Response of Yellow Quality Protein Maize Inbred Lines to Drought stress at Seedling Stage


Department of Agronomy, Faculty of Science and Agriculture, University of Fort Hare, Private Bag X1314, Alice 5700, Eastern Cape, South Africa
*
Author to whom correspondence should be addressed.
Agronomy 2018, 8(12), 287; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120287
Submission received: 1 November 2018
/
Revised: 28 November 2018
/
Accepted: 29 November 2018
/
Published: 1 December 2018
(This article belongs to the Special Issue Gene Expression Regulation during Drought and Salt Stress in Crop Plants)
Abstract
:This study seeks to evaluate the response of 17 yellow Quality Protein Maize (QPM) inbred line seedlings to drought stress (DS), using different morphophysiological traits (plant height (PH), chlorophyll content (CC), stem diameter (SD), proline content (Pro), photochemical efficiency of photosystem II (PS II), canopy temperature (CT) and substomatal carbon dioxide concentration (Ci). The experiment was laid out in a randomized complete block design (RCBD) and replicated three times in a growth chamber. The seedlings were exposed to DS treatment by growing them at 20% field capacity. The control/well-watered (WW) treatments were kept at 80% field capacity throughout the experiment. Highly significant differences (p < 0.001) were observed for PH, SD, and Pro across environments. On the other hand, significant differences (p < 0.05) were observed for CC and PS II, while DS had no significant effects on Ci and CT. Proline content increased under DS compared to WW conditions. Inbred lines L34, L7, L5, L2, L16, and L6 had approximately equal or more Pro than the drought tolerant check (ZM1523). As such, these lines were regarded as drought tolerant. Taking all measured parameters into consideration, L7 performed notably better than the other inbred lines under DS.
1. Introduction
Quality protein maize (Zea mays L.) has almost double the quantities of amino acids lysine and tryptophan when compared to normal maize [1]. In comparison to white maize, yellow maize contains a significant amount of carotenoids, thus making it a good source of Vitamin A [2]. Due to its nutritional value, yellow QPM can be used as a protein and Vitamin A source for most poor people living in third world countries where maize is the staple food. As such, consumption of QPM by young children who are often weaned to soft porridge can perhaps lessen malnutrition diseases such as kwashiorkor and night blindness [3]. Moreover, QPM can be an economically valuable substitute for normal maize in stock feeds as it requires little or no supplementary protein source to balance feeds [4]. However, despite all the nutritional benefits associated with QPM, very little progress has been made towards breeding for tolerance to abiotic stress factors in QPM varieties [5].
One of the major constraints to maize production is drought stress (DS). Due to climate change, drought is projected to be a devastating and more frequent phenomenon affecting sub-Saharan Africa in the future [6]. During the 2015–2016 growing season, South Africa experienced a major drought spell which resulted in 31% maize yield reduction from the previous drought-stricken season [7]. In this case, drought occurred before planting. This resulted in a delay in maize planting by at least 40 days [8]. Such a delay presumably contributed to limited accumulation of heat units in maize plants, resulting in reduced yield throughout the country. As such, great urgency is needed in coming up with sustainable solutions of reducing yield losses elicited by DS.
Drought stress is very unpredictable and can occur at any developmental stage during the growth cycle of a plant. Most literature suggests the flowering stage of maize plant as the most susceptible developmental stage under DS. Therefore, previous breeding projects have been mainly focused on drought tolerance during the flowering and grain filling stages consequently giving less attention to seedling drought stress tolerance. However, Qayyum et al. [9] revealed that DS at seedling stage was equally important as DS at flowering. Drought stress occurring at seedling stage can reduce crop stand [10]. Thus, fewer plants tend to reach physiological maturity, which subsequently reduces final crop yield. Meeks et al. [11] postulated that screening maize cultivars at seedling stage could help identify drought tolerant inbred lines at imminent growth stages. Few studies ascertaining this proposition have been conducted in maize breeding programs [12].
Screening of inbred lines for stress tolerance is one of the most critical steps that should be taken at the start of every hybridization program. The tolerant cultivars generated from these inbred lines can be grown in drought prone areas. Such cultivars are of immense importance to resource-poor farmers who have limited options of alleviating drought. According to Devasirvatham and Tan [13], cultivars with improved tolerance against DS will greatly contribute to food security in the future. Studies aiming at screening for DS tolerance at seedling stage have been conducted in crops such as maize [10,11] and sorghum [14].
Different mechanisms help plants to adapt to dry conditions [15]. Of interest is the accumulation of free proline in drought-stressed plant tissues as first observed by Kemble and MacPherson [16]. Henceforth, free proline accumulation has been used as a marker for DS tolerance [17]. Furthermore, it has been used as a screening technique for DS tolerance in many plant species [18]. For instance, Spoljarevic et al. [19] successfully screened maize cultivars for drought tolerance using the proline determination assay. Accumulation of proline in drought-stressed plants allows plants to survive and recover by maintaining cell protein structure, scavenging for hydroxyl radicals, and balancing cell reduction and oxidation reactions through buffering, among many other functions [20].
When plants are exposed to DS, they usually undergo various morphophysiological changes which are associated with tolerance. Morphophysiological traits that have been associated with tolerance to drought stress include maintenance of chlorophyll content (CC), substomatal carbon dioxide concentration (Ci), photochemical efficiency of photosystem II (PS II), canopy temperature (CT), plant height (PH), and stem diameter (SD) [21,22,23,24], among others. The objective of this study was to assess selected morphophysiological and biochemical responses of yellow QPM inbred line seedlings exposed to drought stress. Tolerant inbred lines from this study can be recommended for use in breeding programs developing cultivars for drought-stricken areas.
