
EuPIP1;2, a Plasma Membrane Aquaporin Gene from Eucommia ulmoides, Enhances Drought and Salt Tolerance in Transgenic Tobacco


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Gene Cloning and Bioinformatic Analysis of EuPIP1;2
2.3. Quantitative RT-PCR and RT-PCR Analysis
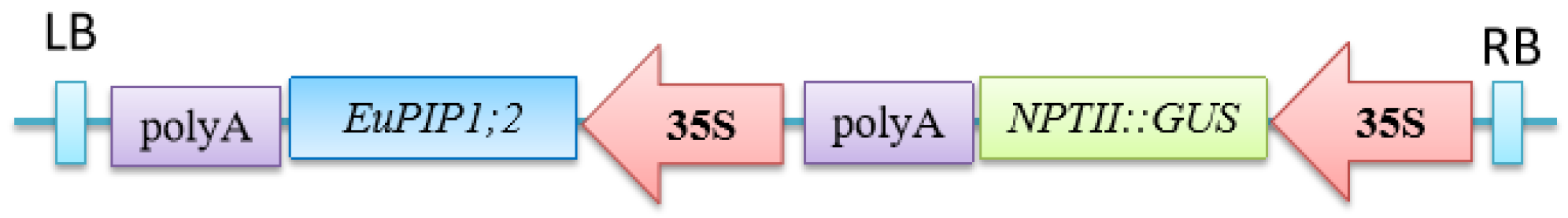
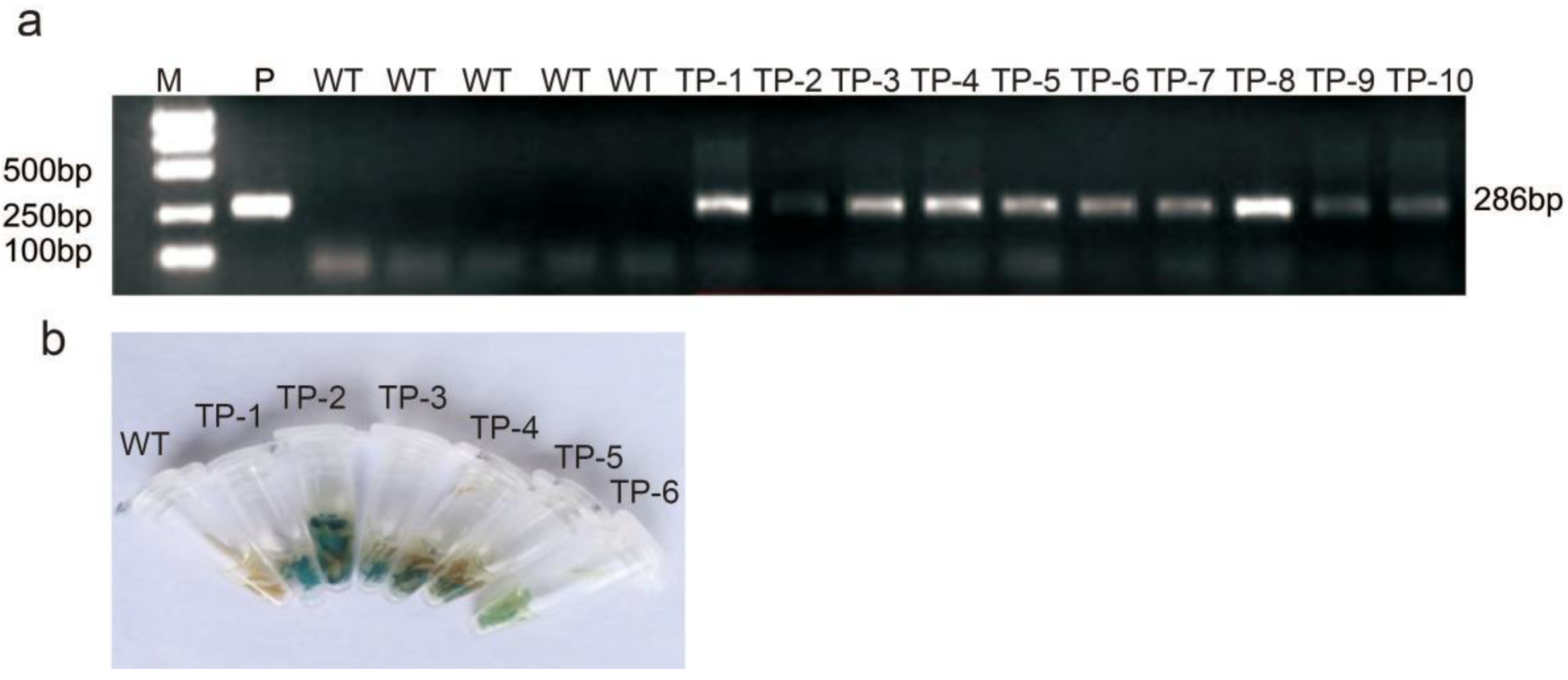
2.4. Vector Construction and Plant Transformation
2.5. Protein Subcellular Localization
2.6. Drought Treatment
2.7. Determination of Water Loss
2.8. Salt Treatment
2.9. Determination of Malondialdehyde and Proline Contents
2.10. Statistical Analysis
3. Results
3.1. Cloning and Characterization of EuPIP1;2
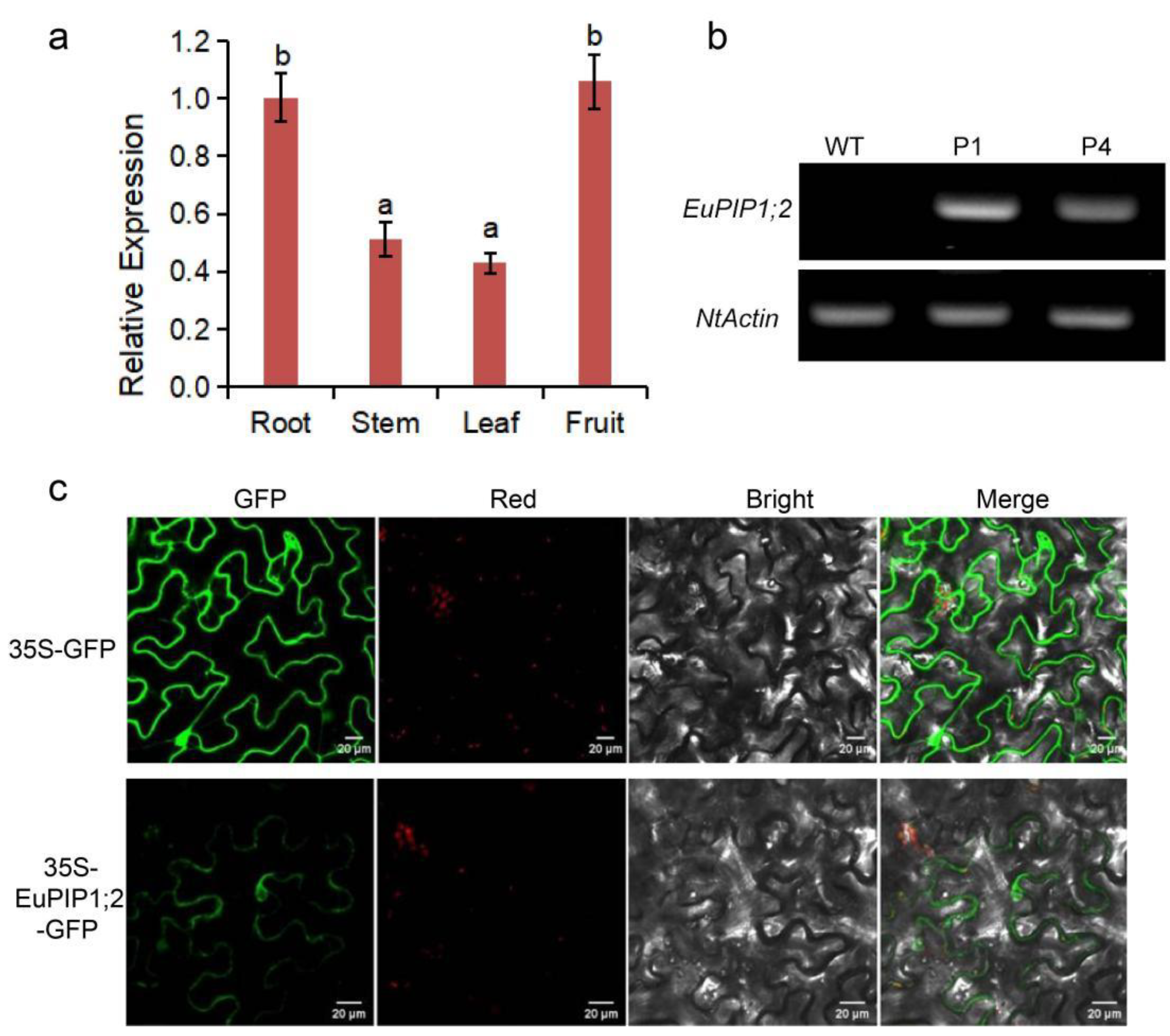
3.2. Expression Pattern of EuPIP1;2
3.3. Subcellular Localization of EuPIP1;2 Protein
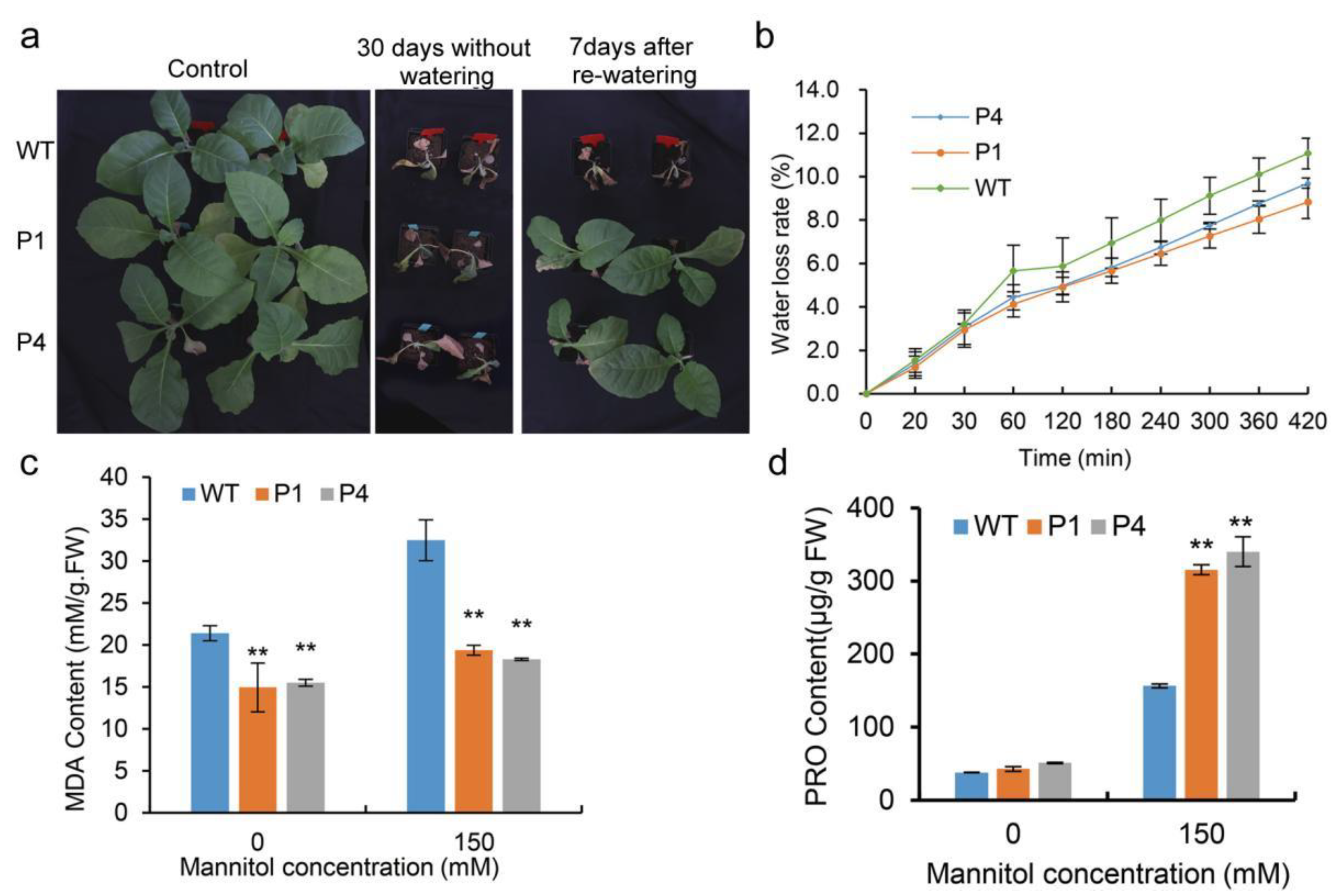
3.4. Overexpression of EuPIP1;2 Improved Drought Tolerance of Transgenic Tobacco
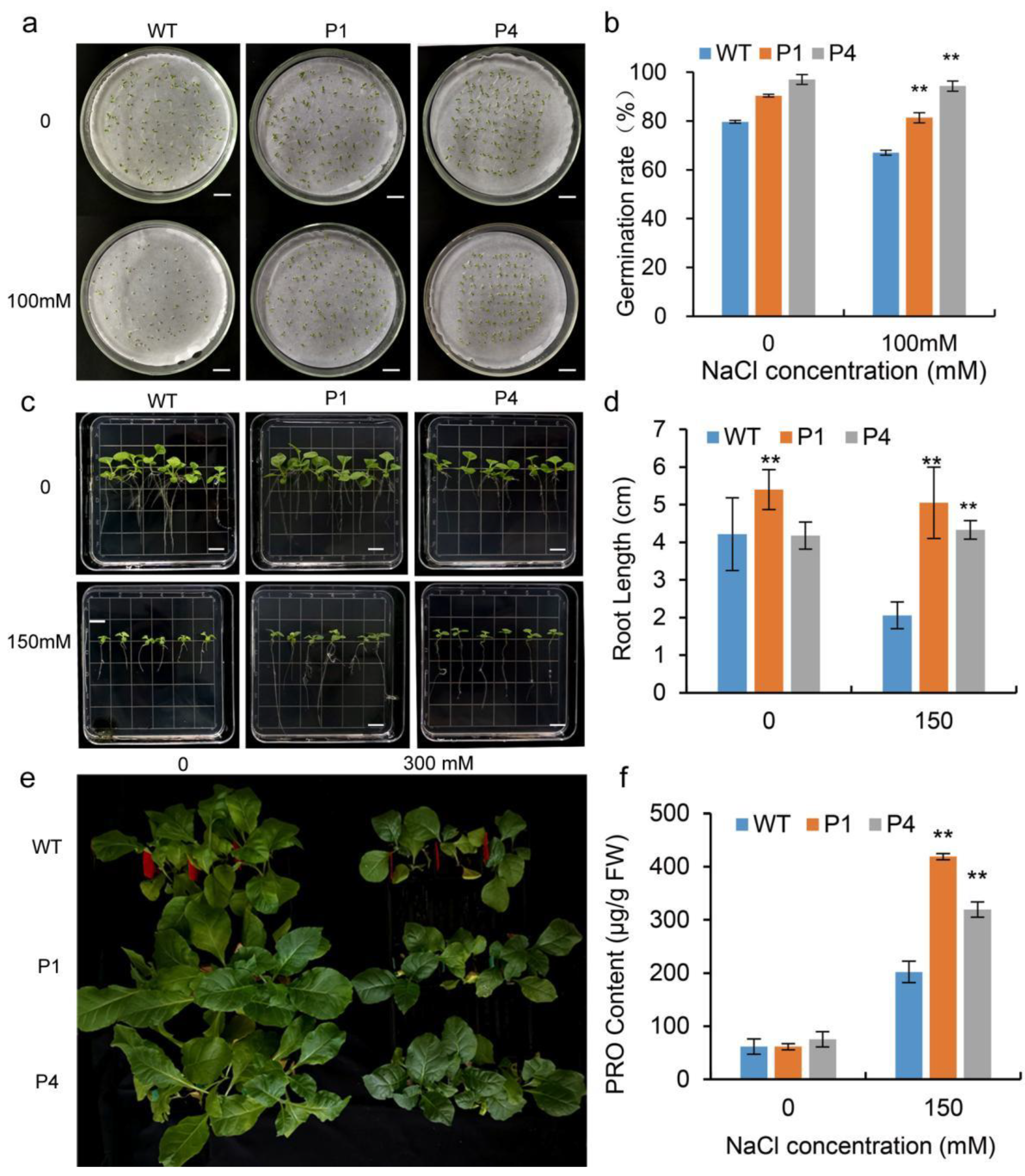
3.5. Effects on Salt Tolerance of Transgenic Tobacco
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R.; Porcel, R.; Lozano, J.M.R. Regulation of root water uptake under abiotic stress conditions. J. Exp. Bot. 2011, 63, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Santoni, V.; Luu, D.-T.; Wudick, M.M.; Verdoucq, L. The cellular dynamics of plant aquaporin expression and functions. Curr. Opin. Plant Biol. 2009, 12, 690–698. [Google Scholar] [CrossRef]
- Quigley, F.; Rosenberg, J.M.; Shachar-Hill, Y.; Bohnert, H.J. From genome to function: The Arabidopsis aquaporins. Genome Biol. 2001, 3, RESEARCH0001. [Google Scholar] [CrossRef] [PubMed]
