
Evaluation of Germplasm and Development of Markers for Resistance to Plasmodiophora brassicae in Radish (Raphanussativus L.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and P. brassicae Inoculation
2.2. DNA Extraction, Gene Cloning, and Sequencing Analysis
3. Results
3.1. Evaluation of Radish Inbred Lines for Resistance to P. brassicae Race 4
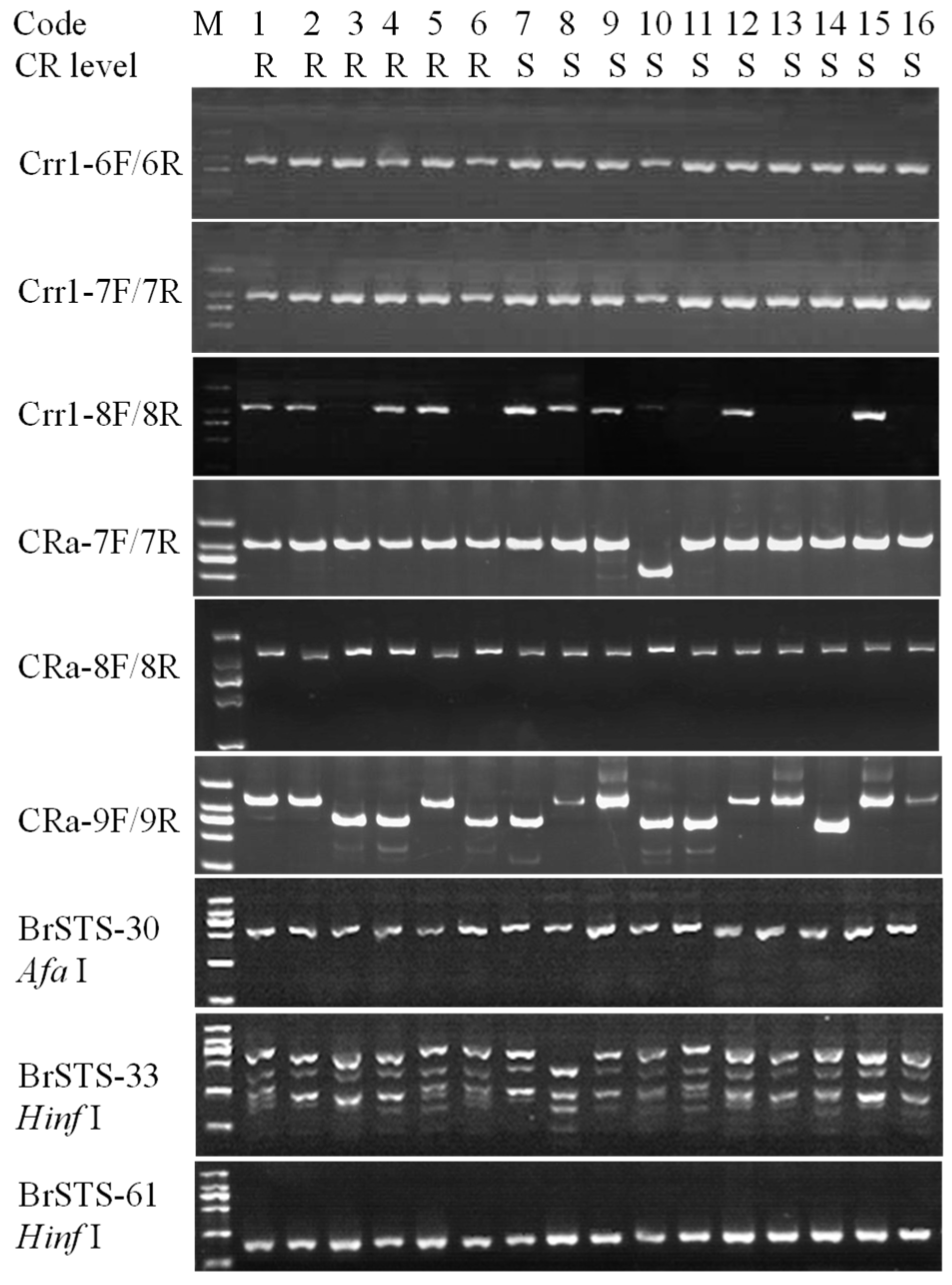
3.2. Homology of Clubroot Resistance Genes and Co-Segregation Analysis of Haplotype and Clubroot Resistance
3.3. Cloning of the RsCRa Allele in Clubroot-Susceptible and -Resistant Radish Inbred Lines
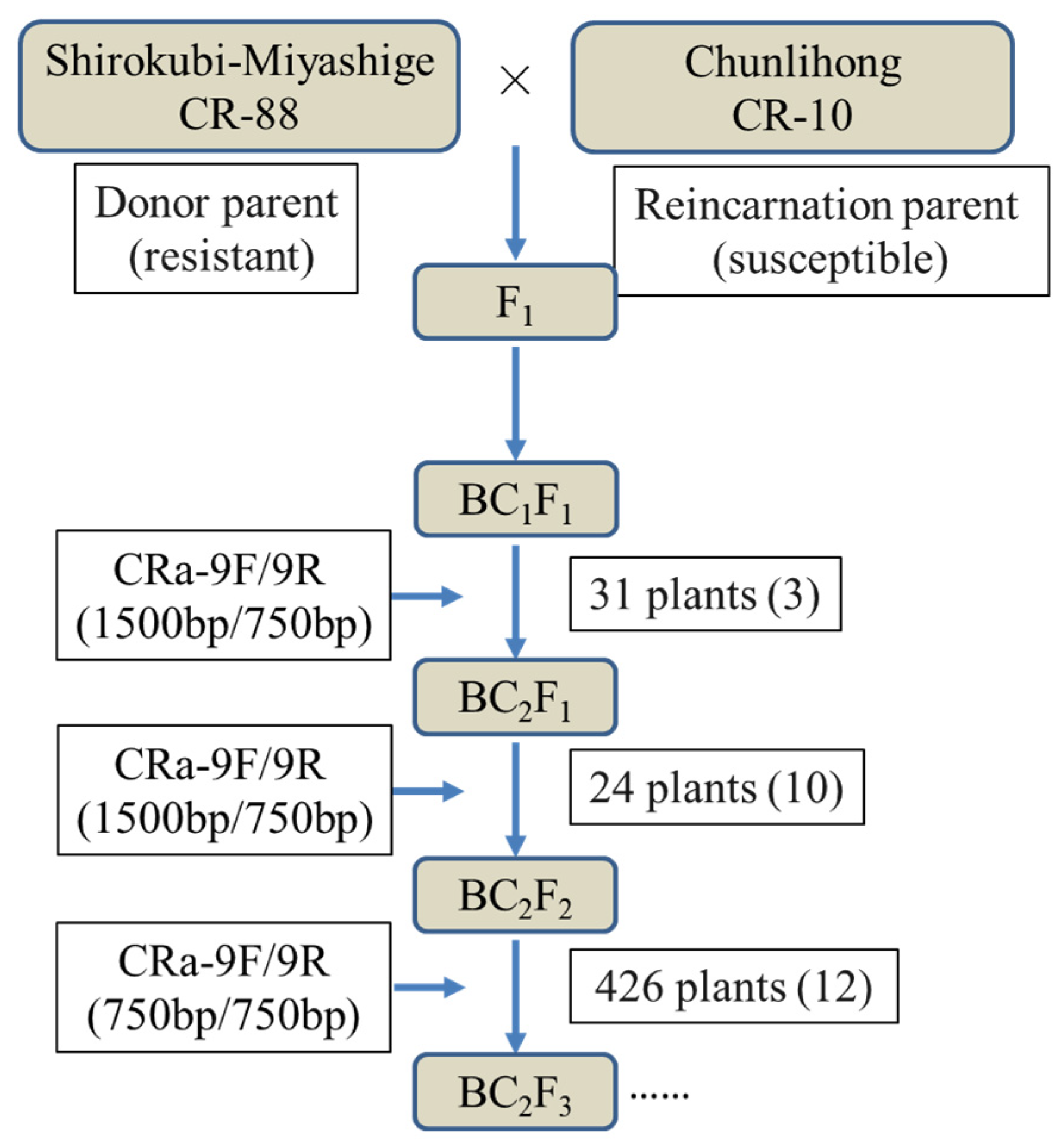
3.4. Verification of Markers for RsCRa and Marker-Assisted Backcrossing for Selection of Clubroot Resistance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hirai, M. Genetic analysis of clubroot resistance in Brassica crops. Breed. Sci. 2006, 56, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Dixon, G.R. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 2009, 28, 19. [Google Scholar] [CrossRef]
- Strelkov, S.E.; Dixon, G.R. Clubroot (Plasmodiophora brassicae) on canola and other Brassica species—Disease development, epidemiology and management. Can. J. of Plant Patho. 2014, 36 (Suppl. S1), 1–4. [Google Scholar] [CrossRef]
- Kageyama, K.; Asano, T. Life cycle of Plasmodiophora brassicae. J. Plant Growth Regul. 2009, 28, 203–211. [Google Scholar] [CrossRef]
- Rolfe, S.A.; Strelkov, S.E.; Links, M.G.; Clarke, W.E.; Robinson, S.J.; Djavaheri, M.; Malinowski, R.; Haddadi, P.; Kagale, S.; Parkin, I.A.P. The compact genome of the plant pathogen Plasmodiophora brassicae is adapted to intracellular interactions with host Brassica spp. BMC Genom. 2016, 17, 272. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.W.; Ou, Y.L.; Wang, Y.J. Occurrence and prevention of clubroot disease on cruciferous crops in Jiangxi. Bull. Plant Prot. 1955, 8, 1–4. (In Chinese) [Google Scholar]
- Wang, J.; Huang, Y.; Li, X.L.; Li, H.Z. Reseach progress in clubroot of crucifers. Plant Prot. 2011, 37, 153–158. [Google Scholar]
- Chai, A.L.; Xie, X.W.; Shi, Y.X.; Li, B.J. Research status of clubroot (Plasmodiophora brassicae) on cruciferous crops in China. Can. J. Plant Pathol. 2014, 36, 142–153. [Google Scholar] [CrossRef]
