
Response of Phytic Acid to Nitrogen Application and Its Relation to Protein Content in Rice Grain

 , , , ,
, , , ,
Abstract
:
1. Introduction
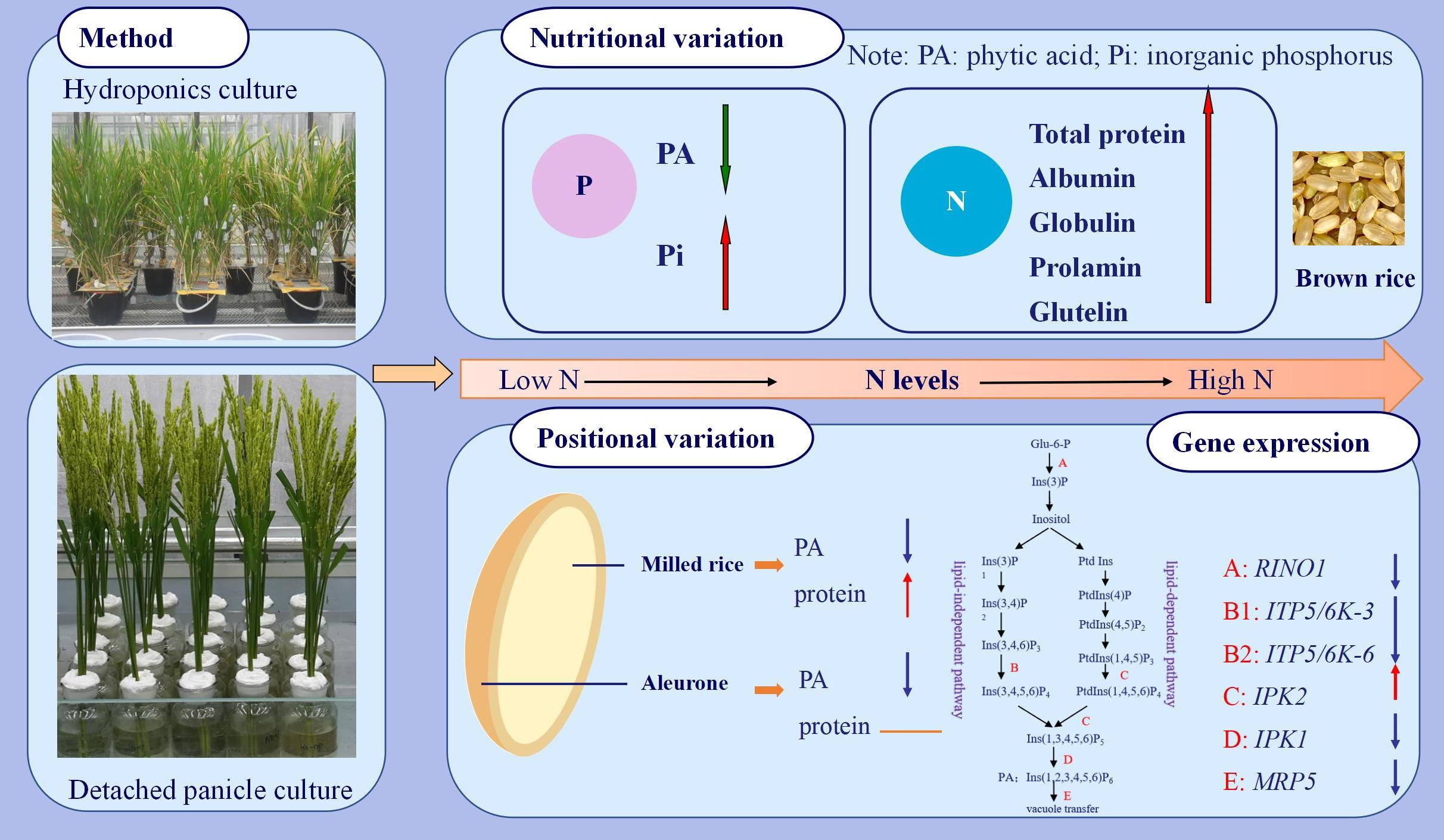
2. Materials and Methods
2.1. Plant Materials
2.2. Hydroponic Culture
2.3. Detached Panicle Culture System
2.4. Chemical Analyses
2.5. Quantitative RT-PCR
2.6. Statistical Analysis
3. Results
3.1. Alterations in Grain Weight and Protein Concentration under Different N Levels
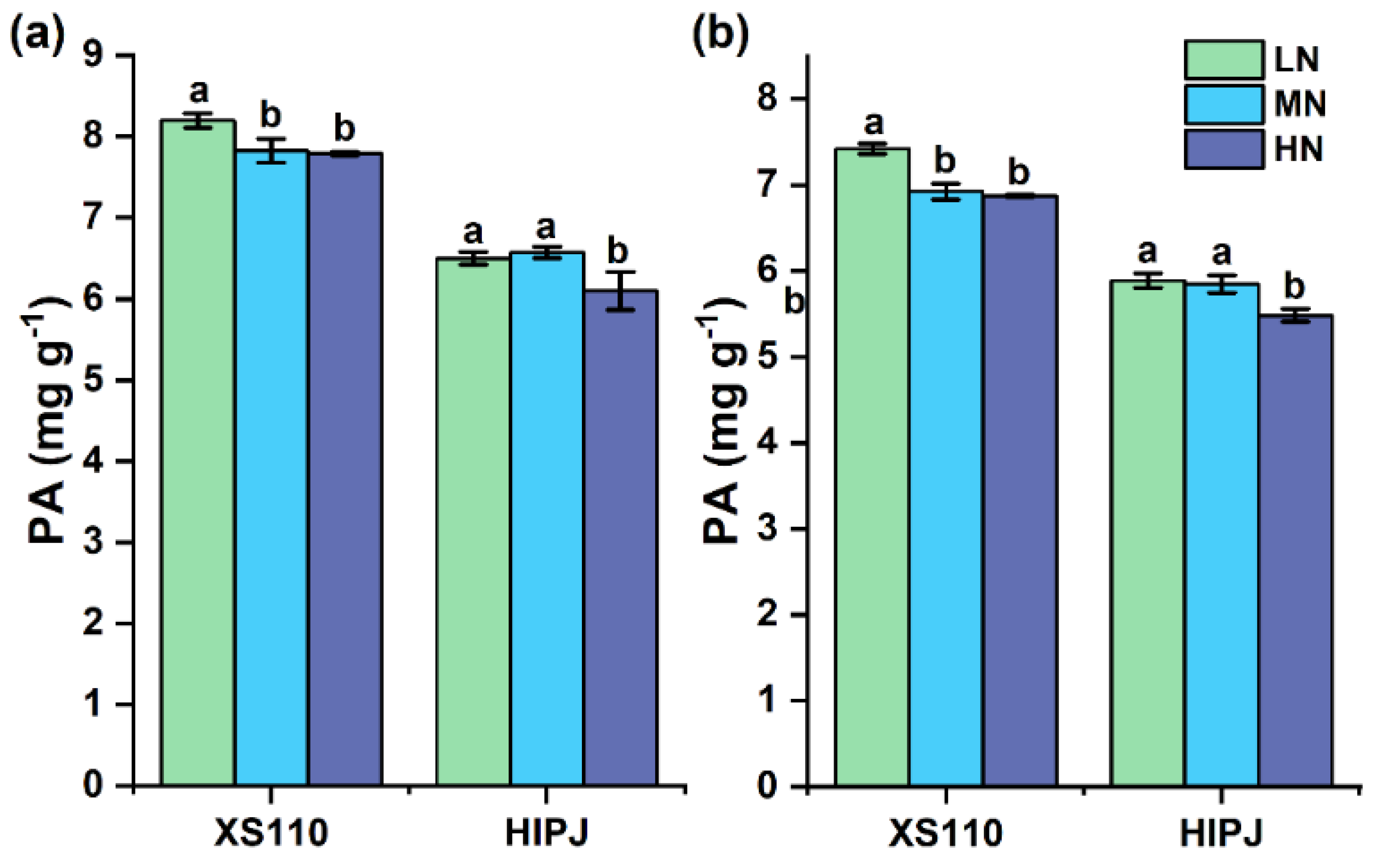
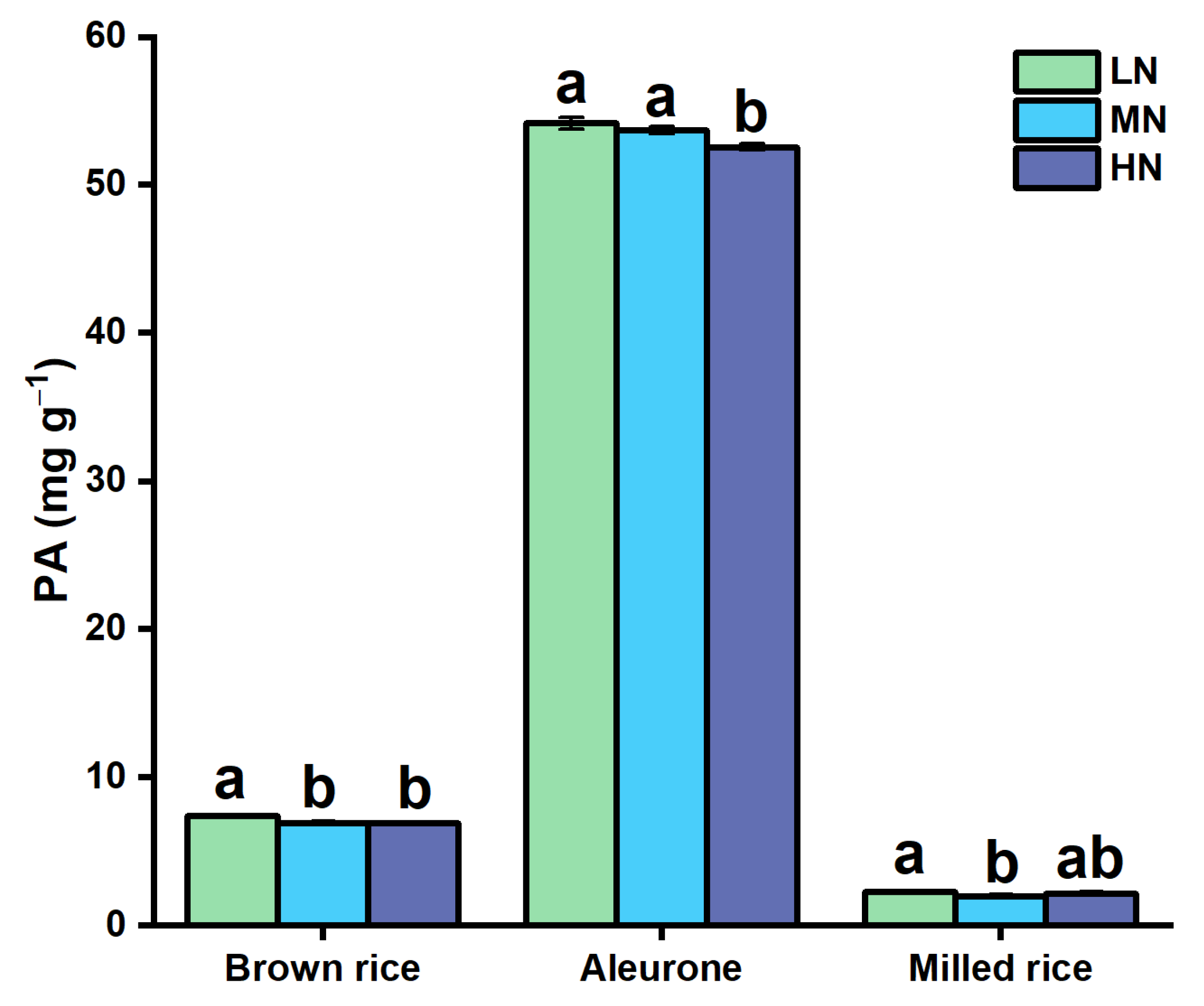
3.2. Alterations in Grain Phytic Acid with N Application
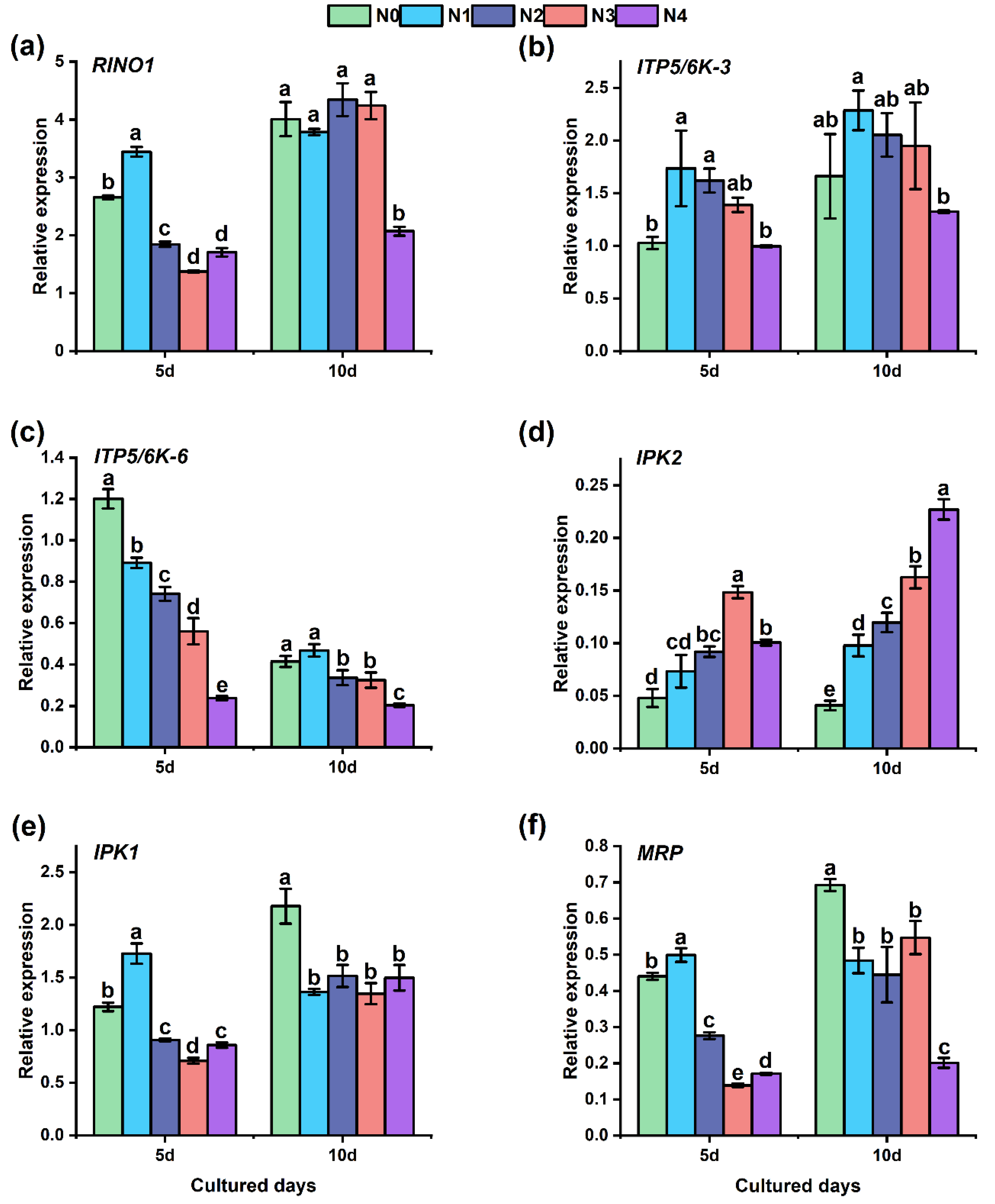
3.3. Transcript Profiling of PA under N Application
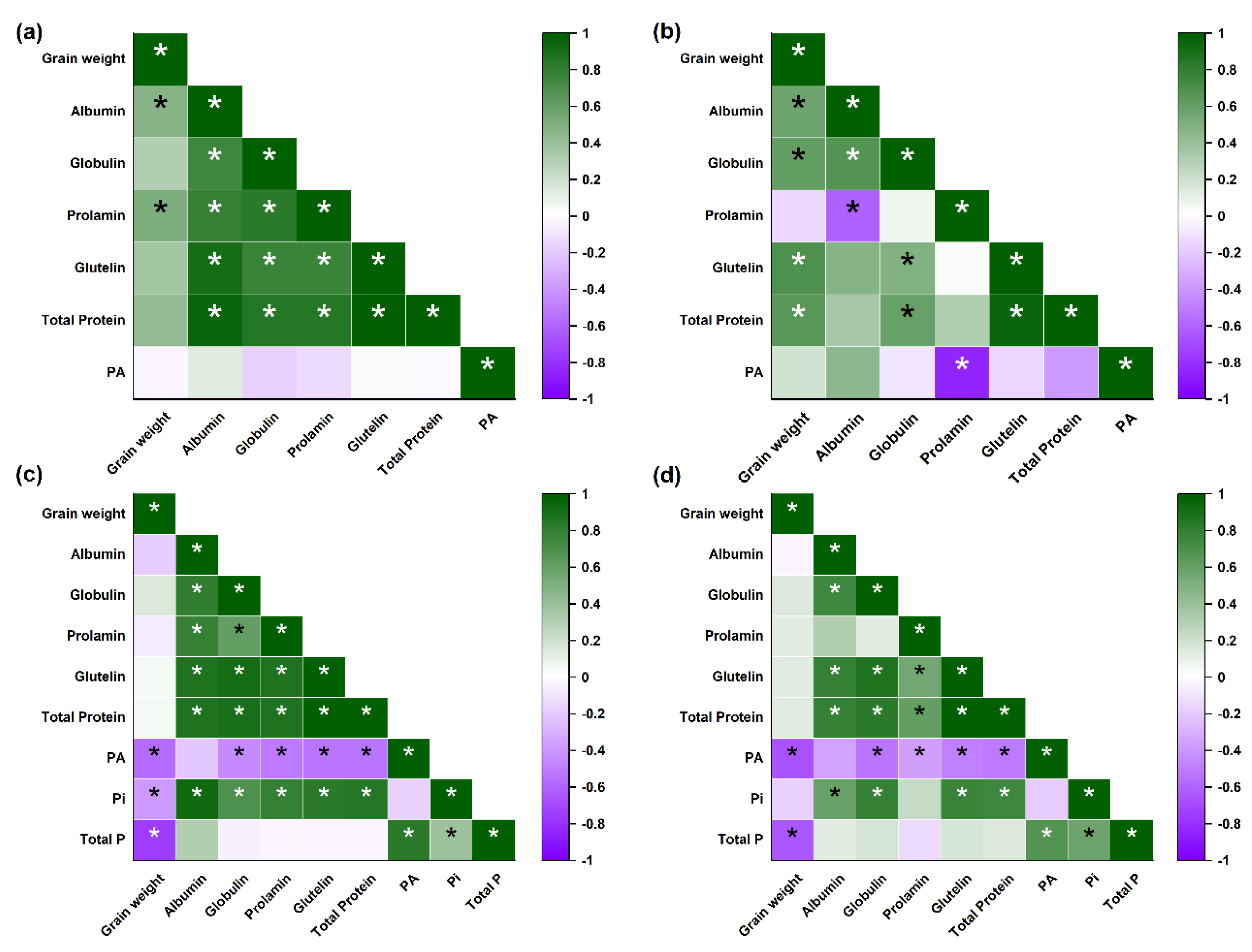
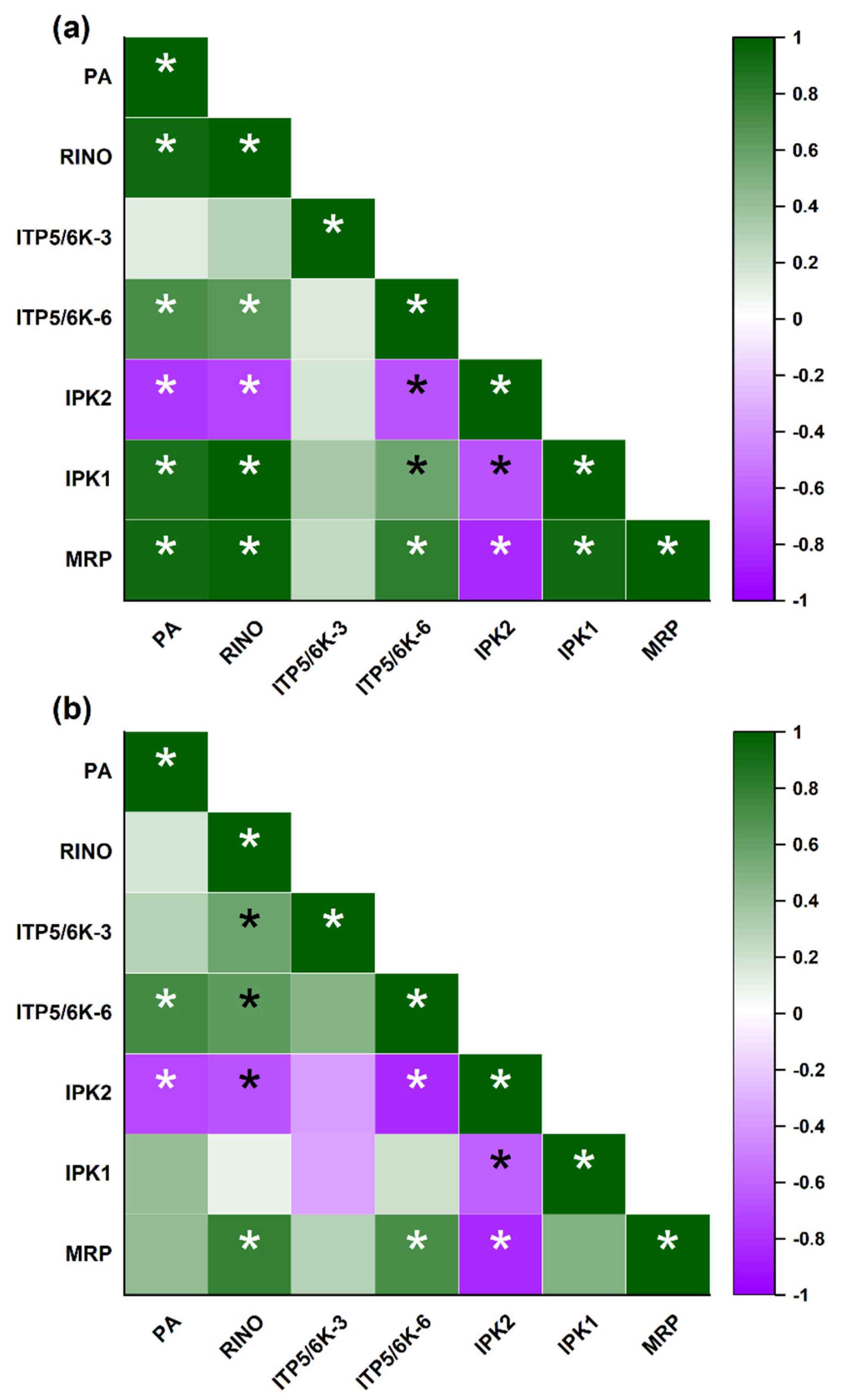
3.4. Correlation Analysis
4. Discussion
4.1. Response of Grain Protein and Its Fractions to N Application
4.2. Response of Grain PA to N Application
4.3. Transcript Profiling of PA under N Application
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schneider, P.; Asch, F. Rice production and food security in Asian Mega deltas-A review on characteristics, vulnerabilities and agricultural adaptation options to cope with climate change. J. Agron. Crop Sci. 2020, 206, 491–503. [Google Scholar] [CrossRef]
- Panigrahi, R.; Kariali, E.; Panda, B.B.; Lafarge, T.; Mohapatra, P.K. Controlling the trade-off between spikelet number and grain filling: The hierarchy of starch synthesis in spikelets of rice panicle in relation to hormone dynamics (vol 46, pg 507, 2019). Funct. Plant Biol. 2019, 46, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Witcombe, J.R.; Gyawali, S.; Subedi, M.; Virk, D.S.; Joshi, K.D. Plant breeding can be made more efficient by having fewer, better crosses. BMC Plant Biol. 2013, 13, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiaz, S.; Ahmad, S.; Noor, M.A.; Wang, X.; Younas, A.; Riaz, A.; Riaz, A.; Ali, F. Applications of the CRISPR/Cas9 System for Rice Grain Quality Improvement: Perspectives and Opportunities. Int. J. Mol. Sci. 2019, 20, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abeysingha, N.S.; Singh, M.; Islam, A.; Sehgal, V.K. Climate change impacts on irrigated rice and wheat production in Gomti River basin of India: A case study. Springerplus 2016, 5, 1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Oort, P.A.J.; Zwart, S.J. Impacts of climate change on rice production in Africa and causes of simulated yield changes. Glob. Chang. Biol. 2018, 24, 1029–1045. [Google Scholar] [CrossRef] [Green Version]
- Perchlik, M.; Tegeder, M. Improving Plant Nitrogen Use Efficiency through Alteration of Amino Acid Transport Processes. Plant Physiol. 2017, 175, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Linquist, B.A.; Liu, L.J.; van Kessel, C.; van Groenigen, K.J. Enhanced efficiency nitrogen fertilizers for rice systems: Meta-analysis of yield and nitrogen uptake. Field Crops Res. 2013, 154, 246–254. [Google Scholar] [CrossRef]
