
Morphophysiological Responses of Two Cool-Season Turfgrasses with Different Shade Tolerances

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Cultivation
2.2. Turf Quality (TQ) and Turf Color Intensity (TCI)
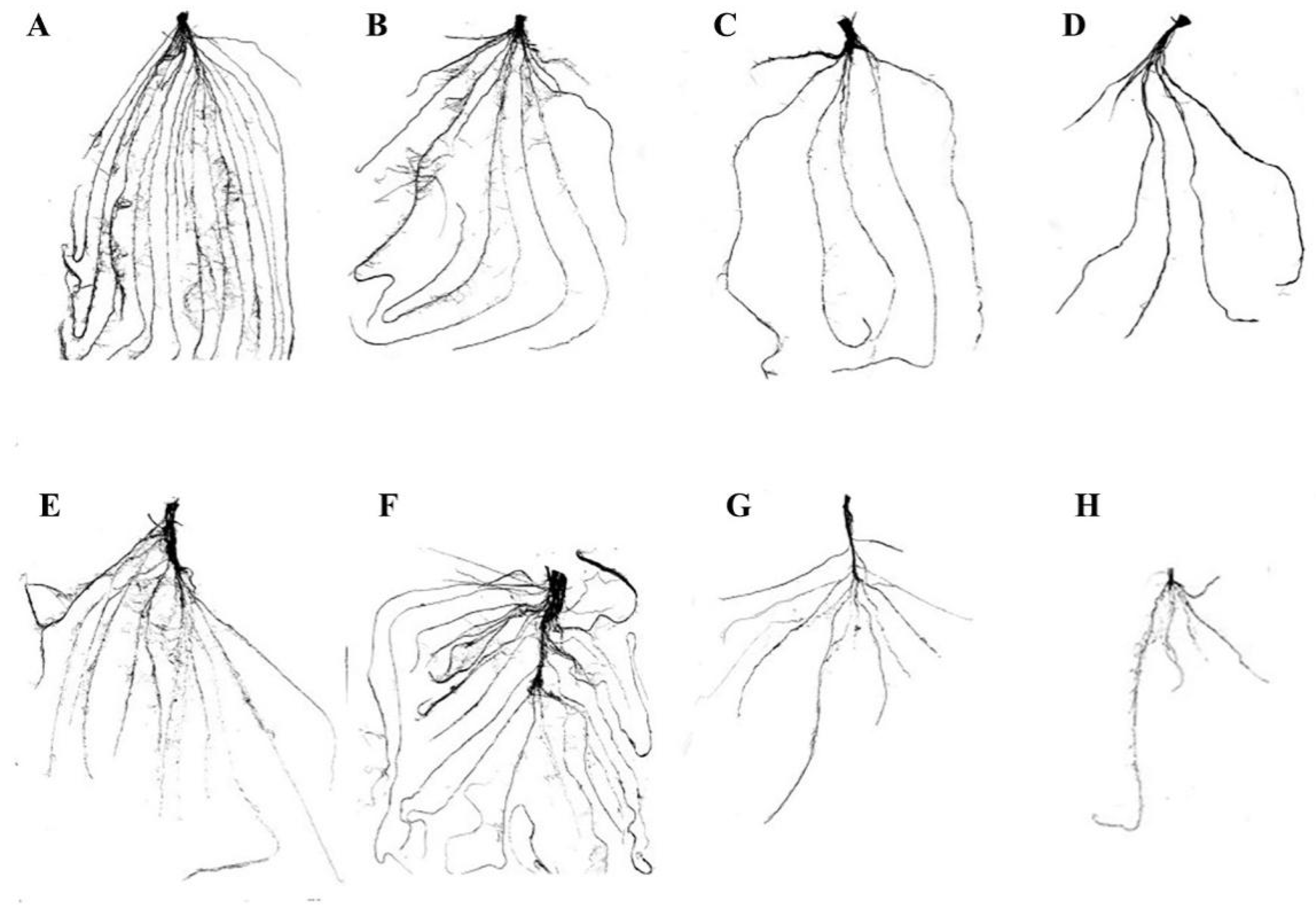
2.3. Root Morphology
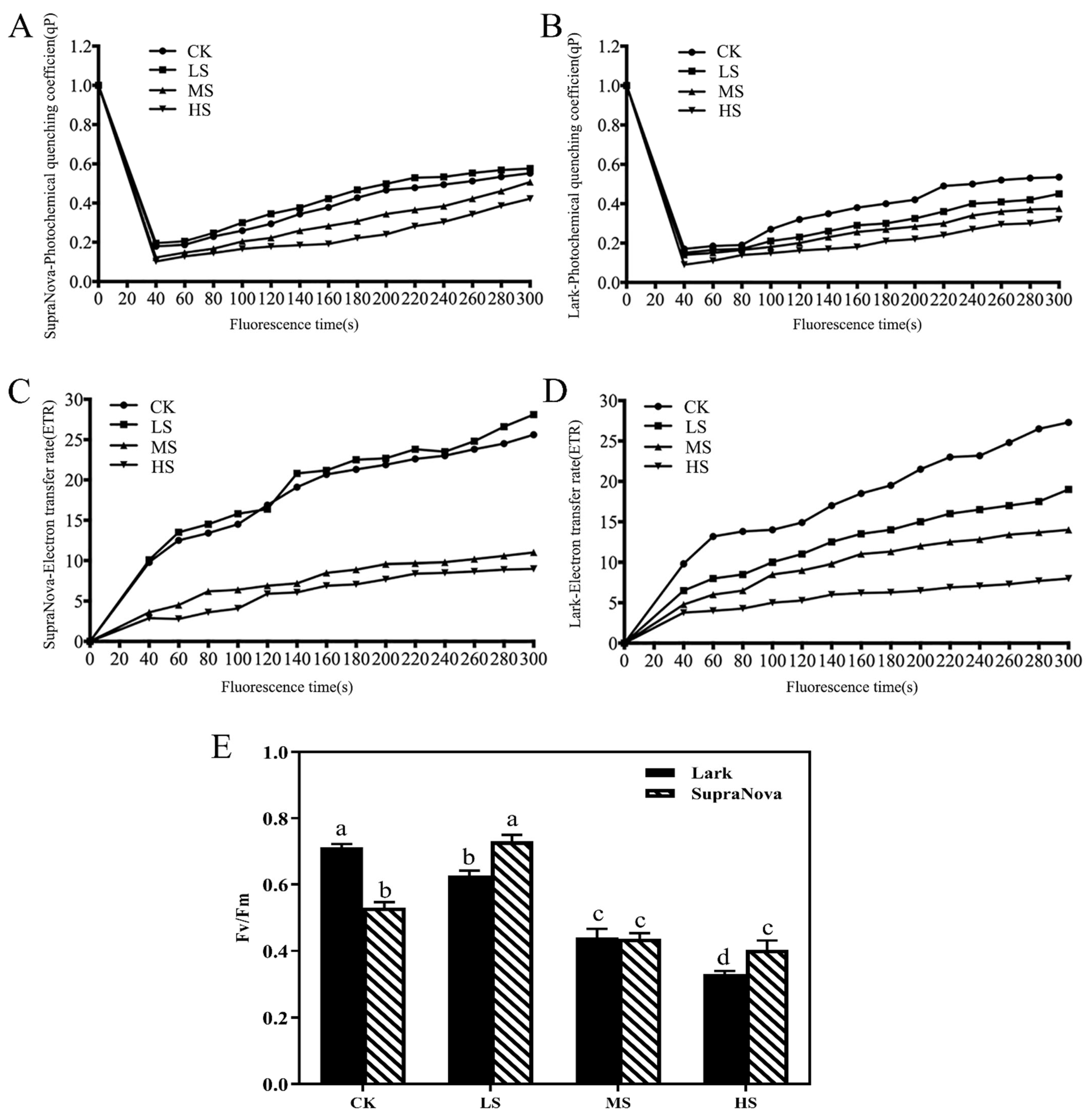
2.4. Chlorophyl a Fluorescence
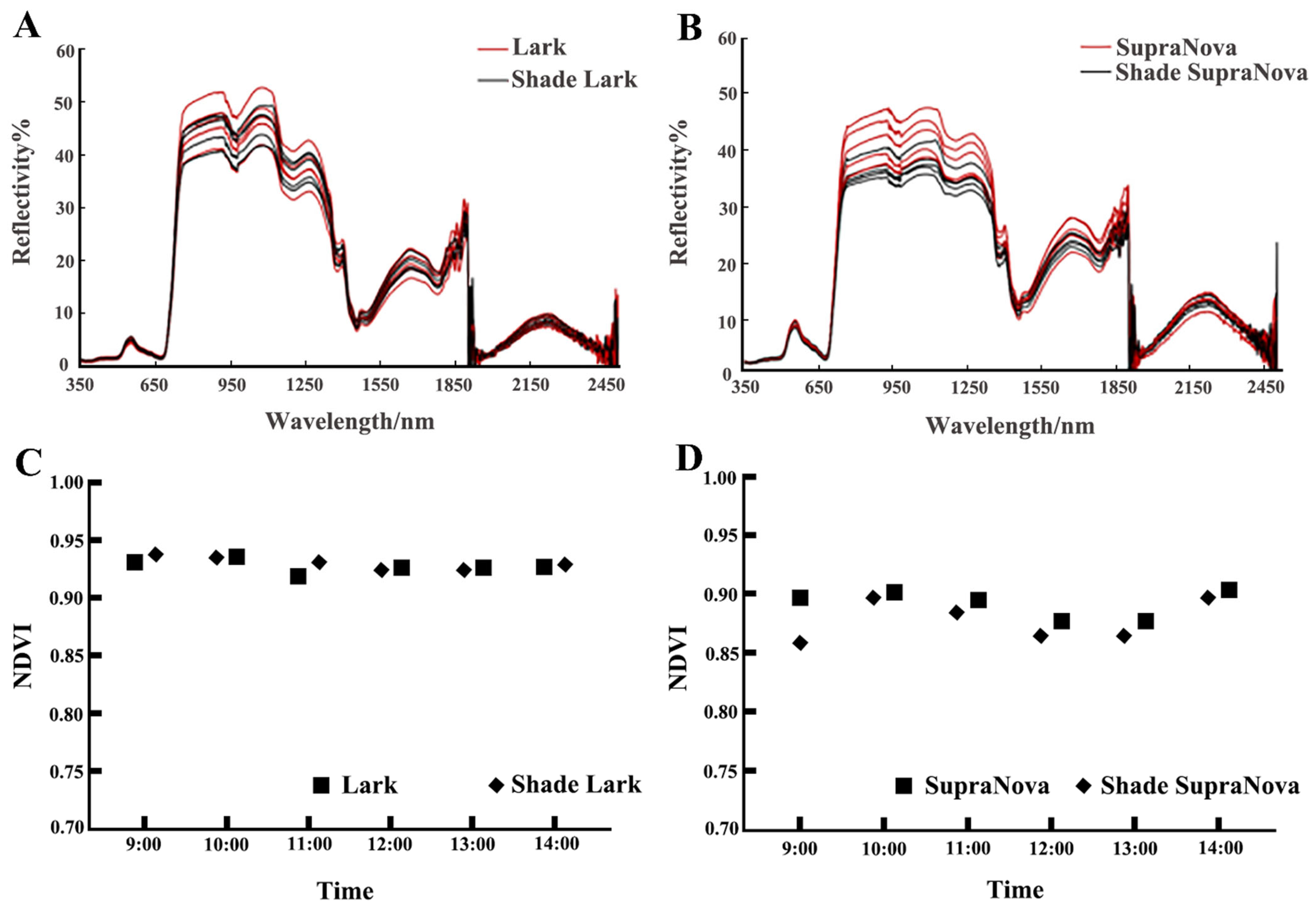
2.5. Spectral Determination
2.6. Physiological Enzyme Assay
2.7. Data Analysis
3. Results
3.1. Turf Quality (TQ) and Turf Color Intensity (TCI)
3.2. Root Morphological Analysis
3.3. Response of Chlorophyll a Fluorescence to Shade
3.4. Response of the Daily Spectral Variation to Shade
3.5. Physiological Responses to Shade
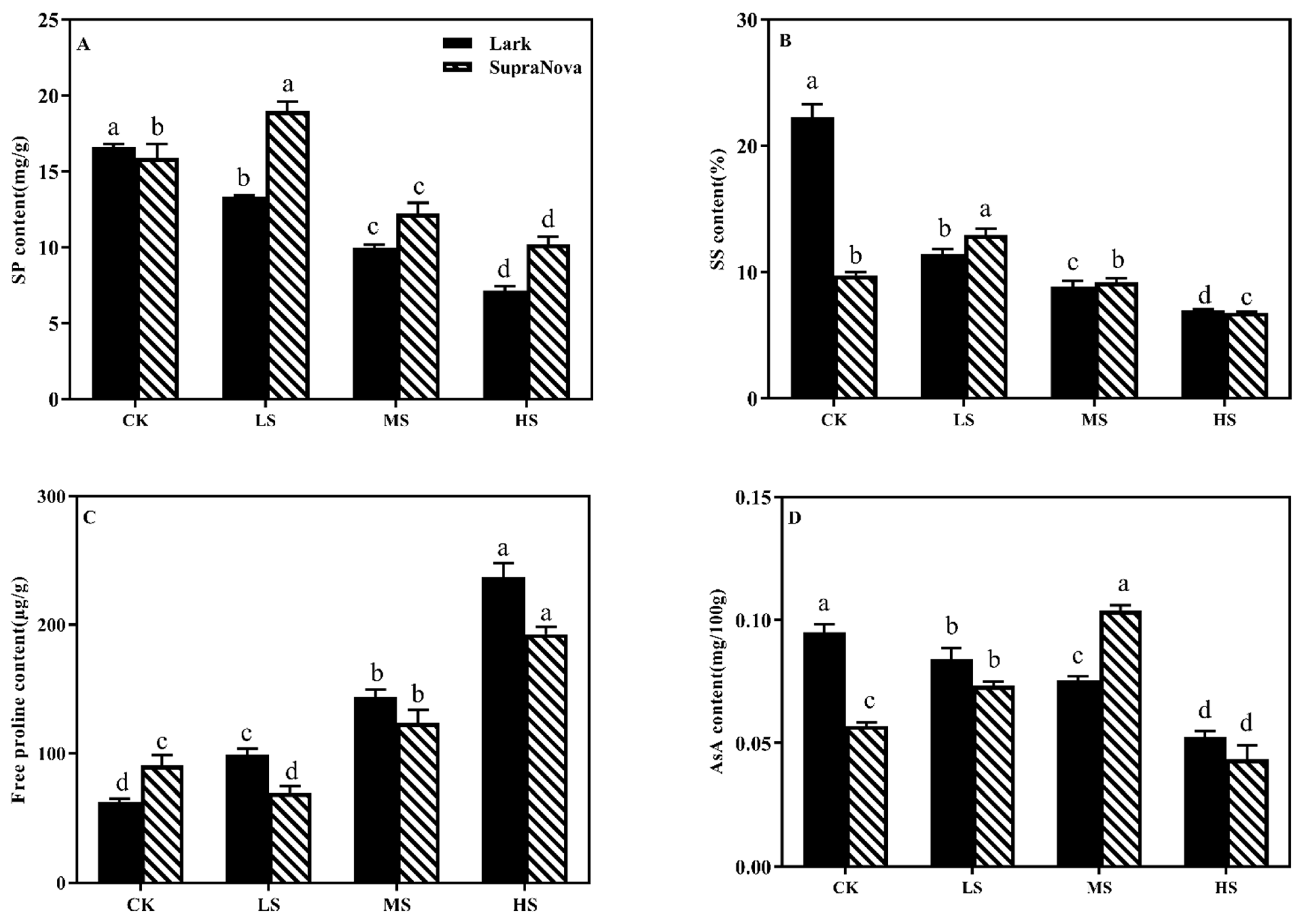
3.5.1. SP, SS, Pro, and AsA Content
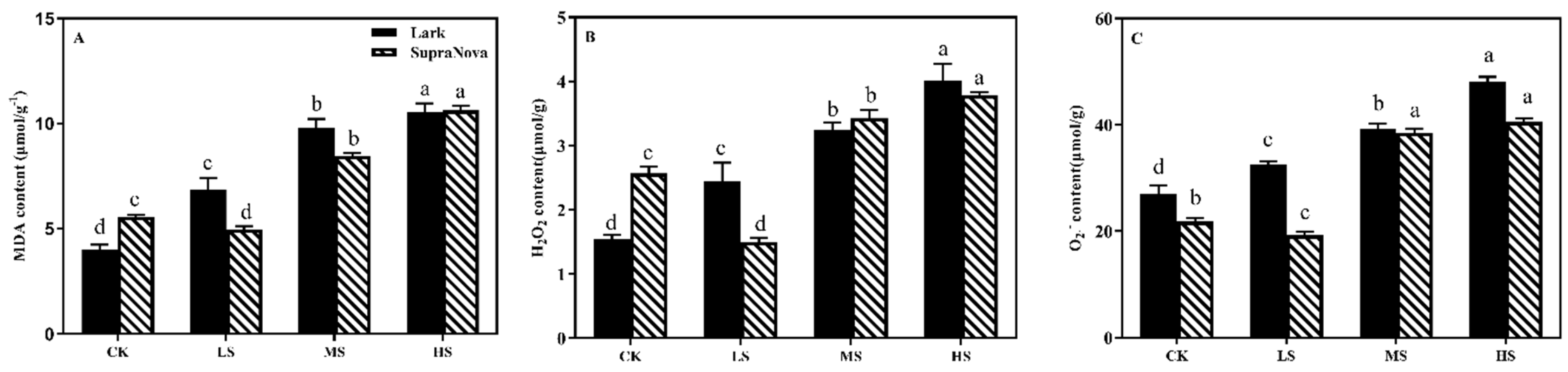
3.5.2. MDA, H2O2, and O2·− Contents
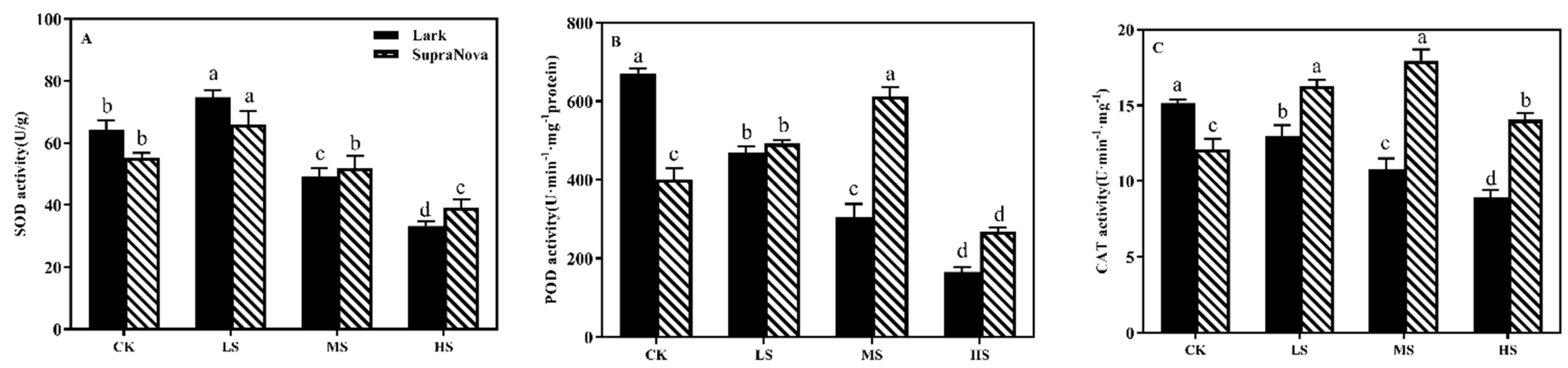
3.5.3. SOD, POD, and CAT Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rafiullah, A.; Tariq, M.; Khan, F.; Shah, A.; Fahad, S.; Wahid, F.; Ali, J.; Adnan, M.; Ahmad, M.; Irfan, M.; et al. Effect of micronutrients foliar supplementation on the production and eminence of plum (Prunus domestica L.). Qual. Assur. Saf. Crop. Foods 2020, 12, 32–40. [Google Scholar] [CrossRef]
- Thomason, W.; Battaglia, M. Early defoliation effects on corn plant stands and grain yield. Agron. J. 2020, 112, 5024–5032. [Google Scholar] [CrossRef]
- Sun, X.; Zheng, Q.; Xiong, L.; Xie, F.; Li, X.; Li, Y.; Zhang, L.; Saud, S.; Guo, Z.; Yan, Y.; et al. Nitrogen assimilation and gene regulation of two Kentucky bluegrass cultivars differing in response to nitrate supply. Sci. Hortic. 2021, 288, 110315. [Google Scholar] [CrossRef]
- Rezaei-Chiyaneh, E.; Mahdavikia, H.; Battaglia, M.L.; Thomason, W.E.; Caruso, G. Intercropping and fertilizer type impact seed productivity and secondary metabolites of dragon’s head and fenugreek. Sci. Hortic. 2021, 287, 110277. [Google Scholar] [CrossRef]
- Dias, A.N.; Siqueira-Silva, A.I.; Souza, J.P.; Kuki, K.N.; Pereira, E.G. Acclimation responses of macaw palm seedlings to contrasting light environments. Sci. Rep. 2018, 8, 15300. [Google Scholar] [CrossRef] [PubMed]