2. Materials and Methods
2.1. Study Site, Plant Materials, Experimental Set-Up, and Management
The experiment was conducted under a controlled environment, in a growth chamber at the University of Fort Hare, Alice, South Africa (32°46′ S latitude, 26°50′ E longitude). Seventeen yellow QPM inbred lines (Table 1), obtained from Quality Seed (PTY) LTD, South Africa, were evaluated. A drought tolerant open pollinated variety (OPV), ZM1523, [25] was used as a tolerant check, mainly for the proline accumulation analysis.
The seeds used were initially sterilized by soaking them in 2% sodium hypochlorite solution for 15 minutes and later rinsed thoroughly using deionized water [26]. Two seeds were sown in well-perforated polyvinyl chloride (PVC) columns containing hygromix growing media. The PVC columns were 11 cm in diameter and 25 cm in length. Three days after emergence, the seedlings were thinned to one plant per column. The experiment was laid out in a split plot design with three replications per treatment (drought or control). The treatments were assigned to main plots while inbred lines were assigned to subplots with each subplot consisting of two PVC columns. The experiment was repeated twice to ensure accuracy of the results.
The protocol used for the drought stress treatment was that of Pfunde and Mutengwa [27], with moderate alterations. Seedlings were exposed to drought stress by growing them at a moisture level of 20% field capacity (FC), while the control was maintained at 80% FC. A SM300 moisture meter (Delta T Devices, Cambridge, UK) was used to maintain the moisture level within the growth media. General conditions within the growth chamber were as follows: temperature was set at 25 °C day/22 °C night, relative humidity of 40% day/60% night, and photosynthetic photon flux density (PPFD) of 650 mol m−2 s−1 under 12 h photoperiod and carbon dioxide gas at 400 ppm [28]. The duration of the experiment was three weeks and no weeds, pests, and diseases were observed.
2.2. Data Collection
Data on different morphophysiological traits and proline accumulation were recorded before terminating the experiment. Canopy temperature and CC were measured using an infrared thermometer (Sentry Optronics Corp, Taipei, Taiwan) and spad-502 chlorophyll meter (Konica Minolta, Hangzhou, Japan) respectively. Photochemical efficiency of photosystem II and Ci measurements were taken in the middle of the third fully developed leaf using an iFL Integrated Fluorometer and Gas Exchange System (ADC Bio Scientific Ltd, Hoddesdon, England). Stem diameter and PH were measured using a calibrated digital Vernier caliper and meter rule respectively. The height considered was from the surface of the media to the tip of the youngest fully developed leaf.
2.3. Proline Accumulation
Proline concentration (Pro) within the leaves was determined using the proline assay as described by Bates et al. [29]. Approximately 0.5 g of fully expanded leaves taken from each plant was homogenized in 10 mL of 3% aqueous sulfosalicylic acid. The homogenate was filtered through a Whatman 2 filter paper. The 2 mL of filtrate was reacted together with 2 mL of acid ninhydrin and 2 mL of glacial acetic acid in a water bath set at 100 °C for one hour. After heat treatment, the reaction was terminated in an ice bath by cooling. Thereafter, 4 mL of cold toluene was added into the reaction mixture. Proline content was measured using a Visible Spectrophotometer (V-1200) (Shanghai Mapada Instruments Co., Ltd, Shanghai, China) with a wavelength range of 325 to 1000 nm set at 520 nm. Toluene was used as the blank. Pure L-Proline in different concentrations was used to produce a standard curve of proline. The standard curve obtained was essential for calculating Pro in every leaf sample tested. The following formula was used to calculate Pro in different leaf samples.
where, 115.5 is the molecular weight of proline [29].
2.4. Data Analysis
The Shapiro–Wilk test was performed to determine the normality of distribution of the recorded data. Data from the two cycles was combined because the Bartlett’s test for homogeneity of error variances was not significant. The values measured for all traits were subjected to combined analysis of variance (ANOVA) using Statistix 10. Treatment means were compared at a probability level of 5% using the Tukey’s test.
3. Results
3.1. Analysis of Variance
Results from the ANOVA showed significant differences among inbred lines, between environments (drought stress and well-watered treatments) and in the interaction between inbred lines and environments. There were significant inbred line by environment interactions for all traits except for Ci and CT. Highly significant differences (p < 0.001) were observed among inbred lines for PH, SD, and Pro across the two environments. Significant differences (p < 0.05) were observed among inbred lines for CC and PS II, while there were no significant differences for Ci and CT across the environments.
3.2. Growth Parameters
3.2.1. Plant Height
Generally, a decrease in PH was observed under DS conditions for all inbred lines (Table 2). The seedlings grown under WW conditions had an average height of 39.0 cm while those grown under DS condition had an average height of 22.6 cm. Under DS, L7 (28.6 cm) recorded the maximum PH, closely followed by L17 (27.4 cm) and L34 (26.6 cm). The minimum PH obtained under DS was 18.5 cm (L5). Although PH of most lines varied across environments, PH of L7 and L17 did not vary across environments. Inbred lines whose height was not significantly reduced by DS can be considered as having a mannerism of drought tolerance.
3.2.2. Stem Diameter
The findings of this investigation showed that DS caused a significant reduction in SD for all inbred lines (Table 2). Generally, SD decreased by an average of 3.6 mm in response to drought stress. The highest SDs recorded under DS and WW conditions were 5.6 mm (L7) and 10.6 mm (L29) respectively. The second ranked SD value under DS was recorded for L17 (5.4 mm), which was closely followed by L16 (5.2 mm). Conversely, the lowest SDs recorded under DS and WW conditions were 3.8 mm (L30) and 6.4 mm (L3), respectively. The stem diameter of L30 did not differ significantly from all lines under DS except that of L7, L17, and L16. No inbred line had similar performance for SD across environments.
3.3. Physiological Performance of Inbred Lines
3.3.1. Photochemical Efficiency of Photosystem II
The mean PS II under DS was not significantly different from that recorded under well-watered conditions (Table 2). Additionally, there was no significant difference in PS II for all inbred lines across the environments except for L3 whose PS II was significantly reduced by DS. Inbred line 5 recorded the highest PS II under DS and WW conditions (0.760 and 0.768, respectively).
