
Combination of Marker-Assisted Backcross Selection of Yr59 and Phenotypic Selection to Improve Stripe Rust Resistance and Agronomic Performance in Four Elite Wheat Cultivars

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
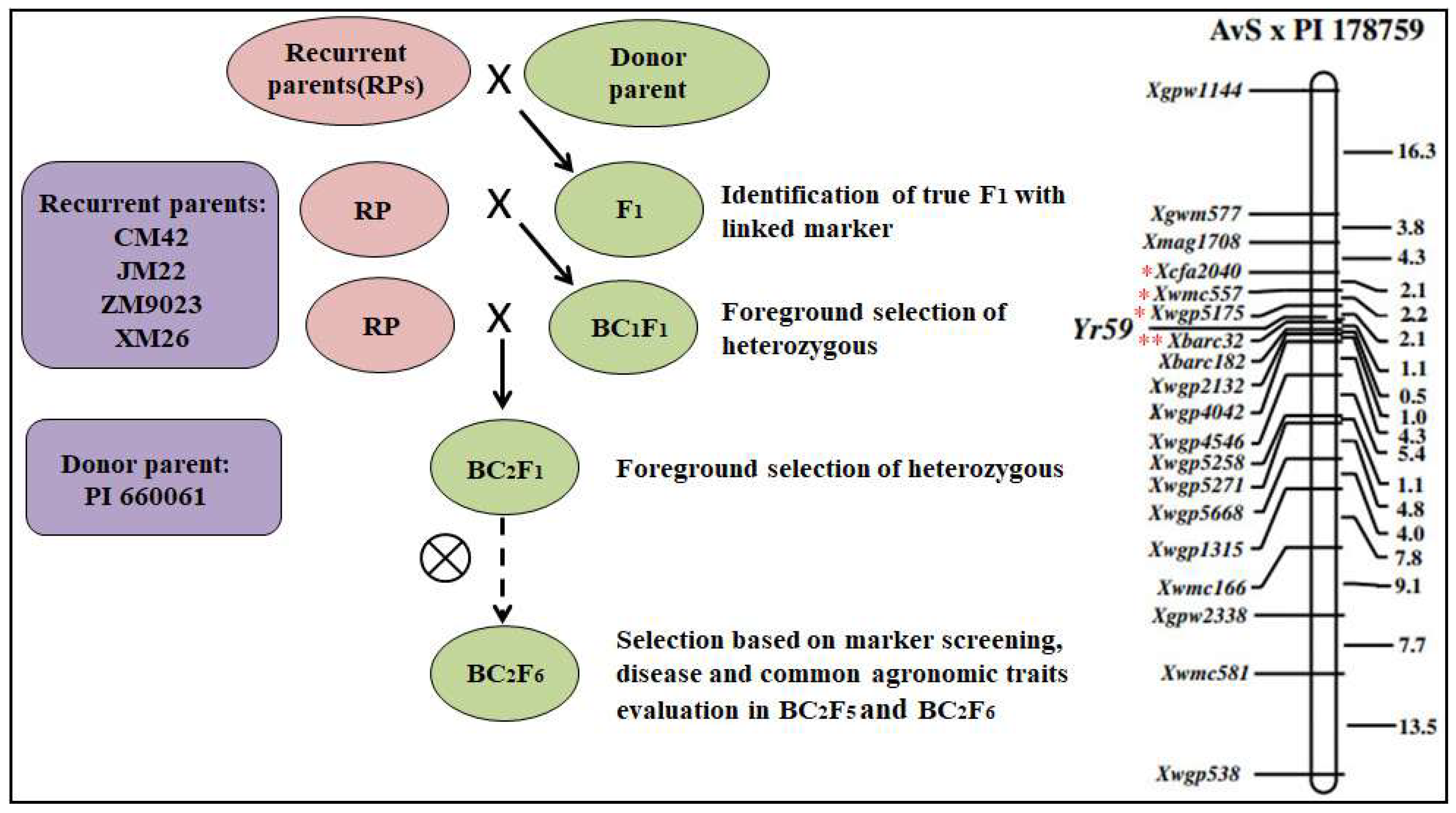
2.1. Plant Material
2.2. Phenotyping for Stripe Rust Reaction and Agronomic Traits in the Fields
2.3. Molecular Marker Genotyping
2.4. Statistical Analysis
3. Results
3.1. Marker-Assisted Backcrossing (MABC)
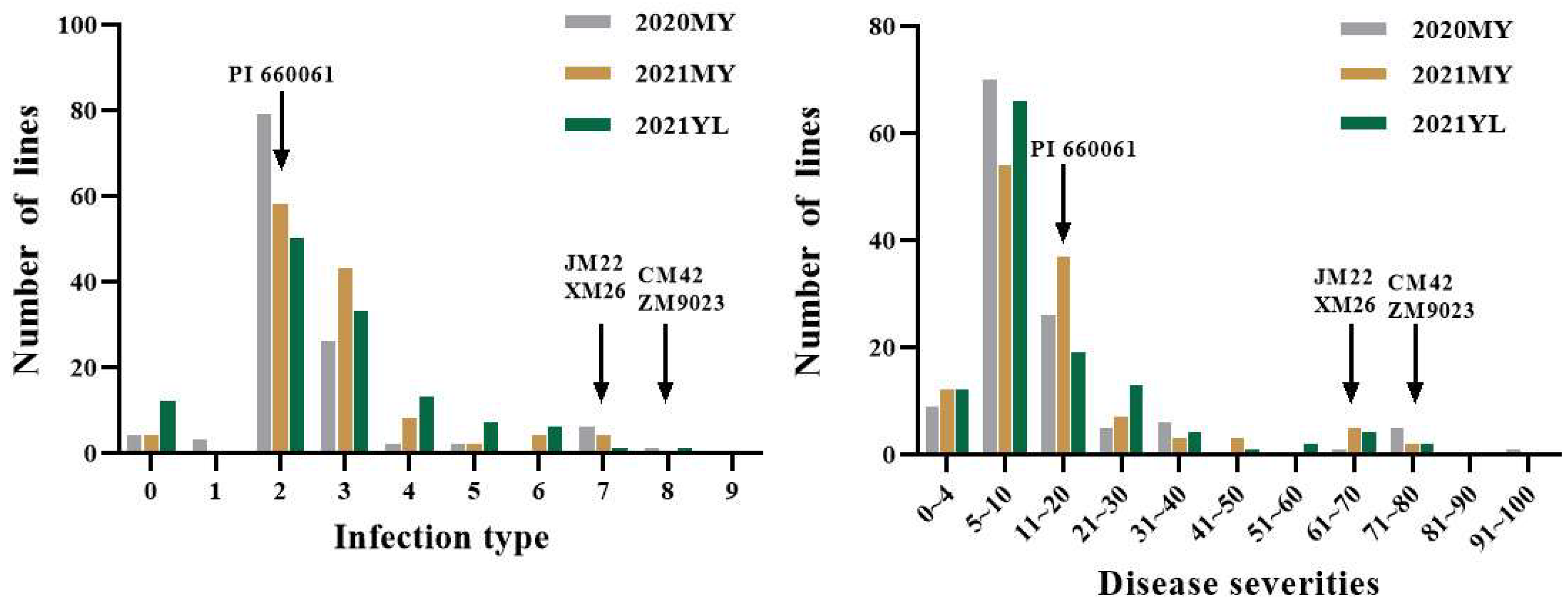
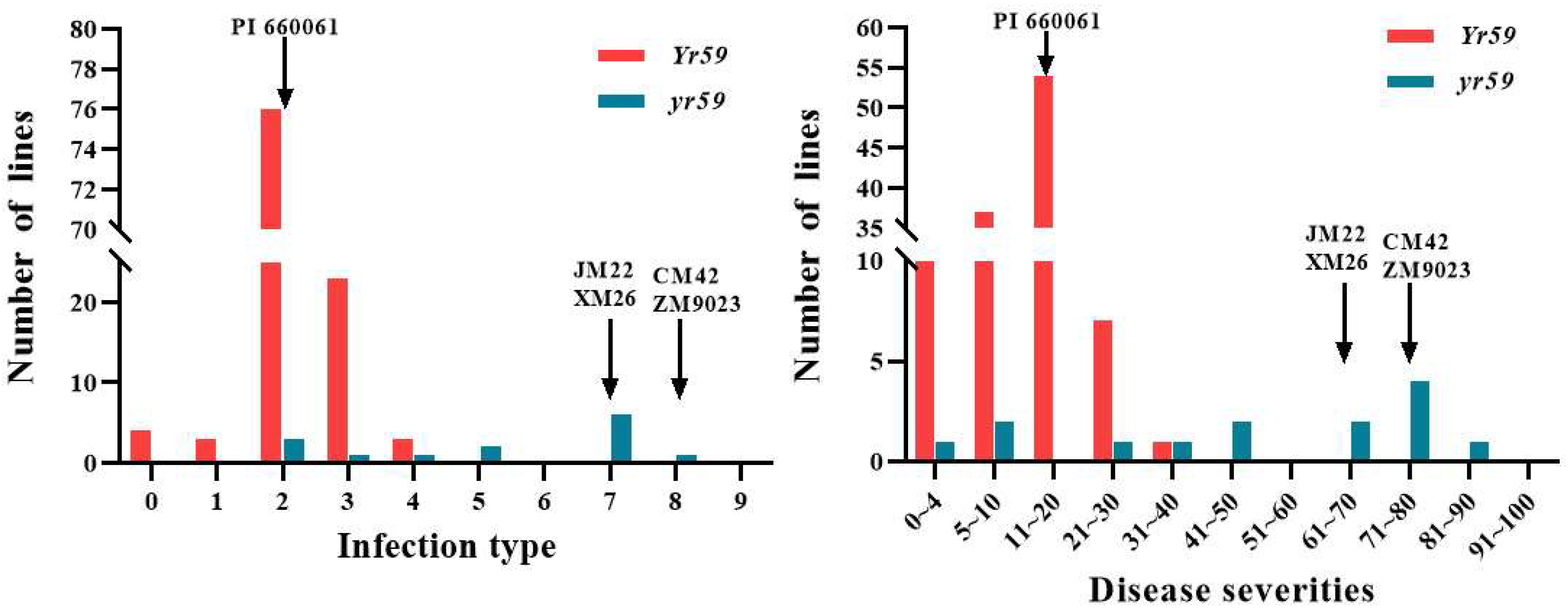
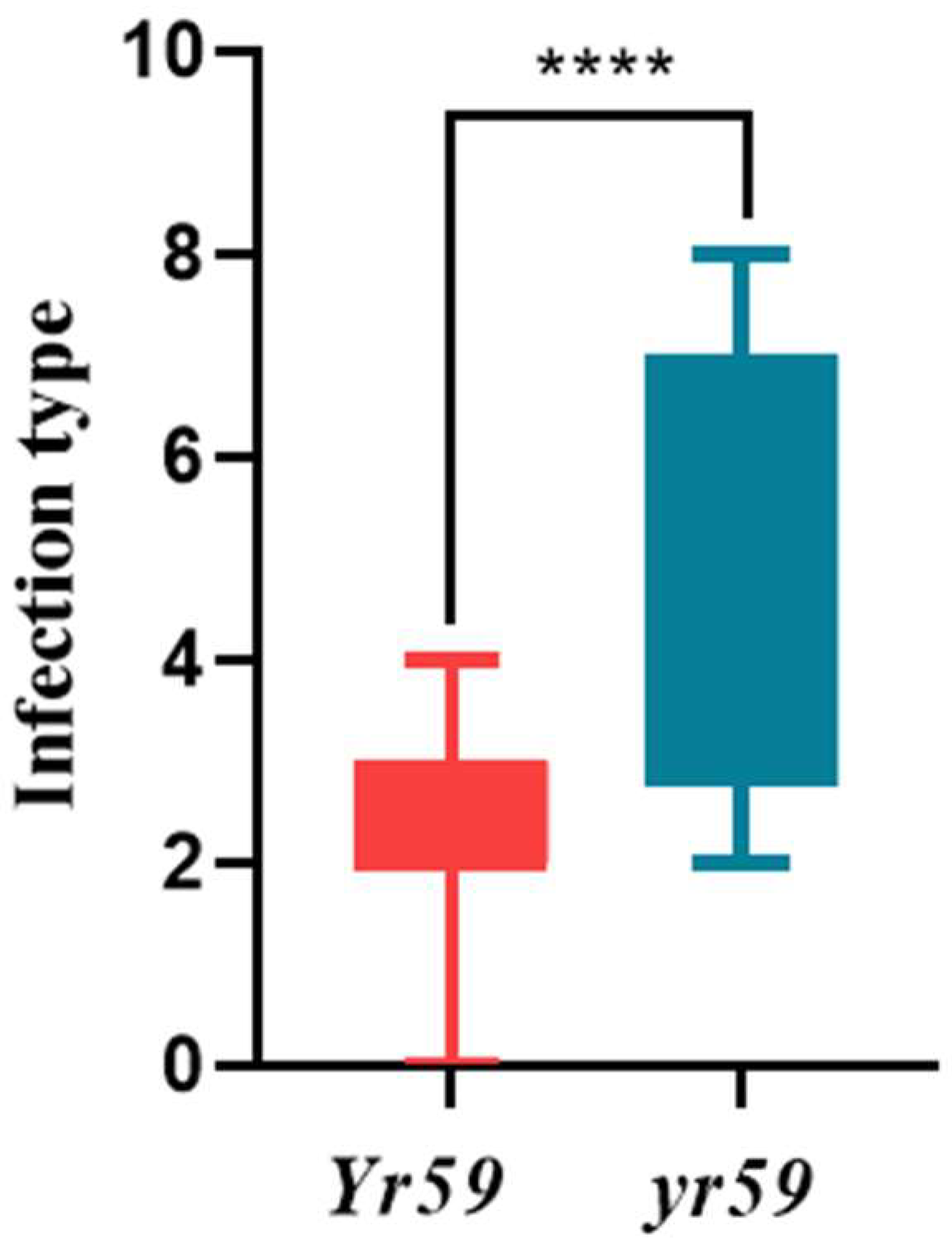
3.2. Evaluation of Disease Resistance
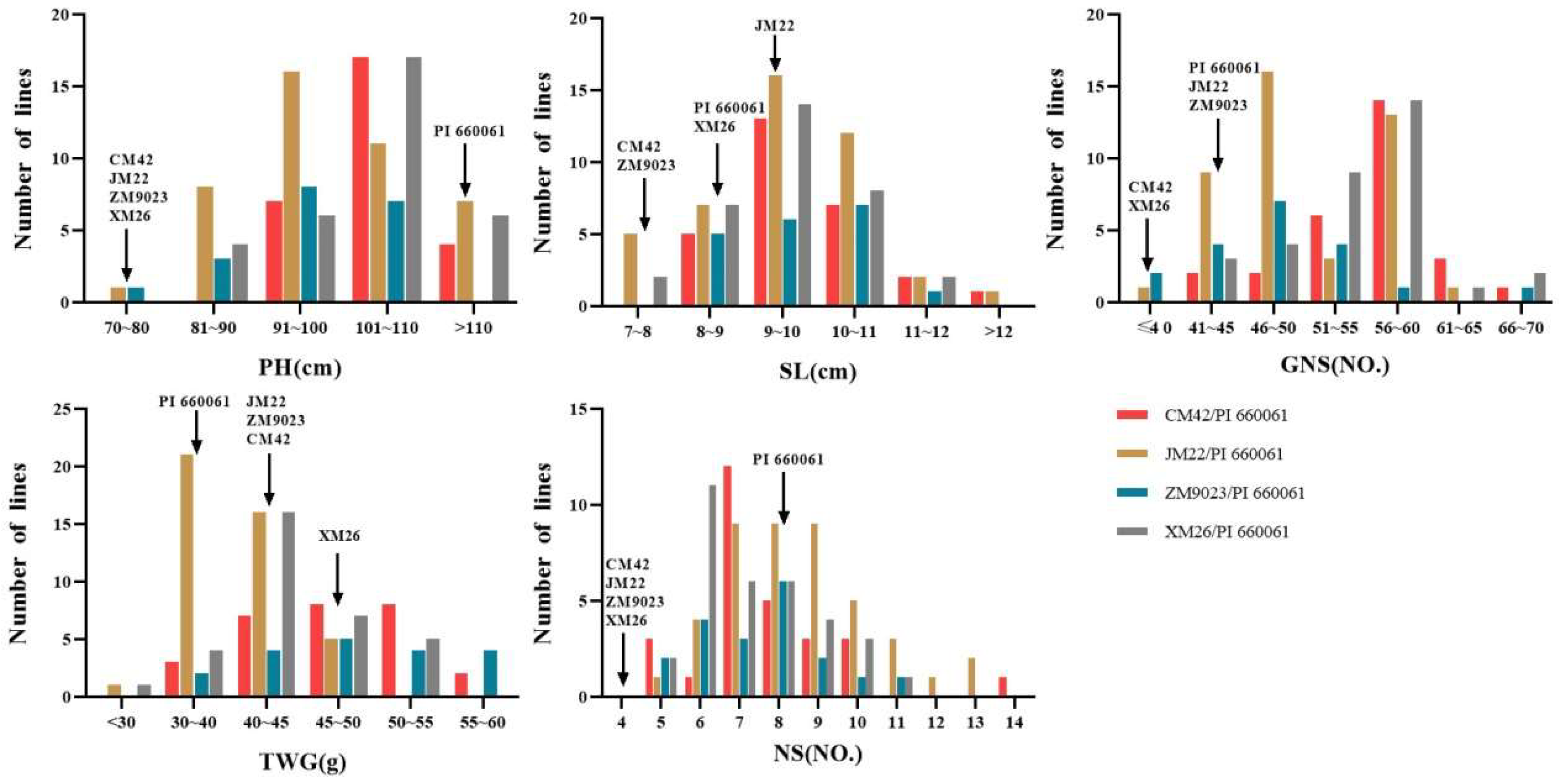
3.3. Evaluation of Agronomic Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, P.K.; Kumar, J.; Mir, R.R.; Kumar, A. Marker-Assisted Selection as a Component of Conventional Plant Breeding. Plant Breed. Rev. 2010, 33, 145–217. [Google Scholar] [CrossRef]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Chen, X. Pathogens which threaten food security: Puccinia striiformis, the wheat stripe rust pathogen. Food Secur. 2020, 12, 239–251. [Google Scholar] [CrossRef]
- Zhou, X.; Fang, T.; Li, K.; Huang, K.; Ma, C.; Zhang, M.; Li, X.; Yang, S.; Ren, R.; Zhang, P. Yield losses associated with different levels of stripe rust resistance of commercial wheat cultivars in China. Phytopathology 2021. [Google Scholar] [CrossRef]
- Collard, B.; Mackill, D.J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 2007, 363, 557–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panigrahi, J.; Mishra, R.R.; Sahu, A.R.; Rath, S.C.; Kole, C.R. Marker-Assisted Breeding for Stress Resistance in Crop Plants. In Molecular Stress Physiology of Plants; Springer: Berlin/Heidelberg, Germany, 2013; pp. 387–426. [Google Scholar]
- Toojinda, T.; Baird, E.; Booth, A.; Broers, L.; Hayes, P.; Powell, W.; Thomas, W.; Vivar, H.; Young, G. Introgression of quantitative trait loci (QTLs) determining stripe rust resistance in barley: An example of marker-assisted line development. Theor. Appl. Genet. 1998, 96, 123–131. [Google Scholar] [CrossRef]
- Dubcovsky, J. Marker-Assisted Selection in Public Breeding Programs. Crop Sci. 2004, 44, 1895–1898. [Google Scholar] [CrossRef]