- Batool, A.; Cheng, Z.-G.; Akram, N.A.; Lv, G.-C.; Xiong, J.-L.; Zhu, Y.; Ashraf, M.; Xiong, Y.-C. Partial and full root-zone drought stresses account for differentiate root-sourced signal and yield formation in primitive wheat. Plant Methods 2019, 15, 75. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Wang, L.; Li, S.; Gao, Y.; Du, Y.; Zhao, L.; Liu, W.; Yang, W. Interactions Between Light Intensity and Phosphorus Nutrition Affect the P Uptake Capacity of Maize and Soybean Seedling in a Low Light Intensity Area. Front. Plant Sci. 2019, 10, 183. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Niu, W. Deficit Alternate Drip Irrigation Increased Root-Soil-Plant Interaction, Tomato Yield, and Quality. Int. J. Environ. Res. Public Health 2020, 17, 781. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.D.O.; Coelho, L.S.; Carvalho, G.R.; Botelho, C.E.; Torres, L.F.; Vilela, D.J.M.; Andrade, A.C.; Silva, V.A. Photochemical efficiency correlated with candidate gene expression promote coffee drought tolerance. Sci. Rep. 2021, 11, 7436. [Google Scholar] [CrossRef]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.; Baenziger, P.S.; Börner, A. Drought Stress Tolerance in Wheat and Barley: Advances in Physiology, Breeding and Genetics Research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [Green Version]
- Liangju, W.; Qi, L.; Wang, L.J. ALA Pretreatment Improves Waterlogging Tolerance of Fig Plants. PLoS ONE 2016, 11, e0147202. [Google Scholar] [CrossRef]
- Niu, Y.-F.; Wang, X.; Hu, D.-X.; Balamurugan, S.; Li, D.-W.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Molecular characterization of a glycerol-3-phosphate acyltransferase reveals key features essential for triacylglycerol production in Phaeodactylum tricornutum. Biotechnol. Biofuels 2016, 9, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, L.; Raza, M.A.; Li, Z.; Chen, Y.; Bin Khalid, M.H.; Du, J.; Liu, W.; Wu, X.; Song, C.; Yu, L.; et al. The Influence of Light Intensity and Leaf Movement on Photosynthesis Characteristics and Carbon Balance of Soybean. Front. Plant Sci. 2019, 9, 1952. [Google Scholar] [CrossRef] [PubMed]
