
Organic Amendment Types Influence Soil Properties, the Soil Bacterial Microbiome, and Tomato Growth


Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Pot Experiment Setup
2.3. Soil Property Analysis
2.4. DNA Extraction and Nanopore Sequencing of the 16S rRNA Gene
2.5. Bioinformatic Data Analysis
2.6. Statistical Analysis
3. Results
3.1. Effects of Organic Amendments on Tomato Growth
3.2. Effects of Organic Amendments on Soil Properties
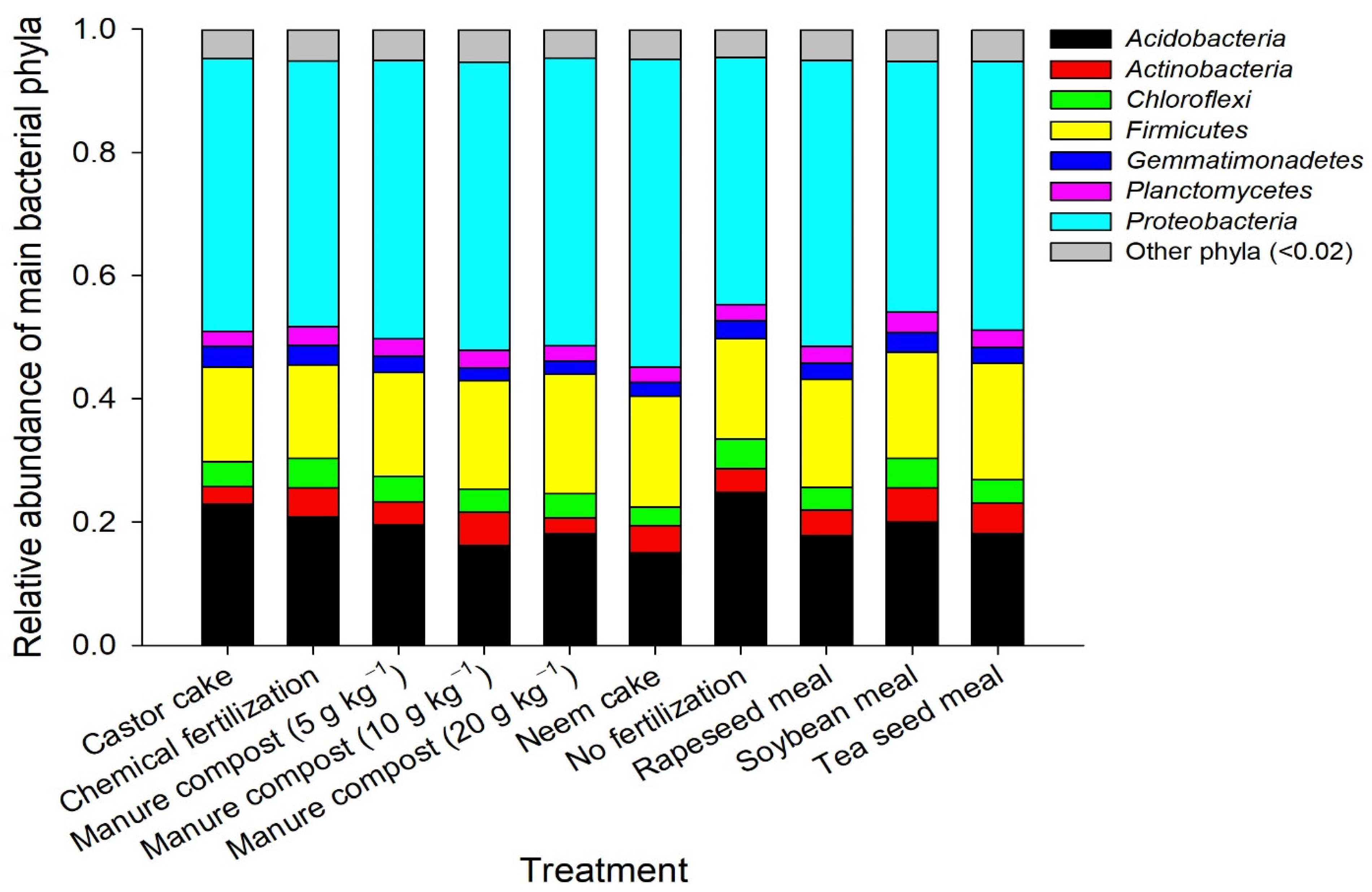
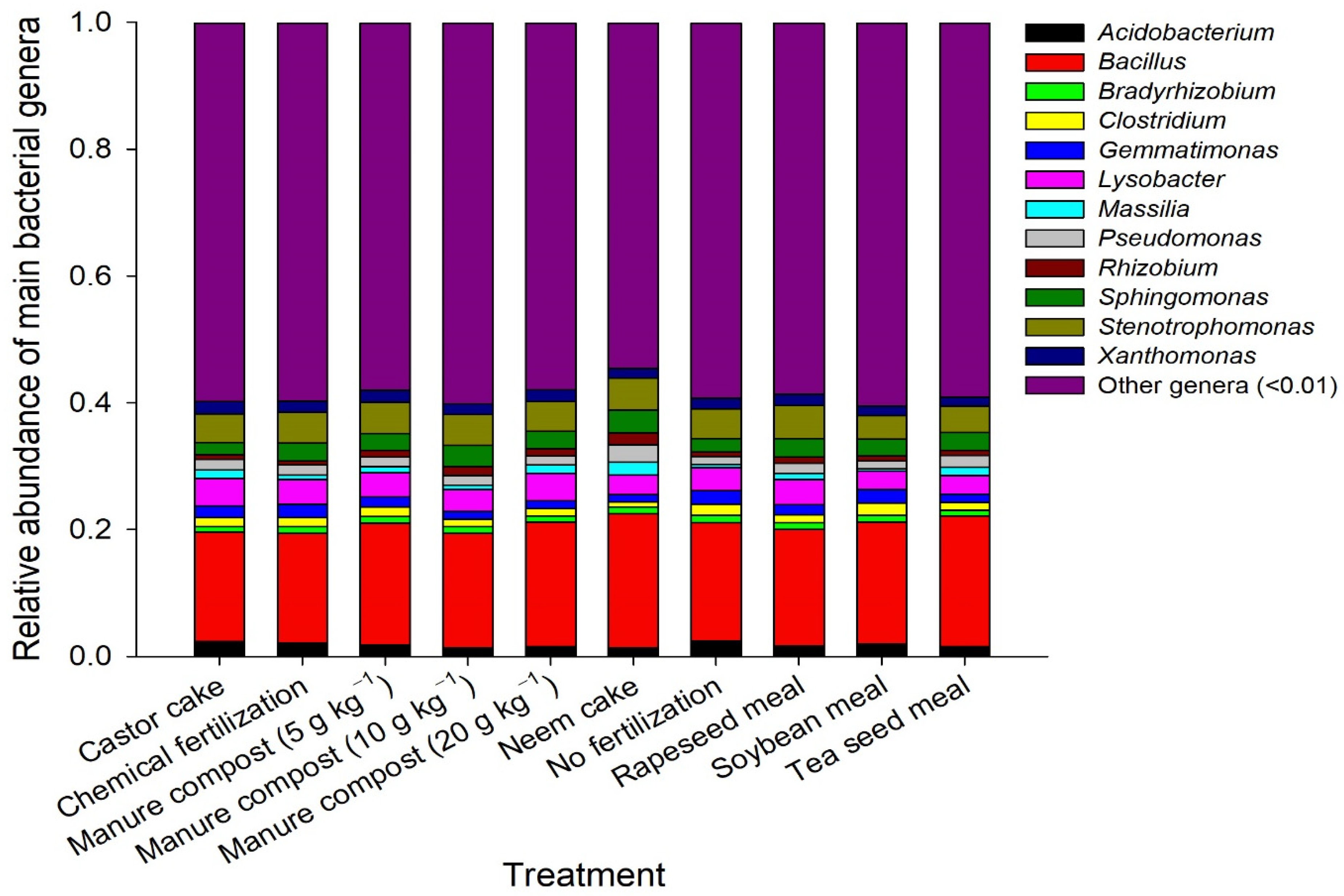
3.3. Microbial Community Composition
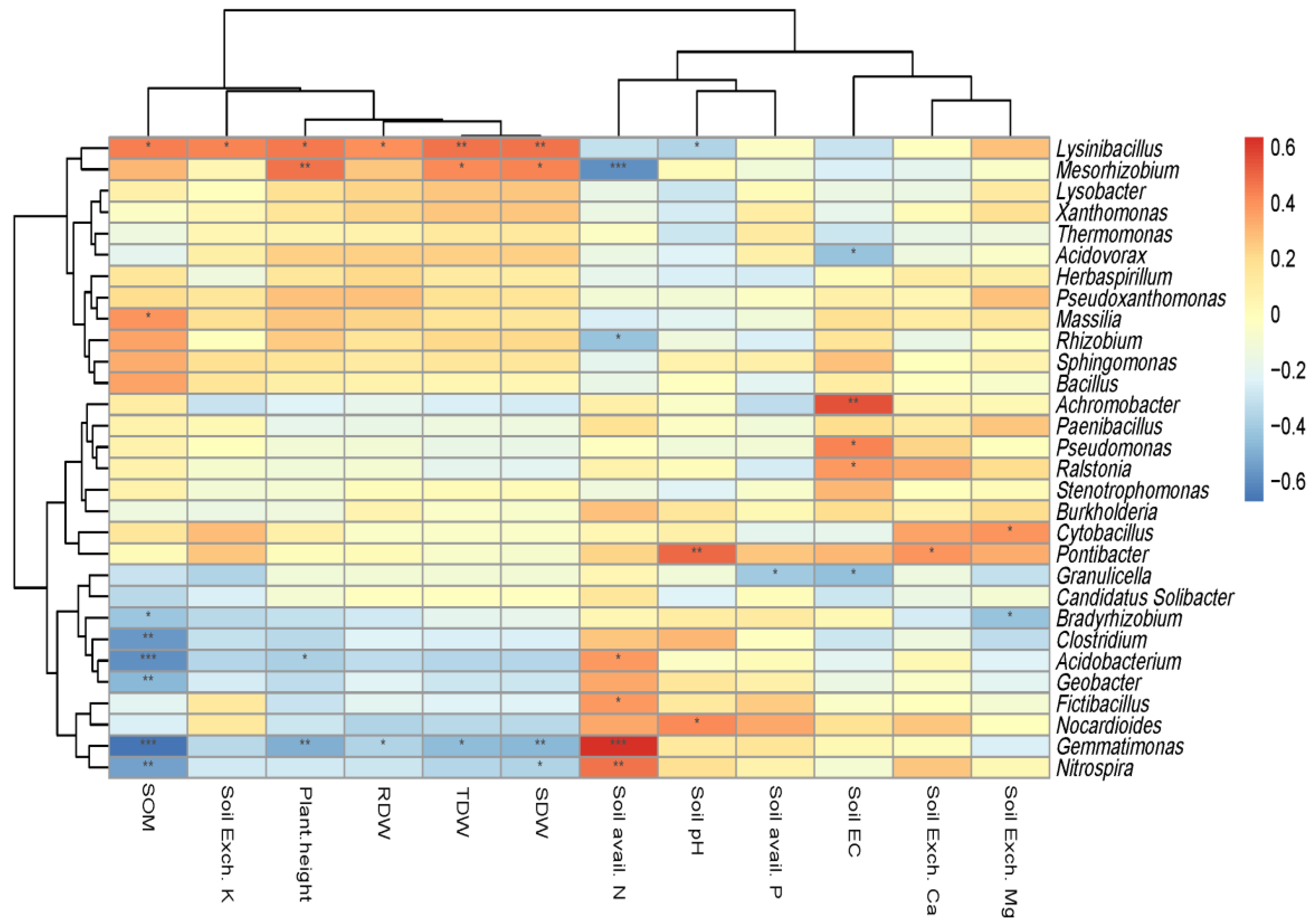
3.4. Relationship between Soil Properties, Tomato Growth, and Soil Microbiome
3.5. Relationship between Organic Amendments and Selected Dominant Genera, Soil Properties, and Tomato Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gryta, A.; Frąc, M.; Oszust, K. Genetic and metabolic diversity of soil microbiome in response to exogenous organic matter amendments. Agronomy 2020, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.; Black, A.L. Quantification of the effect of soil organic matter content on soil productivity. Soil Sci. Soc. Am. J. 1994, 58, 185–193. [Google Scholar] [CrossRef]
- Arthur, E.; Cornelis, W.; Razzaghi, F. Compost amendment to sandy soil affects soil properties and greenhouse tomato productivity. Compost Sci. Util. 2012, 20, 215–221. [Google Scholar] [CrossRef]
- Wang, S.; Chen, H.Y.H.; Tan, Y.; Fan, H.; Ruan, H. Fertilizer regime impacts on abundance and diversity of soil fauna across a poplar plantation chronosequence in coastal Eastern China. Sci. Rep. 2016, 6, 20816. [Google Scholar] [CrossRef] [PubMed]
- Šimon, T.; Javürek, M.; Mikanova, O.; Vach, M. The influence of tillage systems on soil organic matter and soil hydrophobicity. Soil Till. Res. 2009, 105, 44–48. [Google Scholar] [CrossRef]
- Keel, S.G.; Anken, T.; Büchi, L.; Chervet, A.; Fliessbach, A.; Flisch, R.; Huguenin-Elie, O.; Mäder, P.; Mayer, J.; Sinaj, S.; et al. Loss of soil organic carbon in Swiss long-term agricultural experiments over a wide range of management practices. Agric. Ecosyst. Environ. 2019, 286, 106654. [Google Scholar] [CrossRef] [Green Version]
- Cesarano, G.; De Filippis, F.; La Storia, A.; Scala, F.; Bonanomi, G. Organic amendment type and application frequency affect crop yields, soil fertility and microbiome composition. Appl. Soil Ecol. 2017, 120, 254–264. [Google Scholar] [CrossRef]
