Molecular Cloning and Structure–Function Analysis of a Trypsin Inhibitor from Tartary Buckwheat and Its Application in Combating Phytopathogenic Fungi


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Cloning of the FtTI Gene
2.2.1. Nucleic Acid Preparation
2.2.2. cDNA Cloning of the Central Fragment of FtTI
2.2.3. Genome Walking
2.3. Site-Directed Mutagenesis
2.4. Expression of FtTI and FtTI Mutants in E. coli
2.5. Affinity Purification of FtTI and FtTI Mutants
2.6. Molecular Weight Determination and Western Blot Analysis
2.7. Determination of Recombinant FtTI Inhibitory Activity
2.8. Bioassay of Inhibition of Spore Germination and Hyphal Growth by Recombinant FtTI
2.9. Experimental Design and Statistical Analysis
3. Results and Discussion
3.1. Cloning and Characterization of FtTI
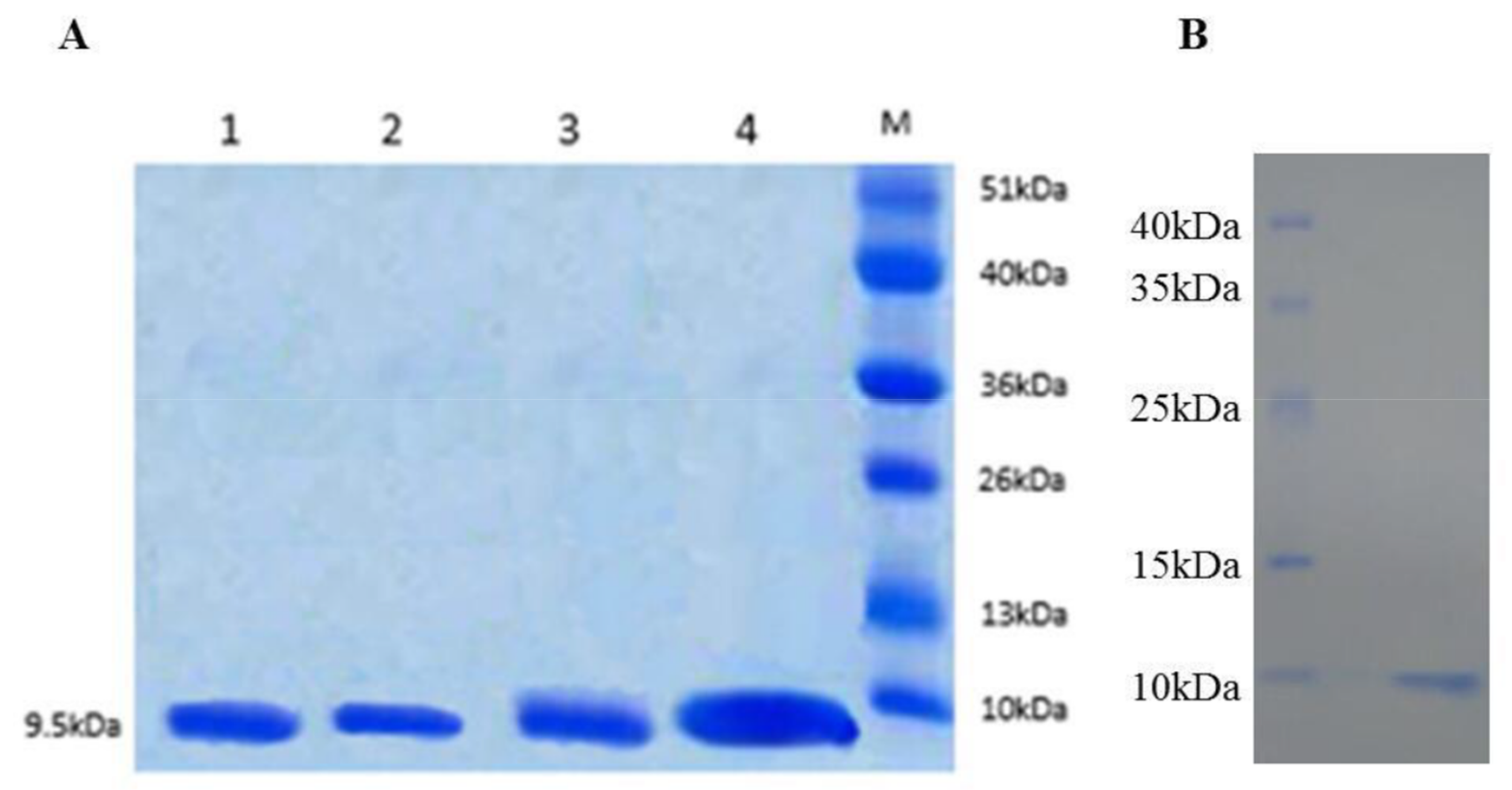
3.2. Expression and Purification of FtTI and FtTI Mutants
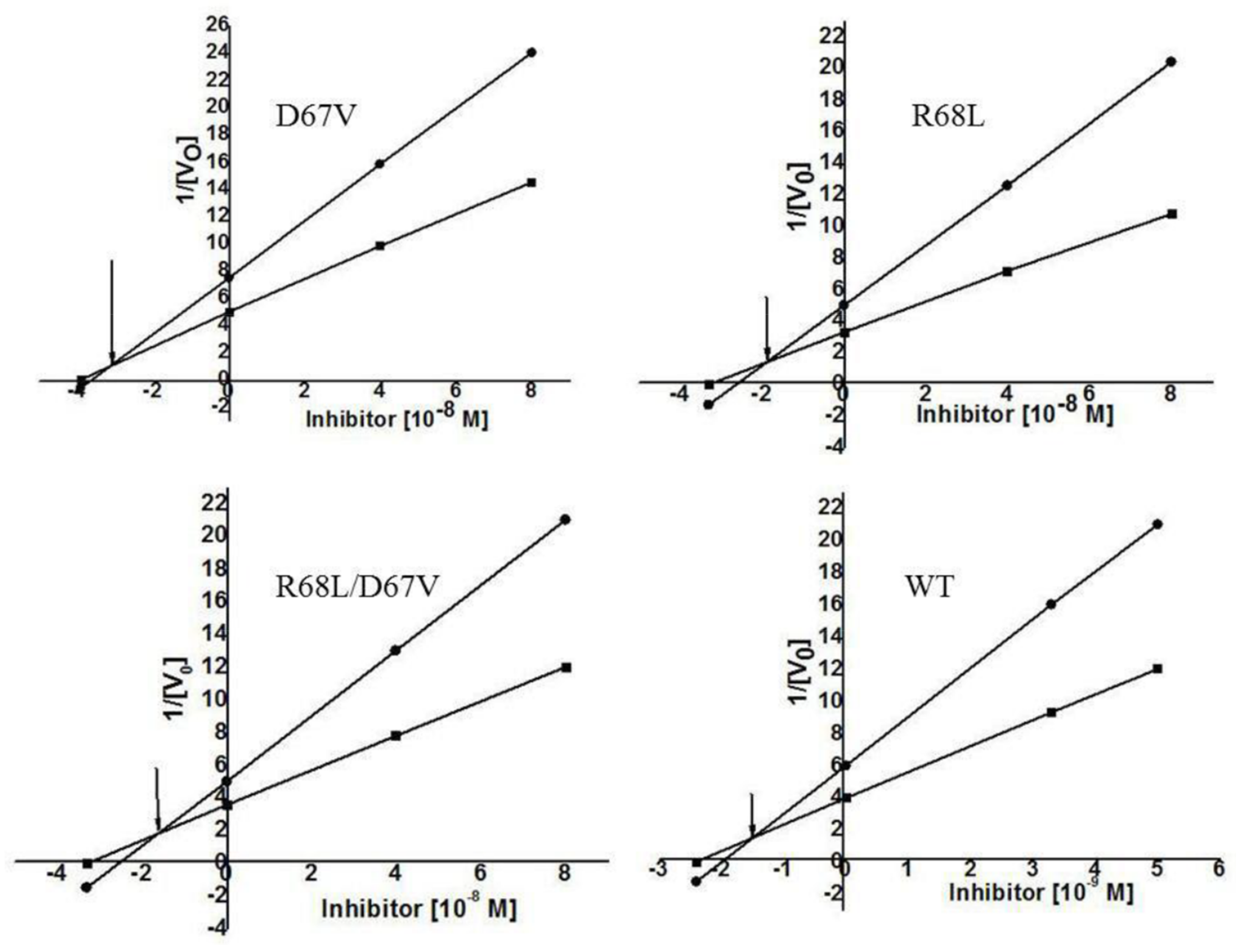
3.3. Analyses of the Characteristics of Recombinant FtTI and Its Mutants
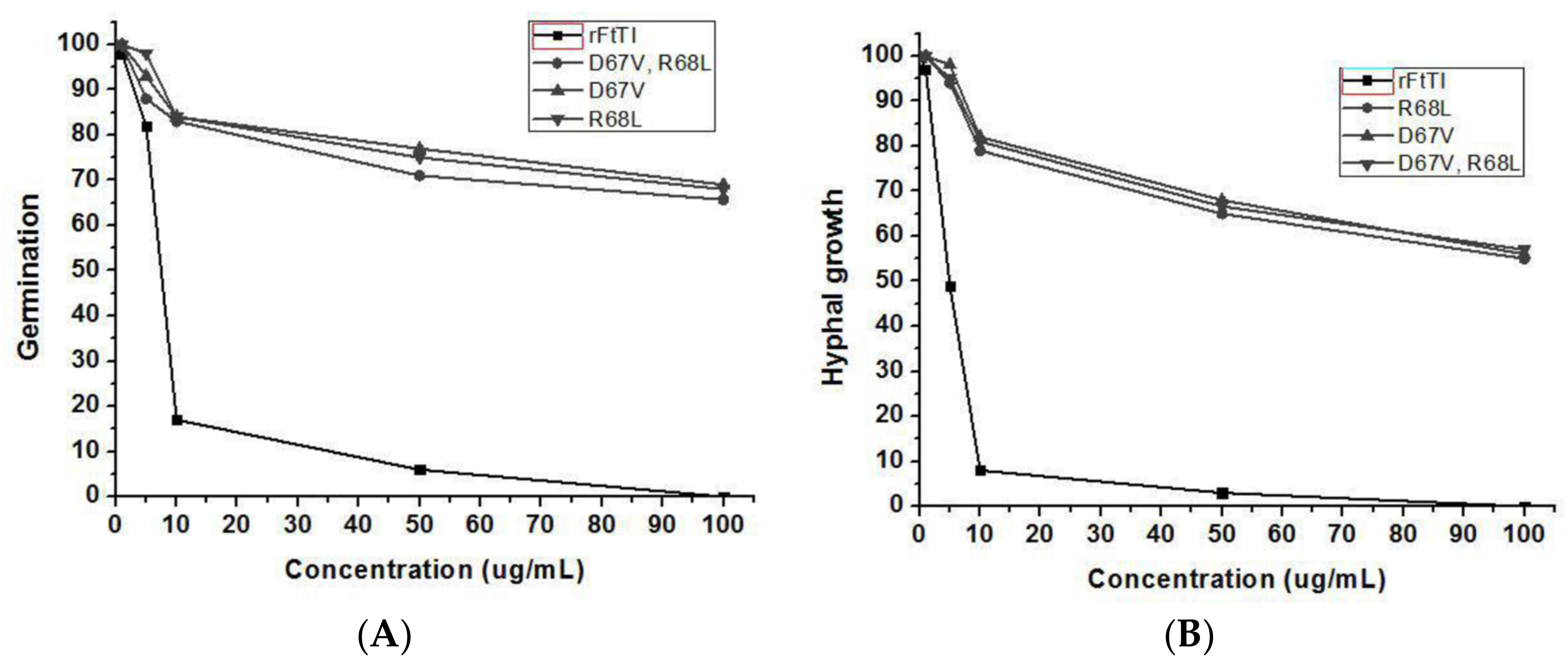
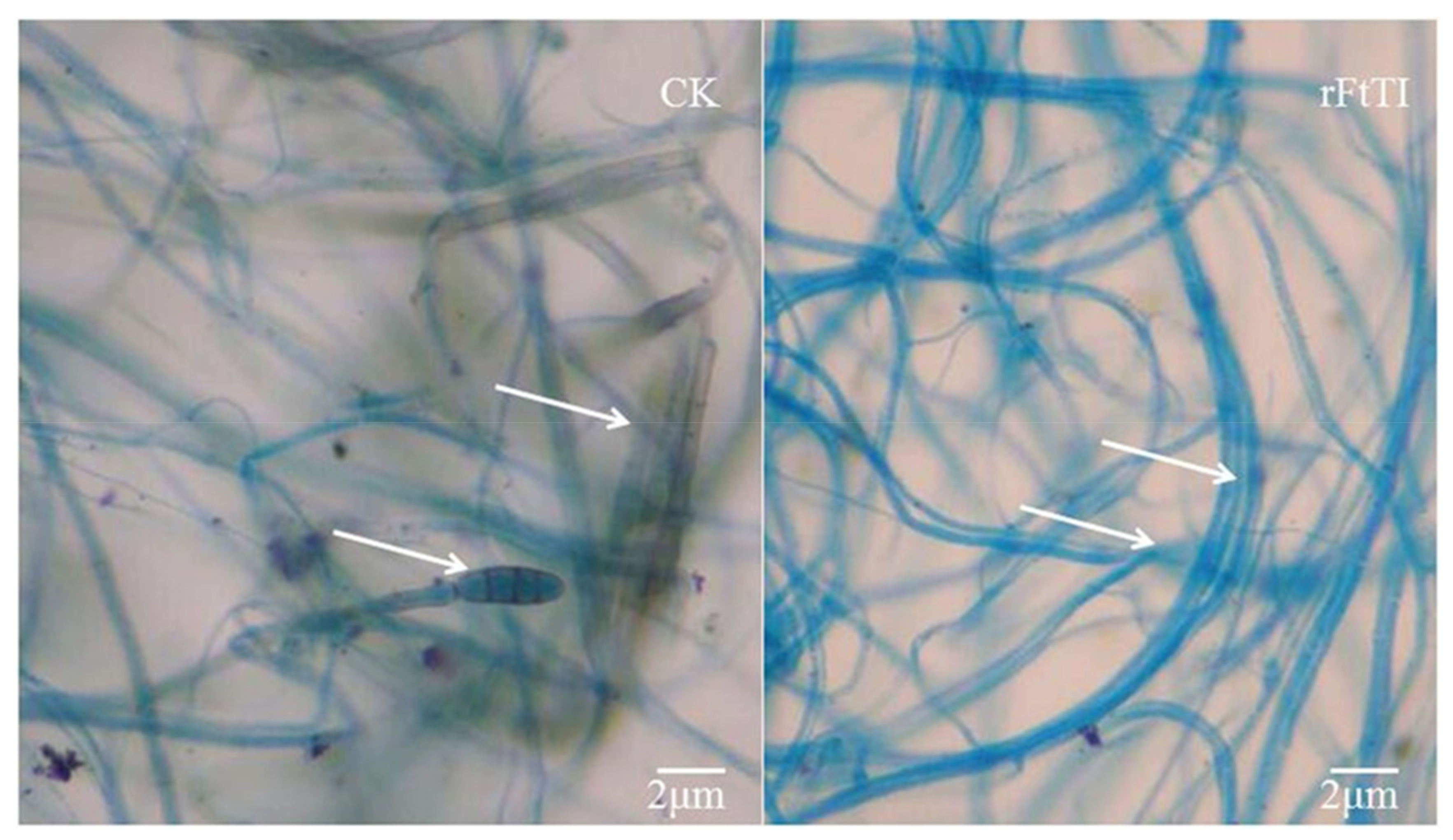
3.4. In-Vitro Antifungal Activity of Recombinant FtTI
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gaspar, Y.M.; McKenna, J.A.; McGinness, B.S.; Hinch, J.; Poon, S.; Connelly, A.A.; Anderson, M.A.; Heath, R.L. Field resistance to Fusarium oxysporum and Verticillium dahliae in transgenic cotton expressing the plant defensin NaD1. J. Exp. Bot. 2014, 65, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Tomazoni, E.Z.; Pauletti, G.F.; da Silva Ribeiro, R.T.; Moura, S.; Schwambach, J. In vitro and in vivo activity of essential oils extracted from Eucalyptus staigeriana, Eucalyptus globulus and Cinnamomum camphora against Alternaria solani Sorauer causing early blight in tomato. Sci. Hortic. 2017, 223, 72–77. [Google Scholar] [CrossRef]
