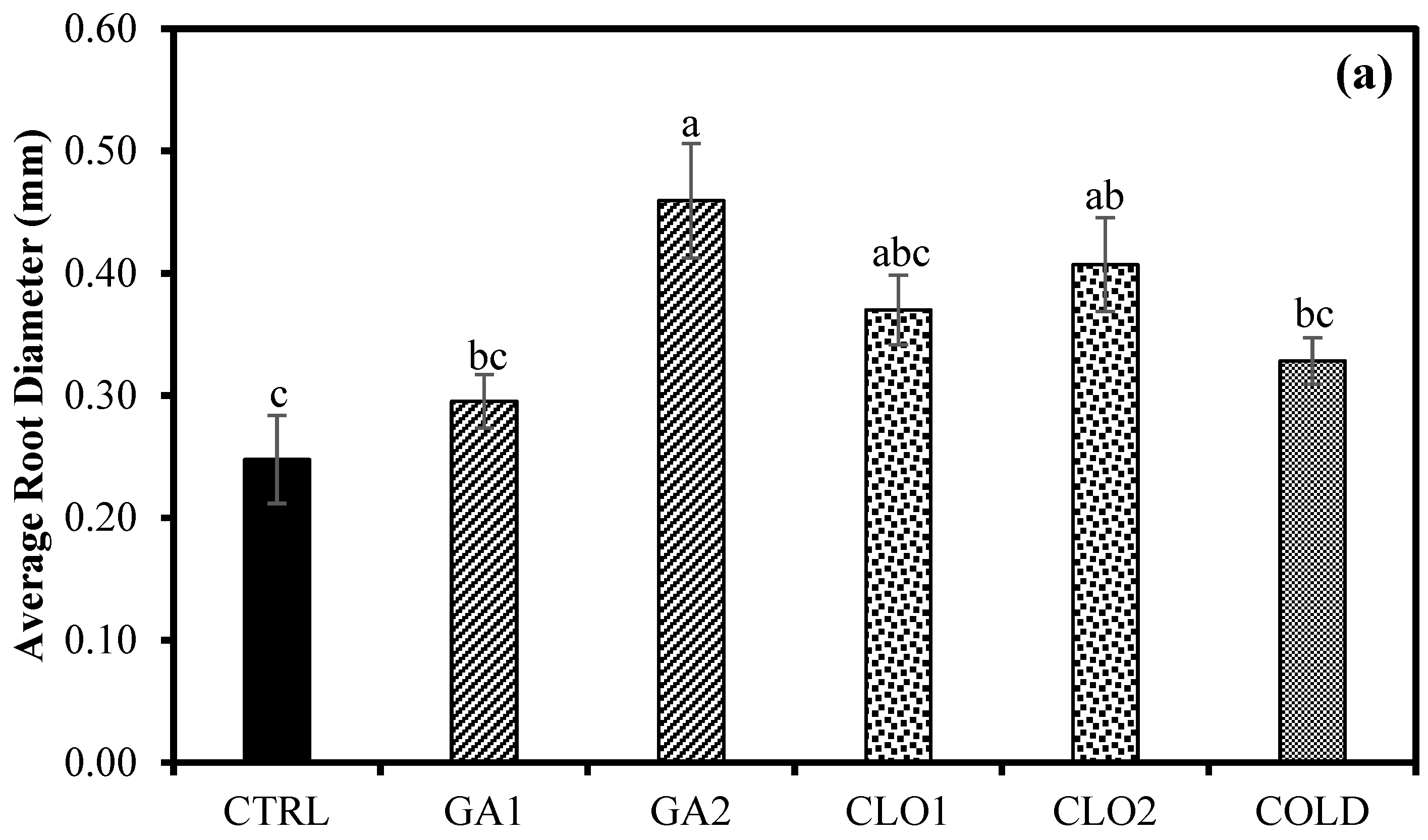
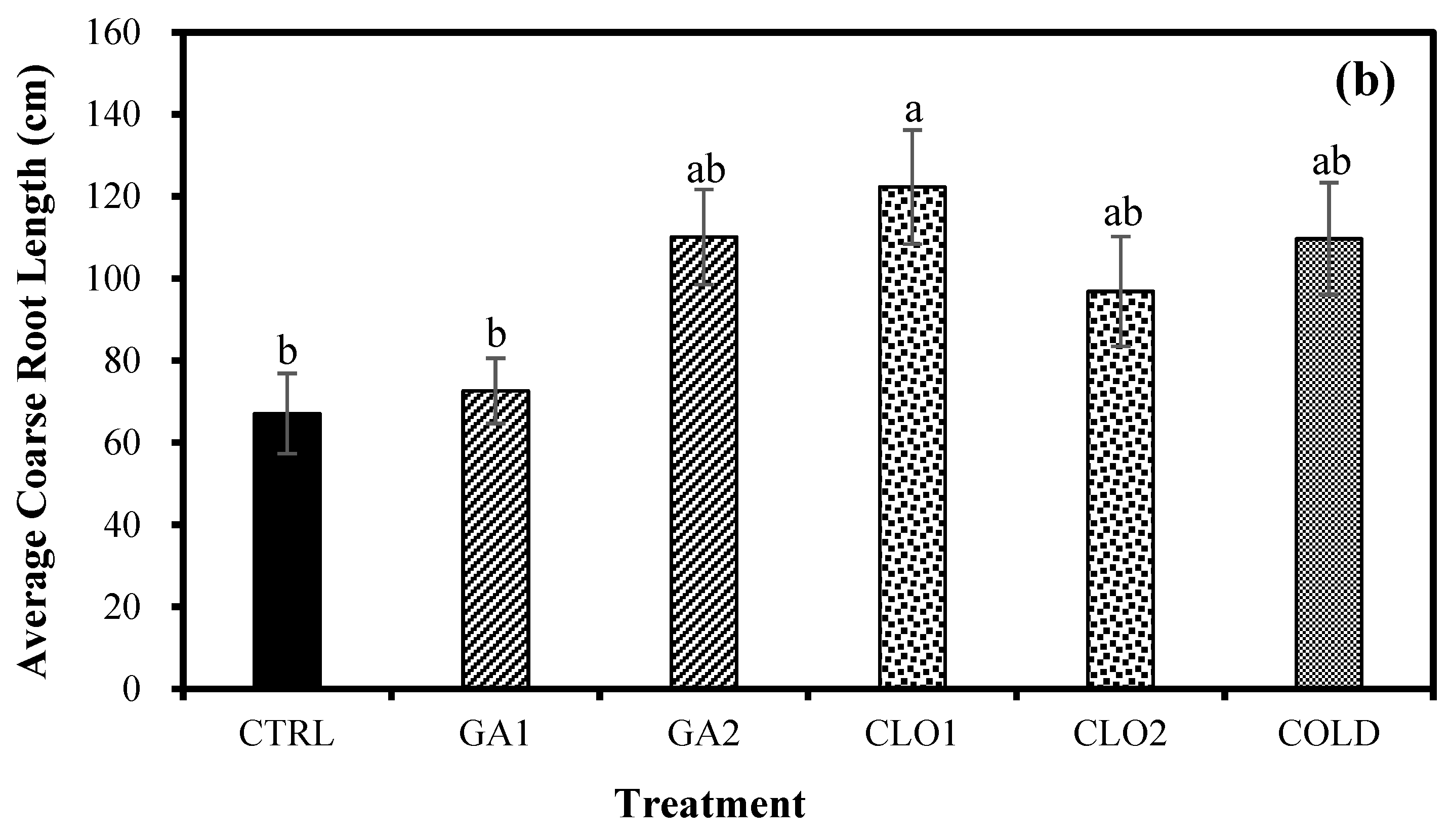
3.7. Root Growth Response after Seed Pre-Treatments (Experiment 3)
4. Discussion
4.1. Varieties Have Impacts on Germination and Early Growth of Industrial Hemp
4.2. Gibberellic Acid May Decrease Seed Germination but May Promote Early Growth of Industrial Hemp
4.3. Chlorine Dioxide Enhances Root Growth of Industrial Hemp Seedlings
4.4. Cold Pre-Treatment Does Not Influence Germination and Early Growth of Industrial Hemp
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TSW | thousand seed weight (g) |
| GP | % germination |
| NS | % normal seedling |
| AS | % abnormal seedling |
| SDW | shoot dry weight plant−1 (mg) |
| RDW | root dry weight plant−1 (mg) |
| SL | shoot length (cm) |
| SGR | shoot growth rate (mm·d−1) |
| SVI | seedling vigour index = % germination × seedling length (cm) |
| ARL | average root length (cm) |
| ARD | average root diameter (mm) |
| AFRL | average fine root length (cm) |
| ACRL | average coarse root length (cm) |
| CTRL | control (no seed pre-treatment) |
| GA1 | 500 mg·L−1 GA3 |
| GA2 | 1000 mg·L−1 GA3 |
| CLO1 | 500 mg·L−1 ClO2 |
| CLO2 | 1000 mg·L−1 ClO2 |
| COLD | cold temperature (4 °C for 72 h). |
References
- Vonapartis, E.; Aubin, M.-P.; Seguin, P.; Mustafa, A.F.; Charron, J.-B. Seed composition of ten industrial hemp cultivars approved for production in Canada. J. Food Compos. Anal. 2015, 39, 8–12. [Google Scholar] [CrossRef]
- Callaway, J. Hempseed as a nutritional resource: An overview. Euphytica 2004, 140, 65–72. [Google Scholar] [CrossRef]
- Cole, C.; Zurbo, B. Industrial hemp—A new crop for NSW. Primefacts 2008, 801, 1–6. [Google Scholar]
- Small, E. Cannabis: A Complete Guide, 1st ed.; Chapman and Hall/CRC: Milton, MA, USA, 2016. [Google Scholar]
- Crawford, F.; Deards, B.; Moir, B.; Thompson, N. Human Consumption of Hemp Seed: Prospects for Consumption and Production; ABARES Report to Client Prepared for FSANZ; ABARES: Canberra, Australia, 2012; pp. 1–15.
- Feng, H.; Nemzer, B.; DeVries, J.W. Sprouted Grains: Nutritional Value, Production and Applications; Woodhead Publishing: Duxford, UK, 2019. [Google Scholar]
- DPIRD. Guidelines to Assist Importation of Industrial Hemp Seed into Western Australia; Department of Primary Industries and Regional Development (DPIRD): South Perth, Australia, 2018; pp. 1–3.
