Silicon Compensates Phosphorus Deficit-Induced Growth Inhibition by Improving Photosynthetic Capacity, Antioxidant Potential, and Nutrient Homeostasis in Tomato


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Treatments
2.2. Determination of Biomass and Root Morphology
2.3. Determination of Photosynthetic Pigment Content and Leaf Gas Exchange Parameters
2.4. Determination of Membrane Lipid Peroxidation, Reactive Oxygen Species, and Antioxidant Enzyme Activity
2.5. Determination of Osmotic Potential and Osmotic Regulatory Substances
2.6. Determination of Element Content
2.7. Statistical Analysis
3. Results
3.1. Exogenous Silicon Improves Plant Growth Under Low Phosphorus Supply
3.2. Exogenous Silicon Increases Chlorophyll Content and Photosynthetic Capacity in Tomato Leaves Under Low Phosphorus Stress
3.3. Effects of Exogenous Silicon and Low Phosphorus on Lipid Peroxidation and ROS Accumulation
3.4. Exogenous Silicon Alleviated Low Phosphorus-Induced Oxidative Stress by Enhancing Antioxidant Enzyme Activity
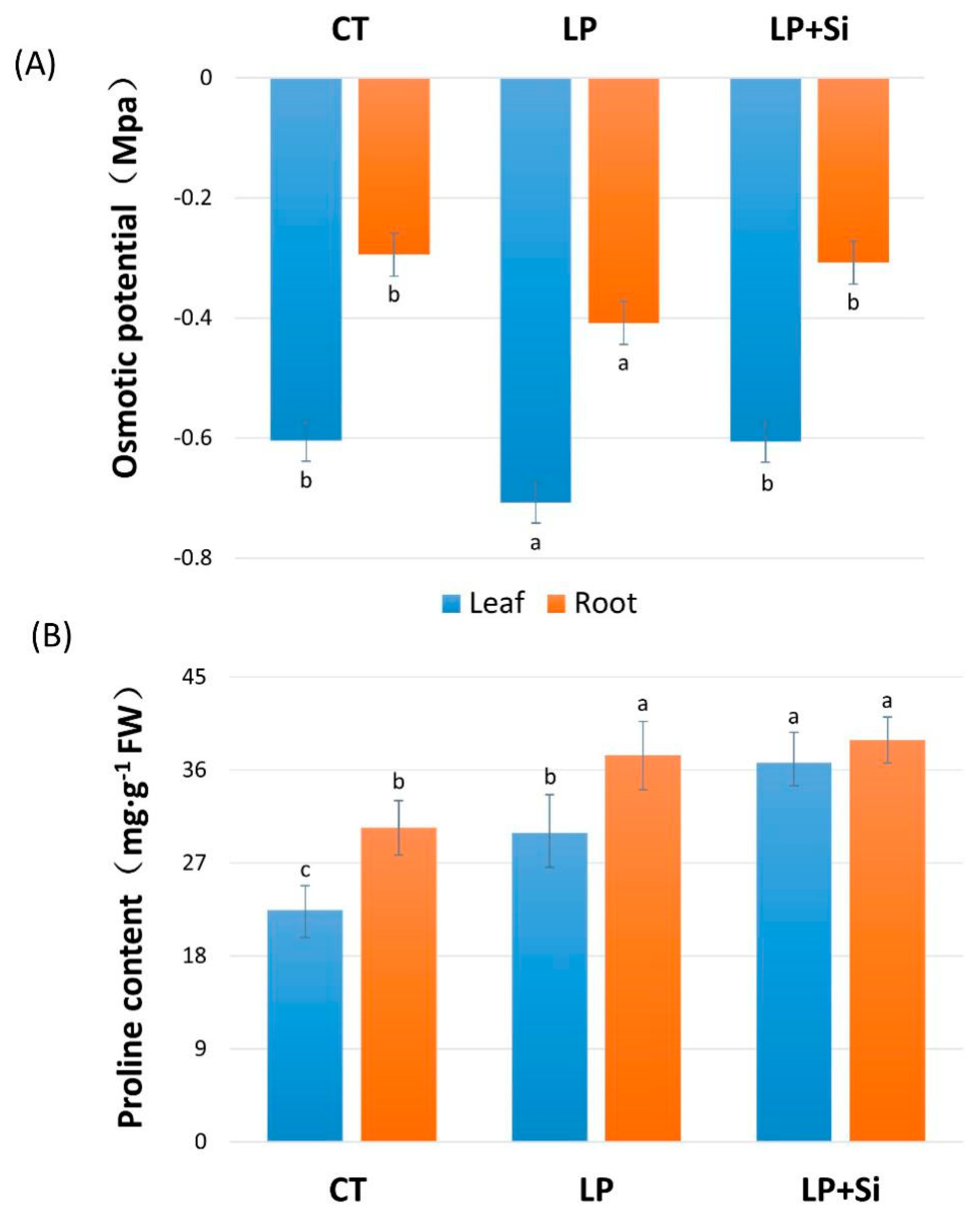
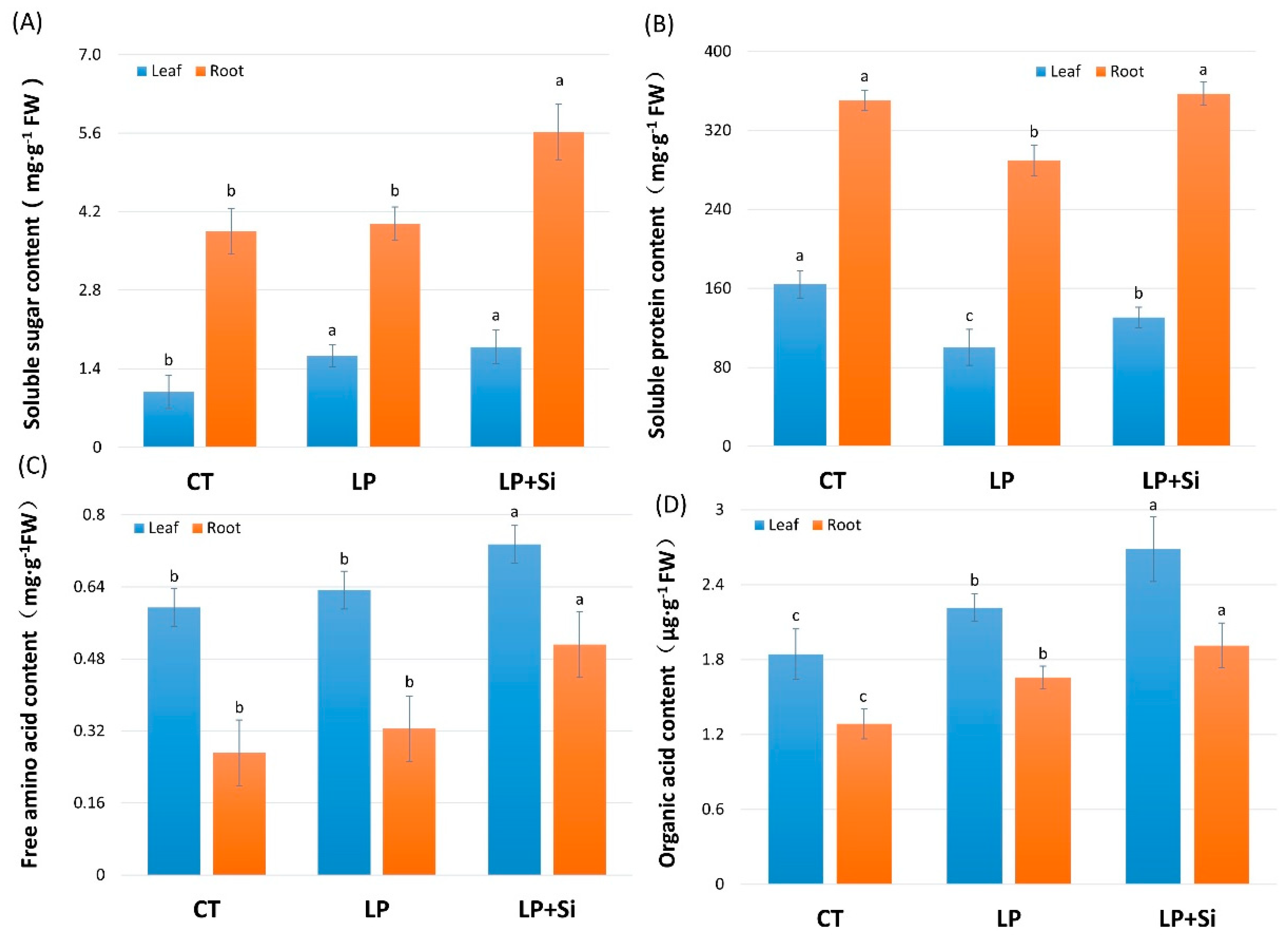
3.5. Effects of Exogenous Silicon on Osmotic Adjustment under Deficit Phosphorus Supply
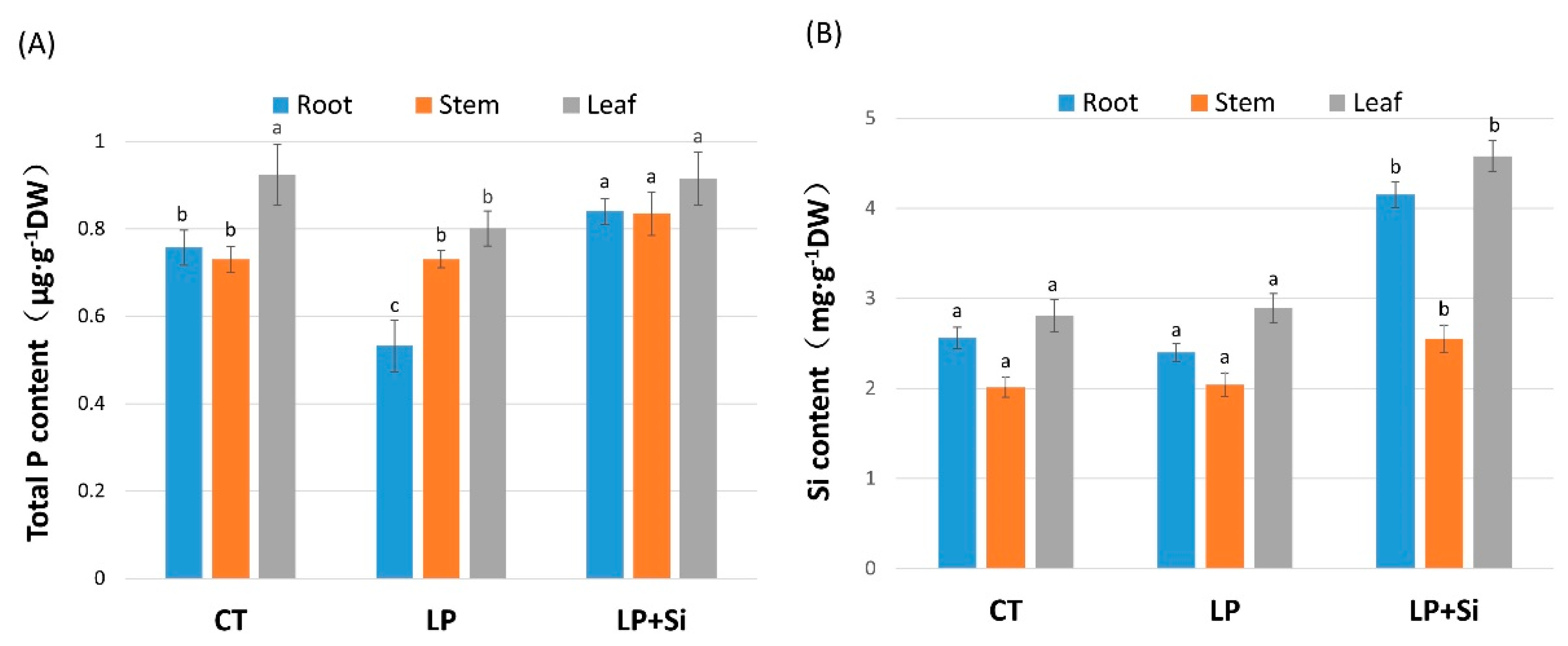
3.6. Effects of Exogenous Silicon and Low Phosphorus on Different Element Content in Tomato Leaves and Roots
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, Z.; Lynch, J.P.; Zhang, B.; Wang, Q. NPK deficiency modulates oxidative stress in plants. In Plant Macronutrient Use Efficiency; Academic Press: Cambridge, MA, USA, 2017; pp. 245–265. [Google Scholar]
- Hu, A.Y.; Che, J.; Shao, J.F.; Yokosho, K.; Zhao, X.Q.; Shen, R.F.; Ma, J.F. Silicon accumulated in the shoots results in down-regulation of phosphorus transporter gene expression and decrease of phosphorus uptake in rice. Plant Soil 2018, 423, 317–325. [Google Scholar] [CrossRef]
- Sulieman, S.; Tran, L.S. Phosphorus homeostasis in legume nodules as an adaptive strategy to phosphorus deficiency. Plant Sci. 2015, 239, 36–43. [Google Scholar] [CrossRef]
- Herrera-Estrella, L.; Lopez-Arredondo, D. Phosphorus: The Underrated Element for Feeding the World. Trends Plant Sci. 2016, 21, 461–463. [Google Scholar] [CrossRef]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2013, 112, 391–408. [Google Scholar] [CrossRef]