- Soriano, G.; Del-Castillo-Alonso, M.Á.; Monforte, L.; Núñez-Olivera, E.; Martínez-Abaigar, J. Acclimation of Bryophytes to Sun Conditions, in Comparison to Shade Conditions, Is Influenced by Both Photosynthetic and Ultraviolet Radiations. Front. Plant Sci. 2019, 10, 998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Guo, X.; Wang, K.; Sun, Y.; Li, W.; Liu, Q.; Liu, Q. Nitrogen deposition does not alleviate the adverse effects of shade on Camellia japonica (Naidong) seedlings. PLoS ONE 2018, 13, e0201896. [Google Scholar] [CrossRef] [Green Version]
- Ajmi, A.; Vázquez, S.; Morales, F.; Chaari, A.; El-Jendoubi, H.; Abadía, A.; Larbi, A. Prolonged artificial shade affects morphological, anatomical, biochemical and ecophysiological behavior of young olive trees (cv. Arbosana). Sci. Hortic. 2018, 241, 275–284. [Google Scholar] [CrossRef]
- Xie, F.; Shi, Z.; Zhang, G.; Zhang, C.; Sun, X.; Yan, Y.; Zhao, W.; Guo, Z.; Zhang, L.; Fahad, S.; et al. Quantitative leaf anatomy and photophysiology systems of C3 and C4 turfgrasses in response to shading. Sci. Hortic. 2020, 274, 109674. [Google Scholar] [CrossRef]
- Kitao, M.; Kitaoka, S.; Harayama, H.; Tobita, H.; Agathokleous, E.; Utsugi, H. Canopy nitrogen distribution is optimized to prevent photoinhibition throughout the canopy during sun flecks. Sci. Rep. 2018, 8, 503. [Google Scholar] [CrossRef]
- Jiang, J.B.; Chen, Y.H.; Huang, W.J. Using hyperspectral derivative index to monitor winter wheat disease. Guang Pu Xue Yu Guang Pu Fen Xi Guang Pu 2007, 27, 2475–2479. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Aslam, M.T.; Alhammad, B.A.; Hassan, M.U.; Maqbool, R.; Chattha, M.U.; Khan, I.; Gitari, H.I.; Uslu, O.S.; Roy, R.; et al. Salinity Stress in Wheat: Effects, Mechanisms and Management Strategies. Phyton Int. J. Exp. Bot. 2021, 91, 667. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Abid, M.; Rizwan, M.; Shahid, M.R.; Alotaibi, M.; et al. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10, 16975. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, J.; Huang, Z.; Mi, L.; Xu, K.; Wu, J.; Fan, Y.; Ma, S.; Jiang, D. Effects of Low Temperature at Booting Stage on Sucrose Metabolism and Endogenous Hormone Contents in Winter Wheat Spikelet. Front. Plant Sci. 2019, 10, 498. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.M. Osmoregulation and Water Stress in Higher Plants. Annu. Rev. Plant Physiol. 1984, 35, 299–319. [Google Scholar] [CrossRef]