- Mur, L.A.J.; Simpson, C.; Kumari, A.; Gupta, A.K.; Gupta, K.J. Moving nitrogen to the centre of plant defence against pathogens. Ann. Bot. 2017, 119, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Verma, A.; Ansari, M.W.; Shukla, A. Physiological response of rice (Oryza sativa L.) genotypes to elevated nitrogen applied under field conditions. Plant Signal Behav. 2014, 9, e29015. [Google Scholar] [CrossRef] [Green Version]
- Xue, L.H.; Yang, L.Z. Recommendations for nitrogen fertiliser topdressing rates in rice using canopy reflectance spectra. Biosyst. Eng. 2008, 100, 524–534. [Google Scholar] [CrossRef]
- Ning, H.F.; Liu, Z.G.; Wang, Q.S.; Lin, Z.M.; Chen, S.J.; Li, G.H.; Wang, S.H.; Ding, Y.F. Effect of nitrogen fertilizer application on grain phytic acid and protein concentrations in japonica rice and its variations with genotypes. J. Cereal Sci. 2009, 50, 49–55. [Google Scholar] [CrossRef]
- Gu, J.F.; Chen, J.; Chen, L.; Wang, Z.Q.; Zhang, H.; Yang, J.C. Grain quality changes and responses to nitrogen fertilizer of japonica rice cultivars released in the Yangtze River Basin from the 1950s to 2000s. Crop J. 2015, 3, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.C.; Huang, Y.C.; Jia, B.Y.; Wang, Y.; Wang, Y.; Xu, Q.; Li, R.F.; Wang, S.; Dou, F.G. Effects of Cultivar, Nitrogen Rate, and Planting Density on Rice-Grain Quality. Agronomy 2018, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.R.; Timsina, Y.N.; Lind, O.C.; Cagno, S.; Janssens, K. Zinc and Iron Concentration as Affected by Nitrogen Fertilization and Their Localization in Wheat Grain. Front. Plant Sci. 2018, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Wood, R.M.; Dunn, B.W.; Balindong, J.L.; Waters, D.L.E.; Blanchard, C.L.; Mawson, A.J.; Oli, P. Effect of agronomic management on rice grain quality Part II: Nitrogen rate and timing. Cereal Chem. 2021, 98, 234–248. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Hirose, S.; Yasuda, H.; Takaiwa, F. Reducing rice seed storage protein accumulation leads to changes in nutrient quality and storage organelle formation. Plant Physiol. 2010, 154, 1842–1854. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.X.; Li, H.; Liang, M.C.; Yang, L. Glutelin and prolamin, different components of rice protein, exert differently in vitro antioxidant activities. J. Cereal Sci. 2016, 72, 108–116. [Google Scholar] [CrossRef]
- Perera, I.; Seneweera, S.; Hirotsu, N. Manipulating the Phytic Acid Content of Rice Grain Toward Improving Micronutrient Bioavailability. Rice 2018, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Ali, N.; Paul, S.; Gayen, D.; Sarkar, S.N.; Datta, S.K.; Datta, K. RNAi mediated down regulation of myo-inositol-3-phosphate synthase to generate low phytate rice. Rice 2013, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Sparvoli, F.; Cominelli, E. Seed Biofortification and Phytic Acid Reduction: A Conflict of Interest for the Plant? Plants 2015, 4, 728–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.F.; Liang, H.M.; Yang, S.Y.; Boch, A.; Clemens, S.; Chen, C.C.; Wu, J.F.; Huang, J.L.; Yeh, K.C. Arabidopsis IRT3 is a zinc-regulated and plasma membrane localized zinc/iron transporter. N. Phytol. 