- Bathige, S.D.N.K.; Umasuthan, N.; Godahewa, G.I.; Jayasinghe, J.D.H.E.; Whang, I.; Noh, J.K.; Lee, J. A homolog of kunitz-type serine protease inhibitor from rock bream, oplegnathus fasciatus: Molecular insights and transcriptional modulation in response to microbial and pamp stimulation, and tissue injury. Fish Shellfish Immunol. 2015, 46, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Qi, R.F.; Liu, Z.X.; Xu, S.Q.; Zhang, L.; Shao, X.X.; Chi, C.W. Small peptides derived from the Lys active fragment of the mung bean trypsin inhibitor are fully active against trypsin. FEBS J. 2010, 277, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Volpicella, M.; Leoni, C.; Arnesano, F.; Gallerani, R.; Ceci, L.R. Analysis by phage display selection and site-directed retromutagenesis of the Mustard Trypsin Inhibitor 2 reactive site. J. Plant Physiol. 2010, 167, 1507–1511. [Google Scholar] [CrossRef] [PubMed]
- Ekchaweng, K.; Khunjan, U.; Churngchow, N. Molecular cloning and characterization of three novel subtilisin-like serine protease genes from Hevea brasiliensis. Physiol. Mol. Plant Pathol. 2017, 97, 79–95. [Google Scholar] [CrossRef]
- Liu, D.; Li, Y.; Man, L.; Wu, Q.; Liu, S.; Liu, Y. Molecular cloning and characterization of PtrZPT2-1, a ZPT2 family gene encoding a Cys2/His2-type zinc finger protein from trifoliate orange (Poncirus trifoliata, (L.) Raf.) that enhances plant tolerance to multiple abiotic stresses. Plant Sci. Int. J. Exp. Plant Biol. 2017, 263, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, T.N.; Parveen, R.; Naz, H.; Haque, M.A.; Fatima, S. Biophysical insight into structure-function relation of Allium sativum Protease Inhibitor by thermal, chemical and pH-induced modulation using comprehensive spectroscopic analysis. Int. J. Biol. Macromol. 