- Zhang, J.; Bei, S.; Li, B.; Zhang, J.; Christie, P.; Li, X. Organic fertilizer, but not heavy liming, enhances banana biomass, increases soil organic carbon and modifies soil microbiota. Appl. Soil Ecol. 2019, 136, 67–79. [Google Scholar] [CrossRef]
- Kheirfam, H. Increasing soil potential for carbon sequestration using microbes from biological soil crusts. J. Arid Environ. 2020, 172, 104022. [Google Scholar] [CrossRef]
- Bei, S.; Zhang, Y.; Li, T.; Christie, P.; Li, X.; Zhang, J. Response of the soil microbial community to different fertilizer inputs in a wheat-maize rotation on a calcareous soil. Agric. Ecosyst. Environ. 2018, 260, 58–69. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Wang, T.; Liu, Y.; Jia, S.; Gao, Y.; Liu, S. Effects of different fertilizer treatments on rhizosphere soil microbiome composition and functions. Land 2020, 9, 329. [Google Scholar] [CrossRef]
- Zhou, J.; Guan, D.; Zhou, B.; Zhao, B.; Ma, M.; Qin, J.; Jiang, X.; Chen, S.; Cao, F.; Shen, D.; et al. Influence of 34-years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China. Soil Biol. Biochem. 2015, 90, 42–51. [Google Scholar] [CrossRef]
- Sun, R.B.; Zhang, X.X.; Guo, X.S.; Wang, D.Z.; Chu, H.Y. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 2015, 88, 9–18. [Google Scholar] [CrossRef]
- Zhao, J.; Ni, T.; Li, J.; Lu, Q.; Fang, Z.; Huang, Q.; Zhang, R.; Li, R.; Shen, B.; Shen, Q. Effects of organic-inorganic compound fertilizer with reduced chemical fertilizer application on crop yields, soil biological activity and bacterial community structure in a rice-wheat cropping system. Appl. Soil Ecol. 2016, 99, 1–12. [Google Scholar] [CrossRef]
- Jiang, S.-Q.; Yu, Y.-N.; Gao, R.-W.; Wang, H.; Zhang, J.; Li, R.; Long, X.-H.; Shen, Q.-R.; Chen, W.; Cai, F. High-throughput absolute quantification sequencing reveals the effect of different fertilizer applications on bacterial community in a tomato cultivated coastal saline soil. Sci. Total Environ. 2019, 687, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, C.; Li, X.; Yang, X.; Zhao, L.; Liu, L.; Zhu, C.; Li, R. Linking plant ecological stoichiometry with soil nutrient and bacterial communities in apple orchards. Appl. Soil Ecol. 2018, 126, 1–10. [Google Scholar] [CrossRef]