- Merrick, L.F.; Burke, A.B.; Chen, X.M.; Carter, A.H. Breeding with major and minor genes: Ge-nomic selection for quantitative disease resistance. Front. Plant Sci. 2021, 12, 713667. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Pixley, K.; Warburton, M.L.; Xie, C.-X. Molecular marker-assisted breeding options for maize improvement in Asia. Mol. Breed. 2010, 26, 339–356. [Google Scholar] [CrossRef]
- Varshney, R.K.; Pandey, M.; Janila, P.; Nigam, S.N.; Sudini, H.; Gowda, M.V.C.; Sriswathi, M.; Radhakrishnan, T.; Manohar, S.S.; Nagesh, P. Marker-assisted introgression of a QTL region to improve rust resistance in three elite and popular varieties of peanut (Arachis hypogaea L.). Theor. Appl. Genet. 2014, 127, 1771–1781. [Google Scholar] [CrossRef] [Green Version]
- Leal-Bertioli, S.C.; Cavalcante, U.; Gouveia, E.G.; Taborda, C.B.; Shirasawa, K.; Guimarães, P.M.; Jackson, S.; Bertioli, D.J.; Moretzsohn, M.C. Identification of QTLs for Rust Resistance in the Peanut Wild Species Arachis magna and the Development of KASP Markers for Marker-Assisted Selection. G3 Genes|Genomes|Genet. 2015, 5, 1403–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujino, K.; Hirayama, Y.; Kaji, R. Marker-assisted selection in rice breeding programs in Hokkaido. Breed. Sci. 2019, 69, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Karunarathna, K.H.T.; Mewan, K.M.; Weerasena, O.V.D.S.J.; Perera, S.A.C.N.; Edirisinghe, E.N.U. A functional molecular marker for detecting blister blight disease resistance in tea (Camellia sinensis L.). Plant Cell Rep. 2021, 40, 351–359. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Zhang, J.; Zhang, X.; He, S.; Xie, D.; Liu, Y.; Li, C.; Wang, Z.; Liu, Y. Development of SSR markers in Paeonia based on De Novo transcriptomic assemblies. PLoS ONE 2020, 15, e0227794. [Google Scholar] [CrossRef] [Green Version]
- Vishwakarma, M.K.; Arun, B.; Mishra, V.K.; Yadav, P.S.; Kumar, H.; Joshi, A.K. Marker-assisted improvement of grain protein content and grain weight in Indian bread wheat. Euphytica 2016, 208, 313–321. [Google Scholar] [CrossRef]
- Yaniv, E.; Raats, D.; Ronin, Y.; Korol, A.B.; Grama, A.; Bariana, H.; Dubcovsky, J.; Schulman, A.H.; Fahima, T. Evaluation of marker-assisted selection for the stripe rust resistance gene Yr15, introgressed from wild emmer wheat. Mol. Breed. 2015, 35, 43. [Google Scholar] [CrossRef] [Green Version]