2017, 103, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Júnior, J.E.M.; Valadares, N.F.; Pereira, H.D.; Dyszy, F.H.; da Costa Filho, A.J.; Uchôa, A.F.; de Oliveira, A.S.; da Silveira Carvalho, C.P.; Grangeiro, T.B. Expression in Escherichia coli of cysteine protease inhibitors from cowpea (Vigna unguiculata): The crystal structure of a single-domain cystatin gives insights on its thermal and pH stability. Int. J. Biol. Macromol. 2017, 102, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.L.; Hwang, M.J.; Chen, Y.M.; Yeh, K.W. Site-directed mutagenesis evidence for a negatively charged trypsin inhibitory loop in sweet potato sporamin. FEBS Lett. 2001, 496, 134–138. [Google Scholar] [CrossRef]
- Yu, Q.; Ryan, E.M.; Allen, T.M.; Birren, B.W.; Henn, M.R.; Lennon, N.J. PriSM: A primer selection and matching tool for amplification and sequencing of viral genomes. Bioinformatics 2011, 27, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Boyce, R.; Chilana, P.; Rose, T.M. iCODEHOP: A new interactive program for designing COnsensus-DEgenerate Hybrid Oligonucleotide Primers from multiply aligned protein sequences. Nucleic Acids Res. 2009, 37, 222–228. [Google Scholar] [CrossRef] [PubMed]
- René, O.; Alix, J.H. Late steps of ribosome assembly in E. coli are sensitive to a severe heat stress but are assisted by the HSP70 chaperone machine. Nucleic Acids Res. 2011, 39, 1855–1867. [Google Scholar] [CrossRef] [PubMed]