- Gommers, C.M.M. Seedling establishment: A dimmer switch-regulated process between dark and light signaling. Plant. Physiol. 2018, 176, 1061–1074. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.H. Seed and seedling vigour in relation to crop growth and yield. Plant. Growth Regul. 1992, 11, 249–255. [Google Scholar] [CrossRef]
- Chacón, P.; Bustamante, R.; Henríquez, C. The effect of seed size on germination and seedling growth of Cryptocarya alba (Lauraceae) in Chile. Rev. Chil. Hist. Nat. 1998, 71, 189–197. [Google Scholar]
- Cicek, E.; Tilki, F. Seed size effects on germination, survival and seedling growth of Castanea sativa Mill. J. Biol. Sci. 2007, 7, 438–441. [Google Scholar] [CrossRef] [Green Version]
- Manonmani, V.; Vanangamudi, K. Effect of seed source and size on seed germination and seedling vigour of sandal (Santalum album). J. Trop. For. Sci. 2002, 14, 150–155. [Google Scholar]
- Aigbe, H.I.; Fredrick, C.; Omokhua, G. Effect of seed source on germination and early seedling growth of Heinsia crinita (Afzel.) G. Taylor. Appl. Trop. Agric. 2016, 21, 180–185. [Google Scholar]
- Gallagher, M.K.; Wagenius, S. Seed source impacts germination and early establishment of dominant grasses in prairie restorations. J. Appl. Ecol. 2016, 53, 251–263. [Google Scholar] [CrossRef]
- Romero, F.R.; Delate, K.; Hannapel, D.J. The effect of seed source, light during germination, and cold-moist stratification on seed germination in three species of Echinacea for organic production. HortScience 2005, 40, 1751–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, R.T.d.; Martins, C.C.; Silva, G.Z.d.; Martins, D. Origin and temperature on the germination of beggartick seeds. Rev. Bras. Eng. Agrícola Ambient. 2017, 21, 448–453. [Google Scholar] [CrossRef] [Green Version]
- Claudia, R.D.; Leandro, D.d.S.; Ademir, G.; Thamiris, G.D. Seed origin, storage conditions, and gibberellic acid on in vitro germination of Campomanesia adamantium (Cambess.) O. Berg. Afr. J. Biotechnol. 2016, 15, 1731–1737. [Google Scholar] [CrossRef] [Green Version]