- Chaumont, F.; Barrieu, F.; Wojcik, E.; Chrispeels, M.J.; Jung, R. Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol. 2001, 125, 1206–1215. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.Y.; Ali, Z.; Wang, C.B.; Xu, L.; Yi, J.X.; Xu, Z.L.; Liu, X.Q.; He, X.L.; Huang, Y.H.; Khan, I.A.; et al. Genome-wide sequence characterization and expression analysis of major intrinsic proteins in soybean (Glycine max L.). PLoS ONE 2013, 8, e56312. [Google Scholar] [CrossRef]
- Reuscher, S.; Akiyama, M.; Mori, C.; Aoki, K.; Shibata, D.; Shiratake, K. Genome-wide identification and expression analysis of aquaporins in tomato. PLoS ONE 2013, 8, e79052. [Google Scholar] [CrossRef] [Green Version]
- Park, W.; Scheffler, B.E.; Bauer, P.J.; Campbell, B.T. Identification of the family of aquaporin genes and their expression in upland cotton (Gossypium hirsutum L.). BMC Plant Biol. 2010, 10, 142. [Google Scholar] [CrossRef] [Green Version]
- Tao, P.; Zhong, X.; Li, B.; Wang, W.; Yue, Z.; Lei, J.; Guo, W.; Huang, X. Genome-wide identification and characterization of aquaporin genes (AQPs) in Chinese cabbage (Brassica rapa ssp. pekinensis). Mol. Genet. Genom. 2014, 289, 1131–1145. [Google Scholar] [CrossRef]
- Chaumont, F.; Tyerman, S. Plant Aquaporins; Springer: Berlin/Heidelberg, Germany, 2017; Volume 667. [Google Scholar]
- Vandeleur, R.K.; Mayo, G.; Shelden, M.C.; Gilliham, M.; Kaiser, B.N.; Tyerman, S.D. The role of plasma membrane intrinsic protein aquaporins in water transport through roots: Diurnal and drought stress responses reveal different strategies be-tween isohydric and anisohydric cultivars of grapevine. Plant Physiol. 