- Brand, G.D.; Pires, D.A.T.; Furtado, J.R.; Cooper, A.; Freitas, S.M.; Bloch, C., Jr. Oligomerization affects the kinetics and thermodynamics of the interaction of a Bowman-Birk inhibitor with proteases. Arch. Biochem. Biophys. 2017, 618, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Gorrón, E.; Rodríguez, F.; Bernal, D.; Rodriguez-Rojas, L.M.; Bernal, A.; Restrepo, S.; Tohme, J. A new method for designing degenerate primers and its use in the identification of sequences in Brachiaria showing similarity to apomixis-associated genes. Bioinformatics 2010, 26, 2053–2054. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.A.; Hamouda, A.; Towner, K.J.; Amyes, S.G. Novel genetic context of multiple bla OXA-58 genes in Acinetobacter genospecies 3. J. Antimicrob. Chemother. 2010, 65, 1586–1588. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F. Chemical composition and health effects of Tartary buckwheat. Food Chem. 2016, 203, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.Z.; Feng, M.L.; Hao, X.L.; Zhao, Z.J.; Li, Y.Y.; Wang, Z.H. Anti-tumoral effects of a trypsin inhibitor derived from buckwheat in vitro and in vivo. Mol. Med. Rep. 2015, 12, 1777–1782. [Google Scholar] [CrossRef] [PubMed]
- Khadeeva, N.V.; Kochieva, E.Z.; Tcherednitchenko, M.Y.; Yakovleva, E.Y.; Sydoruk, K.V.; Bogush, V.G.; Dunaevsky, Y.E.; Belozersky, M.A. Use of buckwheat seed protease inhibitor gene for improvement of tobacco and potato plant resistance to biotic stress. Biochemistry (Mosc.) 2009, 74, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Park, S.S.; Ohba, H. Suppressive activity of protease inhibitors from buckwheat seeds against human T-acute lymphoblastic leukemia cell lines. Appl. Biochem. Biotechnol. 2004, 117, 65–74. [Google Scholar] [CrossRef]
- Wang, Z.; Li, S.; Ren, R.; Li, J.; Cui, X. Recombinant buckwheat trypsin inhibitor induces mitophagy by directly targeting mitochondria and causes mitochondrial dysfunction in Hep G2 cells. J. Agric. Food Chem. 2015, 63, 7795–7804. [Google Scholar] [CrossRef] [PubMed]
