Visual Symptoms, Vegetative Growth, and Mineral Concentration in Fig Tree (Ficus carica L.) Under Macronutrient Deficiencies

Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of the Experiment
2.2. Treatments
2.3. Visual Symptoms of Macronutrient Deficiency
2.4. Vegetative Growth Evaluation
2.5. Minerals Determination
2.6. Statistical Analysis
3. Results and Discussion
3.1. Visual Symptoms of Macronutrient Deficiencies
3.1.1. N Deficiency
3.1.2. P Deficiency
3.1.3. K Deficiency
3.1.4. Ca Deficiency
3.1.5. Mg Deficiency
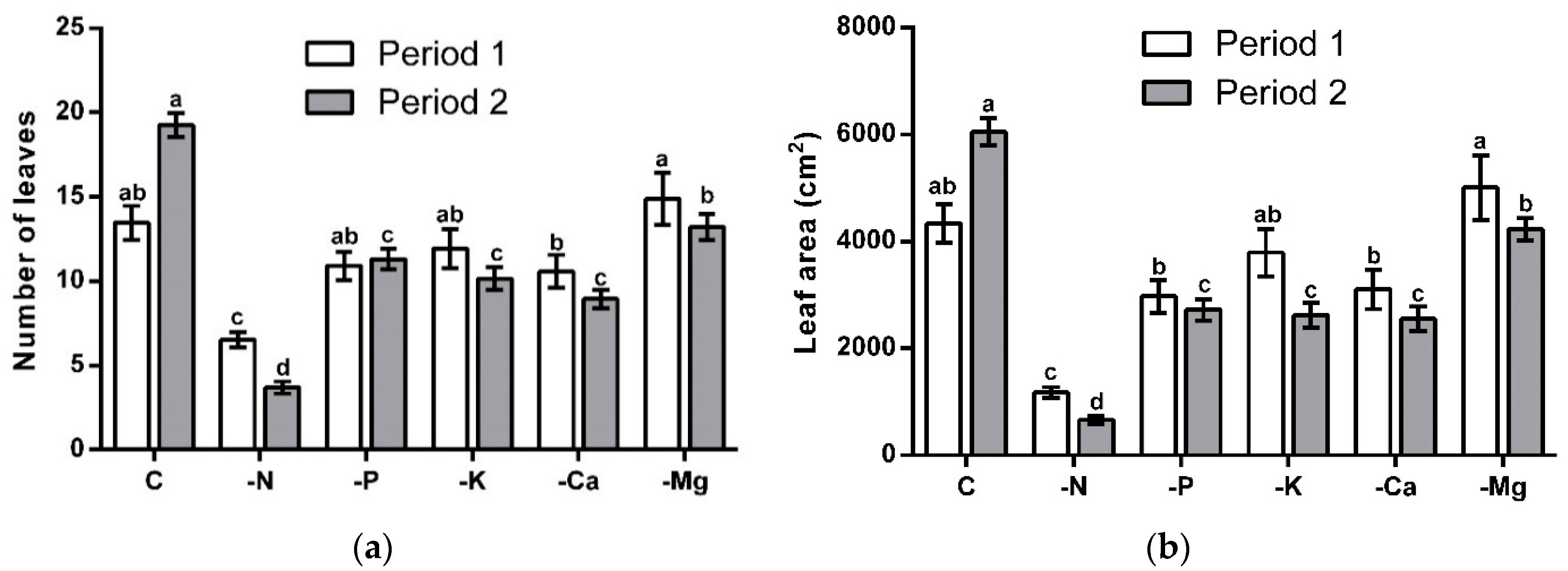
3.2. Vegetative Growth
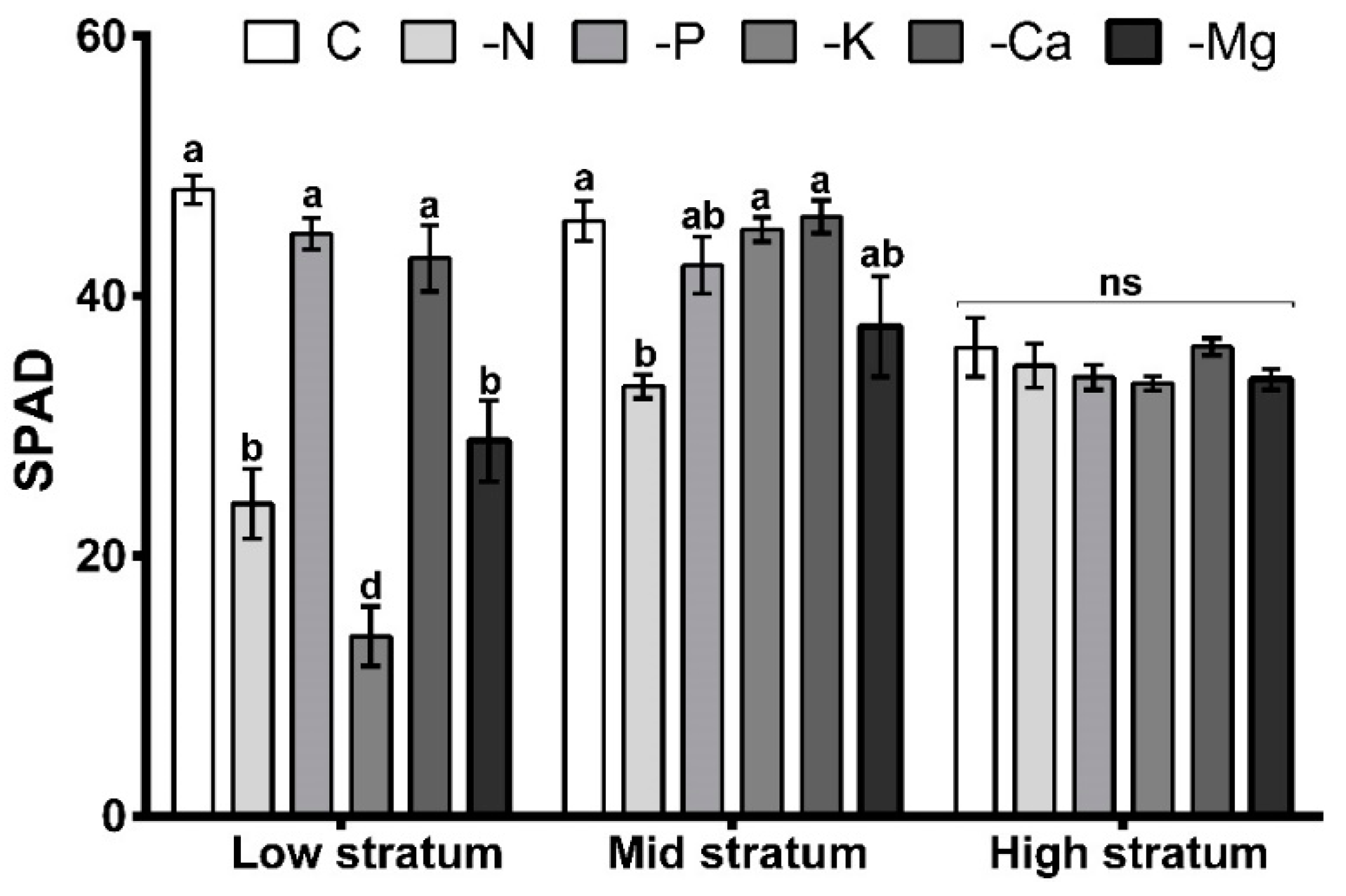
3.3. Mineral Concentration
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agricultural Research Service, United States Department of Agriculture (USDA). Food Composition Databases. Available online: https://ndb.nal.usda.gov/ndb/search/list (accessed on 10 September 2019).
- Deepa, P.; Sowndhararajan, K.; Kim, S.; Park, S.J. A role of Ficus species in the management of diabetes mellitus: A review. J. Ethnopharmacol. 2018, 215, 210–232. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization (FAO). Statistical Yearbook. Agricultural Production. Available online: http://www.fao.org/faostat/es/#data/QC (accessed on 12 September 2019).
- Servicio de Información Agroalimentaria y Pesquera (SIAP). Available online: https://www.gob.mx/siap/acciones-y-programas/produccion-agricola-33119 (accessed on 10 September 2019).