2008, 149, 445–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Feng, M.; Chen, W.; Zhou, X.; Lu, J.; Wang, Y.; Li, Y.; Jiang, C.-Z.; Gan, S.-S.; Ma, N.; et al. In rose, transcription factor PTM balances growth and drought survival via PIP2;1 aquaporin. Nat. Plants 2019, 5, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Lin, R.; Pu, L.; Wang, Z.; Mei, Q.; Zhang, M.; Jian, S. Ectopic expression of CrPIP2;3, a plasma membrane intrinsic protein gene from the halophyte Canavalia rosea, enhances drought and salt-alkali stress tolerance in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 565. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fukumoto, T.; Matsumoto, T.; Gena, P.; Frascaria, D.; Kaneko, T.; Katsuhara, M.; Zhong, S.; Sun, X.; Zhu, Y.; et al. Aquaporin OsPIP1;1 promotes rice salt resistance and seed germination. Plant Physiol. Biochem. 2013, 63, 151–158. [Google Scholar] [CrossRef]
- Zhang, B.; Xie, L.; Sun, T.; Ding, B.; Li, Y.; Zhang, Y. Chrysanthemum morifolium aquaporin genes CmPIP1 and CmPIP2 are involved in tolerance to salt stress. Sci. Hortic. 2019, 256, 108627. [Google Scholar] [CrossRef]
- Xu, Y.; Hu, W.; Liu, J.; Zhang, J.; Jia, C.; Miao, H.; Xu, B.; Jin, Z. A banana aquaporin gene, MaPIP1;1, is involved in tol-erance to drought and salt stresses. BMC Plant Biol. 2014, 14, 59. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Han, N.; Wu, H.; Jia, J.; Guo, J. Plasma membrane intrinsic protein SlPIP1;7 promotes root growth and enhances drought stress tolerance in transgenic tomato (Solanum lycopersicum) plants. Plant Breed. 2021, 140, 1102–1114. [Google Scholar] [CrossRef]
