Variability for Nitrogen Management in Genetically-Distant Maize (Zea mays L.) Lines: Impact of Post-Silking Nitrogen Limiting Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Determination of N Content and 15N-Abundance
2.3. Statistical Analysis
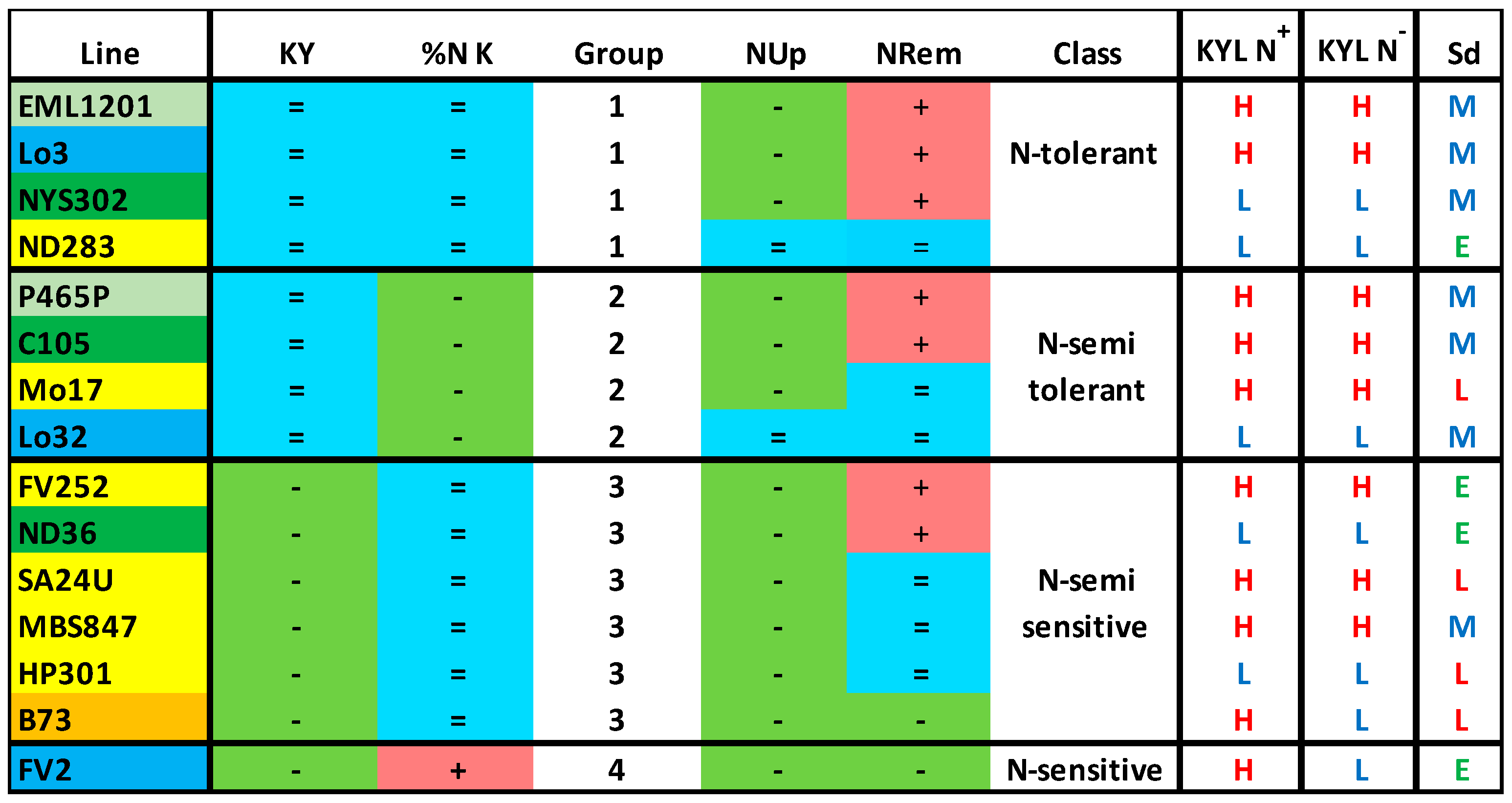
3. Results
3.1. Plant Agronomic Performances
3.2. Plant Nitrogen Management
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Hirel, B.; Tétu, T.; Lea, P.J.; Dubois, F. Improving nitrogen use efficiency in crops for a sustainable agriculture. Sustainability 2011, 3, 1452–1485. [Google Scholar] [CrossRef]
- Galloway, J.N.; Leach, A.M.; Bleeker, A.; Erisman, J.W. A chronology of human understanding of the nitrogen cycle. Philos. Trans. R. Soc. Lond. B 2013, 368. [Google Scholar] [CrossRef] [PubMed]
- Lassaletta, L.; Billen, G.; Garnier, J.; Bouwman, L.; Velazquez, E.; Mueller, N.D.; Gerber, J.S. Nitrogen use in the global food system: Past trends and future trajectories of agronomic performance, pollution, trade, and dietary demand. Environ. Res. Lett. 2016, 11, 095007. [Google Scholar] [CrossRef]
- Pathak, R.R.; Lochab, S.; Raghuram, N. Improving nitrogen-use efficiency. Compr. Biotechnol. 2011, 4, 209–218. [Google Scholar]
- Haegele, J.W.; Cook, K.A.; Nichols, D.M.; Below, F.E. Changes in nitrogen use traits associated with genetic improvement for grain yield of maize hybrids released in different decades. Crop Sci. 2013, 53, 1256–1268. [Google Scholar] [CrossRef]
- Han, M.; Okamoto, M.; Beatty, P.H.; Rothstein, S.J.; Good, A.J. The genetics of nitrogen use efficiency in crop plants. Ann. Rev. Genet. 2015, 49, 269–289. [Google Scholar] [CrossRef]
- Gallais, A.; Coque, M. Genetic variation and selection for nitrogen use efficiency in maize: A synthesis. Maydica 2005, 50, 531–547. [Google Scholar]
- Mastrodomenico, A.T.; Hendrix, C.C.; Below, F.E. Nitrogen use efficiency and the genetic variation of maize expired plant variety protection germplasm. Agriculture 2018, 8, 3. [Google Scholar] [CrossRef]
- Hossard, L.; Archer, D.W.; Bertrand, M.; Colnenne-David, C.; Debraeke, P.; Ernfors, M.; Jeuffroy, M.H.; Munier-Jolain, N.; Nilsson, C.; Sanford, G.R.; et al. A meta-analysis of maize and wheat yields under low-input vs. conventional and organic systems. Agron. J. 2016, 108, 1155–1167. [Google Scholar] [CrossRef]
- Meng, Q.; Cui, Z.; Yang, H.; Zhang, F.; Chen, X. Establishing high-yielding maize systems for sustainable intensification in China. Adv. Agron. 2018, 148, 85–109. [Google Scholar]