- Mendoza, C.V.M.; Vargas, C.J.M.; Calderón, Z.G.; Mendoza, C.M.C.; Santacruz, V.A. Intensive production systems of Fig (Ficus carica L.) under greenhouse conditions. Exp. Agric. 2017, 3, 339–350. [Google Scholar] [CrossRef]
- Kathpalia, R.; Bhatla, S.C. Plant Mineral Nutrition. In Plant Physiology, Development and Metabolism; Bhatla, S.C., Lal, M.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 37–81. [Google Scholar] [CrossRef]
- Alves, F.R.; Montes, N.B.M.; Almeida, H.; Mello, P.R. Growth and nutritional disorders of coffee cultivated in nutrient solutions with suppressed macronutrients. J. Plant Nutr. 2016, 11, 1578–1588. [Google Scholar] [CrossRef]
- Alves, F.R.; Montes, N.B.M.; Almeida, H.; Mello, P.R. Growth and nutritional disorders of eggplant cultivated in nutrients solutions with suppressed macronutrients. J. Plant Nutr. 2015, 7, 1097–1109. [Google Scholar] [CrossRef]
- Chen, L.; Zeng, J.; Xu, D.; Zhao, Z.; Guo, J. Macronutrient deficiency symptoms in Betula alnoides seedlings. Jour. Trop. For. Sci. 2010, 4, 403–413. [Google Scholar]
- Dussán, S.L.; Villegas, D.A.; Miranda, D. Efecto de la deficiencia de N, P, K, Mg, Ca y B sobre la acumulación y distribución de la masa seca en plantas de guayaba (Psidium guajava L.) var. ICA Palmira II en fase de vivero. Rev. Colom. Cienc. Hortic. 2016, 1, 40–52. [Google Scholar] [CrossRef]
- Mota, R.M.J.; Rocha, P.L.G. Physical and Quality Characteristics of Jupi Pineapple Fruits on Macronutrient and Boron Deficiency. Nat. Res. 2014, 8, 359–366. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadística y Geografía (INEGI). Mapa Digital de México V6.3. Available online: http://gaia.inegi.org.mx/mdm6/?coordinates= (accessed on 10 September 2019).
- Servicio Meteorológico Nacional. Available online: https://smn.conagua.gob.mx/es/climatologia/informacion-climatologica/normales-climatologicas-por-estado (accessed on 12 September 2019).
- Papadopoulos, A.P.; Bar-Tal, A.; Silber, A.; Saha, U.K.; Raviv, K. Inorganic and synthetic organic components of soilless culture and potting mixes. In Soilless Culture, Theory and Practices; Raviv, M., Lieth, J.H., Eds.; Elsevier: Amsterdam, NL, USA, 2008; pp. 505–543. ISBN 978-0-444-52975-6. [Google Scholar]
- Sevil, K.S.; Ertan, E.; Seferoglu, S. Effect of different nutrient solution formulations on morphological and biochemical characteristics of nursery fig trees grown in substrate culture. Sci. Hort. 2007, 2, 20–27. [Google Scholar] [CrossRef]
- Muñoz, R.J.J. Manejo del cultivo de tomate en invernadero. In Manual de Producción de Tomate en Invernadero; Castellanos, J.Z., Ed.; Intagri: Celaya/México City, Mexico, 2009; pp. 45–92. ISBN 978-607-95302-0-4. [Google Scholar]
- Giaccone, M.; Pannico, A.; Scognamiglio, C.; Rivera, M.; Cirillo, C.; Rouphael, Y.; Pascale, S.; Basile, B. Regression model for leaf area estimation in Ficus carica L. Acta Hort. 2017, 1163, 163–168. [Google Scholar] [CrossRef]
- Alcántar, G.G.; Sandoval, V.M. Handbook Chemical Analysis of Vegetal Tissue, Especial Publication No. 10; Mexican Society of Soil Science: Chapingo, Mexico, 1999. (In Spanish) [Google Scholar]