2009, 182, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Morgounov, A.; Gomez-Becerra, H.F.; Abugalieva, A.; Dzhunusova, M.; Yessimbekova, M.; Muminjanov, H.; Zelenskiy, Y.; Ozturk, L.; Cakmak, I. Iron and zinc grain density in common wheat grown in Central Asia. Euphytica 2007, 155, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, D.; Liu, Y.; Chen, X.; Zou, C. Overuse of Phosphorus Fertilizer Reduces the Grain and Flour Protein Contents and Zinc Bioavailability of Winter Wheat (Triticum aestivum L.). J. Agric. Food Chem 2017, 65, 1473–1482. [Google Scholar] [CrossRef] [PubMed]
- Simic, M.; Dragicevic, V.; Drinic, S.M.; Vukadinovic, J.; Kresovic, B.; Tabakovic, M.; Brankov, M. The Contribution of Soil Tillage and Nitrogen Rate to the Quality of Maize Grain. Agronomy 2020, 10, 976. [Google Scholar] [CrossRef]
- Kaplan, M.; Karaman, K.; Kardes, Y.M.; Kale, H. Phytic acid content and starch properties of maize (Zea mays L.): Effects of irrigation process and nitrogen fertilizer. Food Chem. 2019, 283, 375–380. [Google Scholar] [CrossRef]
- Wang, S.X.; Li, M.; Tian, X.H.; Li, J.; Li, H.Y.; Ni, Y.J.; Zhao, J.H.; Chen, Y.L.; Guo, C.H.; Zhao, A.Q. Foliar Zinc, Nitrogen, and Phosphorus Application Effect on Micronutrient Concentrations in Winter Wheat. Agron. J. 2015, 107, 61–70. [Google Scholar] [CrossRef]
- Iwai, T.; Takahashi, M.; Oda, K.; Terada, Y.; Yoshida, K.T. Dynamic changes in the distribution of minerals in relation to phytic acid accumulation during rice seed development. Plant Physiol. 2012, 160, 2007–2014. [Google Scholar] [CrossRef] [Green Version]
- Paltridge, N.G.; Palmer, L.J.; Milham, P.J.; Guild, G.E.; Stangoulis, J.C.R. Energy-dispersive X-ray fluorescence analysis of zinc and iron concentration in rice and pearl millet grain. Plant Soil 2012, 361, 251–260. [Google Scholar] [CrossRef]
- Liu, Q.L.; Xu, X.H.; Ren, X.L.; Fu, H.W.; Wu, D.X.; Shu, Q.Y. Generation and characterization of low phytic acid germplasm in rice (Oryza sativa L.). Theor. Appl. Genet. 2007, 114, 803–814. [Google Scholar] [CrossRef]
- Su, D.; Zhou, L.; Zhao, Q.; Pan, G.; Cheng, F. Different Phosphorus Supplies Altered the Accumulations and Quantitative Distributions of Phytic Acid, Zinc, and Iron in Rice (Oryza sativa L.) Grains. J. Agric. Food Chem. 2018, 66, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.M.; Wu, J.G.; Li, G.; Zhang, D.P.; Yang, Z.W.; Shi, C.H. Distribution of phytic acid and mineral elements in three indica rice (Oryza sativa L.) cultivars. J. Cereal Sci. 2011, 54, 116–121. [Google Scholar] [CrossRef]
