Growing Conditions Affect the Phytochemical Composition of Edible Wall Rocket (Diplotaxis erucoides)

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation
2.2. Preparation of Samples
2.3. Traits Measured
2.4. Data Analysis
3. Results
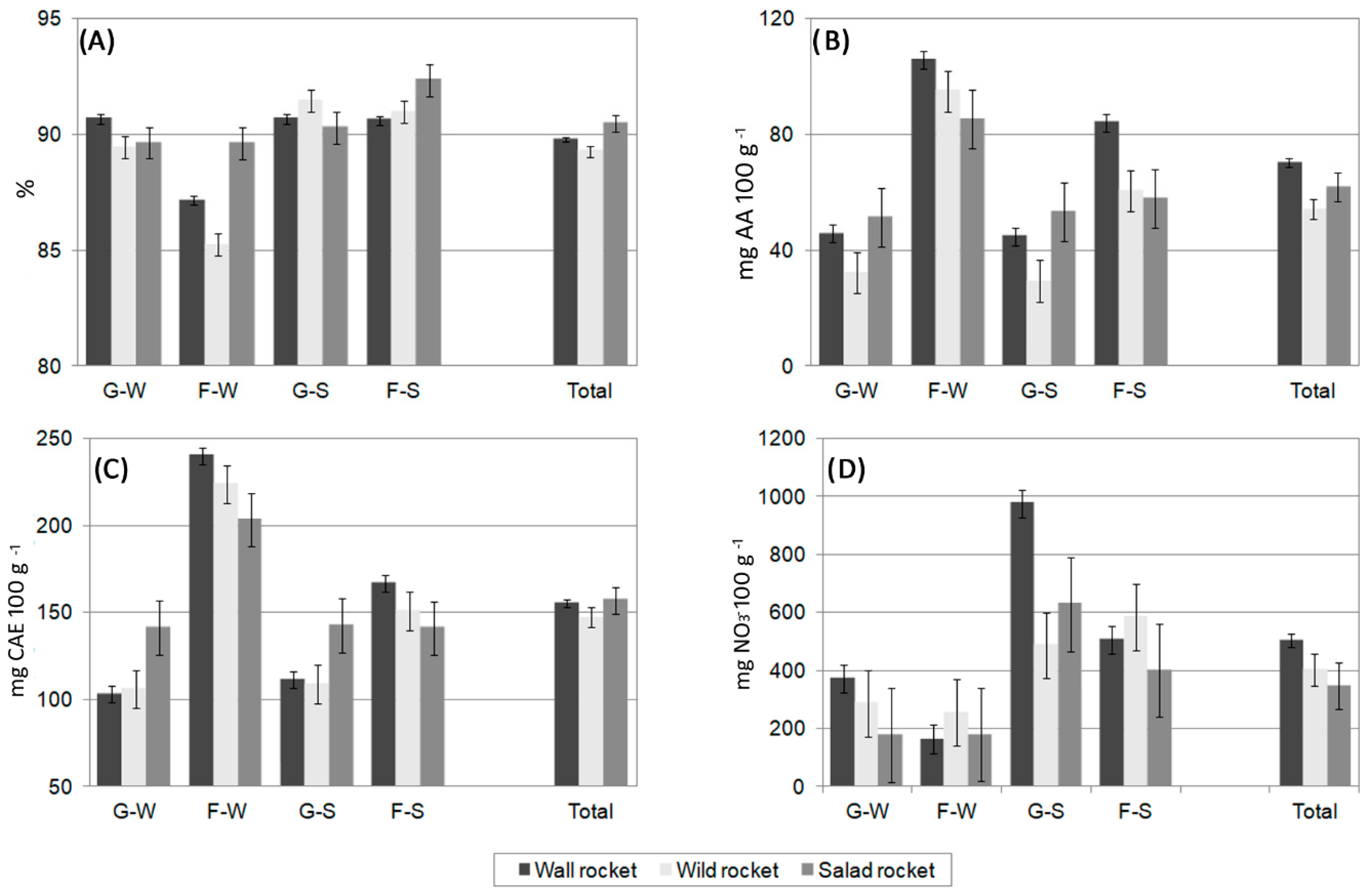
3.1. Differences among Materials of Different Species
3.2. Variation among Wall Rocket Accessions
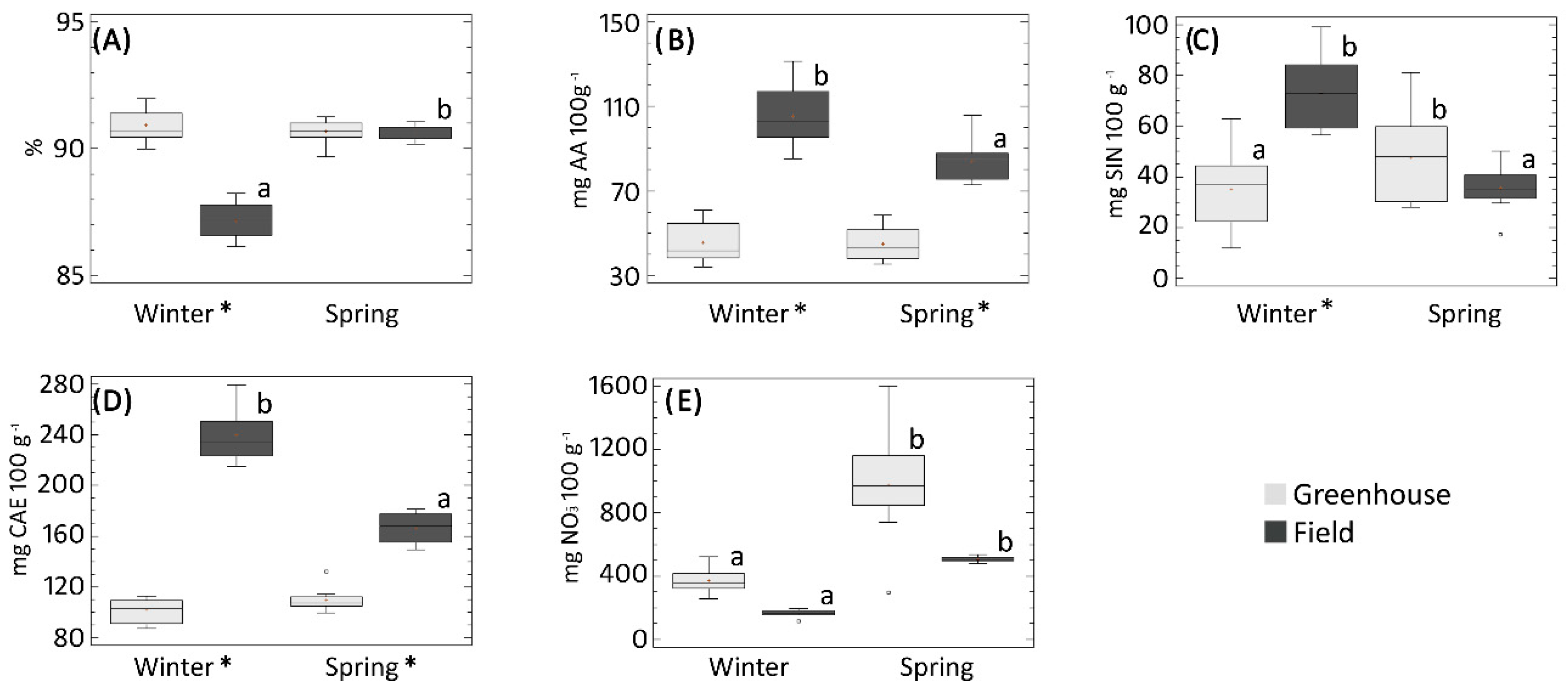
3.2.1. Effects of Accession, Growing System and Interaction
3.2.2. Effects of Accession, Growing System and Interaction
3.2.3. Correlation between Nutritional Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nutrition Society. Scientific concepts of functional foods in Europe. Consensus document. Br. J. Nutr. 1999, 81, S1–S27. [Google Scholar] [CrossRef] [Green Version]
- Pinela, J.; Carocho, M.; Dias, M.I.; Caleja, C.; Barros, L.; Ferreira, I.C.F.R. Wild plant-based functional foods, drugs, and nutraceuticals. In Wild Plants, Mushrooms and Nuts: Functional Food prOperties and Applications; Ferreira, I.C.F.R., Morales, P., Barros, L., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 315–352. [Google Scholar]
- Pinela, J.; Carvalho, A.M.; Ferreira, I.C.F.R. Wild edible plants: Nutritional and toxicological characteristics, retrieval strategies and importance for today’s society. Food Chem. Toxicol. 2017, 110, 165–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceccanti, C.; Landi, M.; Benvenuti, S.; Pardossi, A.; Guidi, L. Mediterranean wild edible plants: Weeds or “New functional crops”? Molecules 2018, 23, 2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarrera, P.M.; Savo, V. Perceived health properties of wild and cultivated food plants in local and popular traditions of Italy: A review. J. Ethnopharmacol. 2013, 146, 659–680. [Google Scholar] [CrossRef] [PubMed]
- Serrasolses, G.; Calvet-Mir, L.; Carrió, E.; D’Ambrosio, U.; Garnatje, T.; Parada, M.; Vallès, J.; Reyes-García, V. A matter of taste: Local explanations for the consumption of wild food plants in the Catalan Pyrenees and the Balearic Islands. Econ. Bot. 2016, 70, 176–189. [Google Scholar] [CrossRef] [Green Version]
- Pignone, D.; Martínez-Laborde, J.B. Diplotaxis. In Wild Crop Relatives: Genomic and Breeding Resources. Oilseeds; Kole, C., Ed.; Springer: Berlin, Germany, 2011; pp. 137–147. [Google Scholar]
- Guarrera, P.M.; Savo, V. Wild food plants used in traditional vegetable mixtures in Italy. J. Ethnopharmacol. 2016, 185, 202–234. [Google Scholar] [CrossRef]