- Bänziger, M.; Edmeades, G.O.; Beck, D.; Bellon, M. Breeding for Drought and Nitrogen Stress Tolerance in Maize: From Theory to Practice; CIMMYT: Mexico City, Mexico, 2000. [Google Scholar]
- Chen, K.; Camberato, J.J.; Tuinstra, M.R.; Kumudini, S.; Tollenaar, M.; Vyn, T.J. Genetic improvement in density and nitrogen stress tolerance traits over 38 years of commercial maize hybrid release. Field Crops Res. 2016, 196, 438–451. [Google Scholar] [CrossRef]
- Cameron, K.C.; Di, H.J.; Moir, J.L. Nitrogen losses from the soil/plant system: A review. Ann. Appl. Biol. 2013, 162, 145–173. [Google Scholar] [CrossRef]
- Moss, B. Water pollution by agriculture. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 659–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Withers, P.J.A.; Neal, C.; Jarvie, H.P.; Doody, D.G. Agriculture and eutrophication: Where do we go from here? Sustainability 2014, 6, 5853–5875. [Google Scholar] [CrossRef]
- Verzeaux, J.; Alahmad, A.; Habbib, H.; Nivelle, E.; Roger, D.; Lacoux, J.; Decocq, G.; Hirel, B.; Catterou, M.; Spicher, F.; et al. Cover crops prevent the deleterious effect of nitrogen fertilization on bacterial diversity by maintaining the carbon concentration of ploughed soil. Geoderma 2016, 281, 49–57. [Google Scholar] [CrossRef]
- Smith, K.A. Changing views of nitrous oxide emissions from agricultural soil: Key controlling processes and assessment at different spatial scales. Eur. J. Soil Sci. 2017, 68, 137–155. [Google Scholar] [CrossRef]
- Gellings, C.W.; Parmenter, K.E. Energy efficiency in fertilizers production and use. In Efficient Use and Conservation of Energy; Gellings, C.W., Ed.; Encyclopedia of Life Support Systems; UNESCO Publications: Oxford, UK, 2016; Volume II, pp. 123–136. [Google Scholar]
- Hirel, B.; Lea, P.J. Genomics of nitrogen use efficiency in maize: From basic approaches to agronomic applications. In The Zea Mays Genome, Compendium of Plant Genomes; Bennetzen, J., Flint-Garcia, S., Hirsch, C., Tuberosa, R., Eds.; Springer: Cham, Switzerland, 2018; ISBN 978-3-319-97427-9. [Google Scholar]
- Oita, A.; Malik, A.; Kanemoto, K.; Geschke, A.; Nishijima, S.; Lenzen, M. Substantial nitrogen pollution embedded in international trade. Nat. Geosci. 2016, 9, 111–115. [Google Scholar] [CrossRef]
- McKenzie, F.C.; Williams, J. Sustainable food production: Constraints, challenges and choices by 2050. Food Secur. 2015, 7, 221–223. [Google Scholar] [CrossRef]