- Bhattacharya, A. Changing Environmental Condition and Phosphorus-Use Efficiency in Plants. In Changing Climate and Resource Use Efficiency in Plants; Academic Press: Cambridge, MA, USA, 2019; pp. 241–305. [Google Scholar]
- Hasan, M.K.; Ahammed, G.J.; Sun, S.; Li, M.; Yin, H.; Zhou, J. Melatonin inhibits cadmium translocation and enhances plant tolerance by regulating sulfur uptake and assimilation in Solanum lycopersicum L. J. Agric. Food Chem. 2019. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Xu, W.; Liu, A.; Chen, S. Endogenous melatonin deficiency aggravates high temperature-induced oxidative stress in Solanum lycopersicum L. Environ. Exp. Bot. 2019, 161, 303–311. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants (Basel) 2019. [Google Scholar] [CrossRef]
- Weih, M.; Westerbergh, A.; Lundquist, P.-O. Role of nutrient-efficient plants for improving crop yields: Bridging plant ecology, physiology, and molecular biology. In Plant Macronutrient Use Efficiency; Academic Press: Cambridge, MA, USA, 2017; pp. 31–44. [Google Scholar]
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants. Plants (Basel) 2019. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. A cooperative system of silicon transport in plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [CrossRef]
- Jang, S.W.; Kim, Y.; Khan, A.L.; Na, C.I.; Lee, I.J. Exogenous short-term silicon application regulates macro-nutrients, endogenous phytohormones, and protein expression in Oryza sativa L. BMC Plant Biol. 2018, 18, 4. [Google Scholar] [CrossRef]
- Chalmardi, Z.K.; Abdolzadeh, A.; Sadeghipour, H.R. Silicon nutrition potentiates the antioxidant metabolism of rice plants under iron toxicity. Acta Physiol. Plant. 2013, 36, 493–502. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Rohman, M.M.; Anee, T.I.; Huang, Y.; Fujita, M. Exogenous Silicon Protects Brassica napus Plants from Salinity-Induced Oxidative Stress Through the Modulation of AsA-GSH Pathway, Thiol-Dependent Antioxidant Enzymes and Glyoxalase Systems. Gesunde Pflanz. 2018, 70, 185–194. [Google Scholar] [CrossRef]