- Shen, X.Q.; Nie, K.; Wu, Q.; Zhang, Y.G.; Meng, X.H. Initial research report on differentiation identification of Chinese cabbage clubroot main physiological races. Chin. Veg. 2009, 8, 59–62. (In Chinese) [Google Scholar]
- Liu, Y.; Luo, Y.F.; Huang, X.Q.; Zhang, L.; Liu, H.Y. Research progress of differential systems for physiological races of Plasmodiophora brassicae Wor. Agric. Sci. Technol. 2012, 13, 2236–2241. [Google Scholar]
- Pang, W.X.; Liang, Y.; Zhan, Z.X.; Li, X.N.; Piao, Z.Y. Development of a Sinitic clubroot differential set for the pathotype classification of Plasmodiophora brassicae. Front. Plant Sci. 2020, 11, 568771. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, E.; Yasui, C.; Ohi, M.; Tsukada, M. Linkage analysis of RFLP markers for clubroot resistance and pigmentation in Chinese cabbage. Euphytica 1998, 104, 79–86. [Google Scholar] [CrossRef]
- Ueno, H.; Matsumoto, E.; Aruga, D.; Kitagawa, S.; Matsumura, H.; Hayashida, N. Molecular characterization of the CRa gene conferring clubroot resistance in Brassica rapa. Plant Mol. Biol. 2012, 80, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, K.; Niwa, T.; Kato, T.; Ohara, T.; Kakizaki, T.; Matsumoto, S. The tandem repeated organization of NB-LRR genes in the clubroot-resistant CRb locus in Brassica rapa L. Mol. Genet. Genom. 2017, 292, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Matsumoto, S.; Hirai, M. Identification of two loci for resistance to clubroot (Plasmodiophora brassicae Woronin) in Brassica rapa L. Theor. Appl. Genet. 2003, 107, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Kondo, M.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Hirai, M.; Matsumoto, S. Simple sequence repeat-based comparative genomics between Brassica rapa and Arabidopsis thaliana: The genetic origin of clubroot resistance. Genetics 2006, 173, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, K.; Suwabe, K.; Tomita, R.N.; Kato, T.; Nunome, T.; Fukuoka, H.; Matsumoto, S. Identification and characterization of Crr1a, a gene for resistance to clubroot disease (Plasmodiophora brassicae Woronin) in Brassica rapa L. PLoS ONE 2013, 8, e54745. [Google Scholar] [CrossRef]
- Hirai, M.; Harada, T.; Kubo, N.; Tsukada, M.; Suwabe, K.; Matsumoto, S. A novel locus for clubroot resistance in Brassica rapa and its linkage markers. Theor. Appl. Genet. 2004, 108, 639–643. [Google Scholar] [CrossRef]