3.3.2. Chlorophyll Content
Lower CC readings were recorded under DS compared to WW conditions for most inbred lines. Additionally, CC did not vary significantly across environments for most of these lines (Table 3). In contrast, CC of L7 and L16 significantly varied across environments. Furthermore, L7 and L16 recorded higher CC under DS conditions (55.2 and 51.8 spad units, respectively), when compared to WW conditions (41.3 and 38.2 spad units, respectively). Inbred lines that performed best under WW were L29 (51.8 spad units), L13 (51.6 spad units), and L18 (49 spad units) while L7 (55.2 spad units), L16 (51.8 spad units), and L13 (49.5 spad units) performed best under DS. Conversely, inbred lines L27 (37.5 spad units) and L16 (38.2 spad units) recorded the lowest CC values under WW conditions while L30 (29.6 spad units), L27 (35.4 spad units), and L29 (36.9 spad units) recorded the lowest CC under DS.
3.3.3. Substomatal CO2 Concentration
Substomatal CO2 concentration was lower under DS compared to under WW conditions (Table 3) with an average concentration of 461 and 474 mol mol−1, respectively. However, Ci did not vary significantly for all lines under DS, under WW conditions, and across environments.
3.3.4. Canopy Temperature
Generally, high CT was recorded under DS compared to WW conditions (Table 3). The average temperature recorded under DS and WW conditions were 29.4 and 27.3 °C, respectively. However, CT for all lines did not significantly differ under DS and WW conditions. Additionally, CT of all lines did not significantly differ across environments except for L3 and L32. A significant increase in CT by 3.7 °C and 3.3 °C was observed for L3 and L32, respectively, under DS.
3.3.5. Proline Content
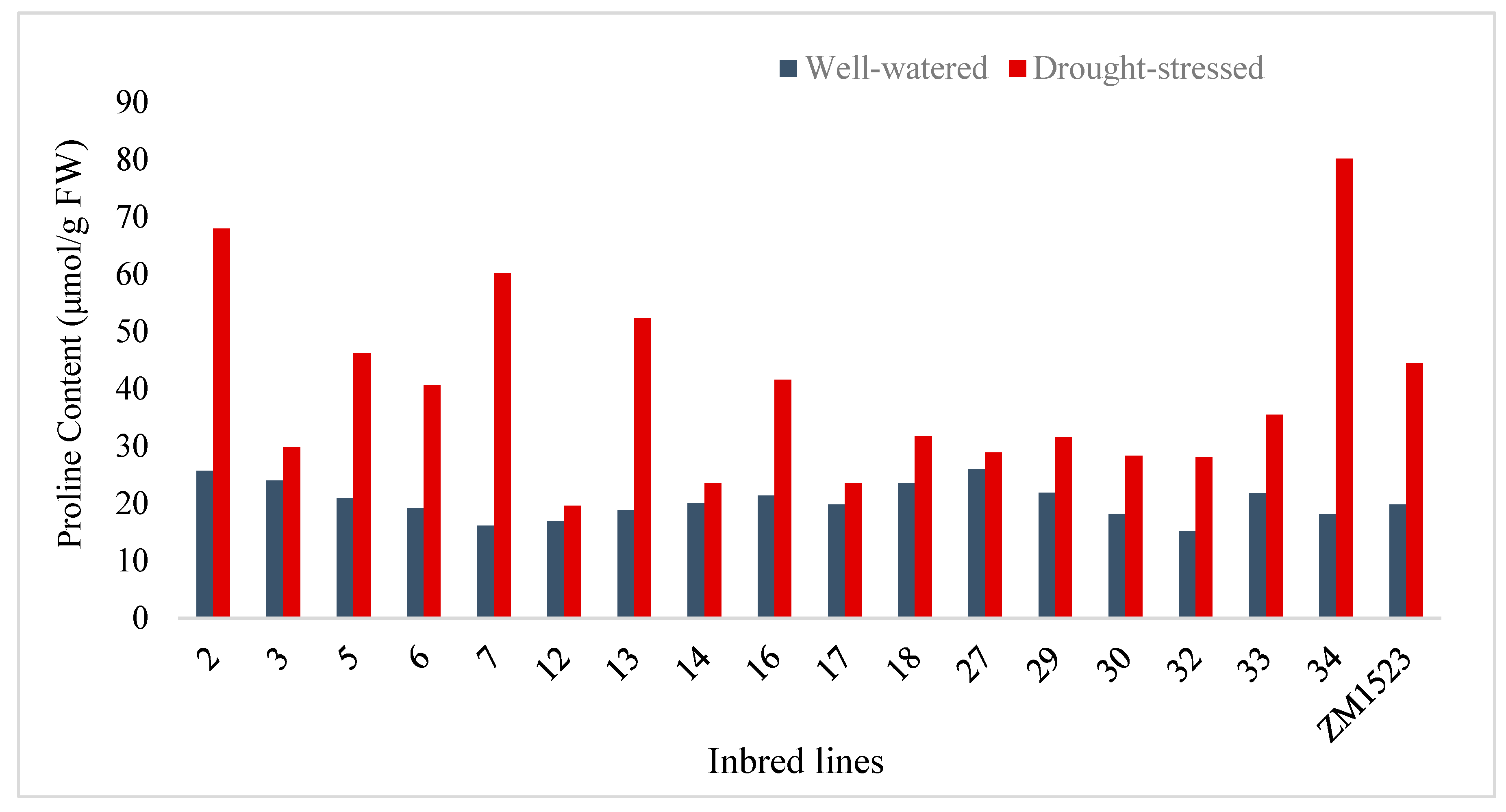
Generally, all inbred lines showed a significant increase in Pro under DS as observed in Table 3. Below is a graphical illustration of how the different yellow QPM inbred lines responded to DS and WW conditions (Figure 1).