- Licata, M.; Tuttolomondo, T.; Leto, C.; Virga, G.; Bonsangue, G.; Cammalleri, I.; Gennaro, M.C.; Bella, S.L. A survey of wild plant species for food use in Sicily (Italy)-results of a 3-year study in four Regional Parks. J. Ethnobiol. Ethnomed. 2016, 12, 12. [Google Scholar] [CrossRef] [Green Version]
- Spadafora, N.D.; Amaro, A.L.; Pereira, M.J.; Müller, C.T.; Pintado, M.; Rogers, H.J. Multi-trait analysis of post-harvest storage in rocket salad (Diplotaxis tenuifolia) links sensorial, volatile and nutritional data. Food Chem. 2016, 211, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Adikwu, E.; Deo, O. Hepatoprotective effect of vitamin C (ascorbic acid). Pharmacol. Pharm. 2013, 4, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Ashor, A.W.; Siervo, M.; Mathers, J.C. Vitamin C, antioxidant status, and cardiovascular aging. In Molecular Basis of Nutrition and Aging: A Volume in the Molecular Nutrition Series; Malavolta, M., Mocchegiani, E., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 609–619. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandl, J.; Szarka, A.; Bánhegyi, G. Vitamin C: Update on physiology and pharmacology. Br. J. Pharmacol. 2009, 157, 1097–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gols, R.; van Dam, N.M.; Reichelt, M.; Gershenzon, J.; Raaijmakers, C.E.; Bullock, J.M.; Harvey, J.A. Seasonal and herbivore-induced dynamics of foliar glucosinolates in wild cabbage (Brassica oleracea). Chemoecology 2018, 28, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Bell, L.; Oruna-Concha, M.J.; Wagstaff, C. Identification and quantification of glucosinolate and flavonol compounds in rocket salad (Eruca sativa, Eruca vesicaria and Diplotaxis tenuifolia) by LC–MS: Highlighting the potential for improving nutritional value of rocket crops. Food Chem. 2015, 172, 852–861. [Google Scholar] [CrossRef] [Green Version]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Schiattone, M.I.; Viggiani, R.; Di Venere, D.; Sergio, L.; Cantore, V.; Todorovic, M.; Perniola, M.; Candido, V. Impact of irrigation regime and nitrogen rate on yield, quality and water use efficiency of wild rocket under greenhouse conditions. Sci. Hortic. 2018, 229, 182–192. [Google Scholar] [CrossRef]