- Saito, M.; Kubo, N.; Matsumoto, S.; Suwabe, K.; Tsukada, M.; Hirai, M. Fine mapping of the clubroot resistance gene, Crr3, in Brassica rapa. Theor. Appl. Genet. 2006, 114, 81–91. [Google Scholar] [CrossRef]
- Sakamoto, K.; Saito, A.; Hayashida, N.; Taguchi, G.; Matsumoto, E. Mapping of isolate-specific QTL for clubroot resistance in Chinese cabbage (Brassica rapa L. ssp. pekinensis). Theor. Appl. Genet. 2008, 117, 759–767. [Google Scholar] [CrossRef]
- Chen, J.; Jing, J.; Zhan, Z.; Zhang, T.; Zhang, C.; Piao, Z. Identification of novel QTLs for isolate-specific partial resistance to Plasmodiophora brassicae in Brassica rapa. PLoS ONE 2013, 8, e85307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, W.; Liang, S.; Li, X.; Li, P.; Yu, S.; Lim, Y.P.; Piao, Z. Genetic detection of clubroot resistance loci in a new population of Brassica rapa. Hortic. Environ. Biotechnol. 2014, 55, 540–547. [Google Scholar] [CrossRef]
- Pang, W.; Fu, P.; Li, X.; Zhan, Z.; Yu, S.; Piao, Z. Identification and mapping of the clubroot resistance gene CRd in Chinese cabbage (Brassica rapa ssp. pekinensis). Front. Plant Sci. 2018, 9, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.R.; Oh, S.H.; Chhapekar, S.S.; Dhandapani, V.; Lee, C.Y.; Rameneni, J.J.; Ma, Y.; Choi, G.J.; Lee, S.S.; Lim, Y.P. Quantitative trait locus mapping of clubroot resistance and Plasmodiophora brassicae pathotype banglim-specific marker development in Brassica rapa. Int. J. Mol. Sci. 2020, 21, 4157. [Google Scholar] [CrossRef] [PubMed]
- Diederichsen, E.; Frauen, M.; Linders, E.G.A.; Hatakeyama, K.; Hirai, M. Status and perspectives of clubroot resistance breeding in crucifer crops. J. Plant Growth Regul. 2009, 28, 265–281. [Google Scholar] [CrossRef]
- Zhang, T.; Wu, D.; Zhao, Z.; Wang, Z.; Piao, Z.Y. Development of near isogenic lines for clubroot resistance in Chinese cabbage and their assessment. Mol. Plant Breed. 2012, 6, 722–730. [Google Scholar]
- Fredua-Agyeman, R.; Yu, Z.; Hwang, S.F.; Strelkov, S.E. Genome-wide mapping of loci associated with resistance to clubroot in Brassica napus ssp. napobrassica (Rutabaga) accessions from Nordic Countries. Front. Plant Sci. 2020, 11, 742. [Google Scholar] [CrossRef]