- Mallick, N.; Vinod, G.P.C.S.J.B.; Sharma, J.B.; Tomar, R.; Sivasamy, M.; Prabhu, K. Marker-assisted backcross breeding to combine multiple rust resistance in wheat. Plant Breed. 2015, 134, 172–177. [Google Scholar] [CrossRef]
- Randhawa, M.S.; Bains, N.S.; Sohu, V.S.; Chhuneja, P.; Trethowan, R.M.; Bariana, H.S.; Bansal, U. Marker Assisted Transfer of Stripe Rust and Stem Rust Resistance Genes into Four Wheat Cultivars. Agronomy 2019, 9, 497. [Google Scholar] [CrossRef] [Green Version]
- Ragoussis, J. Genotyping technologies for all. Drug Discov. Today Technol. 2006, 3, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Kumpatla, S.P.; Buyyarapu, R.; Mammadov, J.; Abdurakhmonov, I. Genomics-Assisted Plant Breeding in the 21st Century: Technological Advances and Progress. In Plant Breeding; InTech: London, UK, 2012; pp. 131–183. [Google Scholar]
- Rafalski, A. Applications of single nucleotide polymorphisms in crop genetics. Curr. Opin. Plant Biol. 2002, 5, 94–100. [Google Scholar] [CrossRef]
- Hyten, D.L.; Cannon, S.B.; Song, Q.; Weeks, N.; Fickus, E.W.; Shoemaker, R.C.; Specht, J.E.; Farmer, A.D.; May, G.D.; Cregan, P.B. High-throughput SNP discovery through deep resequencing of a reduced representation library to anchor and orient scaffolds in the soybean whole genome sequence. BMC Genom. 2010, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.J. High-Throughput SNP Genotyping to Accelerate Crop Improvement. Plant Breed. Biotechnol. 2014, 2, 195–212. [Google Scholar] [CrossRef]
- Mu, J.; Huang, S.; Liu, S.; Zeng, Q.; Dai, M.; Wang, Q.; Wu, J.; Yu, S.; Kang, Z.; Han, D. Genetic architecture of wheat stripe rust resistance revealed by combining QTL mapping using SNP-based genetic maps and bulked segregant analysis. Theor. Appl. Genet. 2018, 132, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Neelam, K.; Brown-Guedira, G.; Huang, L. Development and validation of a breeder-friendly KASPar marker for wheat leaf rust resistance locus Lr21. Mol. Breed. 2012, 31, 233–237. [Google Scholar] [CrossRef]
- Qie, Y.; Liu, Y.; Wang, M.; Li, X.; See, D.R.; An, D.; Chen, X. Development, Validation, and Re-selection of Wheat Lines with Pyramided Genes Yr64 and Yr15 Linked on the Short Arm of Chromosome 1B for Resistance to Stripe Rust. Plant Dis. 2019, 103, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Kaur, B.; Mavi, G.S.; Gill, M.S.; Saini, D.K. Utilization of KASP technology for wheat improvement. Cereal Res. Commun. 2020, 48, 409–421. [Google Scholar] [CrossRef]