- Habermeyer, M.; Roth, A.; Guth, S.; Diel, P.; Engel, K.; Epe, B.; Fürst, P.; Heinz, V.; Humpf, H.U.; Joost, H.G.; et al. Nitrate and nitrite in the diet: How to assess their benefit and risk for human health. Mol. Nutr. Food Res. 2015, 59, 106–128. [Google Scholar] [CrossRef]
- European Comission. Commission Regulation (EU) No. 1258/2011 of 2 December 2011 amending Regulation (EC) No. 1881/2006 as regards maximum levels for nitrates in foodstuffs. Off. J. Eur. Union 2011, L320, 15–17. [Google Scholar]
- Di Gioia, F.; Avato, P.; Serio, F.; Argentieri, M.P. Glucosinolate profile of Eruca sativa, Diplotaxis tenuifolia and Diplotaxis erucoides grown in soil and soilless systems. J. Food Compos. Anal. 2018, 69, 197–204. [Google Scholar] [CrossRef]
- Durazzo, A.; Azzini, E.; Lazzè, M.; Raguzzini, A.; Pizzala, R.; Maiani, G. Italian wild rocket [Diplotaxis tenuifolia (L.) DC.]: Influence of agricultural practices on antioxidant molecules and on cytotoxicity and antiproliferative effects. Agriculture 2013, 3, 285–298. [Google Scholar] [CrossRef] [Green Version]
- Cavaiuolo, M.; Ferrante, A. Nitrates and glucosinolates as strong determinants of the nutritional quality in rocket leafy salads. Nutrients 2014, 6, 1519–1538. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.M.; Carey, E.E.; Rajashekar, C.B. Environmental stresses induce health-promoting phytochemicals in lettuce. Plant Physiol. Biochem. 2009, 47, 578–583. [Google Scholar] [CrossRef]
- Björkman, M.; Klingen, I.; Birch, A.N.E.; Bones, A.M.; Bruce, T.J.A.; Johansen, T.J.; Meadow, R.; Molmann, J.; Seljasen, R.; Smart, L.E.; et al. Phytochemicals of Brassicaceae in plant protection and human health-Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef]
- Sogbohossou, E.O.D.; Achigan-Dako, E.G.; Maundu, P.; Solberg, S.; Deguenon, E.M.S.; Mumm, R.H.; Hale, I.; Van Deynze, A.; Schranz, M.E. A roadmap for breeding orphan leafy vegetable species: A case study of Gynandropsis gynandra (Cleomaceae). Hortic. Res. 2018, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Guijarro-Real, C.; Prohens, J.; Rodriguez-Burruezo, A.; Fita, A. Potential of wall rocket (Diplotaxis erucoides) as a new crop: Influence of the growing conditions on the visual quality of the final product. Sci. Hortic. 2019, 258, 108778. [Google Scholar] [CrossRef]
- Guijarro-Real, C.; Adalid-Martínez, A.M.; Gregori-Montaner, A.; Prohens, J.; Rodríguez-Burruezo, A.; Fita, A. Factors affecting germination of Diplotaxis erucoides and their effect on selected quality properties of the germinated products. Sci. Hortic. 2019. [Google Scholar] [CrossRef]