- Ceccarelli, S. GM crops, organic agriculture and breeding for sustainability. Sustainability 2014, 6, 4273–4286. [Google Scholar] [CrossRef]
- Swain, E.Y.; Rempelos, L.; Orr, C.H.; Hall, G.; Chapman, R.; Almadni, M.; Stockdale, EA.; Kidd, J.; Leifert, C.; Cooper, J.M. Optimizing nitrogen use efficiency in wheat and potatoes: Interaction between genotypes and agronomic practices. Euphytica 2014, 199, 119–136. [Google Scholar] [CrossRef]
- Coque, M.; Gallais, A. Genetic variation among European maize varieties for nitrogen use efficiency under low and high nitrogen fertilization. Maydica 2007, 52, 383–397. [Google Scholar]
- Ciampitti, A.; Vyn, T.J. Grain nitrogen source changes over time in maize: A review. Crop Sci. 2013, 53, 366–377. [Google Scholar] [CrossRef]
- Cañas, R.A.; Yesbergenova-Cuny, Z.; Simons, M.; Chardon, F.; Armengaud, P.; Quilleré, I.; Cukier, C.; Gibon, G.; Limami, A.M.; Nicolas, S.; et al. Exploiting the genetic diversity of maize using a combined metabolomic, enzyme activity profiling, and metabolic modeling approach to link leaf physiology to kernel yield. Plant Cell 2017, 29, 919–943. [Google Scholar] [CrossRef] [PubMed]
- Camus-Kulandaivelu, L.; Veyrieras, J.B.; Madur, D.; Combes, V.; Fourman, M.; Barraud, S.; Dubreuil, P.; Gouesnard, B.; Manicacci, D.; Charcosset, A. Maize adaptation to temperate climate: Relationship between population structure and polymorphism in the Dwarf8 gene. Genetics 2006, 172, 2449–2469. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, S.; Servin, B.; Bertin, P.; Madur, D.; Combes, V.; Dumas, F.; Brunel, D.; Laborde, J.; Charcosset, A.; Nicolas, S. Adaptation of maize to temperate climates: Mid-density genome-wide association genetics and diversity patterns reveal key genomic regions, with a major contribution of the Vgt2 (ZCN8) locus. PLoS ONE 2013, 8, e71377. [Google Scholar] [CrossRef] [PubMed]
- Yesbergenova-Cuny, Z.; Dinant, S.; Martin-Magniette, M.L.; Quillere, I.; Armengaud, P.; Monfalet, P.; Lea, P.J.; Hirel, B. Genetic variability of the phloem sap metabolite content of maize (Zea mays L.) during the kernel-filling period. Plant Sci. 2016, 252, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Coïc, Y.; Lesaint, C. Comment assurer une bonne nutrition en eau et en ions minéraux en horticulture. Hortic. Française 1971, 8, 11–14. [Google Scholar]
