Cladobotryum mycophilum as Potential Biocontrol Agent
by
 ,
,
Mila Santos
1,*, 
Fernando Diánez
1,
Alejandro Moreno-Gavíra
1,
Brenda Sánchez-Montesinos
1 and
Francisco J. Gea
2 1
Departamento de Agronomía, Escuela Superior de Ingeniería, Universidad de Almería, 04120 Almería, Spain
2
Centro de Investigación, Experimentación y Servicios del Champiñón (CIES), Quintanar del Rey, 16220 Cuenca, Spain
*
Author to whom correspondence should be addressed.
Agronomy 2019, 9(12), 891; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120891
Submission received: 10 November 2019
/
Revised: 3 December 2019
/
Accepted: 13 December 2019
/
Published: 15 December 2019
(This article belongs to the Special Issue Integrating Pest Management into Agricultural Production Systems)
Abstract
:A study was conducted to explore the efficacy of potential biocontrol agent Cladobotryum mycophilum against different phytopathogenic fungi. The growth rates of 24 isolates of C. mycophilum were determined, and their antagonistic activity was analysed in vitro and in vivo against Botrytis cinerea, Fusarium oxysporum f. sp. radicis-lycopersici, Fusarium oxysporum f.sp. cucumerinum, Fusarium solani, Phytophthora parasitica, Phytophthora capsici, Pythium aphanidermatum and Mycosphaerella melonis. Most isolates grow rapidly, reaching the opposite end of the Petri dish within 72–96 h. Under dual-culture assays, C. mycophilum showed antagonistic activity in vitro against all phytopathogenic fungi tested, with mycelial growth inhibition ranging from 30 to 90% against all the different phytopathogens tested. Similarly, of all the selected isolates, CL60A, CL17A and CL18A significantly (p < 0.05) reduced the disease incidence and severity in the plant assays compared to the controls for the different pathosystems studied. Based on these results, we conclude that C. mycophilum can be considered as a potential biological control agent in agriculture. This is the first study of Cladobotryum mycophilum as a biological control agent for different diseases caused by highly relevant phytopathogens in horticulture.
Keywords:
biological control; Cladobotryum; Mycosphaerella; Pythium; Phytophthora; Fusarium; Botrytis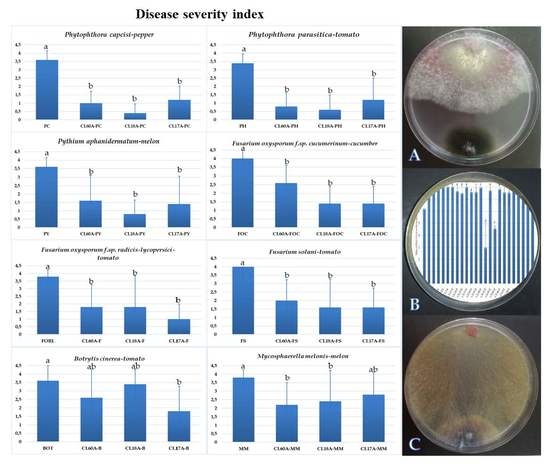
1. Introduction
Cobweb, caused by a mycopathogenic fungus (Oudem.) W. Gams and Hooz (teleomorph Hypomyces odoratus G.R.W. Arnold), is one of the most serious diseases affecting white button mushroom (Agaricus bisporus (Lange) Imbach) cultures worldwide [1,2,3,4]. Its occurrence generally reduces the production and quality, and thus generates economic losses because of mainly cap spotting, the reduction of crop surface and the necessity of early crop termination when the disease becomes epidemic [5]. The main symptoms of cobweb are characterised by the occurrence of a white fluffy mycelium over the surface of the casing layer and attacks on primordia and developing fruitbodies. The pathogen can also cause two types of cap spotting that may even appear post-harvest, affecting the profitability of the product [6]. Recently, C. mycophilum has been identified as the causal agent of cobweb in cultivated king oyster mushroom (Pleurotus eryngii (DC.: Fr.) Quél.) in Spain and Korea [7,8,9] and oyster mushroom (Pleurotus ostreatus (Jacq.) P. Kumm.) in Spain [10]. Methods to control cobweb include strict hygiene measures to prevent dispersion of the conidia and the application of fungicides, mainly prochloraz and metrafenone [11].