A significant difference in Pro was observed on inbred lines exposed to DS (Table 3). Most inbred lines exhibited a significant difference in Pro across environments except for L12, L14, L17, and L27. The ranking of inbred lines in terms of Pro under DS was different from that under WW conditions. For example, L34 which recorded the highest Pro (80.2 µmol/g FW) under DS recorded a much lower Pro (18.1 µmol/g FW-rank 15) under WW conditions compared to other inbred lines. Under DS conditions, the Pro of L34 was closely followed by that of L2, L7, L13, and L5 in that order, with Pro of 68, 60.2, 52.4, and 46.3 µmol/g FW, respectively. Proline content in these five inbred lines significantly differed from each other and were higher than that of ZM1523 (44.2 µmol/g FW) under drought-stressed conditions. However, Pro of L5 was not significantly different from that of ZM1523, L16 (41.6 µmol/g FW), and L6 (40.7 µmol/g FW). Inbred lines with higher Pro than ZM1523, and those whose Pro was lower, but not significantly different from ZM1523 can be considered as showing a mannerism of drought tolerance.
4. Discussion
4.1. Growth Parameters
4.1.1. Plant Height
Based on the results, PH was significantly reduced under DS compared to WW conditions. Plant height of most lines significantly varied across environments except for L7 and L17. Hence, DS negatively affected PH. A decrease in maize PH under DS was also observed by Khodarahmpour and Hamidi [30]. Several reports have attributed inhibition of cell growth and cell elongation and/or expansion, due to reduced turgor pressure, as the main cause of reduction in PH during DS [18]. However, in some inbred lines (L7 and L17) reduction of PH under DS was not significant across environments. This showed that DS did not significantly negatively affect PH of these inbred lines. As such, these lines exhibited stable performance across environments, which is an indicator of drought tolerance.
4.1.2. Stem Diameter
Stem diameter was significantly reduced under DS and no inbred line performed alike across environments. Ali et al. [15] reported the same results and postulated that under drought-stressed conditions, most of the assimilates in maize seedlings were being diverted from the stem to support root growth thus causing a decrease in SD. A decrease of these growth parameters has also been observed in other plant species exposed to drought during various stages of growth [31]. Again, findings from this study showed that L7 and L17 had the highest SD and the least reduction in SD under DS. Hence, based on these observations, these lines were better performing than the other inbred lines evaluated under DS conditions.
4.2. Physiological Performance of Inbred Lines
4.2.1. Photochemical Efficiency of Photosystem II (PS II)
According to Terzi et al. [32], performance of PS II is greatly sensitive to DS compared to other photosynthetic components. In this study, L3 is the only inbred line that showed a significant reduction in PS II under DS compared to WW conditions. Reduction in PS II under DS has been reported in maize [20], in Arabidopsis [33], and in common bean [32]. Gao et al. [24] elucidated that the reduction of PS II observed under limited water stressed conditions could be due to damage caused on the photosystem II reaction centers resulting in overall inefficiency. As such, results from this study for L3 suggest that the reaction centers for photosystem II might have been damaged by DS.
Conversely, other inbred lines showed no significant reduction in PS II when exposed to DS. The performance of these inbred lines was stable across environments. Similar observations were made on birdsfoot trefoil (Lotus japonicus) by Sainz et al. [34], who postulated that DS alone could not effectively reduce performance of PS II.
4.2.2. Chlorophyll Content
Low CC readings were recorded under DS compared to WW conditions for most inbred lines except for L7 and L16. A significant loss in CC was observed on L29 when exposed to DS. Ali et al. [22] observed a loss of CC when maize plants were subjected to various levels of DS. Based on their results, CC loss was directly proportional to stress level. Manivannan et al. [35] and Mafakheri et al. [21] observed the same trend on different sunflower and chick pea cultivars respectively. According to Khayatnezhad and Gholamin [36] and Mafakheri et al. [21] the main cause of CC decrease under DS was due to the damage caused to the chloroplasts by the reactive oxygen species.
In this study, CC did not significantly vary across environments for most inbred lines except L7, L16 and L29. The same observation was made on maize seedlings exposed to DS by withholding irrigation for three days [37]. However, L7 and L16 recorded high CC under DS conditions compared to WW conditions. Similar observations were made by Sephri and Golparvar [38] on canola (Brassica napus L) cultivars exposed to drought. Canola cultivars that showed an increase in CC under DS were depicted as drought tolerant [38]. The probable reason why CC increases in plants exposed to DS is that the plant channels much of its resources towards the production of minerals and biochemical compounds so that it can quickly reach maturity, thereby evading stress.
4.2.3. Substomatal CO2 Concentration
Substomatal CO2 concentration is one of the factors that is greatly affected by different DS treatments and can be used to determine the plant’s ability to tolerate drought. Results from this investigation showed that Ci did not significantly vary for all inbred lines under DS, under WW conditions and across environments. In other words, DS had no effect on Ci. In contrast, Ci was significantly reduced in chickpea cultivars [21], maize [22], and in radish plants [39] that were exposed to DS. A decrease in Ci under DS has been attributed to the fact that stomatal opening is usually inhibited, and this could subsequently result in minute or no entry of CO2 into the leaf [21]. However, studies on wheat revealed that Ci increased upon exposing plants to DS treatments [40]. It has been suggested that such an increase in Ci could be the result of failure of the ‘photosynthesis machinery’ to utilize the internal CO2 [40].
4.2.4. Canopy Temperature
Results obtained in this investigation showed that CT increased by an average of 2.1 °C under DS when compared to WW conditions. However, CT for all inbred lines did not significantly differ under DS or WW conditions. This means that no inbred line responded differently from the other lines under DS or WW conditions. Additionally, CT of all inbred lines did not significantly differ across environments except for L3 and L32. This showed that DS had no effect on the CT of most of these inbred lines. According to Araus et al. [23], CT increment has been used as an effective secondary trait for screening for drought tolerance. Plants which exhibited a significant increase in CT under DS were described as showing a mannerism of drought susceptibility. An increase in CT occurs when the plant is unable to effectively cool itself through transpiration. Ali et al. [22] observed an increase in CT in maize grown under drought-stressed conditions and concluded that reduced transpiration rate under DS might have caused the CT to increase.