- Martin, A.; Belastegui-Macadam, X.; Quilleré, I.; Floriot, M.; Valadier, M.H.; Pommel, B.; Andrieu, B.; Donnison, I.; Hirel, B. Nitrogen management and senescence in two maize hybrids differing in the persistence of leaf greenness. Agronomic, physiological and molecular aspects. New Phytol. 2005, 167, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Gallais, A.; Coque, M.; Quilléré, I.; Le Gouis, J.; Prioul, J.L.; Hirel, B. Estimating proportions of N remobilization and of post-silking N uptake allocated to maize kernels by 15N labelling. Crop Sci. 2007, 47, 685–691. [Google Scholar] [CrossRef]
- Cliquet, J.B.; Deléens, E.; Mariotti, A. C and N mobilization from stalk and leaves during kernel filling by 13C and 15N tracing in Zea mays L. Plant Physiol. 1990, 94, 1547–1553. [Google Scholar] [CrossRef]
- Bertin, P.; Gallais, A. Genetic variation for nitrogen use efficiency in a set a recombinant maize inbred lines I. Agrophysiological results. Maydica 2000, 45, 53–66. [Google Scholar]
- Hirel, B.; Gallais, A. Nitrogen use efficiency—Physiological, molecular and genetic investigations towards crop improvement. In Advances in Maize (Essential Reviews in Experimental Biology); Prioul, J.L., Thévenot, C., Molnar, T., Eds.; Society for Experimental Biology: Cambridge, UK, 2011; Volume 3, pp. 285–310. [Google Scholar]
- Mueller, S.M.; Vyn, T.J. Maize plant resilience to N stress and post-silking N capacity changes over time: A review. Front. Plant Sci. 2016, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Chen, F.; Wu, Q.; Chen, Q.; Wang, J.; Yuan, L.; MI, G. Genetic improvement or root growth increases maize yield via enhanced post-silking uptake. Eur. J. Agron. 2015, 63, 55–61. [Google Scholar] [CrossRef]
- Garnett, T.; Plett, D.; Conn, V.; Conn, S.; Rabie, H.; Rafalski, J.A.; Dhugga, K.; Tester, M.A.; Kaiser, B. Variation for N uptake system in maize: Genotypic response to N supply. Front. Plant Sci. 2015, 6, 936. [Google Scholar] [CrossRef]
- Rajcan, I.; Tollenaar, M. Source: Sink ratio and leaf senescence in maize: II. Nitrogen metabolism during grain filling. Field Crop Res. 1999, 60, 255–265. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, C.; Wu, D.; Xia, T.; Chen, Q.; Chen, F.; Yuan, L.; Mi, G. Effects of nitrogen application rate on grain yield and grain nitrogen concentration in two maize hybrids with contrasting nitrogen remobilization efficiency. Eur. J. Agron. 2015, 62, 79–89. [Google Scholar] [CrossRef]