- Khan, E.; Gupta, M. Arsenic-silicon priming of rice (Oryza sativa L.) seeds influence mineral nutrient uptake and biochemical responses through modulation of Lsi-1, Lsi-2, Lsi-6 and nutrient transporter genes. Sci. Rep. 2018, 8, 10301. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Park, Y.G.; Kim, S.; Jeong, B.R. Foliar or Subirrigation Silicon Supply Mitigates High Temperature Stress in Strawberry by Maintaining Photosynthetic and Stress-Responsive Proteins. J. Plant Growth Regul. 2017, 36, 836–845. [Google Scholar] [CrossRef]
- Imtiaz, M.; Rizwan, M.S.; Mushtaq, M.A.; Ashraf, M.; Shahzad, S.M.; Yousaf, B.; Saeed, D.A.; Rizwan, M.; Nawaz, M.A.; Mehmood, S.; et al. Silicon occurrence, uptake, transport and mechanisms of heavy metals, minerals and salinity enhanced tolerance in plants with future prospects: A review. J. Environ. Manage. 2016, 183, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Chaiwong, N.; Prom, U.T.C.; Bouain, N.; Lacombe, B.; Rouached, H. Individual versus Combinatorial Effects of Silicon, Phosphate, and Iron Deficiency on the Growth of Lowland and Upland Rice Varieties. Int. J. Mol. Sci. 2018. [Google Scholar] [CrossRef]
- Pavlovic, J.; Samardzic, J.; Maksimovic, V.; Timotijevic, G.; Stevic, N.; Laursen, K.H.; Hansen, T.H.; Husted, S.; Schjoerring, J.K.; Liang, Y.; et al. Silicon alleviates iron deficiency in cucumber by promoting mobilization of iron in the root apoplast. New Phytol. 2013. [Google Scholar] [CrossRef]
- Gong, H.J.; Randall, D.P.; Flowers, T.J. Silicon deposition in the root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plant Cell Environ. 2006. [Google Scholar] [CrossRef]
- Yi, Z.; Li, S.; Liang, Y.; Zhao, H.; Hou, L.; Yu, S.; Ahammed, G.J. Effects of Exogenous Spermidine and Elevated CO2 on Physiological and Biochemical Changes in Tomato Plants Under Iso-osmotic Salt Stress. J. Plant Growth Regul. 2018, 37, 1222–1234. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Bailly, C.; Benamar, A.; Corbineau, F.; Come, D. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging. Physiol. Plant. 1996, 97, 104–110. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Van Montagu, M.; Inzé, D.; Van Camp, W. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef] [Green Version]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Ruan, Y.P.; Zhou, J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Brassinosteroid alleviates polychlorinated biphenyls-induced oxidative stress by enhancing antioxidant enzymes activity in tomato. Chemosphere 2013, 90, 2645–2653. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: I. Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Magnesium Deficiency and High Light Intensity Enhance Activities of Superoxide Dismutase, Ascorbate Peroxidase, and Glutathione Reductase in Bean Leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Pérez-López, U.; Robredo, A.; Lacuesta, M.; Mena-Petite, A.; Muñoz-Rueda, A. The impact of salt stress on the water status of barley plants is partially mitigated by elevated CO2. Environ. Exp. Bot. 2009, 66, 463–470. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Iwasaki, K.; Maier, P.; Fecht, M.; Horst, W.J. Leaf apoplastic silicon enhances manganese tolerance of cowpea (Vigna unguiculata). J. Plant Physiol. 2002, 159, 167–173. [Google Scholar] [CrossRef]
- Niu, Y.; Ahammed, G.J.; Tang, C.; Guo, L.; Yu, J. Physiological and Transcriptome Responses to Combinations of Elevated CO2 and Magnesium in Arabidopsis thaliana. PLoS ONE 2016, 11, e0149301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, G.; Li, Y.; Min, J.; Kronzucker, H.J.; Shi, W. Tomato plants ectopically expressing Arabidopsis GRF9 show enhanced resistance to phosphate deficiency and improved fruit production in the field. J. Plant Physiol. 2018, 226, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Farmer, E.E.; Mueller, M.J. ROS-Mediated Lipid Peroxidation and RES-Activated Signaling. Ann. Rev. Plant Biol. 2013, 64, 429–450. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A.; Iwasa, M.; Nakabayashi, R.; Kobayashi, M.; Nishizawa, T.; Okazaki, Y.; Saito, K.; Kusano, M. Effects of Combined Low Glutathione with Mild Oxidative and Low Phosphorus Stress on the Metabolism of Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 1464. [Google Scholar] [CrossRef]
- Maruyama, H.; Wasaki, J. Transgenic approaches for improving phosphorus use efficiency in plants. In Plant Macronutrient Use Efficiency; Academic Press: Cambridge, MA, USA, 2017; pp. 323–338. [Google Scholar]