- Cano, A.; Bermejo, A. Influence of rootstock and cultivar on bioactive compounds in citrus peels. J. Sci. Food Agric. 2011, 91, 1702–1711. [Google Scholar] [CrossRef]
- Grosser, K.; van Dam, N.M. A straightforward method for glucosinolate extraction and analysis with High-Pressure Liquid Chromatography (HPLC). J. Vis. Exp. 2017, 121, e55425. [Google Scholar] [CrossRef] [Green Version]
- Pasini, F.; Verardo, V.; Cerretani, L.; Caboni, M.F.; D’Antuono, L.F. Rocket salad (Diplotaxis and Eruca spp.) sensory analysis and relation with glucosinolate and phenolic content. J. Sci. Food Agric. 2011, 91, 2858–2864. [Google Scholar] [CrossRef]
- Singleton, V.; Rossi, J. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Guijarro-Real, C.; Prohens, J.; Rodriguez-Burruezo, A.; Adalid-Martínez, A.M.; López-Gresa, M.P.; Fita, A. Wild edible fool’s watercress, a potential crop with high nutraceutical properties. PeerJ 2019, 7, e6296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egea-Gilabert, C.; Ruiz-Hernández, M.V.; Parra, M.Á.; Fernández, J.A. Characterization of purslane (Portulaca oleracea L.) accessions: Suitability as ready-to-eat product. Sci. Hortic. 2014, 172, 73–81. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley & Sons, Inc.: Manila, Phillipines, 1984; p. 704. ISBN 978-0-471-87092-0. [Google Scholar]
- Parada, M.; Carrió, E.; Vallès, J. Ethnobotany of food plants in the Alt Empordà region (Catalonia, Iberian Peninsula). J. Appl. Bot. Food Qual. 2011, 84, 11–25. [Google Scholar]
- Egea-Gilabert, C.; Fernández, J.A.; Migliaro, D.; Martínez-Sánchez, J.J.; Vicente, M.J. Genetic variability in wild vs. cultivated Eruca vesicaria populations as assessed by morphological, agronomical and molecular analyses. Sci. Hortic. 2009, 121, 260–266. [Google Scholar] [CrossRef]
- Figàs, M.R.; Prohens, J.; Raigón, M.D.; Pereira-Dias, L.; Casanova, C.; García-Martínez, M.D.; Rosa, E.; Soler, E.; Plazas, M.; Soler, S. Insights into the adaptation to greenhouse cultivation of the traditional Mediterranean long shelf-life tomato carrying the alc mutation: A multi-trait comparison of landraces, selections, and hybrids in open field and greenhouse. Front. Plant Sci. 2018, 9, 1774. [Google Scholar] [CrossRef]