- Chen, X. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Rosewarne, G.M.; Herrera-Foessel, S.A.; Singh, R.P.; Huerta-Espino, J.; Lan, C.X.; He, Z.H. Quantitative trait loci of stripe rust resistance in wheat. Theor. Appl. Genet. 2013, 126, 2427–2449. [Google Scholar] [CrossRef] [Green Version]
- Chen, X. Review Article: High-Temperature Adult-Plant Resistance, Key for Sustainable Control of Stripe Rust. Am. J. Plant Sci. 2013, 04, 608–627. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Penman, L.; Wan, A.; Cheng, P. Virulence races of Puccinia striiformis f. sp. triticiin 2006 and 2007 and development of wheat stripe rust and distributions, dynamics, and evolutionary relationships of races from 2000 to 2007 in the United States. Can. J. Plant Pathol. 2010, 32, 315–333. [Google Scholar] [CrossRef]
- Wan, A.; Chen, X. Virulence Characterization of Puccinia striiformis f. sp. tritici Using a New Set of Yr Single-Gene Line Differentials in the United States in 2010. Plant Dis. 2014, 98, 1534–1542. [Google Scholar] [CrossRef] [Green Version]
- Cheng, P.; Chen, X.M. Molecular mapping of a gene for stripe rust resistance in spring wheat cultivar IDO377s. Theor. Appl. Genet. 2010, 121, 195–204. [Google Scholar] [CrossRef]
- Fu, D.; Uauy, C.; Distelfeld, A.; Blechl, A.; Epstein, L.; Chen, X.; Sela, H.; Fahima, T.; Dubcovsky, J. A Kinase-START Gene Confers Temperature-Dependent Resistance to Wheat Stripe Rust. Sci. 2009, 323, 1357–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.L.; Wang, M.N.; Chen, X.M.; Lu, Y.; Kang, Z.S.; Jing, J.X. Identification of Yr59 conferring high-temperature adult-plant resistance to stripe rust in wheat germplasm PI 178759. Theor. Appl. Genet. 2014, 127, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, X. Stripe Rust Resistance. In Stripe Rust; Springer: Berlin/Heidelberg, Germany, 2017; pp. 353–558. [Google Scholar]
- Li, J.; Dundas, I.; Dong, C.; Li, G.; Trethowan, R.; Yang, Z.; Hoxha, S.; Zhang, P. Identification and characterization of a new stripe rust resistance gene Yr83 on rye chromosome 6R in wheat. Theor. Appl. Genet. 2020, 133, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Langridge, P.; Mir, R.R. Marker-assisted wheat breeding: Present status and future possibilities. Mol. Breed. 2010, 26, 145–161. [Google Scholar] [CrossRef]