- Chen, P.; Liu, P.; Zhang, Q.; Bu, C.; Lu, C.; Srivastava, S.; Zhang, D.; Song, Y. Gene Coexpression Network Analysis Indicates that Hub Genes Related to Photosynthesis and Starch Synthesis Modulate Salt Stress Tolerance in Ulmus pumila. Int. J. Mol. Sci. 2021, 22, 4410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, L.; Chen, B.; Qin, Z.; Xiaoning, Z.; Zhang, Y.; Yao, R.; Liu, H.; Yang, H. Progress in Understanding the Physiological and Molecular Responses of Populus to Salt Stress. Int. J. Mol. Sci. 2019, 20, 1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasilakoglou, I.; Dhima, K.; Giannakoula, A.; Dordas, C.; Skiada, V.; Papadopoulou, K. Carbon Assimilation, Isotope Discrimination, Proline and Lipid Peroxidation Contribution to Barley (Hordeum vulgare) Salinity Tolerance. Plants 2021, 10, 299. [Google Scholar] [CrossRef]
- Liu, C.; Hu, T.X.; Liu, W.T.; Zhang, L.; Shuai, W.; Tang, T.Y.; Li, Q. Physiological bases of herbages shade-tolerance in Eucalyptus grandis and herbage inter-cultivated system. Chin. J. Appl. Ecol. 2008, 19, 1911–1916. [Google Scholar]
- De Sousa, A.; AbdElgawad, H.; Han, A.; Teixeira, J.; Matos, M.; Fidalgo, F. Oxidative Metabolism of Rye (Secale cereale L.) after Short Term Exposure to Aluminum: Uncovering the Glutathione–Ascorbate Redox Network. Front. Plant Sci. 2016, 7, 685. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Sano, S.; Takemoto, T.; Ogihara, A.; Suzuki, K.; Masumura, T.; Satoh, S.; Takano, K.; Mimura, Y.; Morita, S. Stress Responses of Shade-Treated Tea Leaves to High Light Exposure after Removal of Shading. Plants 2020, 9, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bali, S.; Kaur, P.; Jamwal, V.L.; Gandhi, S.G.; Sharma, A.; Ohri, P.; Bhardwaj, R.; Ali, M.A.; Ahmad, P. Seed Priming with Jasmonic Acid Counteracts Root Knot Nematode Infection in Tomato by Modulating the Activity and Expression of Antioxidative Enzymes. Biomolecules 2020, 10, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turgeon, A.J. Turfgrass Management, 8th ed.; Pearson Prentice Hall Press: Hoboken, NJ, USA, 2008. [Google Scholar]