- Bell, L.; Wagstaff, C. Enhancement of glucosinolate and isothiocyanate profiles in Brassicaceae crops: Addressing challenges in breeding for cultivation, storage, and consumer-related Traits. J. Agric. Food Chem. 2017, 65, 9379–9403. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef]
- Weightman, R.; Huckle, A.; Ginsburg, D.; Dyer, C. Factors influencing tissue nitrate concentration in field-grown wild rocket (Diplotaxis tenuifolia) in southern England. Food Addit. Contam. 2012, 29, 1425–1435. [Google Scholar] [CrossRef]
- Guijarro-Real, C.; Rodriguez-Burruezo, A.; Prohens, J.; Adalid-Martínez, A.M.; Fita, A. Influence of the growing conditions in the content of vitamin C in Diplotaxis erucoides. Bull. Univ. Agric. Sci. Vet. Med. Cluj Napoca Hortic. 2017, 74, 144–145. [Google Scholar] [CrossRef] [Green Version]
- D’Antuono, L.F.; Elementi, S.; Neri, R. Glucosinolates in Diplotaxis and Eruca leaves: Diversity, taxonomic relations and applied aspects. Phytochemistry 2008, 69, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.N.; Rosa, E.A.S.; Mellon, F.A.; Kroon, P.A. Ontogenic profiling of glucosinolates, flavonoids, and other secondary metabolites in Eruca sativa (Salad rocket), Diplotaxis erucoides (Wall rocket), Diplotaxis tenuifolia (Wild rocket), and Bunias orientalis (Turkish rocket). J. Agric. Food Chem. 2006, 54, 4005–4015. [Google Scholar] [CrossRef] [PubMed]
- Sans, F.X.; Masalles, R.M. Life-history variation in the annual arable weed Diplotaxis erucoides (Cruciferae). Can. J. Bot. 1994, 72, 10–19. [Google Scholar] [CrossRef]
- Bonasia, A.; Lazzizera, C.; Elia, A.; Conversa, G. Nutritional, biophysical and physiological characteristics of wild rocket genotypes as affected by soilless cultivation system, salinity level of nutrient solution and growing period. Front. Plant Sci. 2017, 8, 300. [Google Scholar] [CrossRef] [Green Version]
- Fita, A.; Rodríguez-Burruezo, A.; Boscaiu, M.; Prohens, J.; Vicente, O. Breeding and domesticating crops adapted to drought and salinity: A new paradigm for increasing food production. Front. Plant Sci. 2015, 6, 978. [Google Scholar] [CrossRef] [Green Version]
- Kissen, R.; Eberl, F.; Winge, P.; Uleberg, E.; Martinussen, I.; Bones, A.M. Effect of growth temperature on glucosinolate profiles in Arabidopsis thaliana accessions. Phytochemistry 2016, 130, 106–118. [Google Scholar] [CrossRef]
- Steindal, A.L.H.; Rdven, R.; Hansen, E.; Mlmann, J. Effects of photoperiod, growth temperature and cold acclimatisation on glucosinolates, sugars and fatty acids in kale. Food Chem. 2015, 174, 44–51. [Google Scholar] [CrossRef]