- Rowe, R.C. Evaluation of radish cultivars for resistance to clubroot (Plasmodiophora brassicae) race 6 for Midwestern United States. Plant Disease 1980, 64, 462–464. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Yuan, Y.; Wei, X.; Zhang, X.; Wang, H.; Song, J.; Li, X. A new identification method reveals the resistance of an extensive-source radish collection to Plasmodiophora brassicae race 4. Agronomy 2021, 11, 792. [Google Scholar] [CrossRef]
- Kamei, A.; Tsuro, M.; Kubo, N.; Hayashi, T.; Wang, N.; Fujimura, T.; Hirai, M. QTL mapping of clubroot resistance in radish (Raphanus sativus L.). Theor. Appl. Genet. 2010, 120, 1021–1027. [Google Scholar] [CrossRef]
- Gan, C.; Deng, X.; Cui, L.; Yu, X.; Yuan, W.; Dai, Z.; Yao, M.; Pang, W.; Ma, Y.; Yu, X.; et al. Construction of a high-density genetic linkage map and identification of quantitative trait loci associated with clubroot resistance in radish (Raphanus sativus L.). Mol. Breed. 2019, 39, 116. [Google Scholar] [CrossRef]
- Scheijgrond, W.; Vos, H. Investigation on the susceptibility to club root. Euphytica 1954, 3, 125–139. [Google Scholar] [CrossRef]
- Ning, Y.; Wang, Y.; Fang, Z.; Zhuang, M.; Zhang, Y.; Lv, H.; Liu, Y.; Li, Z.; Yang, L. Identification and characterization of resistance for Plasmodiophora brassicae race 4 in cabbage (Brassica oleracea var. capitata). Australas Plant Pathol. 2018, 47, 531–541. [Google Scholar] [CrossRef]
- Williams, P.H. A system for the determination of races of Plasmodiophora brassicae that infect cabbage and rutabaga. Phytopathology 1966, 56, 624–626. [Google Scholar]
- Murray, H.; Thompson, W. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.H.; Yue, Z.; Mei, S.Y.; Qiu, Y.; Yang, X.H.; Chen, X.H.; Cheng, F.; Wu, Z.Y.; Sun, Y.Y.; Jing, Y.; et al. A de novo genome of a Chinese radish cultivar. Hort. Plant J. 2015, 1, 155–164. [Google Scholar]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Zhang, L.; Cai, X.; Wu, J.; Liu, M.; Grob, S.; Cheng, F.; Liang, J.; Cai, C.; Liu, Z.; Liu, B.; et al. Improved Brassica rapa reference genome by single-molecule sequencing and chromosome conformation capture technologies. Hortic. Res. 2018, 5, 50. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, H. Studies on breeding of clubroot resistance in cole crops (in Japanese with English summary). Bull. Natl. Res. Inst. Veg. Ornam. Plants Tea Jpn. Ser. A 1993, 7, 1–165. [Google Scholar]