Fungal associations with other fungi are termed fungicolous fungi [12,13]. The aggressive fungicolous taxa, such as Gliocladium spp. and Trichoderma spp., have led to numerous studies on these fungi as biological control agents [14]. Most species of the Cladobotryum genus are characterised by rapid growth and colonisation of the parasitic basidiomycete [4]. Considering their high mycoparasitic capacity, these species are potential candidates for use as biological control agents against key agricultural phytopathogens. However, few in vitro studies have shown the antagonistic activity of Cladobotryum against phytopathogenic fungi. In this context, Bastos et al. [15] describe C. amazonense as a new hyperparasite of Crinipellis perniciosa, that causes ‘witches’ broom disease’ in the cocoa tree. Marzuki et al. [16] assess a 74.8 and 74.7% decrease in the mycelial growth of Ganoderma lucidum and G. boninense, respectively, at the hand of Cladobotryum semicirculare, and showed the in vitro antagonistic activity of C. semicirculare isolates against G. boninense and other fungal pathogens, including Fusarium sp and Phoma sp., assessing 55 and 60% rates of inhibition, respectively. Similarly, Ramos [17] evaluated the in vitro and in vivo antagonistic activity of C. pinarense, C. semicirculare and C. virescens in the control of the phytopathogens Alternaria brassicicola, Corynespora cassiicola and Fusarium chlamydosporum. The findings showed that the three Cladobotryum species had an antagonistic effect on the pathogens studied, with no harm to plants or fruits, highlighting a fungicidal effect on filtered extracts of the three antagonistic isolates.
Biological control of diseases caused by phytopathogenic fungi has not been described for Cladobotryum mycophilum. Therefore, the main objective of this study is to determine: (a) The growth rate of 24 C. mycophilum isolates; (b) the in vitro antagonistic activity of such isolates against eight phytopathogens of interest in agriculture; (c) the in vivo capacity for controlling the development of the disease caused by such phytopathogens in different pathosystems under greenhouse conditions.
2. Materials and Methods
2.1. Fungal Isolates
A total of 24 isolates previously identified as C. mycophilum used in this study were obtained from A. bisporus crops showing cobweb disease symptoms [3]. The phytopathogenic fungi used were: Botrytis cinerea Pers (BOT), Fusarium oxysporum f. sp. radicis-lycopersici Jarvis and Shoem (FORL), Fusarium oxysporum f.sp. cucumerinum Owen (FOC), Fusarium solani (Mart.) Sacc (FL), Phytophthora parasitica Dastur var. nicotianae (Breda de Haan) Tucker (PH), Phytophthora capsici Leonian (PC), Pythium aphanidermatum (Edson) Fitzp (PY) and Mycosphaerella melonis (Pass.)(MM).
Plants infected with the pathogens were collected from different crops in the province of Almeria (Spain) during 2016–2018. Selected stem and leaf sections with active lesions were cut, and tissue pieces from the boundaries between healthy and discoloured areas were planted on potato dextrose agar. Pathogenicity tests were carried out to confirm their pathogenic capacity.
Fungal isolate was grown on PDA for 5 or 15 days at (25–27) ± 2 °C under dark conditions. Spore suspensions of Cladobotryum and phytopathogens isolates were prepared by flooding plates of 10-day-old cultures with sterile distilled water, scraping with a sterile glass rod and filtering and adjusted to a concentration of 1 × 106 spores/mL with a Neubauer haemocytometer.
2.2. Growth Conditions of the Cladobotryum Isolates in PDA
Twenty-four isolates of C. mycophilum were cultured at 25 °C for 7 d in PDA. Mycelium discs (5 mm diameter) were obtained from the edges of the PDA plates and inoculated into new PDA at 0.5 cm from the edge. The plates were incubated for 4–7 d at 25 °C. The colony diameters were recorded every day during culturing. The experiment was completely randomised with five replicates.