Araus et al. [23] suggested that plants showing reduced CT under DS were probably capable of either maintaining high water content in their cells or had enhanced ability to withdraw water from the soil. Such plants were described as showing a mannerism of drought tolerance. The literature suggests that warmer CTs are greatly associated with reduced yield in wheat under DS [41,42]. Thus, L32 and L3 can be described as drought-susceptible lines with regards to CT.
4.2.5. Proline Content
Generally, Pro is released when plant species are subjected to any form of abiotic or biotic stress [21]. It has been postulated that a large amount of Pro is released under DS conditions [18,43]. As such, it was concluded that Pro metabolism is associated with stress tolerance. Generally, plants that produced high concentrations of Pro under DS were regarded as tolerant and vice versa. In this study, most inbred lines exhibited a significant increase in Pro under DS except for L12, L14, L17, and L27. Inbred lines whose Pro concentrations did not significantly vary across environments were described as showing a mannerism of drought susceptibility. On another note, the changes in the ranking of inbred lines when one moves from one environment to the other is normally due to line by environment interaction.
Inbred lines with higher Pro than ZM1523 (L34, L2, L7, L13, and L5) and those whose Pro did not significantly vary from that of ZM1523 (L16 and L6) were described as showing a mannerism of drought tolerance. Results obtained from this study showed that Pro increased by 1.1- to 4.4-times under DS compared to WW conditions. In drought-stressed chickpea, [21] a 10-foldincrease in Pro was recorded in comparison to non-stressed conditions. An et al. [44] observed an increase in Pro of 6.15 times in drought-stressed Periploca sepium relative to non-stressed conditions. This shows that Pro released under drought-stressed conditions could be dependent on the plant species under evaluation, different genotypes within the plant species or the intensity of stress to which the plants were exposed. According to Arjenaki et al. [45], the intensity and duration of DS governs how plants respond to it.
The role of proline under DS is yet to be fully comprehended. However, previous studies postulate that proline is an osmoregulant whose accumulation under DS improves the plant’s ability to withdraw water from drying growing media; it does this by maintaining a water potential gradient [46]. Under drought-stressed conditions, proline can prevent cell membrane damage [47]. Additionally, proline can act as a source of nitrogen and carbon which is needed when plants recover from stress [46]. The role of proline with special reference to maize under drought-stressed conditions has been discussed in detail by Zadebagheri et al. [47].
5. Conclusions
Drought stress caused morphophysiological changes among the yellow QPM inbred lines. The inbred lines responded differently for some traits under drought-stressed conditions. Based on the Pro results, L34, L7, L2, L5, L16, and L6 were considered as drought tolerant. However, when multiple secondary traits were considered, L7 consistently appeared to show mannerisms of drought tolerance. This experiment needs to be repeated under field conditions to confirm the results obtained under the controlled environment. Subsequently, selected drought-tolerant yellow QPM inbred lines can be used in breeding programs focused on developing drought-tolerant, protein-rich maize cultivars.
Author Contributions
N.C. contributed to experimental design, data collection, data analysis and interpretation, writing, editing, and review of the first and final drafts. M.C. contributed to funding acquisition, supervision, reviewing, writing, and editing of the first and final drafts.
Funding
The Govan Mbeki Research and Development Centre (GMRDC) of the University of Fort Hare and the National Research Foundation (NRF), Research and Technology (RTF) fund (Reference: RTF150416117258; Grant No: 98706) are acknowledged for funding the project.
Acknowledgments
Quality Seed (PTY) LTD is acknowledged for supplying the germplasm that was used for this study. We are grateful to the Department of Biochemistry and Microbiology for giving access to facilities used to conduct this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Prasanna, B.M.; Vasal, S.K.; Kassahun, B.; Singh, N.N. Quality protein maize. Curr. Sci. 2001, 81, 1308–1319. [Google Scholar]
- Pillay, K.; Derera, J.; Siwela, M.; Veldman, F.J. Consumer acceptance of yellow provitamin a-biofortified maize in KwaZulu-Natal. S. Afr. J. Clin. Nutr. 2011, 24, 186–191. [Google Scholar] [CrossRef]
- Menkir, A.; Liu, W.; White, W.S.; Maziya-Dixon, B.; Rocheford, T. Carotenoid diversity in tropical-adapted yellow maize inbred lines. Food Chem. 2008, 109, 52–529. [Google Scholar] [CrossRef]
- Tiwari, M.R.; Neopane, D.; Paudel, T.P.; Singh, U.M. Evaluation of Quality Protein Maize (QPM) and normal maize for growth performance of broiler chicken in Nepal. GJSFR 2013, 13, 35–49. [Google Scholar]
- Tandzi, N.L.; Mutengwa, C.; Ngonkeu, E.; Woïn, N.; Gracen, V. Breeding for Quality Protein Maize (QPM) varieties: A review. Agronomy 2018, 7, 80. [Google Scholar] [CrossRef]
- Intergovernmental Panal on Climate Change (IPCC). The Fifth Assessment Report-What is in it for Africa. Available online: https://cdkn.org/ar5-toolkit/ar5-africa/?loclang=en_gb (accessed on 12 November 2015).
- Grain South Africa. Drought. Available online: http://www.grainsa.co.za/pages/industrreports/production-reports (accessed on 16 May 2016).
- United States Department of Agriculture (USDA). Drought Delays 2015/16 Corn Planting in South Africa. Commodity intelligency report. Available online: http://pecad.fas.usda.gov/highlights/2015/11/RSA/index.htm (accessed on 28 May 2016).
