1. Introduction
The loss of agrobiodiversity has coincided in time with the development and widespread application of genetic improvement programs, but the causality of this relationship is being questioned [
1]. This coincidence gave rise to the conservationist movement that aims to retain established combinations of alleles that had become adapted to multiple local environments over the centuries. However, it is important to recall that scientific breeding has developed side-by-side with scientific improvement of environmental conditions and land use and that the Green Revolution was the result of both to understand the entire process [
2,
3].
Our ability to affect ecosystems by applying huge amounts of energy, mostly from fossil fuels [
4], has enabled us to cultivate large tracts of land and to supply them with water and nutrients while also combating the organisms that would decimate the crops. Human intervention converted large areas into homogeneous environments that made it easy for a few varieties that grew optimally in these environments to rapidly supplant diverse varieties that arose through symbiosis between humans and local environments [
5,
6]. Is it then really breeding that is responsible for the decrease in cultivated biodiversity?
Castellfollit del Boix (hereafter, simplified to Castellfollit) is a landrace of the common bean (
Phaseolus vulgaris L.) that is adapted to cultivation without irrigation in the conditions of the Castellfollit del Boix municipality in central Catalonia (northeast of the Iberian Peninsula), where its cultivation is centered. Being of Mesoamerican origin [
7], this bean has been grown in the locality since the eighteenth century [
8]. White seeds, whose shape is between rectangular and oval, characterize Castellfollit landrace (
Figure 1). The plant has an indeterminate growth pattern type III [
9], but it is cultivated without supports because the plants do not grow very high without irrigation. This variety’s main appeal lies in its sensory traits (hardly perceptible seed coat, high creaminess, and delicate but intense dry bean flavor) and culinary qualities (large proportion of whole seeds after cooking).
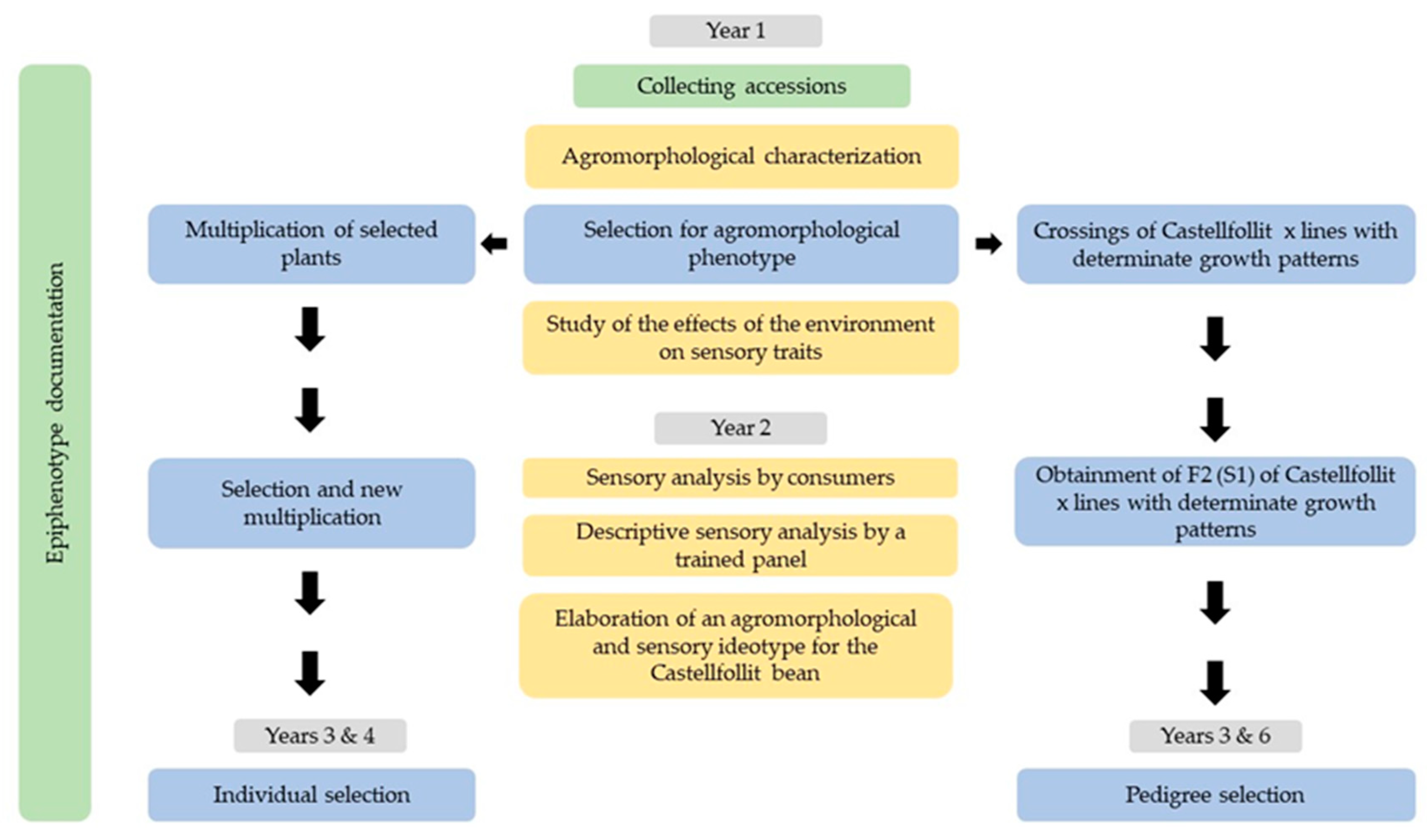
About 10 years ago, the crop had nearly disappeared due to irregular production (despite the existence of a consolidated market). Accordingly, farmers and local authorities fostered the development of a breeding program to boost cultivation by making evolve the landrace to a more determinate growth and higher tolerance to drought, while keeping its sensory quality.