- He, X.; Wang, J.; Li, M.; Hao, D.; Yang, Y.; Zhang, C.; He, R.; Tao, R. Eucommia ulmoides Oliv.: Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2013, 151, 78–92. [Google Scholar] [CrossRef]
- Wuyun, T.-N.; Wang, L.; Liu, H.; Wang, X.; Zhang, L.; Bennetzen, J.L.; Li, T.; Yang, L.; Liu, P.; Du, L.; et al. The hardy rubber tree genome provides insights into the evolution of polyisoprene biosynthesis. Mol. Plant 2017, 11, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.-F.; Zhao, D.-G. Expression of IPT in asakura-sanshoo (Zanthoxylum piperitum (L.) DC. f. inerme makino) alters tree architecture, delays leaf senescence, and changes leaf essential oil composition. Plant Mol. Biol. Rep. 2015, 34, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Gyokusen, M.; Nakazawa, Y.; Gyokusen, K. Selection of housekeeping genes for transgene expression analysis in Eucommia ulmoides oliver using real-time RT-PCR. J. Bot. 2010, 2010, 230961. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yu, Z.; da Silva, J.A.T.; Wen, D. Identification of aquaporin members in Acacia auriculiformis and functional characterization of AaPIP1-2 involved in drought stress. Environ. Exp. Bot. 2021, 185, 104425. [Google Scholar] [CrossRef]
- Zhao, D.; Zeng, X. In vitro regeneration and Agrobacterium tumefaciens-mediated genetic transformation in asakura-sanshoo (Zanthoxylum piperitum (L.) DC. F. inerme Makino) an important medicinal plant. Pharmacogn. Mag. 2015, 11, 374–380. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Liu, Y.; Zhu, Y.; Zhao, D. Selectable gene auto-excision via a cold inducible gene deletor system. Afr. J. Agric. Res. 2010, 5, 2426–2433. [Google Scholar]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Li, M.; Li, M.; Li, D.; Wang, S.-M.; Yin, H. Overexpression of the Zygophyllum xanthoxylum aquaporin, ZxPIP1;3, promotes plant growth and stress tolerance. Int. J. Mol. Sci. 2021, 22, 2112. [Google Scholar] [CrossRef]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; van Arkel, G.; Pereira, A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis [W]. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [Green Version]
- Benga, G. Water channel proteins (later called aquaporins) and relatives: Past, present, and future. IUBMB Life 2009, 61, 112–133. [Google Scholar] [CrossRef]
- Li, S.; Liu, J.; An, Y.; Cao, Y.; Liu, Y.; Zhang, J.; Geng, J.; Hu, T.; Yang, P.; Tianming, H. MsPIP2;2, a novel aquaporin gene from Medicago sativa, confers salt tolerance in transgenic Arabidopsis. Environ. Exp. Bot. 2019, 165, 39–52. [Google Scholar] [CrossRef]
- Leng, H.; Jiang, C.; Song, X.; Lu, M.; Wan, X. Poplar aquaporin PIP1;1 promotes Arabidopsis growth and development. BMC Plant Biol. 2021, 21, 253. [Google Scholar] [CrossRef] [PubMed]
- Siefritz, F.; Tyree, M.T.; Lovisolo, C.; Schubert, A.; Kaldenhoff, R. PIP1 plasma membrane aquaporins in tobacco: From cellular effects to function in plants. Plant Cell 2002, 14, 869–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Gao, F.; Bing, J.; Sun, W.; Feng, X.; Ma, X.; Zhou, Y.; Zhang, G. Overexpression