- Bremner, J.M. Total nitrogen. In Methods for Soil Analysis. Part 2. Chemical and Microbiological Properties; Norman, A.G., Ed.; American Society of Agronomy Inc.: Madison, WI, USA, 1965; pp. 1149–1178. [Google Scholar]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Skrumsager, I.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, H., Ed.; Academic Press: London, UK, 2012; pp. 135–189. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates: Sunderland, MA, USA, 2014; ISBN 978-1-60535-255-8. [Google Scholar]
- Mengel, K.; Kirkby, E.A.; Kosegarten, H.; Appel, T. Principles of Plant Nutrition, 5th ed.; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar] [CrossRef]
- Karthika, K.S.; Rashmi, I.; Parvathi, M.S. Biological functions, uptake and transport of essential nutrients in relation to plant growth. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–49. [Google Scholar] [CrossRef]
- Alcántar, G.G.; Trejo, T.L.I.; Fernández, P.L.; Rodríguez, M.M. Elementos esenciales. In Nutrición de Cultivos, 2nd ed.; Alcántar, G.G.L.I., Trejo, T.Y.F.C., Gómez, M., Eds.; Colegio de Postgraduados: México City, Mexico, 2016; p. 2355. ISBN 978-607-715-324-5. [Google Scholar]
- Romheld, V. Diagnosis of deficiency and toxicity of nutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed; Marschner, H., Ed.; Academic Press: London, UK, 2012; pp. 299–312. [Google Scholar] [CrossRef]
- Tanoi, K.; Kobayashi, N. Leaf senescence by magnesium deficiency. Plants 2015, 4, 756–772. [Google Scholar] [CrossRef]
- Engels, C.; Kirkby, E.; White, P. Mineral Nutrition, Yield and Source-Sink Relationships. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, H., Ed.; Academic Press: London, UK, 2012; pp. 85–133. [Google Scholar] [CrossRef]
- Soares, S.M.P.; Mendoca, F.M.S.; Cesar, S.P.; Cordeiro, A.J.C.; Silva, J.T. Capsicum annuum var. Annuum under macronutrients and boron deficiencies: Leaf content and visual symptomps. J. Plant Nutr. 2019, 5, 417–427. [Google Scholar] [CrossRef]
- White, P.J. Ion uptake mechanisms of individual cells and roots: Short-distance transport. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, H., Ed.; Academic Press: London, UK, 2012; pp. 7–47. [Google Scholar] [CrossRef]
- Farhat, N.; Elkhouni, A.; Zorrig, W.; Smaoui, A.; Abdelly, C.; Rabhi, M. Effects of magnesium deficiency on photosynthesis and carbohydrate partitioning. Acta Physiol. Plant. 2016, 145. [Google Scholar] [CrossRef]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop. J. 2016, 2, 83–91. [Google Scholar] [CrossRef]
- Lima, T.C.; Freitas, M.S.M.; Mendonça, L.V.P.; Gonçalves, Y.D.S.; Pereira, J.A.P.; Lima, K.F. Visual symptoms of macronutrient and boron deficiency in Cymbopogon citratus. J. Plant Nutr. 2019, 13, 1430–1438. [Google Scholar] [CrossRef]
- Rhodes, R.; Miles, N.; Charles, H.J. Interactions between potassium, calcium and magnesium in sugarcane grown on two contrasting soils in South Africa. Field Crop. Res. 2018, 225, 1–11. [Google Scholar] [CrossRef]