This situation provides a good opportunity for studying the effects of a program to make a landrace evolve into its particular ecosystem, on its genetic diversity, and conservation expectatives over time. Their quantitative evaluation can also make it possible to compare situations, study costs, and take decisions about the conservation of agrobiodiversity. The objectives of the present study are to: (i) describe the process of genetic improvement in the Castellfollit landrace to underline the changes that are brought about in the genotypic and phenotypic information and in the diversity of the germplasm conserved in situ and ex situ, (ii) propose an index to enable the quantification of changes in the information regarding a landrace/variety over period of time, and (iii) apply this index to several landraces from diverse crops, including the Castellfollit landrace, before and after the selection processes to evaluate current trends.
3. Results
The main results regarding the epiphenotypic characteristics and the recommendations for optimizing the management of Castellfollit have been compiled in a best practices manual [
24]. All of the previously existing materials that were documented and the newly developed materials are conserved ex situ at the FMA seed germplasm collection, regardless of whether they are currently being used by farmers. All of the information that was collected about the genetic and environmental effects on sensory value will be presented together with the studies of environmental homogeneity and traditional management and the availability of well-defined genetic material to apply for a geographical quality label from the European Union.
3.1. Collecting Accessions, Phenotypic and Epiphenotypic Characterization, Phenotypic Selection, Study of Preferences, and Elaboration of the Ideotype
Searches in Spanish seed banks by name and geographical origin found no accessions of the Castellfollit landrace. Thus, only the 15 accessions that were collected from farmers in the area were used.
The field trial with these accessions revealed considerable phenotypic variation in the traits studied (
Table 2). Some of the variability among plants must be attributed to environmental factors, since the crop is cultivated without irrigation and there were substantial differences in environmental factors in the experimental field.
The sets of plants that were selected for their phenotype within each accession had less variability than the rest of the plants in the accession, since they were selected for their superior characteristics (
Table 2).
Regarding the influence of the environment on sensory and culinary traits (see material and methods
Section 2.3.2) the trained panelists detected significant differences between localities for most parameters studied (
Table 3). Nevertheless, the differences were small if we consider that the sensory traits were measured on a scale that ranged from 0 to 10. The differences in the edaphic and climatic characteristics of the locations were also small (
Table S1), and no significant correlations were found between these characteristics and the differences in sensory and culinary traits. These findings are not unusual, because the environmental effects are diffuse and affected by many interactions [
25].