of the jojoba aquaporin gene, ScPIP1, enhances drought and salt tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2019, 20, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Xiao, Y.; Li, Y.; Hu, P.; Wang, Z.; Yang, G.; Wang, J. Overexpression of CfPIP1-1, CfPIP1-2, and CfPIP1-4 genes of Catalpa fargesii in transgenic Arabidopsis thaliana under drought stress. J. For. Res. 2020, 32, 285–296. [Google Scholar] [CrossRef]
- Wang, L.; Li, Q.; Lei, Q.; Feng, C.; Zheng, X.; Zhou, F.; Li, L.; Liu, X.; Wang, Z.; Kong, J. Ectopically expressing MdPIP1;3, an aquaporin gene, increased fruit size and enhanced drought tolerance of transgenic tomatoes. BMC Plant Biol. 2017, 17, 246. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Tang, H.; Yu, Q.; Li, Z.; Liu, F.; Su, W.; Zhang, C.; Ling, H.; Luo, J.; Su, Y.; Que, Y. A PIP-mediated osmotic stress signaling cascade plays a positive role in the salt tolerance of sugarcane. BMC Plant Biol. 2021, 21, 589. [Google Scholar] [CrossRef]
- Zhou, S.; Hu, W.; Deng, X.; Ma, Z.; Chen, L.; Huang, C.; Wang, C.; Wang, J.; He, Y.; Yang, G.; et al. Overexpression of the wheat aquaporin gene, TaAQP7, enhances drought tolerance in transgenic tobacco. PLoS ONE 2012, 7, e52439. [Google Scholar] [CrossRef] [Green Version]
- Hare, P.; Cress, W. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul. 1997, 21, 79–102. [Google Scholar] [CrossRef]
- Artus, N.N.; Uemura, M.; Steponkus, P.L.; Gilmour, S.J.; Lin, C.; Thomashow, M.F. Constitutive expression of the cold-regulated Arabidopsis thaliana COR15a gene affects both chloroplast and protoplast freezing tolerance. Proc. Natl. Acad. Sci. USA 1996, 93, 13404–13409. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequences | Purpose |