- Kaur, S.; Kaur, J.; Mavi, G.S.; Dhillon, G.S.; Sharma, A.; Singh, R.; Devi, U.; Chhuneja, P. Pyramiding of High Grain Weight With Stripe Rust and Leaf Rust Resistance in Elite Indian Wheat Cultivar Using a Combination of Marker Assisted and Phenotypic Selection. Front. Genet. 2020, 11, 593426. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Li, S.; Liu, Z.; Zhou, Q.; Feng, Y.; Chai, S. Molecular marker assisted gene stacking for disease resistance and quality genes in the dwarf mutant of an elite common wheat cultivar Xiaoyan22. BMC Genet. 2020, 21, 45. [Google Scholar] [CrossRef]
- Yang, Q.; Fang, T.; Li, X.; Ma, C.; Yang, S.; Kang, Z.; Zhou, X. Improving stripe rust resistance and agronomic performance in three elite wheat cultivars using a combination of phenotypic selection and marker detection of Yr48. Crop Prot. 2021, 148, 105752. [Google Scholar] [CrossRef]
- Hu, T.; Zhong, X.; Yang, Q.; Zhou, X.; Li, X.; Yang, S.; Hou, L.; Yao, Q.; Guo, Q.; Kang, Z. Introgression of Two Quantitative Trait Loci for Stripe Rust Resistance into Three Chinese Wheat Cultivars. Agronomy 2020, 10, 483. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Hu, L.; Wang, G.; Zhang, L.; Dong, H.; Li, Y.; Zhang, J.; Zan, X.; Qi, X.; Li, C. Breeding strategies of high quality and wide regional adaptability wheat variety Zhengmai 9023 and relevant thoughts on wheat high yield breeding. J. Henan Agric. Sci. 2009, 9, 14–18. [Google Scholar]
- Tang, Y.-L.; Li, J.; Wu, Y.-Q.; Wei, H.-T.; Li, C.-S.; Yang, W.-Y.; Chen, F. Identification of QTLs for Yield-Related Traits in the ecombinant Inbred Line Population Derived from the Cross Between a Synthetic Hexaploid Wheat-Derived Variety Chuanmai 42 and a Chinese Elite Variety Chuannong 16. Agric. Sci. China 2011, 10, 1665–1680. [Google Scholar] [CrossRef]
- Li, H.; Song, J.; Liu, A.; Guo, D.; Wang, X.; Lin, D.; Zhao, Z.; Liu, J. Effect of sowing time and planting density on yield and components of ’Jimai22’ with super-high yield. Chin. Agric. Sci. Bull. 2011, 27, 243–248. [Google Scholar]
- Liu, X.; Li, M.; Wang, S.; Guo, C. Quality and utilization of new wheat variety Xinmai 26. J. Anhui Agric. Sci. 2018, 46, 31–33. [Google Scholar] [CrossRef]