Significantly more consumers preferred accession CF109 to CF106 (
p > 0.05) in the study of consumers’ preference (see Material and Methods
Section 2.3.3), although the differences in the preferences were not very large (56% CF109; 44% CF106). Although the trained panel’s descriptive analysis (see Material and Methods
Section 2.3.2) found significant differences in most sensory traits, these differences were relatively small (
Table 4). Therefore, it is difficult to attribute consumers’ preference to any particular trait scored by the trained panel.
The information from the farmers and the results of our trials during this first phase of the study led to the following ideotype for Castellfollit beans: upright plant with partially or totally determinate growth habit that is resistant to drought, root disease (especially
Pythium sp.,
Fusarium oxysporum, and
Rhizoctonia sp.), and viruses (bean common mosaic virus), and that provide good yields. Moreover, a large proportion of the seeds shape should be truncate to facilitate the identification of the landrace in the marketplace (with not rounded ends; see the seeds of the right upper part of
Figure 1). A high percentage of the seeds should remain whole after cooking, and very low seed-coat perception, low mealiness, and intense flavor characterize the cooked beans.
3.2. Selection
3.2.1. Individual
No plants with determinate growth habit were identified in the original populations of the landrace. Nevertheless, all of the selected lines had the maximum score for upright bearing. These plants showed considerable variability for the remaining agromorphological traits, being related to yield and seed size and shape (
Table 5). Lines LCF420, with big seeds, and LCF109, with small seeds, had the best yields; there were no differences between these two inbreds in the shape of the seeds that were measured by the quotient width/length. Inbred line LCF181, despite having a significantly lower yield than the two lines above, is interesting, because its seeds are easily distinguished (large projected area and size;
Table 6) from those of the Navy commercial class [
26], with which Castellfollit could be confused.
In the sensory traits (
Table 6), the lines that were closest to the ideotype were LCF122, LCF181, LCF810, and LCF109. In the culinary trait, LCF420 was noteworthy, in a negative sense, because it had a low proportion of whole seeds after cooking.
The chemical composition of the cotyledon did not significantly differ between lines (
Table 7, but there were some differences in the composition of the seed coat. The genotypes with the lowest values for Ca (LCF191) and Mg (LCF 109), indicators of low seed-coat perceptibility [
27], overlapped with other lines. Uronic acid, which is the other trait related with seed-coat perceptibility [
27], did not significantly differ between the genotypes.
In light of these results, future efforts will aim to multiply and cultivate inbred lines LCF109, LCF122, and LCF101; LCF181 has also been included because of the peculiar shape of its seeds.
3.2.2. Pedigree Selection
The main aim of the pedigree selection program was to obtain plants with a totally determinate growth habit and upright bearing. All of the selected lines achieved this objective. On the other hand, these lines differed in most other traits. The line that most closely resembles LCF109, the control with indeterminate growth representative of Castellfollit material is LCF6091, which significantly differs in three of the eight traits; the other three lines significantly differ in four of the eight traits (
Table 8). In general, the inbred line that has donated determinate growth habit and a degree of resistance to drought has conferred a less flat seed shape (smaller projected area), although three of the four lines still have the elongated seeds that clearly differ from the more rounded seeds of Croscat (
Table 8) that are characteristic of the Navy commercial class [
26].
Crossings with Croscat in the pedigree selection program also resulted in changes in sensory traits (
Table 9). Mealiness decreased in lines LCF6071, LCF6001, and LCF6091. In general, the seed-coat roughness and brightness increased. No changes in aroma or flavor were detected. The most negative result of the pedigree selection program was a decrease in the percentage of seeds that remained whole after cooking (
Table 9); this percentage was lower in all of the new lines than in the initial Castellfollit population, but still higher than in the line used to confer determinate growth habit, where it is approximately 20% [
16].
We have decided to multiply the four lines and distribute them to farmers to observe their behavior over several seasons and to gauge consumers’ opinions, since the differences in the sensory and culinary traits between the new lines are small and the differences in agromorphological traits indicate considerable variability.