- Crute, I.R.; Gray, A.R.; Crisp, P.; Buczacki, S.T. Variation in Plasmodiophora brassicae and resistance to clubroot disease in brassicas and allied crops—A critical review. Plant Breed. Abstr. 1980, 50, 91–104. [Google Scholar]
- Chen, X.; Wang, C.; Zhang, X.X.; Wang, S. The inoculation system of cabbage clubroot and Screening of resistant germplasm resources. Plant Prot. 2015, 41, 121–126. [Google Scholar]
- Matsumoto, E.; Ueno, H.; Aruga, D.; Sakamoto, K.; Hayashida, N. Accumulation of three clubroot resistance genes through marker-assisted selection in Chinese cabbage (Brassica rapa spp. pekinensis). J. Jpn. Soc. Hortic. Sci. 2012, 81, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Mehraj, H.; Akter, A.; Miyaji, N.; Miyazaki, J.; Shea, D.J.; Fujimoto, R.; Doullah, M. Genetics of clubroot and fusarium wilt disease resistance in Brassica vegetables: The application of marker assisted breeding for disease resistance. Plants 2020, 9, 726. [Google Scholar] [CrossRef]
- Horikoshi, N.; Tairako, K. Occurrence of clubroot in Japanese radish caused by Plasmodiophora brassicae Woronin in Fukushima. Annu. Rep. Plant Prot. North Jpn. 2002, 53, 58–60. [Google Scholar]
- Piao, Z.Y.; Wu, D.; Wang, M.; Zhang, T. Marker-assisted selection of near isogenic lines for clubroot resistant gene in Chinese cabbage. Acta Hortic. Sin. 2010, 37, 1264–1272. [Google Scholar]
- Chu, M.; Yu, F.; Falk, K.C.; Liu, X.; Zhang, X.; Chang, A.; Peng, G. Identification of the clubroot resistance gene Rpb1 and introgression of the resistance into canola breeding lines using a marker-assisted approach. Acta. Hortic. 2013, 1005, 599–605. [Google Scholar] [CrossRef]
- Tomita, H.; Shimizu, M.; Doullah, M.A.U.; Fujimoto, R.; Okazaki, K. Accumulation of quantitative trait loci conferring broad-spectrum clubroot resistance in Brassica oleracea. Mol. Breed. 2013, 32, 889–900. [Google Scholar] [CrossRef]
- Shah, N.; Sun, J.; Yu, S.; Yang, Z.; Wang, Z.; Huang, F.; Dun, B.; Gong, J.; Liu, Y.; Li, Y.; et al. Genetic variation analysis of field isolates of clubroot and their responses to Brassica napus lines containing resistant genes CRb and PbBa8.1 and their combination in homozygous and heterozygous state. Mol. Breed. 2019, 39, 153. [Google Scholar] [CrossRef]
- Kawasaki, M.; Ohara, T.; Ishida, M.; Takahata, Y.; Hatakeyama, K. Development of novel clubroot resistant rapeseed lines (Brassica napus L.) effective against Japanese field isolates by marker assisted selection. Breed. Sci. 2021, 71, 528–537. [Google Scholar] [CrossRef]