- Veígas, I.D.J.M.; Sousa, G.O.D.; Silva, A.F.D.; Carvalho, J.G.D.; Lima, M.M. Mineral composition and visual symptoms of nutrients deficiencies in long pepper plants (Piper hispidinervum C. DC.). Acta Amazon. 2013, 1, 43–50. [Google Scholar] [CrossRef]
- Veígas, I.J.M.; Silva, L.A.K.; Silva, R.M.F.; Morais, C.R.L.; Capucho, F.D.A.; Oliveira, N.C.F.; Oliveira, C.H.E.; Silva, G.E.M.; Ruffeil, A.G.A.; Poliana, S.S. Visual symptoms and growth parameters linked to deficiency of macronutrients in young Swietenia macrophylla plants. J. Food Agric. Environ. 2012, 1, 937–940. [Google Scholar] [CrossRef]
- Ribeiro, C.A.C.; Gomes, W.L.; Loges, V.; Arruda, C.M.F.; Souza, A.F.A. Macronutrients deficiency in Heliconia psittacorum × Heliconia spathocircinata ‘Golden Torch’. Rev. Ciência Agron. 2015, 2, 258–265. [Google Scholar] [CrossRef]
- Rodríguez, M.M.N.; Tejo, T.L.I.; Alcántar, G.G. Acceso, absorción y transporte nutrimental. In Nutrición de Cultivos, 2nd ed.; Alcántar, G.G.L.I., Trejo, T.Y.F.C., Gómez, M., Eds.; Colegio de Postgraduados: México City, Mexico, 2016; pp. 153–182. ISBN 978-607-715-324-5. [Google Scholar]
- Brown, P. Seasonal variations in fig (Ficus carica L.) leaf nutrient concentrations. HortScience 1994, 8, 871–873. [Google Scholar] [CrossRef]
- Moreno, D.A.; Pulgar, G.; Víllora, G.; Romero, L. Nutritional diagnosis of fig tree leaves. J. Plant Nutr. 1998, 12, 2579–2588. [Google Scholar] [CrossRef]
- Sun, R.; Sun, L.; Jia, M. Analysis of psoralen and mineral elements in the leaves of different fig (Ficus carica L.) cultivars. Acta Hort. 2017, 1173, 293–296. [Google Scholar] [CrossRef]
- Mendoza, C.V.M.; Pineda, P.J.; Vargas, C.J.M.; Hernández, A.E. Nutrition of fig (Ficus carica L.) under hydroponics and greenhouse conditions. J. Plant Nutr. 2019, 11, 1350–1365. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Macronutrients | Micronutrients | ||
|---|---|---|---|
| Element | mmol L−1 | Element | mg L−1 |
| N | 12 | Fe | 2.8 |
| P | 1.3 | Mn | 0.55 |
| K | 4 | B | 0.54 |
| Ca | 4 | Cu | 0.064 |
| Mg | 1.5 | Zn | 0.065 |
| S | 1.5 | Mo | 0.048 |
| Treatment | S.E. (cm week−1) | P.H. (cm) | S.D. (mm) | I.L. (cm) | Dry Weight | ||
|---|---|---|---|---|---|---|---|
| Roots | Stem | Leaves | |||||
| Control | 6.47 a | 102.4 ab | 13.72 b | 3.34 a | 121.1 a | 114.1 a | 87.46 a |
| -N | 1.15 b | 35.08 d | 8.40 d | 1.39 c | 27.47 c | 23.03 b | 10.93 c |
| -P | 4.05 ab | 74.88 c | 11.81 c | 2.64 ab | 66.27 bc | 70.00 ab | 53.56 b |
| -K | 5.70 a | 96.84 b | 12.58 c | 3.19 ab | 44.37 bc | 109.9 a | 55.20 b |
| -Ca | 3.28 ab | 69.31 c | 11.63 c | 2.30 bc | 54.47 bc | 66.93 ab | 36.10 bc |
| -Mg | 5.64 a | 111.0 a | 14.94 a | 3.37 a | 81.14 ab | 70.26 ab | 55.86 b |
| Treatment | Concentration (%) | ||||
|---|---|---|---|---|---|
| N | P | K | Ca | Mg | |
| Leaf | |||||
| Control | 2.54 a | 0.16 c | 1.7 a | 2.90 a | 0.48 b |
| -N | 1.68 b | 0.31 ab | 1.6 a | 2.05 b | 0.32 bc |
| -P | 2.98 a | 0.22 bc | 1.38 a | 2.17 b | 0.47 b |
| -K | 2.77 a | 0.32 a | 0.22 b | 2.33 ab | 0.99 a |