3.2.3. The Selected Germplasm
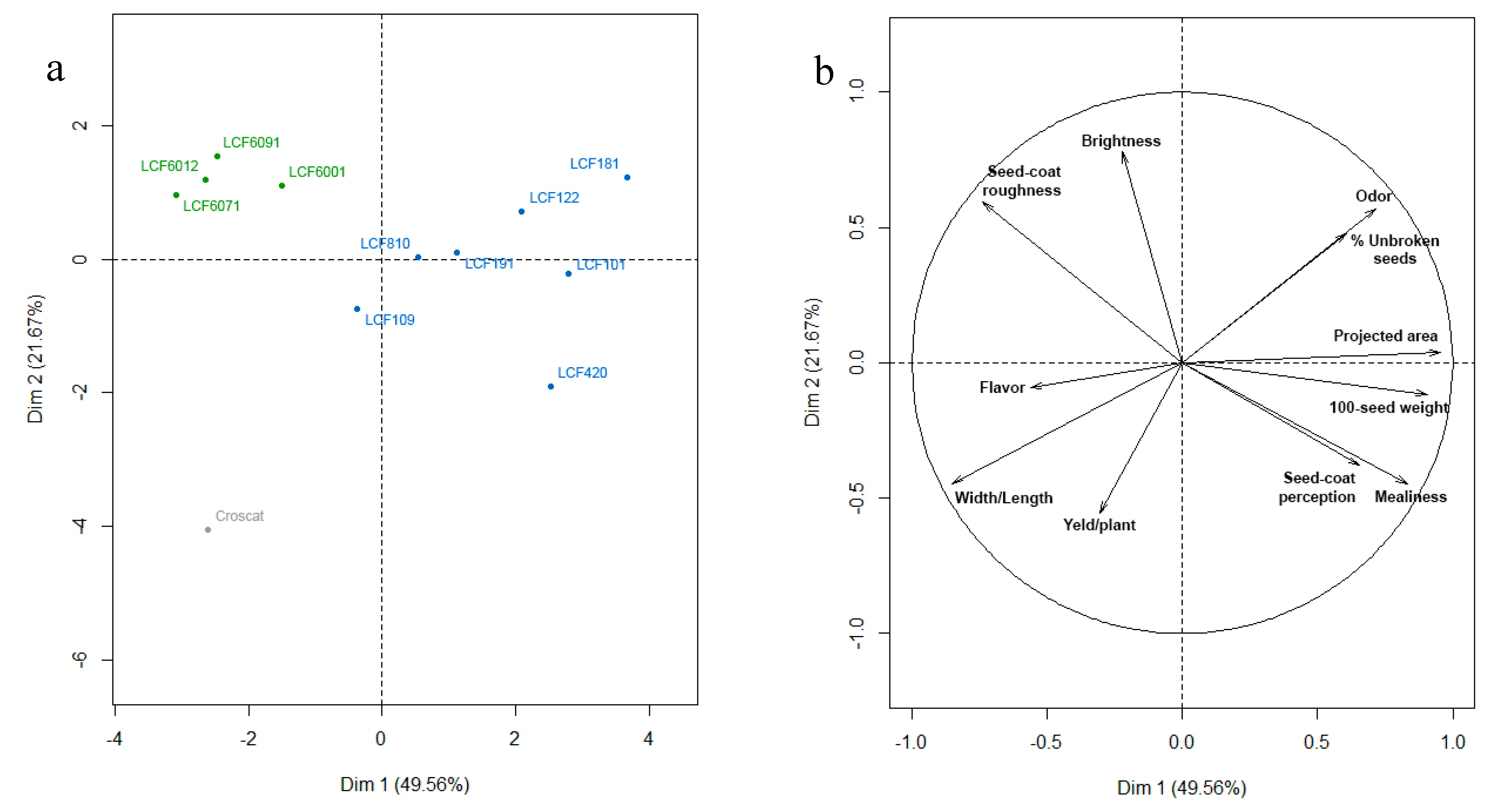
We performed a PCA analysis to evaluate the relationships among the different materials after selection and breeding with the scores of the seven lines coming from the individual selection program (
Table 5,
Table 6 and
Table 7), the four lines of the pedigree selection, and Croscat (
Table 8 and
Table 9) (
Figure 3). Before processing, data were divided by the value of LCF109, present in both experiments, to lower the effects of environmental factors.