- Balaguera-López, H.E.; Cárdenas-Hernández, J.F.; Álvarez-Herrera, J.G. Effect of gibberellic acid (GA3) on seed germination and growth of tomato (Solanum lycopersicum L.). Acta Hortic. 2009, 821, 141–148. [Google Scholar] [CrossRef]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Stejskalová, J.; Kupka, I.; Miltner, S. Effect of gibberellic acid on germination capacity and emergence rate of Sycamore Maple (Acer pseudoplatanus L.) seeds. J. For. Sci. 2015, 61, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Baki, A.A. Pitfalls in using sodium hypochlorite as a seed disinfectant in 14C incorporation studies. Plant. Physiol. 1974, 53, 768. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, A.; Chhetri, V.S.; Bhattacharya, D.; Cason, C.; Luu, P.; Suazo, A. Effectiveness of daily rinsing of alfalfa sprouts with aqueous chlorine dioxide and ozonated water on the growth of Listeria monocytogenes during sprouting. Lett. Appl. Microbiol. 2019, 69, 252–257. [Google Scholar] [CrossRef]
- Hung, N.B.; Park, W.R.; Yun, B.; Seo, D.C.; Kim, W.I.; Kim, H.J.; Han, S.; Kim, S.R. Effect of sequential presoaking and chlorine dioxide treatment on the inactivation of pathogenic Escherichia coli and Salmonella spp. on sprout seeds. Appl. Biol. Chem. 2020, 63, 53. [Google Scholar] [CrossRef]
- Lee, S.; Kim, H.; Beuchat, L.R.; Ryu, J.H. Development of a decontamination method to inactivate Acidovorax citrulli on Cucurbitaceae seeds without loss of seed viability. J. Sci. Food Agric. 2019, 99, 5734–5739. [Google Scholar] [CrossRef]
- Miyoshi, K.; Sato, T. The Effects of ethanol on the germination of seeds of japonica and indica rice (Oryza sativa L.) under anaerobic and aerobic conditions. Ann. Bot. 1997, 79, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Ruiming, W.; Bingcui, C.; Tengfei, W.; Piwu, L.; Feng, D. Effects of chlorine dioxide on the germination, oxidative metabolism and growth of barley seedlings (Hordeum vulgare L.). Sci. Rep. 2019, 9, 5765. [Google Scholar] [CrossRef]
- Salehi, M.R.; Ashiri, F.; Salehi, H. Effect of different ethanol concentrations on seed germination of three turfgrass genera. Adv. Nat. Appl. Sci. 2008, 2, 6–9. [Google Scholar]
- Sauer, D.B. Disinfection of seed surfaces with sodium hypochlorite. Phytopathology 1986, 76, 745. [Google Scholar] [CrossRef]
- Chen, S.Y.; Chou, S.H.; Tsai, C.C.; Hsu, W.Y.; Baskin, C.C.; Baskin, J.M.; Chien, C.T.; Huang, K.; Long, L. Effects of moist cold stratification on germination, plant growth regulators, metabolites and embryo ultrastructure in seeds of Acer morrisonense (Sapindaceae). Plant Physiol. Biochem. 2015, 94, 165–173. [Google Scholar] [CrossRef]