- Liu, Z.H.; Cheng, F.M.; Cheng, W.D.; Zhang, G.P. Positional variations in phytic acid and protein content within a panicle of japonica rice. J. Cereal Sci. 2005, 41, 297–303. [Google Scholar] [CrossRef]
- Jiang, M.; Liu, Y.; Liu, Y.; Tan, Y.; Huang, J.; Shu, Q. Mutation of Inositol 1,3,4-trisphosphate 5/6-kinase6 Impairs Plant Growth and Phytic Acid Synthesis in Rice. Plants 2019, 8, 114. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wu, L.H.; Wang, M.Y. Iron and zinc biofortification in polished rice and accumulation in rice plant (Oryza sativa L.) as affected by nitrogen fertilization. Acta Agric. Scand. Sect. B Soil Plant Sci. 2008, 58, 267–272. [Google Scholar] [CrossRef]
- Liu, P.X.; Ma, X.F.; Wan, H.S.; Zheng, J.M.; Luo, J.T.; Hu, Y.B.; Pu, Z.J. Effects of differential nitrogen application on wheat grain proteome. J. Cereal Sci. 2021, 102, 103367. [Google Scholar] [CrossRef]
- Liu, K.; Deng, J.; Lu, J.; Wang, X.; Lu, B.; Tian, X.; Zhang, Y. High Nitrogen Levels Alleviate Yield Loss of Super Hybrid Rice Caused by High Temperatures During the Flowering Stage. Front. Plant Sci. 2019, 10, 357. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Lal, M.K.; Kar, S.S.; Nayak, L.; Ngangkham, U.; Samantaray, S.; Sharma, G. Bioavailability of iron and zinc as affected by phytic acid content in rice grain. J. Food Biochem. 2017, 41, e12413. [Google Scholar] [CrossRef]
- Wang, Z.H.; Li, S.X.; Malhi, S. Effects of fertilization and other agronomic measures on nutritional quality of crops. J. Sci. Food Agric. 2008, 88, 7–23. [Google Scholar] [CrossRef]
- Shi, R.L.; Zhang, Y.Q.; Chen, X.P.; Sun, Q.P.; Zhang, F.S.; Romheld, V.; Zou, C.Q. Influence of long-term nitrogen fertilization on micronutrient density in grain of winter wheat (Triticum aestivum L.). J. Cereal Sci. 2010, 51, 165–170. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taulemesse, F.; Le Gouis, J.; Gouache, D.; Gibon, Y.; Allard, V. Post-Flowering Nitrate Uptake in Wheat Is Controlled by N Status at Flowering, with a Putative Major Role of Root Nitrate Transporter NRT2.1. PLoS ONE 2015, 10, e0120291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.F.; Sun, M.; Xing, Y.Z.; Hua, J.P.; Sun, X.L.; Zhang, Q.F.; Corke, H. Mapping quantitative trait loci for milling quality, protein content and color characteristics of rice using a recombinant inbred line population derived from an elite rice hybrid. Theor. Appl. Genet. 2001, 103, 1037–1045. [Google Scholar] [CrossRef]
- Liang, W.X.; Zhang, Z.C.; Wen, X.X.; Liao, Y.C.; Liu, Y. Effect of non-structural carbohydrate accumulation in the stem pre-anthesis on grain filling of wheat inferior grain. Field Crops Res. 2017, 211, 66–76. [Google Scholar] [CrossRef]
- Furukawa, S.; Tanaka, K.; Masumura, T.; Ogihara, Y.; Kiyokawa, Y.; Wakai, Y. Influence of rice proteins on eating quality of cooked rice and on aroma and flavor of sake. Cereal Chem. 2006, 83, 439–446. [Google Scholar] [CrossRef]