- Ashouri, S.; Abad, R.F.P.; Zihnioglu, F.; Kocadag, E. Extraction and purification of protease inhibitor (s) from seeds of Helianthus annuus with effects on Leptinotarsa decemlineata digestive cysteine protease. Biocatal. Agric. Biotechnol. 2017, 9, 113–119. [Google Scholar] [CrossRef]
- Qian, C.; Liang, D.; Liu, Y.; Wang, P.; Kausar, S.; Wei, G.; Zhu, B.; Wang, L.; Liu, C. Identification of a small pacifastin protease inhibitor from Nasonia vitripennis venom that inhibits humoral immunity of host (Musca domestica). Toxicon 2017, 131, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.J.; Chen, H.; Shao, J.R.; Wu, Q.; Han, X.Y. An antifungal peptide from Fagopyrum tataricum seeds. Peptides 2011, 32, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Yan, J.; Hou, S.; Chen, H.; Wu, Q.; Han, X. Expression and purification of the trypsin inhibitor from tartary buckwheat in Pichia pastoris and its novel toxic effect on Mamestra brassicae larvae. Mol. Biol. Rep. 2015, 42, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, N.; Banerjee, S.; Dutta, S.K. Cloning, expression and mutational studies of a trypsin inhibitor that retains activity even after cyanogen bromide digestion. Protein Expr. Purif. 2014, 96, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ren, J.; Nocadello, S.; Rice, A.J.; Ojeda, I.; Light, S.; Minasov, G.; Vargas, J.; Nagarathnam, D.; Anderson, W.F.; et al. Identification of novel small molecule inhibitors against NS2B/NS3 serine protease from Zika virus. Antivir. Res. 2017, 139, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, H.; Li, M.; Hu, X.Q.; Li, Y. Functional analysis of truncated and site-directed mutagenesis dextransucrases to produce different type dextrans. Enzyme Microb. Technol. 