- Qayyum, A.; Ahmad, S.; Liaqat, S.; Malik, W.; Noor, E.; Saeed, H.M.; Hanif, M. Screening for drought tolerance in maize (Zea mays L.) hybrids at an early seedling stage. Afr. J. Agric. Res. 2012, 7, 3594–3604. [Google Scholar]
- Khayatnezhad, M.; Gholamin, R.; Jamaatie-Somarin, S.H.; Zabihi-Mahmoodabad, R. Effects of PEG stress on corn cultivars (Zea mays L.) at germination stage. World Appl. Sci. 2010, 11, 504–506. [Google Scholar]
- Meeks, M.; Murray, S.C.; Hague, S.; Hays, D. Measuring maize seedlings drought response in search of tolerant germplasm. Agron. J. 2013, 3, 135–147. [Google Scholar] [CrossRef]
- Djemel, A.; Álvarez-Iglesias, L.; Pedrol, N.; López-Malvar, A.; Ordás, A.; Revilla, P. Identification of drought tolerant populations at multi-stage growth phases in temperate maize germplasm. Euphytica 2018, 214, 138. [Google Scholar] [CrossRef]
- Devasirvatham, V.; Tan, D. Impact of high temperature and drought stresses on chickpea production. Agronomy 2018, 8, 145. [Google Scholar] [CrossRef]
- Bibi, A.; Sadaqat, H.A.; Tahir, M.H.N.; Akram, H.M. Screening of Sorghum (Sorghum bicolor Var Moench) for drought tolerance at seedling stage in polyethylene glycol. J. Anim. Plant Sci. 2012, 22, 671–678. [Google Scholar]
- Gaufichon, L.; Reisdorf-Cren, M.; Rothstein, S.J.; Chardon, F.; Suzuki, A. Biological functions of asparagine synthesis in plants. Plant Sci. 2010, 179, 141–153. [Google Scholar] [CrossRef]
- Kemble, A.R.; MacPherson, H.T. Liberation of amino acids in perennial ryegrass during wilting. Biochem. J. 1954, 53, 46–49. [Google Scholar] [CrossRef]
- Moussa, H.R.; Abdel-Aziz, S.M. Comparative response of drought tolerant and drought sensitive maize inbred lines to limited water stress. Aust. J. Crop Sci. 2008, 1, 31–36. [Google Scholar]
- Liu, H.; Sultan, M.A.F.; Zhao, H.X. The Screening of limited water stress tolerant wheat cultivars with physiological indices. GJESM 2013, 3, 211–218. [Google Scholar]
- Spoljarevic, M.; Agic, D.; Lisjak, M.; Gumze, A.; Wilson, I.D.; Hancock, J.T.; Teklic, T. The relationship of proline content and metabolism on the productivity of maize plants. Plant Signal Behav. 2011, 6, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Ali, Z.; Basra, S.M.A.; Munir, H.; Mahmood, A.; Yousaf, S. Mitigation of drought stress in maize by natural and synthetic growth promoters. J. Agric. Soc. Sci. 2011, 7, 56–62. [Google Scholar]
- Araus, J.L.; Serret, M.D.; Edmeades, G.O. Phenotyping maize for adaptation to drought. Front Physiol. 2012, 3, 305–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Zhang, R.H.; Wang, W.B.; Li, Z.W.; Xue, J.Q. Effects of drought stress on performance of photosystem II in maize seedling stage. J. Appl. Ecol. 2015, 26, 139–1396. [Google Scholar]
- Fischer, K.; Van den Berg, J.; Mutengwa, C. Is Bt maize effective in improving South African smallholder agriculture? S. Afr. J. Sci. 2015, 111, 01–02. [Google Scholar] [CrossRef]
- Kaviani, B. Proline accumulation and growth of soybean callus under salt and limited water stress. Int. J. Agric. Biol. 2008, 10, 221–223. [Google Scholar]
- Pfunde, C.N.; Mutengwa, C.S. Combining ability of quality protein maize inbred line for seedling tolerance to drought stress. Philipp. J. Crop Sci. 2016, 4, 11–12. [Google Scholar]
- Sinsawat, V.; Leipner, J.; Stamp, P.; Franchebound, Y. Effect of heat stress on the photosynthetic apparatus in maize (Zea mays L.) growth at control or high temperature. Environ. Exper. Bot. 2004, 52, 123–129. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldrenand, R.P.; Teare, I.D. Rapid determination off free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Khodarahmpour, Z.; Hamidi, J. Study of yield and yield components of corn (Zea mays L.) inbred lines to drought stress. Afr. J. Biotechnol. 2013, 11, 3099–3105. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.; Wang, L.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Terzi, R.; Saglam, A.; Kutlu, N.; Nar, H.; Kadioglu, A. Impact of soil drought stress on photochemical efficiency of photosystem II and antioxidant enzyme activities of Phaseolus vulgaris cultivars. Turk. J. Bot. 2010, 34, 1–10. [Google Scholar]
- Moustakas, M.; Sperdouli, I.; Kouna, T.; Antonopoulou, C.I.; Therios, I. Exogenous proline induces soluble sugar accumulation and alleviates drought stress effects on photosystem II functioning of Arabidopsis thaliana leaves. Plant Growth Regul. 2011, 65, 315–325. [Google Scholar] [CrossRef]