- Ning, H.F.; Qiao, J.F.; Liu, Z.H.; Lin, Z.M.; Li, G.H.; Wang, Q.S.; Wang, S.H.; Ding, Y.F. Distribution of proteins and amino acids in milled and brown rice as affected by nitrogen fertilization and genotype. J. Cereal Sci. 2010, 52, 90–95. [Google Scholar] [CrossRef]
- Wei, K.; Wang, X.D.; Chen, X.; Wu, F.B.; Zhang, G.P. Hormonal changes in grains of cv. Triumph and its mutant TL43 as affected by nitrogen fertilizer at heading time. J. Cereal Sci. 2009, 49, 246–249. [Google Scholar] [CrossRef]
- Souza, S.R.; Stark, E.M.L.M.; Fernandes, M.S. Foliar spraying of rice with nitrogen: Effect on protein levels, protein fractions, and grain weight. J. Plant Nutr. 1999, 22, 579–588. [Google Scholar] [CrossRef]
- Krishnan, S.; Dayanandan, P. Structural and histochemical studies on grain-filling in the caryopsis of rice (Oryza sativa L.). J. Biosci. 2003, 28, 455–469. [Google Scholar] [CrossRef]
- Tong, C.; Gao, H.; Luo, S.; Liu, L.; Bao, J. Impact of Postharvest Operations on Rice Grain Quality: A Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 626–640. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Xue, Y.; Liu, D.; Kong, W.; Xue, Y.; Tang, Y.; Li, J.; Li, D.; Mei, P. Rational Application of Fertilizer Nitrogen to Soil in Combination with Foliar Zn Spraying Improved Zn Nutritional Quality of Wheat Grains. Front. Plant Sci. 2018, 9, 677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, J.G.; Liu, Z.H.; Lin, Z.M.; Alim, M.A.; Rehmani, M.I.A.; Li, G.H.; Wang, Q.S.; Wang, S.H.; Ding, Y.F. Phosphorus accumulation in grains of japonica rice as affected by nitrogen fertilizer. Plant Soil 2013, 369, 231–240. [Google Scholar] [CrossRef]
- Suzuki, M.; Tanaka, K.; Kuwano, M.; Yoshida, K.T. Expression pattern of inositol phosphate-related enzymes in rice (Oryza sativa L.): Implications for the phytic acid biosynthetic pathway. Gene 2007, 405, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Tai, T.H. Identification of genes necessary for wild-type levels of seed phytic acid in Arabidopsis thaliana using a reverse genetics approach. Mol. Genet. Genom. 2011, 286, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.C.; Vianna, G.R.; Cuneo, F.; Amaya-Farfan, J.; de Capdeville, G.; Rech, E.L.; Aragao, F.J. RNAi-mediated silencing of the myo-inositol-1-phosphate synthase gene (GmMIPS1) in transgenic soybean inhibited seed development and reduced phytate content. Planta 2006, 224, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.X.; Yu, X.M.; Yang, Q.H.; Fu, X.J.; Yuan, F.J. Transcriptome analysis identifies differentially expressed genes in the progenies of a cross between two low phytic acid soybean mutants. Sci. Rep. 2021, 11, 8740. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Primer (Forward, 5′-3′) | Primer (Reverse, 5′-3′) | Tm |
|---|---|---|---|---|
| OsActin | AK100267 | TGCGACGTGGATATTAGGAA | TGCCAGGGAACATAGTGGTA | 59–73 |
| RINO1 | AK103501 | AAACATTCCGATCCAAGGAG | CAAGCTCATAGAGGATGGCA | 59–81 |