2017, 102, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Matsudaira, P. Sequence from picomole quantities of proteins electroblotted onto polyvinylidene difluoride membranes. J. Biol. Chem. 1987, 262, 10035–10038. [Google Scholar] [PubMed]
- Lee, S.C.; Hwang, I.S.; Choi, H.W.; Hwang, B.K. Involvement of the pepper antimicrobial protein CaAMP1 gene in broad spectrum disease resistance. Plant Physiol. 2008, 148, 1004–1020. [Google Scholar] [CrossRef] [PubMed]
- Gibaud, A.; Vogt, N.; Brison, O.; Debatisse, M.; Malfoy, B. Characterization at nucleotide resolution of the homogeneously staining region sites of insertion in two cancer cell lines. Nucleic Acids Res. 2013, 41, 8210–8219. [Google Scholar] [CrossRef] [PubMed]
- Ambadapadi, S.; Zheng, D.; Munaswamy-Ramanujam, G.; Lucas, A. Serine protease inhibitor (serpin) reactive center loop peptides as therapy for inflammatory vasculitis, hemorrhage and acute viral sepsis. Atherosclerosis 2015, 241, e209. [Google Scholar] [CrossRef]
- Bao, R.; Zhou, C.Z.; Jiang, C.; Lin, S.X.; Chi, C.W.; Chen, Y. The ternary structure of the double-headed arrowhead protease inhibitor API-A complexed with two trypsins reveals a novel reactive site conformation. J. Biol. Chem. 2009, 284, 26676–26684. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′→3′) | Purpose |
|---|---|---|
| CAAP1 | TGG CCN GAR CTN GTT GGA | Conservative area DNA |
| CAAP2 | RAC NCA YAC NCG GTC ACA | Conservative area DNA |
| SP1 | TCA CAA CGA AAG TCG AAA GTG C | 5′ Genome walking |
| SP2 | GGT GGC TAG AGT GGT TAA AGG AGG | 5′ Genome walking |
| SP3 | CTT CCC TTT GGT TCC AAC AAG C | 5′ Genome walking |