- Chen, Y.; Pettersen, T.; Kvalbein, A.; Aamlid, T.S. Playing quality, growth rate, thatch accumulation and tolerance to moss and annual bluegrass invasion as influenced by irrigation strategies on red fescue putting greens. J. Agron. Crop Sci. 2018, 204, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Pei, J.; Zhao, Y.; Wang, S. Physiological Changes of Bamboo (Fargesia yunnanensis) Shoots During Storage and the Related Cold Storage Mechanisms. Front. Plant Sci. 2021, 12, 731977. [Google Scholar] [CrossRef]
- Du, J.; Shen, T.; Xiong, Q.; Zhu, C.; Peng, X.; He, X.; Fu, J.; Ouyang, L.; Bian, J.; Hu, L.; et al. Combined proteomics, metabolomics and physiological analyses of rice growth and grain yield with heavy nitrogen application before and after drought. BMC Plant Biol. 2020, 20, 556. [Google Scholar] [CrossRef]
- Guo, Y.; Gao, Z.; Li, L.; Wang, Y.; Zhao, H.; Hu, M.; Li, M.; Zhang, Z. Effect of controlled atmospheres with varying O2/CO2 levels on the postharvest senescence and quality of broccoli (Brassica oleracea L. var. italica) florets. Eur. Food Res. Technol. 2013, 237, 943–950. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alamri, S.A.; Alam, P.; Ashraf, M.; Ahmad, P. Potential of exogenously sourced kinetin in protecting Solanum lycopersicum from NaCl-induced oxidative stress through up-regulation of the antioxidant system, ascorbate-glutathione cycle and glyoxalase system. PLoS ONE 2018, 13, e0202175. [Google Scholar] [CrossRef]
- Wang, W.-R.; Liang, J.-H.; Wang, G.-F.; Sun, M.-X.; Peng, F.-T.; Xiao, Y.-S. Overexpression of PpSnRK1α in tomato enhanced salt tolerance by regulating ABA signaling pathway and reactive oxygen metabolism. BMC Plant Biol. 2020, 20, 128. [Google Scholar] [CrossRef]
- Giannopolotis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Tian, M.; Gu, Q.; Zhu, M. The involvement of hydrogen peroxide and antioxidant enzymes in the process of shoot organogenesis of strawberry callus. Plant Sci. 2003, 165, 701–707. [Google Scholar] [CrossRef]