| ITP5/6K-3 | AK067068 | GCTGAACTTCCTCCAAGACC | GAACCGATCTCTTGTCCCAT | 59–117 |
| ITP5/6K-6 | AK102571 | TTGGGTTTGATGTCGTTGTT | ATGACTGCTTGATTGCATCC | 59–132 |
| IPK1 | AK102842 | TGCCAAAGATTGGGTTTACA | TAGAAGGTGACGAGCCAGTG | 59–74 |
| IPK2 | AK072296 | GGAGCAAACCCTGTACCACT | ACCAGCTTCACCCTTACACC | 59–114 |
| MRP5 | AK121451 | TTGTCGCCGTGATAACATTT | CAGAAAGAACACCTCCAGCA | 59–72 |
| Year | Genotype | N Level | 1000-Grain Weight (g) | Albumin (%) | Globulin (%) | Prolamin (%) | Glutelin (%) | Total Protein (%) | Glutelin /Total Protein | Protein (mg Grain−1) | Glutelin/Prolamin |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2015 | XS110 | LN | 23.8 ± 0.2 b | 0.327 ± 0.030 c | 0.380 ± 0.053 b | 0.611 ± 0.092 b | 7.09 ± 0.16 c | 8.41 ± 0.18 c | 84.3 ± 1.5 a | 2.00 ± 0.04 b | 11.76 ± 1.65 a |
| MN | 26.5 ± 0.2 a | 0.454 ± 0.010 b | 0.444 ± 0.072 ab | 0.759 ± 0.029 ab | 7.62 ± 0.06 b | 9.28 ± 0.11 b | 82.1 ± 0.8 ab | 2.46 ± 0.04 a | 10.05 ± 0.31 b | ||
| HN | 24.2 ± 0.1 b | 0.518 ± 0.008 a | 0.522 ± 0.013 a | 0.786 ± 0.042 a | 8.04 ± 0.22 a | 9.86 ± 0.17 a | 81.5 ± 0.8 b | 2.38 ± 0.03 a | 10.23 ± 0.37 ab | ||
| HIPJ | LN | 23.8 ± 0.5 b | 0.627 ± 0.035 a | 0.533 ± 0.035 a | 0.167 ± 0.031 b | 6.69 ± 0.39 b | 8.01 ± 0.46 b | 83.4 ± 0.4 a | 1.91 ± 0.11 b | 40.73 ± 4.99 a | |
| MN | 25.3 ± 0.0 a | 0.537 ± 0.015 b | 0.500 ± 0.010 a | 0.300 ± 0.072 a | 7.47 ± 0.55 ab | 8.80 ± 0.52 ab | 84.8 ± 1.5 a | 2.22 ± 0.13 ab | 25.67 ± 4.9 b | ||
| HN | 25.1 ± 0.3 a | 0.653 ± 0.021 a | 0.517 ± 0.029 a | 0.240 ± 0.036 ab | 8.61 ± 0.75 a | 10.02 ± 0.70 a | 85.9 ± 1.4 a | 2.51 ± 0.21 a | 36.18 ± 3.73 b | ||
| 2016 | XS110 | LN | 23.7 ± 0.1 b | 0.223 ± 0.060 b | 0.356 ± 0.009 b | 0.584 ± 0.029 c | 6.96 ± 0.09 b | 8.12 ± 0.17 c | 85.7 ± 0.8 a | 1.93 ± 0.03 c | 11.93 ± 0.48 a |
| MN | 24.4 ± 0.5 ab | 0.291 ± 0.005 b | 0.418 ± 0.027 ab | 0.685 ± 0.028 b | 7.18 ± 0.12 b | 8.57 ± 0.11 b | 83.7 ± 0.3 b | 2.09 ± 0.02 b | 10.48 ± 0.28 b | ||
| HN | 24.9 ± 0.0 a | 0.455 ± 0.059 a | 0.493 ± 0.057 a | 0.754 ± 0.014 a | 7.64 ± 0.05 a | 9.35 ± 0.12 a | 81.8 ± 0.5 c | 2.33 ± 0.03 a | 10.14 ± 0.13 b | ||
| HIPJ | LN | 22.7 ± 0.1 b | 0.326 ± 0.038 b | 0.353 ± 0.053 b | 0.708 ± 0.042 a | 6.40 ± 0.45 b | 7.79 ± 0.37 b | 82.1 ± 1.9 a | 1.77 ± 0.08 c | 9.04 ± 0.33 a | |
| MN | 24.7 ± 0.2 a | 0.488 ± 0.022 a | 0.512 ± 0.021 a | 0.828 ± 0.056 a | 7.37 ± 0.37 a | 9.20 ± 0.32 a | 80.1 ± 1.3 a | 2.27 ± 0.08 b | 8.91 ± 0.27 a | ||
| HN | 24.6 ± 0.3 a | 0.494 ± 0.025 a | 0.580 ± 0.036 a | 1.086 ± 0.287 a | 8.03 ± 0.23 a | 10.19 ± 0.51 a | 78.9 ± 2.0 a | 2.51 ± 0.12 a | 7.68 ± 1.62 a | ||
| CV | 3.9 | 29.6 | 16.1 | 42.8 | 8.3 | 9.0 | 2.0 | 11.6 | 71.4 |
| Year | Genotype | N Level | 1000-Grain Weight (g) | Albumin (%) | Globulin (%) | Prolamin (%) | Glutelin (%) | Total Protein (%) | Glutelin/Total Protein | Protein (mg Grain−1) | Glutelin/Prolamin |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2015 | XS110 | N0 | 16.33 ± 0.17 c | 0.350 ± 0.012 e | 0.156 ± 0.005 e | 0.180 ± 0.003 e | 2.24 ± 0.12 e | 2.92 ± 0.14 e | 76.5 ± 0.7 c | 0.48 ± 0.03 e | 12.43 ± 0.51 d |