- De Corato, U. Disease-suppressive compost enhances natural soil suppressiveness against soil-borne plant pathogens: A critical review. Rhizosphere 2020, 13, 100192. [Google Scholar] [CrossRef]
- Erika, C.; Griebel, S.; Naumann, M.; Pawelzik, E. Biodiversity in tomatoes: Is it reflected in nutrient density and nutritional yields under organic outdoor production? Front. Plant Sci. 2020, 11, 589692. [Google Scholar] [CrossRef]
- Maureira, F.; Rajagopalan, K.; Stöckle, C.O. Evaluating tomato production in open-field and high-tech greenhouse systems. J. Clean. Prod. 2022, 337, 130459. [Google Scholar] [CrossRef]
- Liu, E.; Yan, C.; Mei, X.; He, W.; Bing, S.H.; Ding, L.; Liu, Q.; Liu, S.; Fan, T. Long-term effect of chemical fertilizer, straw, and manure on soil chemical and biological properties in northwest China. Geoderma 2010, 158, 173–180. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, X.; Bao, E.C.; Li, J.S.; Zou, Z.R.; Cao, K. Bio-organic fertilizer with reduced rates of chemical fertilization improves soil fertility and enhances tomato yield and quality. Sci. Rep. 2020, 10, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasnain, M.; Chen, J.; Ahmed, N.; Memon, S.; Wang, L.; Wang, Y.; Wang, P. The effects of fertilizer type and application time on soil properties, plant traits, yield and quality of tomato. Sustainability 2020, 12, 9065. [Google Scholar] [CrossRef]
- Zhang, H.; Ding, W.; Yu, H.; He, X. Linking organic carbon accumulation to microbial community dynamics in a sandy loam soil: Result of 20 years compost and inorganic fertilizers repeated application experiment. Biol. Fertil. Soils 2015, 51, 137–150. [Google Scholar] [CrossRef]
- Vaughn, S.F.; Deppe, N.A.; Berhow, M.A.; Evangelista, R.L. Lesquerella press cake as an organic fertilizer for greenhouse tomatoes. Ind. Crops Prod. 2010, 32, 164–168. [Google Scholar] [CrossRef]
- De Corato, U. Effect of value-added organic co-products from four industrial chains on functioning of plant disease suppressive soil and their potentiality to enhance soil quality: A review from the perspective of a circular economy. Appl. Soil Ecol. 2021, 168, 104221. [Google Scholar] [CrossRef]