- Cho, J.S.; Jang, B.K.; Lee, C.H. Breaking combinational dormancy of Rhus javanica L. seeds in South Korea: Effect of mechanical scarification and cold-moist stratification. S. Afr. J. Bot. 2020, 133, 174–177. [Google Scholar] [CrossRef]
- Li, W.; Liu, X.; Hanada, A.; Khan, M.A. Effect of cold stratification, scarification and hormones on germination of dimorphic seeds of Atriplex centralasiatica under saline conditions. Seed Sci. Technol. 2011, 39, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Mattana, E.; Picciau, R.; Puddu, S.; Cuena Lombraña, A.; Bacchetta, G. Effect of temperature and cold stratification on seed germination of the Mediterranean wild aromatic Clinopodium sandalioticum (Lamiaceae). Plant Biosyst. 2016, 150, 846–850. [Google Scholar] [CrossRef]
- Sun, B.; Liu, G.-L.; Phan, T.T.; Yang, L.-T.; Li, Y.-R.; Xing, Y.-X. Effects of cold stress on root growth and physiological metabolisms in seedlings of different sugarcane varieties. Sugar Tech. 2017, 19, 165–175. [Google Scholar] [CrossRef]
- Hu, H.R.; Liu, H.; Du, G.H.; Yang, F.; Deng, G.; Yang, Y.; Liu, F.H. Fiber and seed type of hemp (Cannabis sativa L.) responded differently to salt-alkali stress in seedling growth and physiological indices. Ind. Crop. Prod. 2019, 129, 624–630. [Google Scholar] [CrossRef]
- Hu, H.R.; Liu, H.; Liu, F.H. Seed germination of hemp (Cannabis sativa L.) cultivars responds differently to the stress of salt type and concentration. Ind. Crop. Prod. 2018, 123, 254–261. [Google Scholar] [CrossRef]
- Mi, R.; Taylor, A.G.; Smart, L.B.; Mattson, N.S. Developing production guidelines for baby leaf hemp (Cannabis sativa L.) as an edible salad green: Cultivar, sowing density and seed size. Agriculture 2020, 10, 617. [Google Scholar] [CrossRef]
- Sera, B.; Gajdova, I.; Gavril, B.; Hnatiuc, E.; Sery, M.; Spatenka, P. Hemp (Cannabis sativa L.) seeds after plasma treatment. In Proceedings of the 13th International Conference on Optimization of Electrical and Electronic Equipment, Brasov, Romania, 24–26 May 2012; Volumes 1–5, pp. 1371–1374. [Google Scholar]
- Sera, B.; Sery, M.; Gavril, B.; Gajdova, I. Seed germination and early growth responses to seed pre-treatment by non-thermal plasma in hemp cultivars (Cannabis sativa L.). Plasma Chem. Plasma Process. 2017, 37, 207–221. [Google Scholar] [CrossRef]