- Sainz, M.; Diaz, P.; Monza, J.; Borsani, O. Heat stress results in loss of chloroplast Cu/Zn superoxide dismutase and increased damage to Photosystem II in combined drought-heat stressed Lotus japonicus. Physiol. Plant. 2010, 140, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Manivannan, P.; Jaleel, C.A.; Sankar, B.; Kishorekumar, A.; Somasundaram, R.; Alagu, A.; Lakshmanan, G.M.; Panneerselvam, R. Growth, biochemical modifications and proline metabolism in Helianthus annuus L. as induced by drought stress. Colloid Surf. B. 2007, 59, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Khayatnezhad, M.; Gholamin, R. The effect of drought stress on leaf chlorophyll content and stress resistance in maize cultivars (Zea mays L.). Afr. J. Microbiol. Res. 2012, 6, 2844–2848. [Google Scholar] [CrossRef]
- Schlemmer, M.R.; Francis, D.D.; Shanahan, J.F.; Schepers, J.S. Remotely measuring chlorophyll content in corn leaves with differing nitrogen levels and relative water content. Agron J. 2005, 97, 106–112. [Google Scholar] [CrossRef]
- Sepehri, A.A.; Golparvar, R. The effect of drought stress on water relations, chlorophyll content and leaf area in canola cultivars (Brassica napus L.). Electron. J. Biol. 2011, 7, 49–53. [Google Scholar]
- Akram, N.A.; Waseem, M.; Ameen, R.; Ashraf, M. Trehalose pre-treatment induces drought tolerance in radish (Raphanus sativus L.) plants: Some key physio-biochemical traits. Acta. Physiol. Plant. 2016, 38, 1–10. [Google Scholar] [CrossRef]
- Saeidi, M.; Abdoli, M. Effect of drought stress during grain filling on yield and its components, gas exchange variables, and some physiological traits of wheat cultivars. J. Agric. Sci. Tech. 2015, 17, 885–898. [Google Scholar]
- Gutierrez, M.; Reynolds, M.P.; Raun, W.R.; Stone, M.L.; Klatt, A.R. Spectral water indices for assessing yield in elite bread wheat inbred lines under well-irrigated, water-stressed, and high-temperature conditions. Crop Sci. 2010, 50, 197–214. [Google Scholar] [CrossRef]
- Mason, R.E.; Singh, R.P. Considerations when deploying canopy temperature to select high yielding wheat breeding lines under drought and heat stress. Agron. J. 2014, 4, 191–201. [Google Scholar] [CrossRef]
- Nimisha, A.; Bahadur, S.N.; Kavita, Y. Interactive effects of rice residue and limited water stress on growth and metabolism of wheat seedlings. J. Stress Physiol. Biochem. 2014, 10, 232–245. [Google Scholar]
- An, Y.; Zhang, M.; Liu, G.; Han, R.; Liang, Z. Proline accumulation in leaves of Periploca sepium via both biosynthesis up-regulation and transport during recovery from severe drought. PLoS ONE. 2013, 8, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Arjenaki, F.G.; Jabbari, R.; Morshedi, A. Evaluation of drought stress on relative water content, chlorophyll content and mineral elements of wheat (Triticum aestivum L.) varieties. Int. J. Agri. Crop Sci. 2012, 4, 726–729. [Google Scholar]
- Rutherford, R.S. The assessment of proline accumulation as a mechanism of drought resistance in sugarcane. In Proceedings of the South African Sugar Technologists’ Association, Durban and Mount Edgecomb, KwaZulu-Natal, South Africa, 5–8 June 1989. [Google Scholar]
- Zadebagheri, M.; Azarpanah, A.; Javanmardi, S. Proline metabolite transport an efficient approach in corn yield improvement as response to drought conditions. Int. J. Farm Alli. Sci. 2014, 3, 453–461. [Google Scholar]
Figure 1.
Means of proline content of quality protein maize inbred lines exposed to drought stressed and well-watered conditions.
Figure 1.
Means of proline content of quality protein maize inbred lines exposed to drought stressed and well-watered conditions.


{kind=link}
Table 1.
Name and origin of 17 inbred lines used as plant materials.
| Inbred Line Number | Name | Origin * |
|---|---|---|
| 2 | D0620Y | M |
| 3 | K0315Y | F |
| 5 | S0178Y | M |
| 6 | S0249Y | M |
| 7 | S0825Y | M |
| 12 | A0595Y | F |
| 13 | R0445Y | F |
| 14 | CM132Y | M |
| 16 | CM321Y | P |
| 17 | EM86Y | B |
| 18 | EM88Y | B |
| 27 | HM46Y | F |
| 29 | HM267Y | F |
| 30 | HM48Y | L |
| 32 | HM63Y | L |
| 33 | HM1472Y | B |
| 34 | F155Y | L |
* Inbred lines were obtained from the following sources: M = M37W; P = Potchefstroom; F = F2834T; B = Brazil; L = Unknown.
Table 2.
Mean comparison among quality protein maize inbred lines based on plant height, stem diameter and photochemical efficiency of photosystem II under drought-stressed and well-watered conditions.
Table 2.
Mean comparison among quality protein maize inbred lines based on plant height, stem diameter and photochemical efficiency of photosystem II under drought-stressed and well-watered conditions.