- Smith, J.L.; Doran, J.W. Measurement and use of pH and electrical conductivity. In Methods for Assessing Soil Quality; Doran, J.W., Jones, A.J., Eds.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 169–185. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining organic carbon in soils: Effect of variations in digestion conditions and of inorganic soil constituents. Soil Sci. 1934, 63, 251–263. [Google Scholar] [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen–total. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; Soil Science Society of America: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Bray, H.B.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–45. [Google Scholar] [CrossRef]
- Tan, K.H. Soil Sampling, Preparation, and Analysis; Marcel Dekker: New York, NY, USA, 1996; p. 408. [Google Scholar]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Song, L.; Breitwieser, F.P.; Salzberg, S.L. Centrifuge: Rapid and sensitive classification of metagenomic sequences. Genome Res. 2016, 26, 1721–1729. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Ren, H. TaxonKit: A practical and efficient NCBI taxonomy toolkit. J. Genet. Genom. 2021, 48, 844–850. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajene, I.J.; Khamis, F.M.; van Asch, B.; Pietersen, G.; Rasowo, B.A.; Ombura, F.L.; Wairimu, A.W.; Akutse, K.S.; Setamou, M.; Mohamed, S.; et al. Microbiome diversity in Diaphorina citri populations from Kenya and Tanzania shows links to China. PLoS ONE 2020, 15, e0235348. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Lee, S.-M.; Chen, T.-C. A generalized Good’s nonparametric coverage estimator. Chin. J. Math. 1988, 16, 189–199. [Google Scholar]
- Havlin, J.L.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. Soil Fertility and Fertilizers, 7th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Nikolajsen, M.T.; Pacholski, A.S.; Sommer, S.G. Urea ammonium nitrate solution treated with inhibitor technology: Effects on ammonia emission reduction, wheat yield, and inorganic N in soil. Agronomy 2020, 10, 161. [Google Scholar] [CrossRef] [Green Version]
- Cuartero, J.; Fernández-Muñoz, R. Tomato and salinity. Sci. Hortic. 1999, 78, 83–125. [Google Scholar] [CrossRef]
- Hu, Y.C.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Berry, P.M.; Sylvester-Bradley, R.; Philipps, L.; Hatch, D.J.; Cuttle, S.P.; Rayns, F.W.; Gosling, P. Is the productivity of organic farms restricted by the supply of available nitrogen? Soil Use Manag. 2002, 18, 248–255. [Google Scholar] [CrossRef]