- Li, G.Q.; Li, Z.F.; Yang, W.Y.; Zhang, Y.; He, Z.H.; Xu, S.C.; Singh, R.P.; Qu, Y.Y.; Xia, X.C. Molecular mapping of stripe rust resistance gene YrCH42 in Chinese wheat cultivar Chuanmai 42 and its allelism with Yr24 and Yr26. Theor. Appl. Genet. 2006, 112, 1434–1440. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, Y.; Han, D.; Kang, Z.; Li, G.; Cao, A.; Chen, P. SSR and STS markers for wheat stripe rust resistance gene Yr26. Euphytica 2007, 159, 359–366. [Google Scholar] [CrossRef]
- Chen, C.; He, Z.; Lu, J.; Li, J.; Ren, Y.; Ma, C.; Xia, X. Molecular mapping of stripe rust resistance gene YrJ22 in Chinese wheat cultivar Jimai 22. Mol. Breed. 2016, 36, 118. [Google Scholar] [CrossRef]
- Yin, G.-H.; Li, G.-Y.; He, Z.-H.; Liu, J.-J.; Wang, H.; Xia, X.-C. Molecular Mapping of Powdery Mildew Resistance Gene in Wheat Cultivar Jimai 22. Acta Agron. Sin. 2009, 35, 1425–1431. [Google Scholar] [CrossRef]
- Zhang, P.; Guo, C.; Liu, Z.; Bernardo, A.; Ma, H.; Jiang, P.; Song, G.; Bai, G. Quantitative trait loci for Fusarium head blight resistance in wheat cultivars Yangmai 158 and Zhengmai 9023. Crop J. 2021, 9, 143–153. [Google Scholar] [CrossRef]
- Gao, L.; Yan, X.; Li, X.; Guo, G.; Hu, Y.; Ma, W.; Yan, Y. Proteome analysis of wheat leaf under salt stress by two-dimensional difference gel electrophoresis (2D-DIGE). Phytochemistry 2011, 72, 1180–1191. [Google Scholar] [CrossRef]
- Liu, T.G.; Peng, Y.L.; Chen, W.Q.; Zhang, Z.Y. First Detection of Virulence in Puccinia striiformis f. sp. tritici in China to Resistance Genes Yr24 (=Yr26) Present in Wheat Cultivar Chuanmai 42. Plant Dis. 2010, 94, 1163. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, T.; Zhang, Z.; Jia, Q.; Wang, B.; Gao, L.; Peng, Y.; Jin, S.; Chen, W. Discovery and pathogenicity of CYR34, a new race of Puccinia striiformis f. sp. tritici in China. Acta Phytopathol. Sin. 2017, 47, 681–687. [Google Scholar] [CrossRef]
- Wang, M.; Chen, X.; Xu, L.; Cheng, P.; Bockelman, H.E. Registration of 70 Common Spring Wheat Germplasm Lines Resistant to Stripe Rust. J. Plant Regist. 2012, 6, 104–110. [Google Scholar] [CrossRef]
- Chen, W.; Wellings, C.; Chen, X.; Kang, Z.; Liu, T. Wheat stripe (yellow) rust caused by Puccinia striiformisf. sp. tritici. Mol. Plant Pathol. 2014, 15, 433–446. [Google Scholar] [CrossRef]
- Line, R.F.; Qayoum, A. Virulence, Aggressiveness, Evolution, and Distribution of Races of Puccinia Striiformis (the Cause Of Stripe Rust of Wheat) in North America 1968–1987; US Department of Agriculture: Washington, DC, USA, 1992.