- Cooke, J.; Leishman, M.R. Is plant ecology more siliceous than we realise? Trends Plant Sci. 2011, 16, 61–68. [Google Scholar] [CrossRef]
- Soo Won, J.; Hamayun, M.; Sohn, E.-Y.; Shin, D.-H.; Kim, K.-U.; Lee, I.-J. Studies on the effect of Silicon nutrition on plant growth, mineral contents and endogenous gibberellins of three rice cultivars. J. Crop Sci. Biotech. 2007, 10, 47–51. [Google Scholar]
- Yamaji, N.; Mitatni, N.; Ma, J.F. A Transporter Regulating Silicon Distribution in Rice Shoots. Plant Cell 2008, 20, 1381–1389. [Google Scholar] [CrossRef] [Green Version]
- Shimojima, M.; Madoka, Y.; Fujiwara, R.; Murakawa, M.; Yoshitake, Y.; Ikeda, K.; Koizumi, R.; Endo, K.; Ozaki, K.; Ohta, H. An engineered lipid remodeling system using a galactolipid synthase promoter during phosphate starvation enhances oil accumulation in plants. Front. Plant Sci. 2015, 6, 664. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, C.; Clement, M.; Thibaud, M.C.; Javot, H.; Chiarenza, S.; Delannoy, E.; Revol, J.; Soreau, P.; Balzergue, S.; Block, M.A.; et al. Identification of phosphatin, a drug alleviating phosphate starvation responses in Arabidopsis. Plant Physiol. 2014, 166, 1479–1491. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, Y.; Yao, H.; Wu, J.; Sun, H.; Gong, H. Silicon improves seed germination and alleviates oxidative stress of bud seedlings in tomato under water deficit stress. Plant Physiol. Biochem. 2014, 78, 27–36. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Andujar, C.; Ruiz-Lozano, J.M.; Dodd, I.C.; Albacete, A.; Perez-Alfocea, F. Hormonal and Nutritional Features in Contrasting Rootstock-mediated Tomato Growth under Low-phosphorus Nutrition. Front. Plant Sci. 2017, 8, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, N.; Chiba, Y.; Mitani-Ueno, N.; Feng Ma, J. Functional Characterization of a Silicon Transporter Gene Implicated in Silicon Distribution in Barley. Plant Physiol. 2012, 160, 1491–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, R.; Bélanger, R.R. Molecular evolution of aquaporins and silicon influx in plants. Funct. Ecology 2016, 30, 1277–1285. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Shoot Fresh Weight (g) | Root Fresh Weight (g) | Shoot Dry Weight (g) | Root Dry Weight (g) | Total Root Length (cm) | Total Root Surface Area (cm2) | Total Root Volume (cm3) | Average Root Diameter (mm) |