- Ros, M.; Klammer, S.; Knapp, B.; Aichberger, K.; Insam, H. Long-term effects of compost amendment of soil on functional and structural diversity and microbial activity. Soil Use Manag. 2006, 22, 209–218. [Google Scholar] [CrossRef]
- Marschner, P.; Kandeler, E.; Marschner, B. Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biol. Biochem. 2003, 35, 453–461. [Google Scholar] [CrossRef]
- Fu, L.; Penton, C.R.; Ruan, Y.Z.; Shen, Z.Z.; Xue, C.; Li, R.; Shen, Q.R. Inducing the rhizosphere microbiome by biofertilizer application to suppress banana Fusarium wilt disease. Soil Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Pershina, E.; Valkonen, J.; Kurki, P.; Ivanova, E.; Chirak, E.; Korvigo, I.; Provorov, N.; Andronov, E. Comparative analysis of prokaryotic communities associated with organic and conventional farming systems. PLoS ONE 2015, 10, e0145072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El-Enshasy, H.A.; Dailin, D.J.; Suriani, N.L. Recent understanding of soil Acidobacteria and their ecological significance: A critical review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; Di Lonardo, D.P.; Bodelier, P.L.E. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. 2017, 93, fix006. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, I.; Yokota, A.; Yamazoe, A.; Fujiwara, T. Proposal of Lysinibacillus boronitolerans gen. nov sp nov., and transfer of Bacillus fusiformis to Lysinibacillus fusiformis comb. nov. and Bacillus sphaericus to Lysinibacillus sphaericus comb. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 1117–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahsan, N.; Marian, M.; Suga, H.; Shimizu, M. Lysinibacillus xylanilyticus strain GIC41 as a potential plant biostimulant. Microbes Environ. 2021, 36, ME21047. [Google Scholar] [CrossRef] [PubMed]
- Passera, A.; Rossato, M.; Oliver, J.S.; Battelli, G.; Shahzad, G.; Cosentino, E.; Sage, J.M.; Toffolatti, S.L.; Lopatriello, G.; Davis, J.R.; et al. Characterization of Lysinibacillus fusiformis strain S4C11: In vitro, in planta, and in silico analyses reveal a plant-beneficial microbe. Microbiol. Res. 2021, 244, 126665. [Google Scholar] [CrossRef] [PubMed]