| -Ca | 2.95 a | 0.21 bc | 1.47 a | 2.01 b | 0.49 b |
| -Mg | 2.67 a | 0.22 bc | 1.62 a | 2.18 b | 0.26 c |
| Stem | |||||
| Control | 1.65 b | 0.22 a | 0.65 bc | 0.65 ab | 0.25 ab |
| -N | 0.82 c | 0.27 a | 0.54 cd | 0.68 ab | 0.19 c |
| -P | 1.30 b | 0.13 b | 0.70 b | 0.71 ab | 0.22 bc |
| -K | 1.52 b | 0.29 a | 0.26 e | 0.77 a | 0.23 abc |
| -Ca | 2.18 a | 0.28 a | 0.46 d | 0.56 b | 0.28 a |
| -Mg | 1.48 b | 0.26 a | 0.84 a | 0.78 a | 0.20 bc |
| Root | |||||
| Control | 2.46 ns | 0.51 b | 0.97 ab | 1.43 a | 0.38 a |
| -N | 1.92 ns | 0.49 b | 0.74 cd | 1.14 ab | 0.25 b |
| -P | 2.26 ns | 0.23 c | 0.88 bc | 1.41 a | 0.39 a |
| -K | 2.24 ns | 0.52 b | 0.28 e | 1.17 ab | 0.28 b |
| -Ca | 2.44 ns | 0.58 b | 0.68 d | 0.65 b | 0.29 b |
| -Mg | 2.06 ns | 0.86 a | 1.12 a | 1.35 a | 0.22 b |
| Treatment | Concentration (mg kg−1 DW) | ||||
|---|---|---|---|---|---|
| Fe | B | Mn | Zn | Cu | |
| Leaf | |||||
| Control | 914.2 a | 151.5 ab | 148.9 b | 45.15 a | 6.16 ab |
| -N | 456.5 b | 143.7 ab | 92.59 b | 28.93 b | 4.47 c |
| -P | 600.6 ab | 105.7 b | 107.9 b | 38.15 ab | 6.85 a |
| -K | 497.3 b | 111.7 b | 253.4 a | 34.51 ab | 5.17 bc |
| -Ca | 660.0 ab | 175.4 a | 275.5 a | 43.84 a | 4.80 bc |
| -Mg | 655.7 a | 126.6 ab | 130.3 b | 36.71 ab | 4.77 bc |
| Stem | |||||
| Control | 118.1 b | 23.66 ns | 31.85 bc | 9.66 ab | 2.84 cd |
| -N | 112.9 b | 23.85 ns | 14.04 d | 8.71 b | 4.47 a |
| -P | 96.15 b | 25.55 ns | 16.32 cd | 12.37 a | 3.73 ab |
| -K | 205.4 a | 25.39 ns | 65.81 a | 10.84 ab | 4.08 a |
| -Ca | 156.6 ab | 25.58 ns | 80.50 a | 11.75 ab | 2.30 d |
| -Mg | 157.9 ab | 29.95 ns | 34.96 b | 9.66 ab | 3.15 bc |
| Root | |||||
| Control | 1135.1 ns | 40.70 a | 103.4 cd | 30.09 ns | 18.78 bc |
| -N | 1377.8 ns | 21.95 b | 56.86 cd | 44.03 ns | 33.45 a |
| -P | 1061.1 ns | 38.46 a | 39.86 d | 42.83 ns | 27.36 ab |
| -K | 1434.3 ns | 29.61 ab | 221.8 a | 39.47 ns | 28.26 ab |
| -Ca | 1263.4 ns | 29.88 ab | 191.4 ab | 47.88 ns | 20.33 bc |
| -Mg | 890.3 ns | 30.58 ab | 131.9 bc | 34.97 ns | 13.73 c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garza-Alonso, C.A.; Olivares-Sáenz, E.; Gutiérrez-Díez, A.; Vázquez-Alvarado, R.E.; López-Jiménez, A. Visual Symptoms, Vegetative Growth, and Mineral Concentration in Fig Tree (Ficus carica L.) Under Macronutrient Deficiencies. Agronomy 2019, 9, 787. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120787
Garza-Alonso CA, Olivares-Sáenz E, Gutiérrez-Díez A, Vázquez-Alvarado RE, López-Jiménez A. Visual Symptoms, Vegetative Growth, and Mineral Concentration in Fig Tree (Ficus carica L.) Under Macronutrient Deficiencies. Agronomy. 2019; 9(12):787. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120787
Chicago/Turabian StyleGarza-Alonso, Carlos Alberto, Emilio Olivares-Sáenz, Adriana Gutiérrez-Díez, Rigoberto E. Vázquez-Alvarado, and Alfredo López-Jiménez. 2019. "Visual Symptoms, Vegetative Growth, and Mineral Concentration in Fig Tree (Ficus carica L.) Under Macronutrient Deficiencies" Agronomy 9, no. 12: 787. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120787