|---|---|---|
| EuPIP1;2C-F | 5′-GGGAGTTTCTAGAACGAACAACA-3′ | Gene cloning |
| EuPIP1;2C-R | 5′-TCAAGAGGAGCTCTTAAATGGGA-3′ | |
| EuPIP1;2-F | 5′-TGAACCACGGCTACACCA-3′ | qRT-PCR and RT-PCR |
| EuPIP1;2-R | 5′-AACCCAATCGGAAGAGGC-3′ | |
| EuActin-F | 5′-TGACGGAGCGTGGTTACTCATTCA-3′ | qRT-PCR |
| EuActin-R | 5′-TCTTGGCAGTCTCCATTTCCTGGT-3′ | |
| EuPIP1;2J-F | 5′-TGAACCACGGCTACACCA-3′ | Transgenic plant PCR detection |
| EuPIP1;2J-R | 5′-AAGGGACCCACCCAGAAA-3′ | |
| NtActin-F | 5′-TGGTTAAGGCTGGATTTGCT-3′ | RT-PCR |
| NtActin-R | 5′-TGCATCCTTTTGACCCATAC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Li, J.; Huang, Y.; Li, Y.; Su, C.; Zeng, X. EuPIP1;2, a Plasma Membrane Aquaporin Gene from Eucommia ulmoides, Enhances Drought and Salt Tolerance in Transgenic Tobacco. Agronomy 2022, 12, 615. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030615
Chen J, Li J, Huang Y, Li Y, Su C, Zeng X. EuPIP1;2, a Plasma Membrane Aquaporin Gene from Eucommia ulmoides, Enhances Drought and Salt Tolerance in Transgenic Tobacco. Agronomy. 2022; 12(3):615. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030615
Chicago/Turabian StyleChen, Jiajia, Jianrong Li, Yanhui Huang, Yan Li, Changfeng Su, and Xiaofang Zeng. 2022. "EuPIP1;2, a Plasma Membrane Aquaporin Gene from Eucommia ulmoides, Enhances Drought and Salt Tolerance in Transgenic Tobacco" Agronomy 12, no. 3: 615. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030615