| PH (cm) | SD (mm) | PS II | ||||
|---|---|---|---|---|---|---|
| Inbred line | Drought-stressed | Well-watered | Drought-stressed | Well-watered | Drought-stressed | Well-watered |
| 2 | 22.9 fg | 38.3 abc | 4.2 ijk | 7.8 cd | 0.733 abc | 0.764 ab |
| 3 | 18.6 g | 33.1 bcdef | 4.1 ijk | 6.4 efg | 0.597 d | 0.750 abc |
| 5 | 18.5 g | 42.1 ab | 4.9 hijk | 8.4 c | 0.760 abc | 0.768 a |
| 6 | 21.1 g | 38.4 abc | 4.0 jk | 8.8 bc | 0.701 abc | 0.718 abc |
| 7 | 28.6 cdefg | 36.2 abcd | 5.6 fgh | 7.9 cd | 0.677 cd | 0.751 abc |
| 12 | 22.2 g | 40.7 ab | 4.6 hijk | 8.0 cd | 0.729 abc | 0.752 abc |
| 13 | 20.5 g | 38.5 abc | 4.4 hijk | 7.6 cde | 0.726 abc | 0.765 ab |
| 14 | 21.9 g | 34.8 abcde | 4.2 ijk | 9.8 ab | 0.733 abc | 0.751 abc |
| 16 | 25.0 efg | 39.6 ab | 5.2 ghij | 7.9 cd | 0.708 abc | 0.750 abc |
| 17 | 27.4 defg | 33.1 bcdef | 5.4 ghi | 6.8 def | 0.731 abc | 0.746 abc |
| 18 | 24.6 efg | 43.1 ab | 4.4 hijk | 8.8 bc | 0.744 abc | 0.757 abc |
| 27 | 18.8 g | 41.1 ab | 4.0 jk | 8.6 bc | 0.751 abc | 0.764 ab |
| 29 | 24.0 fg | 36.9 abcd | 4.0 jk | 10.6 a | 0.737 abc | 0.743 abc |
| 30 | 19.0 g | 41.5 ab | 3.8 k | 8.0 cd | 0.752 abc | 0.763 ab |
| 32 | 20.5 g | 40.0 ab | 4.7 hijk | 7.8 cd | 0.739 abc | 0.753 abc |
| 33 | 23.5 fg | 45.2 a | 4.6 hijk | 6.9 def | 0.746 abc | 0.761 ab |
| 34 | 26.6 defg | 39.5 ab | 4.9 hijk | 8.0 cd | 0.683 bc | 0.739 abc |
PH = plant height, SD = stem diameter, and PS II = photochemical efficiency of photosystem II. Means followed by the same letter within the same column and across the environments (drought stress and well-watered) for a given trait are not significantly different according to Tukey’s test at p = 0.05.
Table 3.
Mean comparison among quality protein maize inbred lines based on chlorophyll content, substomatal CO2 concentration, canopy temperature, and proline content under drought-stressed and well-watered conditions.
Table 3.
Mean comparison among quality protein maize inbred lines based on chlorophyll content, substomatal CO2 concentration, canopy temperature, and proline content under drought-stressed and well-watered conditions.
| CC (Spad Units) | Ci (mol mol−1) | CT (°C) | PRO (µmol/g FW) | |||||
|---|---|---|---|---|---|---|---|---|
| Genotype | Drought-stressed | Well-watered | Drought-stressed | Well-watered | Drought-stressed | Well-watered | Drought-stressed | Well-watered |
| 2 | 44.5 abcde | 45.4 abcde | 465.0 | 480.7 | 27.6 | 27.2 | 68.0 b | 25.7 ijkl |
| 3 | 46.7 abcde | 47.7 abcde | 459.0 | 471.3 | 30.0 | 26.7 | 29.8 ghi | 24.0 jklm |
| 5 | 45.7 abcde | 48.9 abcd | 458.3 | 470.3 | 29.7 | 27.7 | 46.2 e | 20.9 klmnop |
| 6 | 43.1 abcde | 43.7 abcde | 457.3 | 470.7 | 29.5 | 27.3 | 40.7 ef | 19.2 mnopq |
| 7 | 55.2 a | 41.3 bcdef | 459.7 | 482.0 | 29.1 | 26.8 | 60.2 c | 16.1 pq |
| 12 | 38.5 cdef | 40.9 bcdef | 456.0 | 473.3 | 29.8 | 27.5 | 19.6 mnopq | 16.9 opq |
| 13 | 49.5 abcd | 51.6 abc | 467.7 | 474.7 | 28.6 | 27.1 | 52.4 d | 18.8 mnopq |
| 14 | 43.7 abcde | 46.3 abcde | 469.3 | 470.7 | 29.6 | 27.8 | 23.6 jkmn | 20.1 lmnopq |
| 16 | 51.8 ab | 38.2 def | 458.0 | 469.7 | 29.1 | 26.8 | 41.6 e | 21.4 klmnop |
| 17 | 42.2 abcdef | 42.8 abcde | 454.0 | 471.3 | 28.8 | 27.5 | 23.5 jklmn | 19.8 mnopq |
| 18 | 46.2 abcde | 49.0 abcd | 459.0 | 473.3 | 30.3 | 27.5 | 31.7 gh | 23.5 jklmn |
| 27 | 35.4 ef | 37.5 def | 462.0 | 471.3 | 28.5 | 27.4 | 28.9 hij | 26.0 hijk |
| 29 | 36.9 def | 51.8 ab | 459.0 | 482.0 | 30.2 | 27.5 | 31.5 gh | 21.9 klmno |
| 30 | 29.6 f | 41.1 bcdef | 473.7 | 476.0 | 29.2 | 27.6 | 28.3 hij | 18.2 nopq |
| 32 | 44.7 abcde | 45.5 abcde | 452.7 | 470.7 | 30.5 | 26.8 | 28.1 hij | 15.1 q |
| 33 | 41.5 bcdef | 43.1 abcde | 467.3 | 470.7 | 29.9 | 27.4 | 35.5 fg | 21.8 klmnop |
| 34 | 39.9 bcdef | 41.2 bcdef | 457.0 | 476.0 | 29.3 | 27.4 | 80.2 a | 18.1nopq |
| ZM1523 | - | - | - | - | - | - | 44.5 e | 19.8 mnopq |
CC = chlorophyll content, Ci = Substomatal CO2 concentration, CT = canopy temperature, and Pro = proline content. Means followed by the same letter within the same column and across the environments (drought stress and well-watered) for a given trait are not significantly different according to Tukey’s test at p = 0.05.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chiuta, N.E.; Mutengwa, C.S. Response of Yellow Quality Protein Maize Inbred Lines to Drought stress at Seedling Stage. Agronomy 2018, 8, 287. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120287
AMA Style
Chiuta NE, Mutengwa CS. Response of Yellow Quality Protein Maize Inbred Lines to Drought stress at Seedling Stage. Agronomy. 2018; 8(12):287. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120287
Chicago/Turabian StyleChiuta, Nyasha E., and Charles S. Mutengwa. 2018. "Response of Yellow Quality Protein Maize Inbred Lines to Drought stress at Seedling Stage" Agronomy 8, no. 12: 287. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120287
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.