- Salvatore, S.; Pellegrini, N.; Brenna, O.V.; Del Rio, D.; Frasca, G.; Brighenti, F.; Tumino, R. Antioxidant characterization of some sicilian edible wild greens. J. Agric. Food Chem. 2005, 53, 9465–9471. [Google Scholar] [CrossRef]
- Bianco, V.V.; Santamaria, P. Nutritional value and nitrate content in edible wild species used in Southern Italy. Acta Hortic. 1998, 476, 71–90. [Google Scholar] [CrossRef]
- Disciglio, G.; Tarantino, A.; Frabboni, L.; Gagliardi, A.; Giuliani, M.M.; Tarantino, E.; Gatta, G. Qualitative characterisation of cultivated and wild edible plants: Mineral elements, phenols content and antioxidant capacity. Ital. J. Agron. 2017, 12, 1036. [Google Scholar] [CrossRef] [Green Version]
- Orsini, F.; Maggio, A.; Rouphael, Y.; De Pascale, S. “Physiological quality” of organically grown vegetables. Sci. Hortic. 2016, 208, 131–139. [Google Scholar] [CrossRef]
- Król, A.; Amarowicz, R.; Weidner, S. The effects of cold stress on the phenolic compounds and antioxidant capacity of grapevine (Vitis vinifera L.) leaves. J. Plant Physiol. 2015, 189, 97–104. [Google Scholar] [CrossRef]
- Cárdenas-Navarro, R.; Adamowicz, S.; Robin, P. Nitrate accumulation in plants: A role for water. J. Exp. Bot. 1999, 50, 613–624. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Sum of Squares (%) | |||
|---|---|---|---|---|
| S | E | S × E | Residual | |
| d.f | 2 | 3 | 6 | 44 |
| Moisture | 4.21 * | 56.22 * | 17.69 *** | 21.88 |
| AA | 10.22 *** | 59.10 ** | 5.57 ns | 25.12 |
| TP | 0.83 ns | 72.61 ** | 7.74 * | 18.82 |
| NO3− | 3.86 ns | 52.78 * | 13.98 ** | 29.38 |
| Cycle | Trait | A | GS | A × GS | Block | Residual |
|---|---|---|---|---|---|---|
| d.f | 10 | 1 | 10 | 8 | 80 | |
| Winter | Moisture | 3.63 ns | 60.62 *** | 2.92 ns | 6.36 | 26.47 |
| AA | 3.81 * | 63.37 *** | 3.26 ns | 14.92 | 14.64 | |
| SIN | 10.00 * | 35.89 ** | 4.19 ns | 11.44 | 38.48 | |
| TP | 1.21 ns | 81.11 *** | 1.33 ns | 5.42 | 10.93 | |
| NO3− | 2.84 ns | 16.55 ns | 3.04 ns | 34.22 | 43.35 | |
| Spring | Moisture | 1.72 ns | 0.07 ns | 6.14 ns | 47.30 | 44.76 |
| AA | 2.75 ns | 52.85 *** | 7.00 * | 13.63 | 23.77 | |
| SIN | 11.63 ns | 5.10 ns | 14.96 ns | 9.06 | 59.25 | |
| TP | 2.97 ns | 57.35 ** | 3.54 * | 22.76 | 13.38 | |
| NO3− | 9.53 ns | 5.04 ns | 9.49 ns | 32.99 | 42.96 |