The cumulative variation was explained by the first two axes was 71,23% (
Figure 3). The traits that best correlated with the first axis were projected area and 100 seeds weight (
R = 0.95 and
R = 0.90, respectively), and width/length (
R = −0.85). With the second axis the highest correlations were those of brightness (
R = 0.77), seed-coat roughness (
R = 0.59), intensity of odor (
R = 0.56), and yield (
R = −0.55). Croscat, which is the donor of the determinate growth, has the most extreme position and it is clearly different from the rest of inbreds. The lines that were selected into Castellfollit landrace show a considerable dispersion across the two axes, while the inbreds coming from the pedigree process mainly differ in their position with respect to the first axis (
Figure 3). Our intervention had the aim of simulating a spontaneous process of crossing between two varieties and the posterior selection made in favor of the forms closer to the original landrace while taking advantage of new properties, as is usual in farmers’ more or less conscious procedures. The result is that the new inbreds of complete determinate growth habit are in the range of variation of the landrace with respect to sensory properties and yield (axis 2,
Figure 3), but they have broadened the variation in size and shape of the seeds (axis 1,
Figure 3).
3.3. Effects on Conservation and the Predisposition to Conservation during the Intervention Process
At the start of the intervention, there were 15 accessions of the Castellfollit landrace (variety population) that were conserved in situ (but in high risk of extinction), and none were conserved ex situ. The CPI score at that time was 0.15 (
Table 9; details of calculations can be consulted as
Table S2), which indicated that there was little documented information about the landrace and its environment. At the end of the intervention, fewer of the old accessions were being cultivated because those farthest from the ideotype had been abandoned; however, the four lines selected within the accessions and four lines improved by the introgression of genes conferring determinate growth habit and other favorable traits were also being cultivated. Moreover, all of the accessions collected were also being conserved ex situ. After the intervention, the CPI score was 0.59 (
Table 10; details of calculations can be consulted as
Table S2); the increase in the score reflects the documentation of the material and the increased vigor of its use.
We calculated the CPI for 13 other landraces of diverse species that our team has worked with (
Table 10; details of calculations can be consulted as
Table S2). One of these (maize “d’escairar”, used to make soups) was included, because the only activities to date have been collecting accessions and some local promotion, but no other efforts to characterize the landrace have been undertaken. In the remaining cases, the CPI score ranged from 0.07 to 0.36 before intervention and from 0.29 to 0.80 after the intervention. The lowest initial CPI score was for the Mandó tomato, a landrace about which very little information had been documented when we started to work with it. The highest final CPI score was for the Ganxet and Tavella Brisa landraces of common beans, which our team has been researching, breeding, and promoting for 25 years and 20 years, respectively.
In each case, one or two researchers working with each landrace were asked to quickly assess the items in the CPI based on their knowledge; in fact, on average calculating the CPI only took 10 min. Despite the inevitable risk of error related to the personal bias of the estimations, in the cases where two independent assessors calculated the CPI for a landrace, the results were very similar (
Table 10), which suggested the confirmation of the hypothesis proposed in the material and methods section that the deviations of all the items would add up to zero. Without doubt, in its present form, the index is far from achieving the efficiency and predictive capacity of established indices for cardiovascular risk [
28], diet quality [
29], or food safety [
30], but it might be a starting point for further development.
Disregarding the above-mentioned case of maize “d’escairar”, the CPI score increases during the intervention, but this increase is not linear (mean correlation coefficient between the two series [excluding the maize landrace], R = 0.54, nearly significant at p < 0.05). The differences among the situations of the landraces before the intervention, in the time between T0 and T1, in the investments in each case, etc. could explain the differences among cases.
It is also noteworthy that the CPI scores are independent of the number of accessions conserved in situ and/or ex situ (correlation coefficient between the CPI and the number of accessions conserved ex situ after the intervention, R = 0.21, not significant at p < 0.05). This is important, because the objective of the CPI is to complement the information that is obtained in a simple inventory of the resources conserved.
4. Discussion
At the start of this intervention in the Castellfollit bean, the cultivation of the landrace had substantially decreased, despite the greater ease of cultivating large fields of beans facilitated by mechanization and the presence of a market demand that exceeded the supply. It seems paradoxical that the dilution of the autochthonous seed with improved materials, mainly by spontaneous crossings with Great Northern [
7], combined with increased capacity to transform fields leading to the abandonment of traditional methods of production in an ecosystem that was very fragile, due to a lack of water, led to the near disappearance of the cultivation of dry beans in the area of Castellfollit del Boix due to decreasing yields.