- Suriyong, S.; Krittigamas, N.; Pinmanee, S.; Punyalue, A.; Vearasilp, S. Influence of storage conditions on change of hemp seed quality. In Proceedings of the 1st International Conference on Asian Highland Natural Resources Management, Chiang Mai, Thailand, 7–9 January 2015; Volume 5, pp. 170–176. [Google Scholar]
- Gupta, U.; Solanki, H. Boron: Impact on Seed Germination and Growth of Solanum melongena L.; Lambert Academic Publishing GmbH & Co. KG: Saarbrücken, Germany, 2012; p. 136. [Google Scholar]
- Corns, W.G. Effects of gibberellin treatments on germination of various species of weed seeds. Can. J. Plant. Sci. 1960, 40, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Groot, S.; Karssen, C. Gibberellins regulate seed germination in tomato by endosperm weakening: A study with gibberellin-deficient mutants. Planta 1987, 171, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Machado de Mello, A.; Streck, N.A.; Blankenship, E.E.; Paparozzi, E.T. Gibberellic acid promotes aeed germination in Penstemon digitalis cv. Husker Red. HortScience 2009, 44, 870–873. [Google Scholar] [CrossRef] [Green Version]
- Chunmei, Z.; Hao, X.; Siting, Y.; Shiyi, W.; Lingfei, L.; Shengchun, Z.; Xiaojing, W. Gibberellic Acid-stimulated arabidopsis6 serves as an integrator of gibberellin, abscisic acid, and glucose signaling during seed germination in Arabidopsis. Plant. Physiol. 2015, 169, 2288–2303. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; An, L.; Jiao, X.; Chen, X.; Zhou, G.; McLaughlin, N. Effects of gibberellic acid on water uptake and germination of sweet sorghum seeds under salinity stress. Chil. J. Agric. Res. 2019, 79, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Cornea-Cipcigan, M.; Pamfil, D.; Sisea, C.R.; Margaoan, R. Gibberellic acid can improve seed germination and ornamental quality of selected Cyclamen species grown under short and long days. Agronomy 2020, 10, 516. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Sex Type | Country | Seed Importer |
|---|---|---|---|
| Ferimon | Monoecious | France | WA Hemp Growers’ Co-op Ltd. (HempGro) |
| Fedora 17 | Monoecious | France | WA Hemp Growers’ Co-op Ltd. (HempGro) |
| Santhica | Monoecious | France | WA Hemp Growers’ Co-op Ltd. (HempGro) |
| Felina 32 | Monoecious | France | Premium Hemp Australia and DPIRD * |
| Morpeth | Monoecious | Denmark | Food, Fibre and Land International Group (FFLI) |
| Bama 4 | Dioecious | China | Premium Hemp Australia and DPIRD |
| Han NE | Dioecious | China | Premium Hemp Australia and DPIRD |