- Shabanamol, S.; Thampi, M.; Sajana, P.; Varghese, S.; Karthika, S.; George, T.K.; Jisha, M.S. Characterization of the major antifungal extrolite from rice endophyte Lysinibacillus sphaericus against Rhizoctonia solani. Arch. Microbiol. 2021, 203, 2605–2613. [Google Scholar] [CrossRef]
- Naureen, Z.; Rehman, N.U.; Hussain, H.; Hussain, J.; Gilani, S.A.; Al Housni, S.K.; Mabood, F.; Khan, A.L.; Farooq, S.; Abbas, G.; et al. Exploring the potentials of Lysinibacillus sphaericus ZA9 for plant growth promotion and biocontrol activities against phytopathogenic fungi. Front. Microbiol. 2017, 8, 1477. [Google Scholar] [CrossRef]
- Menéndez, E.; Pérez-Yépez, J.; Hernández, M.; Rodríguez-Pérez, A.; Velázquez, E.; León-Barrios, M. Plant growth promotion abilities of phylogenetically diverse Mesorhizobium strains: Effect in the root colonization and development of tomato seedlings. Microorganisms 2020, 8, 412. [Google Scholar] [CrossRef] [Green Version]
- Prudent, M.; Causse, M.; Génard, M.; Tripodi, P.; Grandillo, S.; Bertin, N. Genetic and physiological analysis of tomato fruit weight and composition: Influence of carbon availability on QTL detection. J. Exp. Bot. 2009, 60, 923–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuvelink, E. Growth, development and yield of a tomato crop: Periodic destructive measurements in a greenhouse. Sci. Hortic. 1995, 61, 77–99. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Height (cm) | Shoot Dry Weight (g Plant−1) | Root Dry Weight (g Plant−1) |
|---|---|---|---|
| Castor cake | 14.3 ± 1.96 bc a | 0.56 ± 0.15 c | 0.11 ± 0.03 b |
| Chemical fertilization | 11.3 ± 1.87 ef | 0.50 ± 0.18 cd | 0.09 ± 0.03 b–d |
| Manure compost (5 g kg−1) | 14.0 ± 2.03 c | 0.65 ± 0.22 bc | 0.10 ± 0.02 bc |
| Manure compost (10 g kg−1) | 15.9 ± 1.98 b | 0.77 ± 0.12 b | 0.11 ± 0.03 b |
| Manure compost (20 g kg−1) | 20.1 ± 1.13 a | 1.34 ± 0.19 a | 0.17 ± 0.03 a |
| Neem cake | 11.7 ± 1.27 de | 0.34 ± 0.10 de | 0.07 ± 0.02 cd |
| No fertilization | 8.05 ± 0.67 g | 0.25 ± 0.11 e | 0.04 ± 0.01 e |
| Rapeseed meal | 9.38 ± 2.37 fg | 0.32 ± 0.17 e | 0.06 ± 0.03 de |
| Soybean meal | 11.1 ± 1.17 ef | 0.30 ± 0.08 e | 0.06 ± 0.02 de |
| Tea seed meal | 13.6 ± 1.86 cd | 0.52 ± 0.13 cd | 0.10 ± 0.02 bc |
| Treatment | pH | EC a | OM b | Avail. N c | Bray-1P | Exch. K | Exch. Ca | Exch. Mg |