|---|---|---|---|---|---|---|---|---|
| CT | 28.7 ± 1.55a | 9.97 ± 1.70a | 2.35 ± 0.12a | 0.63 ± 0.06a | 2158 ± 173a | 495 ± 23a | 9.72 ± 1.02a | 0.71 ± 0.04a |
| LP | 14.2 ± 3.37b | 4.01 ± 1.44b | 1.05 ± 0.22b | 0.26 ± 0.09c | 1469 ± 305b | 255 ± 36c | 4.73 ± 0.12c | 0.60 ± 0.02b |
| LP + Si | 26.0 ± 1.43a | 8.05 ± 0.36a | 2.04 ± 0.11a | 0.48 ± 0.02b | 2306 ± 65a | 408 ± 45b | 7.64 ± 0.47b | 0.66 ± 0.03a |
| Treatment | Chl a | Chl b | Carotenoids | Chl (a + b) | Net Photosynthetic Rate (μmol CO2·m−2·S−1) | Stomatal Conductance (molH2O·m−2·s−1) | Intercellular CO2 Concentration (μmol CO2·mol−1 air) | Transpiration Rate (mmolH2O·m−2·s−1) | Water Use Efficiency (mmol CO2·mol−1·H2O) |
|---|---|---|---|---|---|---|---|---|---|
| CT | 1.94 ± 0.04a | 0.28 ± 0.01a | 0.51 ± 0.01a | 2.22 ± 0.04a | 24.1 ± 1.60a | 0.13 ± 0.04a | 246 ± 24.7c | 4.62 ± 0.32a | 2.97 ± 0.10a |
| LP | 1.83 ± 0.01b | 0.25 ± 0.01b | 0.46 ± 0.01b | 2.08 ± 0.03b | 13.8 ± 0.89b | 0.11 ± 0.03c | 293 ± 14.3a | 2.55 ± 0.34c | 2.48 ± 0.05c |
| LP + Si | 1.91 ± 0.03a | 0.30 ± 0.03a | 0.50 ± 0.02a | 2.21 ± 0.04a | 22.9 ± 1.72a | 0.12 ± 0.07b | 273 ± 14.9b | 3.21 ± 0.27b | 2.73 ± 0.06b |
| Antioxidant Enzymes | Tissue | CT | LP | LP + Si |
|---|---|---|---|---|
| SOD activity (U·g−1·FW·h−1) | Leaf | 35.5 ± 2.37a | 28.6 ± 1.79b | 36.6 ± 2.20a |
| Root | 14.8 ± 1.68a | 11.9 ± 1.09b | 15.25 ± 1.61a | |
| POD activity (U·g−1 FW) | Leaf | 2617 ± 85.0a | 2338 ± 78.4b | 2489 ± 83.2b |
| Root | 21,964 ± 311a | 14,452 ± 457c | 20,037 ± 731b | |
| CAT activity (U·g−1 FW) | Leaf | 88.1 ± 7.42a | 65.7 ± 5.16b | 90.1 ± 8.51a |
| Root | 98.4 ± 6.67a | 63.8 ± 7.74b | 100 ± 5.59a |
| Element | Tissue | CT | LP | LP + Si |
|---|---|---|---|---|
| K content (g·kg−1 DW) | Root | 36.0 ± 0.9a | 29.8 ± 1.0c | 32.9 ± 0.1b |
| Stem | 125 ± 6.4a | 113 ± 1.8b | 123 ± 3.4a | |
| Leaf | 39.4 ± 0.5a | 32.3 ± 1.2b | 32.5 ± 1.8b | |
| Na content (g·kg−1 DW) | Root | 17.1 ± 0.5a | 5.4 ± 0.3c | 7.1 ± 1.3b |