- Jeong, Y.M.; Kim, N.; Ahn, B.O.; Oh, M.; Chung, W.H.; Chung, H.; Jeong, S.; Lim, K.B.; Hwang, Y.J.; Kim, G.B. Elucidating the triplicated ancestral genome structure of radish based on chromosome-level comparison with the Brassica genomes. Theor. Appl. Genet. 2016, 129, 1357–1372. [Google Scholar] [CrossRef]
- Xing, G.M.; Long, M.H.; Tanaka, S.; Fujieda, K. Clubroot resistance in Brassicoraphanus [hybrid of cabbage × radish]. J. Fac. Agric. Kyushu Univ. 1989, 33, 189–194. [Google Scholar] [CrossRef]
- Hagimori, M.; Nagaoka, M.; Kato, N.; Yoshikawa, H. Production and characterization of somatic hybrids between the Japanese radish and cauliflower. Theor. Appl. Genet. 1992, 84, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Akaba, M.; Kaneko, Y.; Hatakeyama, K.; Ishida, M.; Bang, S.W.; Matsuzawa, Y. Identification and evaluation of clubroot resistance of radish chromosome using a Brassica napus-Raphanus sativus monosomic addition line. Breed. Sci. 2009, 59, 203–206. [Google Scholar] [CrossRef] [Green Version]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef]
- Mun, J.H.; Yu, H.J.; Park, S.; Park, B.S. Genome-wide identification of NBS-encoding resistance genes in Brassica rapa. Mol. Genet. Genom. 2009, 282, 617–631. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Tehrim, S.; Zhang, F.; Tong, C.; Huang, J.; Cheng, X.; Dong, C.; Zhou, Y.; Qin, R.; Hua, W.; et al. Genome-wide comparative analysis of NBS-encoding genes between Brassica species and Arabidopsis thaliana. BMC Genom. 2014, 15, 3. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Hu, T.; Wang, W.; Hu, H.; Wei, Q.; Bao, C. Investigation of evolutionary and expressional relationships in the function of the leucine-rich repeat receptor-like protein kinase gene family (LRR-RLK) in the radish (Raphanus sativus L.). Sci. Rep. 2019, 9, 6937. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Song, Z.; Zhang, W.; Xu, L.; Su, X.; Muleke, E.M.M.; Liu, L. Identification and characterization of expressed TIR-and non-TIR-NBS-LRR resistance gene analogous sequences from radish (Raphanus sativus L.) de novo transcriptome. Sci. Hortic. 2017, 216, 284–292. [Google Scholar] [CrossRef]