- Peterson, R.F.; Campbell, A.B.; Hannah, A.E. A diagrammatic scale for estimating rust intensity on leaves and stems of cereals. Can. J. Res. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Somers, D.J.; Isaac, P.; Edwards, K. A high-density microsatellite consensus map for bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2004, 109, 1105–1114. [Google Scholar] [CrossRef]
- Rajapakse, S. GENETICS | Molecular Markers. Encycl. Rose Sci. 2003, 149, 334–341. [Google Scholar] [CrossRef]
- Cuc, L.M.; Mace, E.S.; Crouch, J.H.; Quang, V.D.; Long, T.D.; Varshney, R.K. Isolation and characterization of novel microsatellite markers and their application for diversity assessment in cultivated groundnut (Arachis hypogaea). BMC Plant Biol. 2008, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Bassam, B.J.; Caetano-Anollés, G.; Gresshoff, P.M. Fast and sensitive silver staining of DNA in polyacrylamide gels. Anal. Biochem. 1991, 196, 80–83. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, Z.; Liu, B.; Gao, L.; Peng, Y.; Chen, W. Detection of virulence to Yr26 and patho-genicity to Chinese commercial winter wheat cultivars at seedling stage. Acta Phytopathol. Sin. 2015, 45, 41–47. [Google Scholar] [CrossRef]
- Liu, R.; Lu, J.; Zhou, M.; Zheng, S.; Liu, Z.; Zhang, C.; Du, M.; Wang, M.; Li, Y.; Wu, Y.; et al. Developing stripe rust resistant wheat (Triticum aestivum L.) lines with gene pyramiding strategy and marker-assisted selection. Genet. Resour. Crop Evol. 2020, 67, 381–391. [Google Scholar] [CrossRef]
- Gautam, T.; Dhillon, G.; Saripalli, G.; Rakhi; Singh, V.P.; Prasad, P.; Kaur, S.; Chhuneja, P.; Sharma, P.K.; Balyan, H.S.; et al. Marker-assisted pyramiding of genes/QTL for grain quality and rust resistance in wheat (Triticum aestivum L.). Mol. Breed. 2020, 40, 49. [Google Scholar] [CrossRef]
- Ren, M.; Xu, R. Molecular marker-assisted selection and identification of Guinong 19, a new wheat variety. Guizhou Agric. Sci. 2009, 37, 10–13. [Google Scholar]
- Zhang, B.; Jia, Q.; Huang, J.; Cao, S.; Sun, Z.; Luo, H.; Wang, X.; Jin, S. Trends and toxicity analysis of new strains G22-9 and G22-14 in puccinia striiformis f. sp. Tritici. Acta Agric. Boreali Occident. Sin. 2015, 24, 125–130. [Google Scholar]
- Li, W.; Song, G.; Li, J.; Li, Y.; Zhang, S.; Zhang, Z. Molecular detection of four pleiotropic disease resistance genes in wheat. J. Triticeae Crops. 2020, 40, 395–400. [Google Scholar]
- Chen, F.; Mu, J.; Huang, C.; Liu, J.; Yao, J.; Meng, J. Comparison between pedigree and bulk method in wheat breeding. J. Jinling Inst. Technol. 2003. [Google Scholar] [CrossRef]
- Liu, L.; Wang, M.N.; Feng, J.Y.; See, D.R.; Chao, S.M.; Chen, X.M. Combination of all-stage and high-temperature adult-plant resistance QTL confers high-level, durable resistance to stripe rust in winter wheat cultivar Madsen. Theor. Appl. Genet. 2018, 131, 1835–1849. [Google Scholar] [CrossRef]
- Liu, L.; Yuan, C.Y.; Wang, M.N.; See, D.R.; Zemetra, R.S.; Chen, X.M. QTL analysis of durable stripe rust resistance in the North American winter wheat cultivar Skiles. Theor. Appl. Genet. 2019, 132, 1677–1691. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Primer Sequence | Tm (°C) | Linked to Yr Gene | References |
|---|---|---|---|---|
| Xbarc32 | GCGTGAATCCGGAAACCCAATCTGTG | 60.5 | Yr59 | [36,60] |
| TGGAGAACCTTCGCATTGTGTCATTA | ||||
| Xwgp5175 | GGAGGCTTAGGGAAG | 49.0 | Yr59 | [36,60] |
| TGGTAGGTCCTTGTA | ||||
| Xwmc557 | GGTGCTTGTTCATACGGGCT | 57.6 | Yr59 | [36,61] |
| AGGTCCTCGATCCGCTCAT | ||||
| Xcfa2040 | TCAAATGATTTCAGGTAACCACTA | 52.2 | Yr59 | [36,60] |
| TTCCTGATCCCACCAAACAT | ||||
| Xwmc658 | CTCATCGTCCTCCTCCACTTTG | 57.6 | YrJ22 | [50,60] |
| GCCATCCGTTGACTTGAGGTTA | ||||
| WE173 | GGGACAAGGGGAGTTGAAGC | 58.0 | Yr26 | [49,60] |
| GAGAGTTCCAAGCAGAACAC | ||||
| Xbarc181 | CGCTGGAGGGGGTAAGTCATCAC | 58.0 | Yr26 | [49,60] |
| CGCAAATCAAGAACACGGGAGAAAGAA |
| Number of Plants/Lines | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Generations | CM42/PI660061 | JM22/PI660061 | ZM9023/PI660061 | XM26/PI660061 | Total | |||||
| Tested | Positive | Tested | Positive | Tested | Positive | Tested | Positive | Tested | Positive | |
| F1 | 10 | 8 | 13 | 9 | 9 | 7 | 11 | 9 | 43 | 33 |