- Ye, S.; Shao, Q.; Xu, M.; Li, S.; Wu, M.; Tan, X.; Su, L. Effects of Light Quality on Morphology, Enzyme Activities, and Bioactive Compound Contents in Anoectochilus roxburghii. Front. Plant Sci. 2017, 8, 857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, A.; Bordoloi, S.; Ganesan, S.P.; Sekharan, S.; Sahoo, L. A relook into plant wilting: Observational evidence based on unsaturated soil–plant-photosynthesis interaction. Sci. Rep. 2020, 10, 22064. [Google Scholar] [CrossRef] [PubMed]
- Villanova, I.R.; Martínez-García, J.F. Plant Responses to Vegetation Proximity: A Whole Life Avoiding Shade. Front. Plant Sci. 2016, 7, 236. [Google Scholar] [CrossRef] [Green Version]
- Hochholdinger, F.; Marcon, C.; Baldauf, J.A.; Yu, P.; Frey, F.P. Proteomics of Maize Root Development. Front. Plant Sci. 2018, 9, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Wu, X.; Sun, M.; Peng, F. Hydrogen Sulfide Alleviates Waterlogging-Induced Damage in Peach Seedlings via Enhancing Antioxidative System and Inhibiting Ethylene Synthesis. Front. Plant Sci. 2020, 11, 696. [Google Scholar] [CrossRef]
- Elkins, C.; Van Iersel, M.W. Longer Photoperiods with the Same Daily Light Integral Increase Daily Electron Transport through Photosystem II in Lettuce. Plants 2020, 9, 1172. [Google Scholar] [CrossRef]
- Mo, Y.; Wang, Y.; Yang, R.; Zheng, J.; Liu, C.; Li, H.; Ma, J.; Zhang, Y.; Wei, C.; Zhang, X. Regulation of Plant Growth, Photosynthesis, Antioxidation and Osmosis by an Arbuscular Mycorrhizal Fungus in Watermelon Seedlings under Well-Watered and Drought Conditions. Front. Plant Sci. 2016, 7, 644. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Wei, X.; Wan, D.; He, W.; Wang, X.; Xiong, Y. Effect of Molybdenum on Plant Physiology and Cadmium Uptake and Translocation in Rape (Brassica napus L.) under Different Levels of Cadmium Stress. Int. J. Environ. Res. Public Health 2020, 17, 2355. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Chen, H.; Guo, B.; Liu, C. Understanding the molecular mechanisms underlying the effects of light intensity on flavonoid production by RNA-seq analysis in Epimedium pseudowushanense B.L.Guo. PLoS ONE 2017, 12, e0182348. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Lai, L.; Zhou, J.; Li, Q.; Yi, S.; Sun, Q.; Zheng, Y. Changes in levels of enzymes and osmotic adjustment compounds in key species and their relevance to vegetation succession in abandoned croplands of a semiarid sandy region. Ecol. Evol. 2020, 10, 2269–2280. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Numan, M.; Khan, A.L.; Lee, I.-J.; Imran, M.; Asaf, S.; Al-Harrasi, A. Melatonin: Awakening the Defense Mechanisms during Plant Oxidative Stress. Plants 2020, 9, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Song, L.; Yu, W.; Hu, Y.; Liu, Y.; Wu, J.; Ying, Y. Growth, physiological, and biochemical responses of Camptotheca acuminata seedlings to different light environments. Front. Plant Sci. 2015, 6, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Schellhorn, H.E. Can ageing-related degenerative diseases be ameliorated through administration of vitamin C at pharmacological levels? Med. Hypotheses 2007, 68, 1315–1317. [Google Scholar] [CrossRef] [PubMed]
- Munir, S.; Mumtaz, M.A.; Ahiakpa, J.K.; Liu, G.; Chen, W.; Zhou, G.; Zheng, W.; Ye, Z.; Zhang, Y. Genome-wide analysis of Myo-inositol oxygenase gene family in tomato reveals their involvement in ascorbic acid accumulation. BMC Genom. 2020, 21, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Xie, J.; Yu, J. Physiological and transcriptomic responses of Lanzhou Lily (Lilium davidii, var. unicolor) to cold stress. PLoS ONE 2020, 15, e0227921. [Google Scholar] [CrossRef] [PubMed]
- Bao, G.; Tang, W.; An, Q.; Liu, Y.; Tian, J.; Zhao, N.; Zhu, S. Physiological effects of the combined stresses of freezing-thawing, acid precipitation and deicing salt on alfalfa seedlings. BMC Plant Biol. 2020, 20, 204. [Google Scholar] [CrossRef]
- Tyburski, J.; Dunajska, K.; Tretyn, A. Reactive oxygen species localization in roots of Arabidopsis thaliana seedlings grown under phosphate deficiency. Plant Growth Regul. 2010, 59, 27–36. [Google Scholar] [CrossRef]
- Li, Y.; Qin, T.; Wei, C.; Sun, J.; Dong, T.; Zhou, R.; Chen, Q.; Wang, Q. Using Transcriptome Analysis to Screen for Key Genes and Pathways Related to Cytoplasmic Male Sterility in Cotton (Gossypium hirsutum L.). Int. J. Mol. Sci. 2019, 20, 5120. [Google Scholar] [CrossRef] [Green Version]