| Han FNH | Dioecious | China | Premium Hemp Australia and DPIRD |
| Yuma 1 | Dioecious | China | Premium Hemp Australia and DPIRD |
| Han FNQ | Dioecious | China | Premium Hemp Australia and DPIRD |
| Han NW | Dioecious | China | Premium Hemp Australia and DPIRD |
| Puma 3 | Dioecious | China | Premium Hemp Australia and DPIRD |
| Han COLD | Dioecious | China | Premium Hemp Australia and DPIRD |
| SI 1 | Dioecious | China | Premium Hemp Australia and DPIRD |
| Seed Origin | Variety | TSW | GP (Petri Dish) | GP (Glasshouse) |
|---|---|---|---|---|
| Europe | Ferimon | 18.5 ef | 18.0 e | 46.0 cd |
| Fedora 17 | 21.1 de | 52.0 abcd | 74.0 abc | |
| Santhica | 19.2 def | 28.0 de | 62.0 abcd | |
| Felina 32 | 15.1 f | 42.0 abcde | 48.0 cd | |
| Morpeth | 21.2 de | 38.0 bcde | 48.0 cd | |
| Mean | 19.0 | 36.0 | 56.0 | |
| China | Bama 4 | 34.6 b | 40.0 bcde | 60.0 bcd |
| Han NE | 28.8 c | 62.0 ab | 64.0 abcd | |
| Han FNH | 23.7 cde | 66.0 ab | 82.0 ab | |
| Yuma 1 | 35.3 b | 54.0 abcd | 70.0 abc | |
| Han FNQ | 24.0 cd | 70.0 a | 92.0 a | |
| Han NW | 26.8 c | 26.0 de | 38.0 de | |
| Puma 3 | 35.6 b | 32.0 cde | 66.0 abcd | |
| Han COLD | 35.5 b | 58.0 abc | 66.0 abcd | |
| SI 1 | 62.3 a | 17.0 e | 13.0 e | |
| Mean | 34.1 | 47.0 | 61.0 | |
| Minimum | 15.1 | 17.0 | 13.0 | |
| Maximum | 21.2 | 70.0 | 92.0 | |
| Mean | 28.7 | 43.0 | 59.0 | |
| p-value | <0.001 *** | <0.001 *** | <0.001 *** |
| Varieties | NS | AS | SDW | RDW | SL | SGR | SVI |
|---|---|---|---|---|---|---|---|
| Ferimon | 88.0 abcd | 12.0 abc | 69.0 | 6.4 b | 5.9 def | 3.0 def | 269.4 cde |
| Fedora 17 | 95.0 abc | 5.0 bc | 76.8 | 6.6 b | 81 abcd | 4.1 abcd | 598.6 ab |
| Santhica | 91.0 abc | 9.0 bc | 86.7 | 8.8 b | 8.6 abc | 4.1 abc | 537.4 bc |
| Felina 32 | 100.0 a | 0.0 c | 89.4 | 10.8 b | 7.8 abcd | 3.9 abcd | 390.8 bcde |
| Morpeth | 100.0 a | 0.0 c | 64.3 | 11.9 b | 5.1 ef | 2.6 ef | 252.2 cde |
| Bama 4 | 68.0 d | 32.0 a | 79.1 | 6.3 b | 6.1 cdef | 3.0 cdef | 372.0 bcde |
| Han NE | 97.0 abc | 3.0 bc | 97.0 | 11.6 b | 10.5 a | 5.2 a | 666.6 ab |
| Han FNH | 97.0 ab | 3.0 bc | 62.1 | 7.9 b | 7.2 bcdef | 3.6 bcdef | 593.0 ab |
| Yuma 1 | 94.0 abc | 6.0 bc | 87.0 | 9.6 b | 7.7 bcde | 3.9 bcde | 543.0 abc |
| Han FNQ | 100.0 a | 0.0 c | 71.7 | 7.7 b | 9.3 ab | 4.6 ab | 853.8 a |
| Han NW | 79.0 abcd | 21.0 abc | 59.3 | 7.8 b | 5.1 f | 2.5 f | 206.6 de |