| BC1F1 | 13 | 6 | 15 | 7 | 10 | 4 | 12 | 6 | 50 | 23 |
| BC2F1 | 24 | 12 | 31 | 16 | 20 | 10 | 23 | 11 | 98 | 49 |
| BC2F5 | 28 | 28 | 43 | 39 | 19 | 16 | 33 | 26 | 123 | 109 |
| BC2F6 | 28 | 28 | 43 | 39 | 19 | 16 | 33 | 26 | 123 | 109 |
| Cross | Resistant (IT = 0–3) | Intermediate (IT = 4–6) | Susceptible (IT = 7–9) |
|---|---|---|---|
| CM42/PI660061 | 26 | 2 | 0 |
| JM22/PI660061 | 39 | 2 | 2 |
| ZM9023/PI660061 | 17 | 1 | 1 |
| XM26/PI660061 | 28 | 4 | 1 |
| Stripe Rust | Agronomic Trait | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Line | Parent/Crosses | Gene | IT | DS (%) | PH (cm) | NS | GNS | SL (cm) | TGW (g) |
| M169 | Susc. check | No | 9 | 100 | 116.0 | 5.0 | 39.0 | 10.3 | 38.1 |
| PI 660061 | Res. donor | Yr59 | 2 | 20 | 125.1 | 8.0 | 45.0 | 8.7 | 39.5 |
| CM42 | Elite parent | Yr26 | 6 | 75 | 91.3 | 5.0 | 40.0 | 7.1 | 42.8 |
| JM22 | Elite parent | YrJ22 | 5 | 60 | 82.6 | 4.0 | 44.0 | 9.3 | 43.5 |
| ZM9023 | Elite parent | Unknown | 5 | 70 | 86.4 | 4.0 | 41.0 | 7.1 | 42.4 |
| XM26 | Elite parent | Unknown | 5 | 60 | 83.2 | 5.0 | 39.0 | 8.1 | 48.2 |
| BC2F6-7 | JM22/PI660061 | Yr59+YrJ22 | 1 | 5 | 92.1 | 13.0 | 49.0 | 9.6 | 42.2 |
| BC2F6-8 | JM22/PI660061 | Yr59+YrJ22 | 2 | 8 | 89.3 | 7.0 | 49.0 | 9.8 | 41.8 |
| BC2F6-21 * | JM22/PI660061 | Yr59 | 1 | 7 | 93.5 | 9.0 | 55.0 | 9.0 | 46.6 |
| BC2F6-27 | JM22/PI660061 | Yr59 | 3 | 20 | 95.3 | 10.0 | 47.0 | 11.1 | 42.3 |
| BC2F6-28 * | JM22/PI660061 | Yr59 | 2 | 13 | 97.2 | 7.0 | 58.0 | 9.4 | 43.1 |
| BC2F6-33 * | JM22/PI660061 | Yr59+YrJ22 | 3 | 5 | 88.1 | 9.0 | 59.0 | 10.0 | 44.7 |
| BC2F6-40 * | JM22/PI660061 | Yr59+YrJ22 | 3 | 15 | 94.8 | 7.0 | 56.0 | 10.2 | 42.0 |
| BC2F6-48 * | CM42/PI660061 | Yr59+Yr26 | 2 | 15 | 95.6 | 8.0 | 56.0 | 10.5 | 43.6 |
| BC2F6-59 * | CM42/PI660061 | Yr59 | 2 | 8 | 97.8 | 7.0 | 56.0 | 10.1 | 47.5 |
| BC2F6-61 * | CM42/PI660061 | Yr59+Yr26 | 3 | 18 | 99.7 | 7.0 | 61.0 | 10.0 | 46.0 |
| BC2F6-65 * | CM42/PI660061 | Yr59 | 2 | 13 | 99.3 | 5.0 | 56.0 | 9.0 | 52.7 |
| BC2F6-69 | CM42/PI660061 | Yr59+Yr26 | 3 | 17 | 90.6 | 7.0 | 51.0 | 9.4 | 43.8 |
| BC2F6-72 | ZM9023/PI660061 | Yr59 | 2 | 8 | 85.1 | 6.0 | 43.0 | 8.9 | 44.2 |
| BC2F6-73 * | ZM9023/PI660061 | Yr59 | 0 | 0 | 88.7 | 6.0 | 46.0 | 8.3 | 57.6 |
| BC2F6-78 | ZM9023/PI660061 | Yr59 | 2 | 13 | 97.0 | 8.0 | 46.0 | 8.4 | 40.3 |
| BC2F6-84 * | ZM9023/PI660061 | Yr59 | 2 | 4 | 97.7 | 8.0 | 45.0 | 10.2 | 58.1 |
| BC2F6-85 * | ZM9023/PI660061 | Yr59 | 2 | 17 | 86.1 | 7.0 | 45.0 | 9.3 | 50.0 |
| BC2F6-86 * | ZM9023/PI660061 | Yr59 | 1 | 5 | 96.2 | 7.0 | 42.0 | 10.3 | 57.8 |
| BC2F6-96 * | XM26/PI660061 | Yr59 | 3 | 12 | 82.8 | 7.0 | 51.0 | 9.2 | 49.4 |
| BC2F6-97 * | XM26/PI660061 | Yr59 | 2 | 5 | 99.4 | 7.0 | 43.0 | 10.2 | 53.7 |
| BC2F6-98 | XM26/PI660061 | Yr59 | 2 | 4 | 87.1 | 5.0 | 43.0 | 8.7 | 54.8 |
| BC2F6-102 | XM26/PI660061 | Yr59 | 3 | 15 | 96.8 | 6.0 | 56.0 | 8.5 | 44.1 |
| BC2F6-106 * | XM26/PI660061 | Yr59 | 3 | 20 | 96.5 | 6.0 | 61.0 | 10.5 | 44.1 |
| BC2F6-107 * | XM26/PI660061 | Yr59 | 3 | 13 | 88.6 | 7.0 | 60.0 | 9.3 | 43.0 |
| BC2F6-116 | XM26/PI660061 | Yr59 | 3 | 12 | 94.8 | 6.0 | 58.0 | 9.2 | 42.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Fang, T.; Zhou, X.; Chen, X.; Li, X.; Feng, J.; Yang, S.; Kang, Z. Combination of Marker-Assisted Backcross Selection of Yr59 and Phenotypic Selection to Improve Stripe Rust Resistance and Agronomic Performance in Four Elite Wheat Cultivars. Agronomy 2022, 12, 497. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020497
Zhang M, Fang T, Zhou X, Chen X, Li X, Feng J, Yang S, Kang Z. Combination of Marker-Assisted Backcross Selection of Yr59 and Phenotypic Selection to Improve Stripe Rust Resistance and Agronomic Performance in Four Elite Wheat Cultivars. Agronomy. 2022; 12(2):497. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020497
Chicago/Turabian StyleZhang, Min, Taohong Fang, Xinli Zhou, Xianming Chen, Xin Li, Junyan Feng, Suizhuang Yang, and Zhensheng Kang. 2022. "Combination of Marker-Assisted Backcross Selection of Yr59 and Phenotypic Selection to Improve Stripe Rust Resistance and Agronomic Performance in Four Elite Wheat Cultivars" Agronomy 12, no. 2: 497. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020497