- Yang, L.; Guo, S.; Chen, Q.; Chen, F.; Yuan, L.; Mi, G. Use of stable nitrogen isotope to reveal the source-sink regulation of nitrogen uptake and remobilization during grain filling phase in maize. PLoS ONE 2016. [Google Scholar] [CrossRef] [PubMed]
- Coque, M.; Martin, A.; Veyrieras, J.B.; Hirel, B.; Gallais, A. Genetic variation for N-remobilization and post-silking N uptake in a set of maize recombinant inbred lines. 3. QTL detection and coincidences. Theor. Appl. Genet. 2008, 117, 729–747. [Google Scholar] [CrossRef]


{kind=link}
| Group | Line | KY (g) ± SE | KN ± SE | %NK ± SE | SDW (g) ± SE | Nt (mg) ± SE | 15NHI ± SE | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N+ | N− | t-test | N+ | N− | t-test | N+ | N− | t-test | N+ | N− | t-test | N+ | N− | t-test | N+ | N− | t-test | ||
| 1 | EML1201 | 92.6 ± 3.2 | 80.5 ± 8.5 | ns | 266 ± 8.4 | 216.5 ± 15.5 | * | 2.06 ± 0.12 | 1.94 ± 0.16 | ns | 136.2 ± 6.6 | 141.8 ± 14.3 | ns | 4.35 ± 0.19 | 3.07 ± 0.09 | ** | 0.36 ± 0.01 | 0.47 ± 0.03 | * |
| 1 | Lo3 | 86.1 ± 5.6 | 85.8 ± 6.2 | ns | 465 ± 12.9 | 469.3 ± 22.6 | ns | 1.86 ± 0.06 | 1.78 ± 0.03 | ns | 106.4 ± 12.8 | 91.4 ± 9.0 | ns | 3.09 ± 0.19 | 2.62 ± 0.16 | (*) | 0.46 ± 0.03 | 0.55 ± 0.02 | * |
| 1 | NYS302 | 35.2 ± 4.0 | 29.8 ± 4.1 | ns | 238 ± 20.3 | 174.5 ± 11.5 | ns | 2.56 ± 0.13 | 2.36 ± 0.10 | ns | 133.3 ± 3.2 | 90.7 ± 7.9 | ** | 3.69 ± 0.28 | 1.94 ± 0.13 | * | 0.21 ± 0.04 | 0.36 ± 0.04 | (*) |
| 1 | ND283 | 34.1 ± 2.0 | 35.3 ± 1.5 | ns | 177.7 ± 10.5 | 197.8 ± 3.8 | ns | 2.14 ± 0.15 | 2.14 ± 0.20 | ns | 43.1 ± 1.5 | 48.9 ± 4.0 | ns | 1.19 ± 0.08 | 1.20 ± 0.12 | ns | 0.61 ± 0.03 | 0.67 ± 0.05 | ns |
| 2 | P465P | 56.7 ± 7.6 | 70.4 ± 1.3 | ns | 298.5 ± 60.1 | 311.7 ± 13.3 | ns | 2.33 ± 0.08 | 2.04 ± 0.10 | (*) | 155.4 ± 17.7 | 134.7 ± 14.3 | ns | 3.33 ± 0.22 | 2.44 ± 0.13 | * | 0.38 ± 0.08 | 0.61 ± 0.04 | (*) |
| 2 | C105 | 52.2 ± 4.1 | 62.3 ± 8.3 | ns | 209 ± 14.0 | 227.3 ± 28.6 | ns | 2.17 ± 0.08 | 1.85 ± 0.07 | * | 81.8 ± 3.0 | 61.7 ± 7.8 | * | 2.68 ± 0.11 | 1.87 ± 0.11 | ** | 0.37 ± 0.01 | 0.62 ± 0.04 | ** |
| 2 | Mo17 | 94.4 ± 5.5 | 87.9 ± 15.7 | ns | 298.1 ± 21.3 | 295.8 ± 46.6 | ns | 2.31 ± 0.04 | 1.99 ± 0.13 | (*) | 155.3 ± 11.0 | 144.9 ± 7.5 | ns | 4.35 ± 0.23 | 3.31 ± 0.09 | ** | 0.44 ± 0.03 | 0.47 ± 0.07 | ns |
| 2 | Lo32 | 27.3 ± 0.4 | 24.8 ± 6.4 | ns | 223 ± 15.0 | 189 ± 47.8 | ns | 2.39 ± 0.04 | 2.15 ± 0.04 | * | 126.9 ± 13.0 | 121.0 ± 18.1 | ns | 3.28 ± 0.49 | 2.27 ± 0.20 | ns | 0.17 ± 0.04 | 0.24 ± 0.08 | ns |