| N1 | 18.81 ± 0.06 b | 0.397 ± 0.007 d | 0.358 ± 0.005 d | 0.266 ± 0.003 d | 4.45 ± 0.11 d | 5.47 ± 0.10 d | 81.3 ± 0.4 b | 1.03 ± 0.02 d | 16.74 ± 0.22 b | ||
| N2 | 18.84 ± 0.11 b | 0.424 ± 0.004 c | 0.458 ± 0.005 c | 0.376 ± 0.014 c | 6.99 ± 0.08 c | 8.24 ± 0.06 c | 84.7 ± 0.3 a | 1.55 ± 0.02 c | 18.58 ± 0.62 a | ||
| N3 | 20.57 ± 0.08 a | 0.456 ± 0.011 b | 0.499 ± 0.003 b | 0.507 ± 0.022 b | 8.87 ± 0.17 b | 10.33 ± 0.17 b | 85.8 ± 0.3 a | 2.12 ± 0.03 a | 17.49 ± 0.43 ab | ||
| N4 | 15.43 ± 0.35 d | 0.588 ± 0.008 a | 0.589 ± 0.015 a | 0.696 ± 0.021 a | 10.75 ± 0.03 a | 12.62 ± 0.06 a | 85.2 ± 0.2 a | 1.95 ± 0.04 b | 15.46 ± 0.43 c | ||
| HIPJ | N0 | 17.36 ± 0.35 ab | 0.352 ± 0.007 d | 0.413 ± 0.002 c | 0.207 ± 0.038 c | 4.67 ± 0.30 d | 5.65 ± 0.28 e | 82.7 ± 1.3 b | 0.98 ± 0.03 d | 22.9 ± 2.55 ab | |
| N1 | 16.67 ± 1.13 bc | 0.424 ± 0.033 c | 0.516 ± 0.012 b | 0.256 ± 0.023 c | 6.87 ± 0.27 c | 8.07 ± 0.27 d | 85.2 ± 0.5 a | 1.34 ± 0.06 c | 26.89 ± 1.36 a | ||
| N2 | 18.92 ± 0.45 a | 0.508 ± 0.034 b | 0.610 ± 0.008 a | 0.368 ± 0.042 b | 7.39 ± 0.13 c | 8.87 ± 0.15 c | 83.3 ± 0.5 ab | 1.68 ± 0.03 b | 20.23 ± 2.01 bc | ||
| N3 | 17.54 ± 0.59 ab | 0.510 ± 0.019 b | 0.635 ± 0.010 a | 0.528 ± 0.013 a | 8.61 ± 0.51 b | 10.28 ± 0.52 b | 83.7 ± 0.8 ab | 1.80 ± 0.03 a | 16.29 ± 0.58 c | ||
| N4 | 15.21 ± 0.49 c | 0.577 ± 0.005 a | 0.622 ± 0.019 a | 0.536 ± 0.030 a | 9.56 ± 0.10 a | 11.30 ± 0.09 a | 84.6 ± 0.2 ab | 1.72 ± 0.04 ab | 17.88 ± 0.8 c | ||
| 2016 | XS110 | N0 | 17.22 ± 0.11 cd | 0.403 ± 0.006 d | 0.181 ± 0.010 d | 0.431 ± 0.011 d | 4.19 ± 0.09 e | 5.21 ± 0.09 e | 80.5 ± 0.3 c | 0.90 ± 0.01 d | 9.73 ± 0.05 c |
| N1 | 17.71 ± 0.26 c | 0.443 ± 0.006 c | 0.292 ± 0.010 c | 0.532 ± 0.036 c | 6.01 ± 0.21 d | 7.28 ± 0.23 d | 82.6 ± 0.4 b | 1.29 ± 0.03 c | 11.32 ± 0.44 b | ||
| N2 | 18.74 ± 0.57 b | 0.447 ± 0.006 bc | 0.382 ± 0.018 b | 0.631 ± 0.039 b | 7.10 ± 0.04 c | 8.56 ± 0.09 c | 82.9 ± 0.6 b | 1.60 ± 0.03 b | 11.28 ± 0.64 b | ||
| N3 | 20.18 ± 0.42 a | 0.470 ± 0.010 a | 0.401 ± 0.010 b | 0.640 ± 0.031 b | 8.48 ± 0.68 b | 10.00 ± 0.65 b | 84.8 ± 1.3 a | 2.02 ± 0.09 a | 13.25 ± 0.67 a | ||
| N4 | 16.52 ± 0.24 d | 0.467 ± 0.012 ab | 0.453 ± 0.005 a | 0.764 ± 0.035 a | 10.01 ± 0.09 a | 11.70 ± 0.11 a | 85.6 ± 0.2 a | 1.93 ± 0.01 a | 13.12 ± 0.49 a | ||
| HIPJ | N0 | 15.71 ± 0.27 d | 0.403 ± 0.006 c | 0.246 ± 0.005 d | 0.546 ± 0.026 d | 3.71 ± 0.05 d | 4.91 ± 0.06 d | 75.7 ± 0.5 b | 0.77 ± 0.01 e | 6.81 ± 0.25 c | |
| N1 | 16.51 ± 0.12 c | 0.440 ± 0.010 b | 0.424 ± 0.007 c | 0.653 ± 0.032 c | 6.64 ± 0.09 c | 8.15 ± 0.05 c | 81.4 ± 0.6 a | 1.35 ± 0.01 d | 10.18 ± 0.4 a | ||
| N2 | 19.21 ± 0.28 b | 0.453 ± 0.006 b | 0.474 ± 0.013 b | 0.744 ± 0.016 c | 7.67 ± 0.04 b | 9.34 ± 0.07 b | 82.1 ± 0.2 a | 1.79 ± 0.01 c | 10.3 ± 0.17 a | ||