The project to recover and improve the Castellfollit landrace has enabled us to collect, characterize, multiply, and conserve the historic material. We assessed farmers’ needs and consumers’ preferences, which were obtained inbred lines within the historic material, and introduced genes that favor management and avoid yield loses while keeping modifications to the sensory profile to a minimum. It is true that we have not limited our genetic intervention to the introduction of a single gene to confer determinate the growth habit through backcrossing, so the new varieties with determinate growth habit probably have a considerable number of genes from the donors’ genome. Nevertheless, this situation is not unlike the process that has occurred throughout history in landraces through fortuitous crossings with other varieties [
31,
32]. We selected the crossings with determinate growth that gave the highest yields of seeds most similar to the historical variety, analogously to what farmers have always done (selecting the plants whose appearance they found most interesting from the appearance of these fortuitous hybrids and continuing this process until they obtained totally homozygous materials). We are also aware that the number of accessions and plants involved in the selection programs is limited, but this kind of interventions must reach a consensus between effectiveness and funding, as the application of the results cover a limited geographical area. Nevertheless, the results show that even dealing with a reduced sample of the variability significant progress is possible.
Throughout this process, we also documented the epiphenotype of the landrace, and proposed methods of cultivation that respected the traditional values while combining them with modern techniques [
24]. We also rejected the currently very common, inefficient practice of cultivation in shallow fields, which is acceptable for cereals, but not for beans. Finally, the farmers now have all the tools that are necessary to apply for a quality label, such as a protected denomination of origin (PDO) from the European Union [
33].
All of the changes brought about in the intervention have been used to develop an index that quantifies the aspects that our experience tells us favors the conservation of landraces. We consider it to be especially important that the proposed index takes the information collected about a landrace and the efforts to promote it into account, because information regarding the germplasm is fundamental for fomenting its use and thus for its conservation [
10,
34].
When comparing both the conservation of the landrace and its derivatives before and after the process and comparing the CPI scores at the two points in time indicate that the prospects for conserving the landrace have improved, as has also occurred in other species and situations in which our team has intervened. In every case, the interventions have had a very positive effect on the conservation of the landraces and aspects that are related to their use. The proposed CPI has proven to be easy to calculate and useful in explaining the state of conservation of landraces, complementing simple data regarding the number of accessions conserved in situ and ex situ and diversity indices derived from these data [
35,
36]. The simplicity with which a person who knows the landrace and its environment can calculate the CPI score makes it, together with the list of accessions, especially appropriate for evaluating the achievements of interventions to conserve plant genetic resources.
Our projects have shown us that landraces lose ground, because it is difficult for them to compete with improved materials, mostly because of their low response to the more intensive cultivation methods that have become generalized. This is fundamentally a matter of objectives, because the results favor biodiversity when the goal is to improve the materials in a specific environment rather than in a generalized environment [
37]. Scientific breeding of landraces, which should be the natural evolution of these varieties in the twenty-first century [
5], normally involves collecting materials and characterizing them, followed by selection within the collected materials, sometimes incorporating genes from crop wild relatives or other cultivated varieties, a process that significantly increases the genetic variability of the materials. As a result, the ex situ conservation of the previously characterized (often from many points of view) material is ensured, and new variants that are adapted to diverse conditions can be obtained. The number of varieties that have evolved from initial populations depends on the diversity of environments in the area of cultivation and the intended uses for the obtained varieties. The more microenvironments there are, the more it makes sense to develop many varieties. It is evident that we can use our knowledge of genetics and agronomy to recover some of the diversity and wealth of traditional varieties that arose before the advent of scientific crop management and to make these varieties competitive if we opt for low-input crops for a sustainable agriculture [
38,
39]. Thus, restoring diversity is mainly a matter of appreciating environmental diversity; in this way, we can save and even expand genetic diversity. We can achieve new varieties that are adapted to local conditions very quickly by applying our knowledge of the genome. Perhaps by refocusing the aims of breeding programs, we can transform what Louwaars [
1] aptly describes as “troubled relationships” between plant breeding and diversity into a true friendship.

 ,
, 




{kind=link}
{kind=link}
{kind=link}