| Puma 3 | 90.0 abc | 10.0 bc | 117.8 | 10.0 b | 8.8 ab | 4.4 ab | 602.2 ab |
| Han COLD | 77.0 bcd | 19.0 abc | 90.9 | 9.0 b | 7.3 bcdef | 3.7 bcdef | 505.4 bcd |
| SI 1 | 75.0 cd | 25.0 ab | 117.9 | 25.4 a | 6.1 cdef | 3.7 cdef | 83.0 e |
| Minimum | 68.0 | 0.0 | 59.3 | 6.3 | 5.1 | 2.5 | 83.0 |
| Maximum | 100.0 | 32.0 | 117.9 | 25.4 | 10.5 | 5.2 | 853.8 |
| Mean | 89.0 | 10.0 | 83.5 | 10.0 | 7.4 | 3.7 | 462.4 |
| p-value | 0.008 ** | 0.017 * | 0.121 ns | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| Varieties | ARL | ARD | AFRL | ACRL |
|---|---|---|---|---|
| Ferimon | 82.6 bc | 0.27 abc | 65.3 | 17.3 bc |
| Fedora 17 | 83.5 bc | 0.24 abc | 64.1 | 19.4 bc |
| Santhica | 129.3 abc | 0.22 bc | 99.1 | 30.2 bc |
| Felina 32 | 102.3 bc | 0.19 c | 87.0 | 15.3 bc |
| Morphet | 246.3 a | 0.26 abc | 166.9 | 79.4 a |
| Bama 4 | 43.2 c | 0.26 abc | 33.2 | 10.0 c |
| Han NE | 128.5 abc | 0.22 bc | 98.1 | 30.4 bc |
| Han FNH | 131.6 abc | 0.20 bc | 107.1 | 24.5 bc |
| Yuma 1 | 147.9 abc | 0.24 abc | 106.7 | 41.2 abc |
| Han FNQ | 121.0 abc | 0.26 abc | 87.5 | 33.5 bc |
| Han NW | 143.9 abc | 0.28 ab | 96.3 | 47.6 abc |
| Puma 3 | 200.1 ab | 0.25 abc | 142.7 | 57.4 ab |
| Han COLD | 153.2 abc | 0.28 ab | 110.9 | 42.3 abc |
| SI 1 | 255.5 a | 0.31 a | 176.6 | 78.9 a |
| Minimum | 43.2 | 0.19 | 33.19 | 10.0 |
| Maximum | 255.5 | 0.31 | 176.62 | 79.4 |
| Mean | 140.6 | 0.25 | 102.97 | 37.7 |
| p-value | 0.037 ** | <0.001 *** | 0.077 ns | 0.006 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.M.; Rengel, Z.; Storer, P.; Siddique, K.H.M.; Solaiman, Z.M. Industrial Hemp (Cannabis sativa L.) Varieties and Seed Pre-Treatments Affect Seed Germination and Early Growth of Seedlings. Agronomy 2022, 12, 6. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010006
Islam MM, Rengel Z, Storer P, Siddique KHM, Solaiman ZM. Industrial Hemp (Cannabis sativa L.) Varieties and Seed Pre-Treatments Affect Seed Germination and Early Growth of Seedlings. Agronomy. 2022; 12(1):6. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010006
Chicago/Turabian StyleIslam, Mohammad Moinul, Zed Rengel, Paul Storer, Kadambot H. M. Siddique, and Zakaria M. Solaiman. 2022. "Industrial Hemp (Cannabis sativa L.) Varieties and Seed Pre-Treatments Affect Seed Germination and Early Growth of Seedlings" Agronomy 12, no. 1: 6. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010006