| N3 | 20.58 ± 0.14 a | 0.477 ± 0.006 a | 0.519 ± 0.012 b | 0.877 ± 0.061 b | 8.51 ± 0.28 b | 10.39 ± 0.22 b | 82 ± 0.9 a | 2.14 ± 0.06 a | 9.73 ± 0.54 ab | ||
| N4 | 16.12 ± 0.43 cd | 0.480 ± 0.010 a | 0.588 ± 0.003 a | 1.102 ± 0.020 a | 9.77 ± 0.33 a | 11.94 ± 0.31 a | 81.8 ± 0.7 a | 1.92 ± 0.01 b | 8.86 ± 0.14 b | ||
| CV | 9.53 | 13.61 | 32.17 | 43.22 | 32.63 | 31.03 | 3.31 | 31.78 | 35.59 |
| Year | Genotype | N Level | PA Concentration mg kg−1 | Pi Concentration mg g−1 | TP Concentration mg kg−1 | PA-P/TP |
|---|---|---|---|---|---|---|
| 2015 | XS110 | N0 | 7.87 ± 0.18 a | 0.28 ± 0.01 c | 2.50 ± 0.06 b | 88.6 ± 0.2 a |
| N1 | 7.42 ± 0.11 ab | 0.29 ± 0.02 c | 2.39 ± 0.05 bc | 87.6 ± 0.8 a | ||
| N2 | 7.19 ± 0.07 bc | 0.33 ± 0.01 b | 2.36 ± 0.02 bc | 85.7 ± 0.3 b | ||
| N3 | 6.83 ± 0.31 c | 0.37 ± 0.01 b | 2.29 ± 0.09 c | 83.7 ± 0.5 c | ||
| N4 | 7.68 ± 0.12 a | 0.59 ± 0.01 a | 2.75 ± 0.04 a | 78.6 ± 0.3 d | ||
| HIPJ | N0 | 6.12 ± 0.24 a | 1.04 ± 0.03 b | 2.76 ± 0.09 b | 62.4 ± 0.7 ab | |
| N1 | 6.06 ± 0.38 a | 1.00 ± 0.06 b | 2.71 ± 0.05 b | 62.9 ± 2.9 a | ||
| N2 | 4.95 ± 0.24 b | 1.02 ± 0.07 b | 2.41 ± 0.14 c | 57.8 ± 0.6 abc | ||
| N3 | 4.45 ± 0.05 b | 1.09 ± 0.13 b | 2.35 ± 0.12 c | 53.5 ± 3.3 c | ||
| N4 | 6.20 ± 0.18 a | 1.32 ± 0.02 a | 3.07 ± 0.05 a | 56.9 ± 1 bc | ||
| 2016 | XS110 | N0 | 7.67 ± 0.22 a | 0.36 ± 0.01 c | 2.52 ± 0.06 ab | 85.8 ± 0.6 a |
| N1 | 7.91 ± 0.21 a | 0.35 ± 0.01 c | 2.58 ± 0.06 a | 86.5 ± 0.5 a | ||
| N2 | 6.78 ± 0.16 b | 0.36 ± 0.01 c | 2.27 ± 0.04 c | 84.2 ± 0.7 b | ||
| N3 | 6.56 ± 0.09 b | 0.42 ± 0.01 b | 2.26 ± 0.03 c | 81.5 ± 0.3 c | ||
| N4 | 6.71 ± 0.33 b | 0.47 ± 0.01 a | 2.36 ± 0.10 bc | 80.1 ± 0.6 c | ||
| HIPJ | N0 | 6.31 ± 0.31 a | 0.68 ± 0.01 d | 2.46 ± 0.08 b | 72.3 ± 1.2 a | |
| N1 | 5.84 ± 0.13 b | 0.87 ± 0.03 c | 2.51 ± 0.04 b | 65.5 ± 1.1 b | ||
| N2 | 5.64 ± 0.03 bc | 0.94 ± 0.02 bc | 2.53 ± 0.03 b | 62.9 ± 0.4 b | ||
| N3 | 4.32 ± 0.11 d | 1.03 ± 0.11 b | 2.25 ± 0.13 c | 54.2 ± 2 c | ||
| N4 | 5.38 ± 0.03 c | 1.27 ± 0.02 a | 2.79 ± 0.03 a | 54.3 ± 0.4 c | ||
| CV | 16.91 | 51.12 | 8.63 | 18.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, D.; Muneer, M.A.; Chen, X.; Rasmussen, S.K.; Wu, L.; Cai, Y.; Cheng, F. Response of Phytic Acid to Nitrogen Application and Its Relation to Protein Content in Rice Grain. Agronomy 2022, 12, 1234. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051234
Su D, Muneer MA, Chen X, Rasmussen SK, Wu L, Cai Y, Cheng F. Response of Phytic Acid to Nitrogen Application and Its Relation to Protein Content in Rice Grain. Agronomy. 2022; 12(5):1234. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051234
Chicago/Turabian StyleSu, Da, Muhammad Atif Muneer, Xiaohui Chen, Søren K. Rasmussen, Liangquan Wu, Yuanyang Cai, and Fangmin Cheng. 2022. "Response of Phytic Acid to Nitrogen Application and Its Relation to Protein Content in Rice Grain" Agronomy 12, no. 5: 1234. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051234