|---|---|---|---|---|---|---|---|---|
| (dS m−1) | (g kg−1) | (mg kg−1) | ||||||
| Castor cake | 7.47 ± 0.41 ab d | 0.32 ± 0.03 d | 26.8 ± 0.62 d | 73.8 ± 10.3 c | 25.1 ± 3.81 b–d | 165 ± 16.3 d–f | 6491 ± 154 a | 431 ± 9.32 a–c |
| Chemical fertilization | 6.88 ± 0.04 c | 0.52 ± 0.01 a | 27.8 ± 0.89 cd | 114 ± 13.3 a | 38.8 ± 4.21 a | 185 ± 12.9 b–d | 6192 ± 152 a–d | 423 ± 11.5 b–d |
| Manure compost (5 g kg−1) | 7.09 ± 0.35 bc | 0.36 ± 0.04 cd | 27.6 ± 0.53 cd | 45.6 ± 10.4 ef | 25.2 ± 8.98 b–d | 178 ± 21.5 c–e | 5877 ± 105 c–e | 406 ± 11.7 de |
| Manure compost (10 g kg−1) | 6.94 ± 0.66 c | 0.36 ± 0.03 cd | 30.5 ± 1.38 b | 33.1 ± 5.85 fg | 26.0 ± 5.65 bc | 191 ± 11.9 bc | 6025 ± 381 b–e | 421 ± 17.8 b–d |
| Manure compost (20 g kg−1) | 7.03 ± 0.45 c | 0.40 ± 0.08 bc | 35.7 ± 1.96 a | 30.7 ± 10.1 g | 31.3 ± 6.40 b | 233 ± 18.1 a | 5723 ± 89.7 e | 443 ± 14.9 a |
| Neem cake | 7.00 ± 0.09 c | 0.56 ± 0.07 a | 29.1 ± 1.86 cd | 79.0 ± 17.0 bc | 21.9 ± 9.94 cd | 174 ± 10.8 c–e | 6226 ± 244 a–c | 431 ± 24.2 a–c |
| No fertilization | 6.94 ± 0.06 c | 0.40 ± 0.06 bc | 26.4 ± 0.68 d | 59.3 ± 8.04 de | 15.0 ± 2.80 e | 150 ± 17.2 f | 5863 ± 96.3 de | 392 ± 6.48 e |
| Rapeseed meal | 7.09 ± 0.23 bc | 0.45 ± 0.08 b | 26.8 ± 3.18 d | 92.6 ± 11.7 b | 18.6 ± 2.56 de | 161 ± 8.53 ef | 6299 ± 98.2 ab | 437 ± 11.0 ab |
| Soybean meal | 7.63 ± 0.23 a | 0.41 ± 0.05 bc | 27.7 ± 0.88 cd | 108 ± 18.7 a | 24.5 ± 3.41 cd | 174 ± 14.5 c–e | 6271 ± 98.3 ab | 426 ± 13.0 a–c |
| Tea seed meal | 7.48 ± 0.27 ab | 0.44 ± 0.03 b | 30.1 ± 1.55 b | 71.7 ± 14.9 cd | 25.4 ± 4.99 bc | 202 ± 35.1 b | 6210 ± 686 a–d | 419 ± 20.9 cd |
| Treatment | Chao1 | ACE | Shannon | Chao’s Coverage |
|---|---|---|---|---|
| Castor cake | 2152 ± 98.7 a–d a | 2344 ± 82.5 ab | 5.03 ± 0.07 de | 0.970 ± 0.003 |
| Chemical fertilization | 2089 ± 24.5 cd | 2296 ± 25.0 ab | 5.03 ± 0.05 de | 0.967 ± 0.005 |
| Manure compost (5 g kg−1) | 2251 ± 54.6 a | 2397 ± 51.8 a | 5.18 ± 0.07 bc | 0.981 ± 0.003 |
| Manure compost (10 g kg−1) | 2175 ± 42.5 a–d | 2372 ± 24.1 a | 5.36 ± 0.02 a | 0.977 ± 0.004 |
| Manure compost (20 g kg−1) | 2065 ± 112 d | 2256 ± 83.9 b | 5.26 ± 0.05 ab | 0.964 ± 0.006 |
| Neem cake | 2218 ± 83.4 ab | 2366 ± 68.6 a | 5.39 ± 0.10 a | 0.968 ± 0.008 |