- El-Beltagi, H.S.; Mohamed, H.I.; Sofy, M.R. Role of Ascorbic acid, Glutathione and Proline Applied as Singly or in Sequence Combination in Improving Chickpea Plant through Physiological Change and Antioxidant Defense under Different Levels of Irrigation Intervals. Molecules 2020, 25, 1702. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Shading Level (%) | 15D | 30D | 45D | |||
|---|---|---|---|---|---|---|---|
| TCI | TQ | TCI | TQ | TCI | TQ | ||
| SupraNova | CK | 7.1 a | 7.6 a | 7.3 a | 7.5 a | 7.1 a | 7.6 a |
| LS | 8.1 a | 8.6 a | 7.8 a | 7.8 a | 7.5 a | 7.7 a | |
| MS | 6.0 b | 7.2 a | 3.2 c | 3.5 c | 2.1 d | 2.8 d | |
| HS | 4.8 b | 4.4 b | 2.2 d | 3.0 c | 1.3 d | 1.7 d | |
| Lark | CK | 8.3 a | 8.0 a | 9.0 a | 8.2 a | 9.0 a | 8.2 a |
| LS | 6.2 b | 7.2 a | 4.7 b | 3.5 c | 3.5 c | 4.2 b | |
| MS | 4.6 b | 4.7 b | 2.7 d | 3.2 c | 1.5 d | 2.3 d | |
| HS | 2.7 d | 3.1 c | 2.5 d | 1.7 d | 1.3 d | 1.7 d | |
| Species | Shading Level (%) | Root Length (cm) | Root Surface Area (cm2) |
|---|---|---|---|
| SupraNova | CK | 401.8 ± 5.2 b | 52.8 ± 3.4 b |
| LS | 675.0 ± 8.7 a | 70.7 ± 2.4 a | |
| MS | 222.5 ± 3.2 c | 41.8 ± 4.2 c | |
| HS | 103.5 ± 7.4 d | 24.7 ± 1.2 d | |
| Lark | CK | 1162.4 ± 17.5 a | 111.2 ± 7.3 a |
| LS | 512.9 ± 6.8 b | 52.5 ± 4.8 b | |
| MS | 354.8 ± 3.4 c | 56.2 ± 4.5 b | |
| HS | 309.3 ± 4.3 d | 53.6 ± 1.4 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Xiong, L.; Sun, X.; Shah, S.; Guo, Z.; Zhao, X.; Liu, L.; Cheng, L.; Tian, Z.; Xie, F.; et al. Morphophysiological Responses of Two Cool-Season Turfgrasses with Different Shade Tolerances. Agronomy 2022, 12, 959. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12040959
Dong L, Xiong L, Sun X, Shah S, Guo Z, Zhao X, Liu L, Cheng L, Tian Z, Xie F, et al. Morphophysiological Responses of Two Cool-Season Turfgrasses with Different Shade Tolerances. Agronomy. 2022; 12(4):959. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12040959
Chicago/Turabian StyleDong, Lili, Liangbing Xiong, Xiaoyang Sun, Saud Shah, Zhixin Guo, Xueying Zhao, Lizhi Liu, Lintao Cheng, Zhi Tian, Fuchun Xie, and et al. 2022. "Morphophysiological Responses of Two Cool-Season Turfgrasses with Different Shade Tolerances" Agronomy 12, no. 4: 959. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12040959