- Hulbert, S.H.; Webb, C.A.; Smith, S.M.; Sun, Q. Resistance gene complexes: Evolution and utilization. Annu. Rev. Phytopathol. 2001, 39, 285–312. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Provenance | Total | Immune DI = 0 | Highly Resistant 0 < DI ≤ 1 | Slightly Susceptible 1 < DI ≤ 2 | Moderately Susceptible 2 < DI ≤ 3 | Highly Susceptible 3 < DI ≤ 4 |
|---|---|---|---|---|---|---|
| China | 40 | 6 | 1 | 3 | 16 | 14 |
| South Korea | 33 | 11 | 3 | 8 | 5 | 6 |
| Japan | 16 | 10 | 1 | 5 | 0 | 0 |
| Other | 6 | 0 | 1 | 5 | 0 | 0 |
| Total | 95 | 27 | 6 | 21 | 21 | 20 |
| B. rapa | R. sativus | |||||||
|---|---|---|---|---|---|---|---|---|
| Resistant Gene | GenBank ID | Gene/QTL Location | Flanking Regin (±20~30 kb) | Homologous Gene (Marker) | Gene/QTL Location | Query Cov. | Per. Ident. | Synteny Regin |
| Crr1 | AB605024 | A08: 12271628–12276052 | A08: 12241628–12306052 | Rsa10003637 | R08: 27134943–27138766 | 86% | 91.69% | R08: 27084498–27148246 |
| Rsa10003639 | R08: 27124803–27128501 | 42% | 83.38% | |||||
| CRa | AB751516 | A03: 25523235–25546244 | A03: 25503710–25565309 | Rsa10025569 | R04: 5211349–5232999 | 74% | 86.79% | R04: 5197160–5238084 |
| Rsa10025571 | R04: 5204303–5209454 | 81% | 85.93% | |||||
| Crr3 (markers) | ||||||||
| BrSTS-54 STS | AB265763 | A03: 16156827–16157418 | A03: 16048124–16156827 | BrSTS-33 (QTL marker) | R05: 847681–848363 | - | - | R05: 798486–891819 |
| BrSTS-61 STS | AB265769 | A03: 16099477–16098797 | BrSTS-30 (QTL marker) | |||||
| BrSTS-78 SSR | AB265777 | A03: 16048124–16048942 | BrSTS-61 (QTL marker) | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Wang, Y.; Qian, H.; Zhang, Z.; Zhang, L. Evaluation of Germplasm and Development of Markers for Resistance to Plasmodiophora brassicae in Radish (Raphanussativus L.). Agronomy 2022, 12, 554. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030554
Wang Q, Wang Y, Qian H, Zhang Z, Zhang L. Evaluation of Germplasm and Development of Markers for Resistance to Plasmodiophora brassicae in Radish (Raphanussativus L.). Agronomy. 2022; 12(3):554. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030554
Chicago/Turabian StyleWang, Qingbiao, Yanping Wang, Huihui Qian, Ziye Zhang, and Li Zhang. 2022. "Evaluation of Germplasm and Development of Markers for Resistance to Plasmodiophora brassicae in Radish (Raphanussativus L.)" Agronomy 12, no. 3: 554. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030554