| 3 | FV252 | 76.3 ± 3.5 | 59.1 ± 5.4 | * | 372.2 ± 20.1 | 295.4 ± 25.8 | * | 2.04 ± 0.13 | 1.93 ± 0.10 | ns | 99.9 ± 6.7 | 80.0 ± 6.6 | (*) | 3.07 ± 0.08 | 1.98 ± 0.15 | *** | 0.48 ± 0.03 | 0.59 ± 0.04 | * |
| 3 | ND36 | 37.2 ± 3.2 | 24.5 ± 0.8 | (*) | 245.7 ± 15.0 | 152.3 ± 3.8 | * | 2.58 ± 0.11 | 2.38 ± 0.27 | ns | 99.1 ± 16.9 | 62.0 ± 8.6 | ns | 3.03 ± 0.40 | 1.32 ± 0.19 | (*) | 0.30 ± 0.03 | 0.48 ± 0.02 | * |
| 3 | SA24U | 122.5 ± 2.5 | 85.1 ± 8.6 | (*) | 793.3 ± 0.3 | 553.8 ± 81.3 | (*) | 1.97 ± 0.08 | 2.02 ± 0.04 | ns | 191.2 ± 1.5 | 126.3 ± 29.0 | ns | 4.73 ± 0.02 | 2.99 ± 0.34 | * | 0.43 ± 0.01 | 0.60 ± 0.14 | ns |
| 3 | MBS847 | 105.9 ± 8.7 | 59.9 ± 7.4 | ** | 543 ± 32.3 | 311.4 ± 41.3 | ** | 2.12 ± 0.08 | 2.15 ± 0.07 | ns | 119.3 ± 11.1 | 107.9 ± 12.3 | ns | 3.94 ± 0.31 | 2.56 ± 0.28 | * | 0.52 ± 0.04 | 0.49 ± 0.04 | ns |
| 3 | HP301 | 49.3 ± 4.9 | 34.1 ± 3.6 | (*) | 468.9 ± 49.5 | 306.3 ± 39.1 | (*) | 2.32 ± 0.15 | 2.02 ± 0.17 | ns | 130.6 ± 2.1 | 160.2 ± 18.4 | ns | 3.12 ± 0.12 | 2.50 ± 0.10 | * | 0.32 ± 0.00 | 0.26 ± 0.04 | ns |
| 3 | B73 | 85.4 ± 19.0 | 19.2 ± 3.0 | (*) | 391.5 ± 54.0 | 97.8 ± 4.3 | * | 2.29 ± 0.27 | 2.45 ± 0.22 | ns | 159.2 ± 12.7 | 156.5 ± 27.5 | ns | 4.38 ± 0.14 | 3.03 ± 0.72 | (*) | 0.38 ± 0.05 | 0.16 ± 0.03 | * |
| 4 | FV2 | 54.4 ± 5.7 | 15.8 ± 2.6 | ** | 258.6 ± 19.8 | 76.7 ± 10.3 | *** | 2.04 ± 0.06 | 2.42 ± 0.16 | * | 75.5 ± 7.3 | 71.8 ± 7.0 | ns | 2.23 ± 0.20 | 1.55 ± 0.20 | (*) | 0.48 ± 0.04 | 0.24 ± 0.05 | * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quilleré, I.; Dargel-Graffin, C.; Lea, P.J.; Hirel, B. Variability for Nitrogen Management in Genetically-Distant Maize (Zea mays L.) Lines: Impact of Post-Silking Nitrogen Limiting Conditions. Agronomy 2018, 8, 309. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120309
Quilleré I, Dargel-Graffin C, Lea PJ, Hirel B. Variability for Nitrogen Management in Genetically-Distant Maize (Zea mays L.) Lines: Impact of Post-Silking Nitrogen Limiting Conditions. Agronomy. 2018; 8(12):309. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120309
Chicago/Turabian StyleQuilleré, Isabelle, Céline Dargel-Graffin, Peter J. Lea, and Bertrand Hirel. 2018. "Variability for Nitrogen Management in Genetically-Distant Maize (Zea mays L.) Lines: Impact of Post-Silking Nitrogen Limiting Conditions" Agronomy 8, no. 12: 309. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120309