| No fertilization | 2084 ± 79.2 cd | 2255 ± 54.5 b | 4.90 ± 0.11 e | 0.972 ± 0.005 |
| Rapeseed meal | 2207 ± 74.5 a–c | 2358 ± 70.5 ab | 5.30 ± 0.13 ab | 0.970 ± 0.006 |
| Soybean meal | 2117 ± 36.5 b–d | 2308 ± 15.5 ab | 5.10 ± 0.03 cd | 0.967 ± 0.004 |
| Tea seed meal | 2164 ± 99.5 a–d | 2346 ± 83.7 ab | 5.29 ± 0.07 ab | 0.966 ± 0.003 |
| pH | EC a | SOM b | Avail. N c | Bray-1P | Exch. K | Exch. Ca | Exch. Mg | |
|---|---|---|---|---|---|---|---|---|
| Plant height | −0.11 c | −0.57 *** | 0.64 *** | −0.66 *** | 0.17 | 0.53 ** | −0.24 | 0.31 |
| Shoot dry weight | −0.19 | −0.56 ** | 0.60 *** | −0.71 *** | 0.18 | 0.46 ** | −0.38 * | 0.15 |
| Root dry weight | −0.05 | −0.46 ** | 0.52 *** | −0.49 ** | 0.27 | 0.48 ** | −0.29 | 0.17 |
| Chao1 richness | 0.05 | 0.10 | −0.22 | −0.12 | −0.20 | −0.18 | 0.13 | 0.03 |
| ACE | 0.11 | 0.04 | −0.21 | −0.10 | −0.11 | −0.09 | 0.21 | 0.12 |
| Shannon | 0.05 | 0.20 | 0.56 ** | −0.36 * | −0.07 | 0.31 | 0.08 | 0.37* |
| Acidobacteria | −0.11 | −0.26 | −0.52 ** | 0.27 | −0.06 | −0.31 | −0.03 | −0.30 |
| Actinobacteria | 0.48 ** | 0.30 | −0.16 | 0.22 | 0.23 | 0.10 | 0.29 | 0.03 |
| Chloroflexi | 0.14 | −0.17 | 0.43 * | 0.38 * | 0.29 | −0.09 | −0.09 | −0.22 |
| Firmicutes | −0.02 | −0.06 | 0.71 *** | −0.37 * | −0.21 | 0.44 * | −0.05 | 0.27 |
| Gemmatimonadetes | 0.03 | 0.02 | −0.61 *** | 0.62 * | 0.10 | −0.32 | 0.13 | −0.11 |
| Planctomycetes | 0.49 ** | 0.08 | −0.26 | 0.34 | 0.40 * | 0.06 | 0.06 | 0.02 |
| Proteobacteria | −0.21 | 0.18 | 0.45* | −0.39 * | −0.07 | 0.10 | −0.08 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, J.-Y.; Liu, C.-H.; Tampus, K.; Lin, Y.-C.; Huang, C.-H. Organic Amendment Types Influence Soil Properties, the Soil Bacterial Microbiome, and Tomato Growth. Agronomy 2022, 12, 1236. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051236
Su J-Y, Liu C-H, Tampus K, Lin Y-C, Huang C-H. Organic Amendment Types Influence Soil Properties, the Soil Bacterial Microbiome, and Tomato Growth. Agronomy. 2022; 12(5):1236. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051236
Chicago/Turabian StyleSu, Jia-Yang, Cheng-Huan Liu, Kimberly Tampus, Ya-Chi Lin, and Cheng-Hua Huang. 2022. "Organic Amendment Types Influence Soil Properties, the Soil Bacterial Microbiome, and Tomato Growth" Agronomy 12, no. 5: 1236. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051236