| Accession | n | AA | SIN | TP | NO3− | Rank | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | CV | Mean | CV | Mean | CV | Mean | CV | |||
| DER001-1 | 4 | 70.21 | 35.6 | 67.96 | 31.7 | 163.88 | 39.9 | 573.32 | 73.4 | 1 |
| DER055-1 | 4 | 77.45 | 40.1 | 48.17 | 18.9 | 153.33 | 35.3 | 470.74 | 63.7 | 2.5 |
| cv. Wasabi | 4 | 73.61 | 54.5 | 47.93 | 51.1 | 168.10 | 48.7 | 319.33 | 49.8 | 2.5 |
| DER006-1 | 4 | 68.91 | 40.9 | 55.29 | 47.2 | 158.84 | 39.3 | 494.37 | 76.0 | 4 |
| DER031-1 | 4 | 67.70 | 32.9 | 59.76 | 38.8 | 149.84 | 39.3 | 527.23 | 87.2 | 5.5 |
| DER089-1 | 4 | 64.21 | 54.3 | 44.01 | 48.9 | 143.00 | 39.5 | 542.74 | 61.3 | 5.5 |
| DER081-1 | 4 | 64.50 | 41.2 | 48.52 | 38.2 | 152.74 | 40.8 | 656.95 | 97.8 | 7 |
| DER045-1 | 4 | 62.30 | 44.9 | 43.10 | 52.4 | 141.96 | 39.3 | 493.40 | 55.1 | 8 |
| DER067-1 | 4 | 69.55 | 55.2 | 33.99 | 55.9 | 159.66 | 51.1 | 438.57 | 54.2 | 9 |
| DER064-1 | 4 | 79.71 | 47.5 | 37.40 | 56.6 | 155.16 | 46.8 | 522.47 | 75.2 | 10 |
| DER085-1 | 4 | 71.82 | 39.6 | 39.62 | 29.8 | 148.97 | 38.2 | 502.18 | 66.6 | 11 |
| Total | 44 | 70.00 | 40.1 | 47.79 | 42.7 | 154.13 | 37.3 | 503.75 | 67.5 | |
| Moisture | AA | TP | SIN | NO3− | |
|---|---|---|---|---|---|
| Moisture | −0.7032 *** | −0.7832 *** | −0.7820 *** | 0.4981 *** | |
| AA | −0.5712 *** | 0.9209 *** | 0.4554 ** | −0.5549 *** | |
| TP | −0.8546 *** | 0.6488 *** | 0.6211 *** | −0.4898 ** | |
| SIN | −0.5878 *** | 0.3670 *** | 0.5545 *** | −0.3489 * | |
| NO3− | 0.2374 ** | −0.2149 ** | −0.3252 *** | −0.2489 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guijarro-Real, C.; Adalid-Martínez, A.M.; Aguirre, K.; Prohens, J.; Rodríguez-Burruezo, A.; Fita, A. Growing Conditions Affect the Phytochemical Composition of Edible Wall Rocket (Diplotaxis erucoides). Agronomy 2019, 9, 858. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120858
Guijarro-Real C, Adalid-Martínez AM, Aguirre K, Prohens J, Rodríguez-Burruezo A, Fita A. Growing Conditions Affect the Phytochemical Composition of Edible Wall Rocket (Diplotaxis erucoides). Agronomy. 2019; 9(12):858. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120858
Chicago/Turabian StyleGuijarro-Real, Carla, Ana M. Adalid-Martínez, Katherine Aguirre, Jaime Prohens, Adrián Rodríguez-Burruezo, and Ana Fita. 2019. "Growing Conditions Affect the Phytochemical Composition of Edible Wall Rocket (Diplotaxis erucoides)" Agronomy 9, no. 12: 858. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120858