| Stem | 11.6 ± 0.7a | 3.7 ± 0.4c | 7.6 ± 0.3b | |
| Leaf | 2.1 ± 0.4a | 1.4 ± 0.1b | 2.2 ± 0.4a | |
| Ca content (g·kg−1 DW) | Root | 12.2 ± 2.3a | 4.0 ± 0.1c | 5.0 ± 0.7b |
| Stem | 11.1 ± 2.6a | 3.5 ± 0.6b | 14.0 ± 0.7a | |
| Leaf | 15.8 ± 0.9a | 11.7 ± 0.8b | 12.8 ± 2.5ab | |
| Mg content (g·kg−1 DW) | Root | 5.9 ± 0.6a | 4.4 ± 0.4c | 5.1 ± 0.1b |
| Stem | 7.2 ± 1.3a | 5.6 ± 1.1b | 6.5 ± 0.1b | |
| Leaf | 8.7 ± 0.9a | 6.4 ± 0.3c | 7.2 ± 0.3b | |
| Fe content (mg·kg−1 DW) | Root | 4562 ± 3571a | 3171 ± 206b | 4292 ± 355a |
| Stem | 632 ± 53.4a | 231 ± 60.4b | 584 ± 72.5a | |
| Leaf | 459 ± 18.6a | 381 ± 19.0b | 434 ± 18.8a | |
| Mn content (mg·kg−1 DW) | Root | 936 ± 17.2a | 615 ± 42.8b | 862 ± 13.3a |
| Stem | 63.2 ± 2.7a | 55.0 ± 3.0c | 58.7 ± 2.1b | |
| Leaf | 73.6 ± 2.5a | 58.3 ± 3.9b | 60.4 ± 1.6b | |
| Zn content (mg·kg−1 DW) | Root | 87.2 ± 1.4a | 71.8 ± 6.4b | 76.5 ± 3.3b |
| Stem | 34.5 ± 2.1a | 21.3 ± 0.8b | 32.8 ± 3.1a | |
| Leaf | 21.9 ± 4.5a | 15.5 ± 0.5b | 13.3 ± 0.5c | |
| Cu content (mg·kg−1 DW) | Root | 87.9 ± 2.4a | 79.9 ± 3.6a | 82.6 ± 6.2a |
| Stem | 31.2 ± 7.5a | 16.7 ± 1.8b | 26.7 ± 7.4a | |
| Leaf | 27.9 ± 3.0a | 25.3 ± 0.01a | 26.1 ± 2.9a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liang, Y.; Zhao, X.; Jin, X.; Hou, L.; Shi, Y.; Ahammed, G.J. Silicon Compensates Phosphorus Deficit-Induced Growth Inhibition by Improving Photosynthetic Capacity, Antioxidant Potential, and Nutrient Homeostasis in Tomato. Agronomy 2019, 9, 733. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9110733
Zhang Y, Liang Y, Zhao X, Jin X, Hou L, Shi Y, Ahammed GJ. Silicon Compensates Phosphorus Deficit-Induced Growth Inhibition by Improving Photosynthetic Capacity, Antioxidant Potential, and Nutrient Homeostasis in Tomato. Agronomy. 2019; 9(11):733. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9110733
Chicago/Turabian StyleZhang, Yi, Ying Liang, Xin Zhao, Xiu Jin, Leiping Hou, Yu Shi, and Golam Jalal Ahammed. 2019. "Silicon Compensates Phosphorus Deficit-Induced Growth Inhibition by Improving Photosynthetic Capacity, Antioxidant Potential, and Nutrient Homeostasis in Tomato" Agronomy 9, no. 11: 733. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9110733