| SP4 | ACC CGG TCA CAA CGA AAG TC | 3′ Genome walking |
| SP5 | CAC AAC GAA AGT CGA AAG GTC | 3′ Genome walking |
| SP6 | GTG CGG CAG GTG GCT AGA GT | 3′ Genome walking |
| Fusion expression P1 | CCG GAA TTC CAT GTT AAT TTA CGC TAA AGT T | Fusion expression of gene |
| Fusion expression P2 | GAA TGC GGC CGC ACC GAC GGT TGG GGT | Fusion expression of gene |
| SMP1 | CGT TGT GAC CTG GTG CGT GTT TTA A | Site mutation, R68L |
| SMP2 | AAA GTG CGG CAG GTG GCT AGA GTG G | Site mutation, R68L |
| SMP3 | CTT TCG TTG TGT CCG GGT GCG TGT T | Site mutation, D67V |
| SMP4 | TCG AAA GTG CGG CAG GTG GCT AGA G | Site mutation, D67V |
| SMP5 | CTT TCG TTG TGT CCT GGT GCG TGT T | Site mutation, D67V, R68L |
| SMP6 | TCG AAA GTG CGG CAG GTG GCT AGA G | Site mutation, D67V, R68L |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, J.-j.; Tian, S.-j.; Yan, J.; Chen, H.; Xu, R.-h.; Cheng, J.-p. Molecular Cloning and Structure–Function Analysis of a Trypsin Inhibitor from Tartary Buckwheat and Its Application in Combating Phytopathogenic Fungi. Agronomy 2018, 8, 46. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8040046
Ruan J-j, Tian S-j, Yan J, Chen H, Xu R-h, Cheng J-p. Molecular Cloning and Structure–Function Analysis of a Trypsin Inhibitor from Tartary Buckwheat and Its Application in Combating Phytopathogenic Fungi. Agronomy. 2018; 8(4):46. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8040046
Chicago/Turabian StyleRuan, Jing-jun, Shan-jun Tian, Jun Yan, Hui Chen, Ru-hong Xu, and Jian-ping Cheng. 2018. "Molecular Cloning and Structure–Function Analysis of a Trypsin Inhibitor from Tartary Buckwheat and Its Application in Combating Phytopathogenic Fungi" Agronomy 8, no. 4: 46. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8040046