2.3. Screening Test of Antagonistic Isolates
The 24 isolates of C. mycophilum were screened for their antagonism in vitro against eight phytopathogens by adopting the confrontation assay of Santos et al. [18].
Petri dishes (9 cm diameter) containing 17 mL of PDA (Bioxon, Becton Dickinson, Mexico) were prepared. In this test, 0.5-cm plugs of mycelium with active growth of each isolate were placed at the ends of Petri dishes with a distance of 8 cm between the two fungi, antagonist-phytopathogen.
A plug of PDA medium was used as control treatment while the pathogen plug was placed at the other side. The plates were then assessed by measuring the distances between pathogen and fungal cultures. Results were transformed into percentages of mycelium growth inhibition. These tests were carried out in quintuplicate.
2.4. Greenhouse Evaluation of Selected Antagonists of C. mycophilum on the Severity of Eight Phytopathogens
Biocontrol activity of selected Cladobotryum mycophilum isolates on different pathosystems was determined: B. cinerea-tomato, F. oxysporum f. sp. radicis-lycopersici-tomato, Fusarium oxysporum f.sp. cucumerinum-cucumber, Fusarium solani-tomato, Phytophthora parasitica-tomato, Phytophthora capsica-pepper, Pythium aphanidermatum-melon and Mycosphaerella melonis-melon. All seeds were disinfected with 2% hypochlorite for 3 min and washed liberally with tap water to eliminate residues. Subsequently, the seeds were pregerminated in darkness in a humid room at 25 °C and transplanted to 500 mL pots into a commercial peat mix, with one seed per pot. Simultaneously to sowing, 5 mL of water (T0) or 5 mL spore suspension of each isolate of Cladobotryum was placed in each pot at 1 × 106 propagules/plant.
To prepare Phytophthora parasitica, Phytophthora capsici and Pythium aphanidermatum inocula, the procedure described by Diánez et al. was followed [19]. To obtain the inoculum, isolates were grown on PDA for 7 d at 25°C. Then, plates were incubated under fluorescent light for 3 d at 25 °C to induce sporangium formation. Twenty-four hours before inoculation, fungal colonies were covered with 20 mL of sterilized tap water and incubated under light overnight. During the inoculation day, Petri dishes were placed at 4 °C for 30 min, followed by 60 min at room temperature to enhance zoospore release from sporangia. The flooding water, containing zoospores and mycelium was filtered through two layers of cheesecloth. The concentration of the zoospore suspension was adjusted to approx. 104 zoospores mL−1 using a haemocytometer. The inoculum was used immediately. Inocula of the other phytopathogens were prepared by scraping and subsequent filtration, as previously indicated.
The phytopathogen was inoculated when the plant had a second true leaf and was performed using a sterile micropipette, by pouring the zoospores/conidias suspension (5 mL) uniformly over the surface of the peat in each pot at a concentration of 104 propagules/mL. In the case of B. cinerea and M. melonis, the pathogen was applied by spraying five times at the same concentration throughout the plant. Previously, the first true leaf had been cut to facilitate pathogen entry.
All pathogenicity tests were performed under greenhouse conditions and at different seasons (for B. cinerea the experiment were performed in winter; for the rest of the phytopathogens in spring) to provide the ideal environmental conditions for each pathogen. The experimental units consisted of four repetitions with 24 plants per pathosystem. The experiment were conducted using completely randomized block designs.
Symptom severity was rated periodically, and 30–60 days after inoculation final disease severity index was estimated according to the following scale: 0 = healthy plant; 1 = symptoms beginning; 2 = moderate symptoms; 3 = severely affected plant; and 4 = dead plant. Similarly, possible symptoms caused by the application of different C. mycophilum isolates without pathogen were observed in melon, tomato, pepper and cucumber plants.
2.5. Statistical Analysis
Data were analysed with analysis of variance (ANOVA), carried out using the Statgraphics Centurion ver. (XVI) program to evaluate the parameter values differences. Differences between treatments were determined by Duncan’s multiple range test at 5% of significance level.
3. Results
3.1. Colony Growth of Cladobotryum Isolates
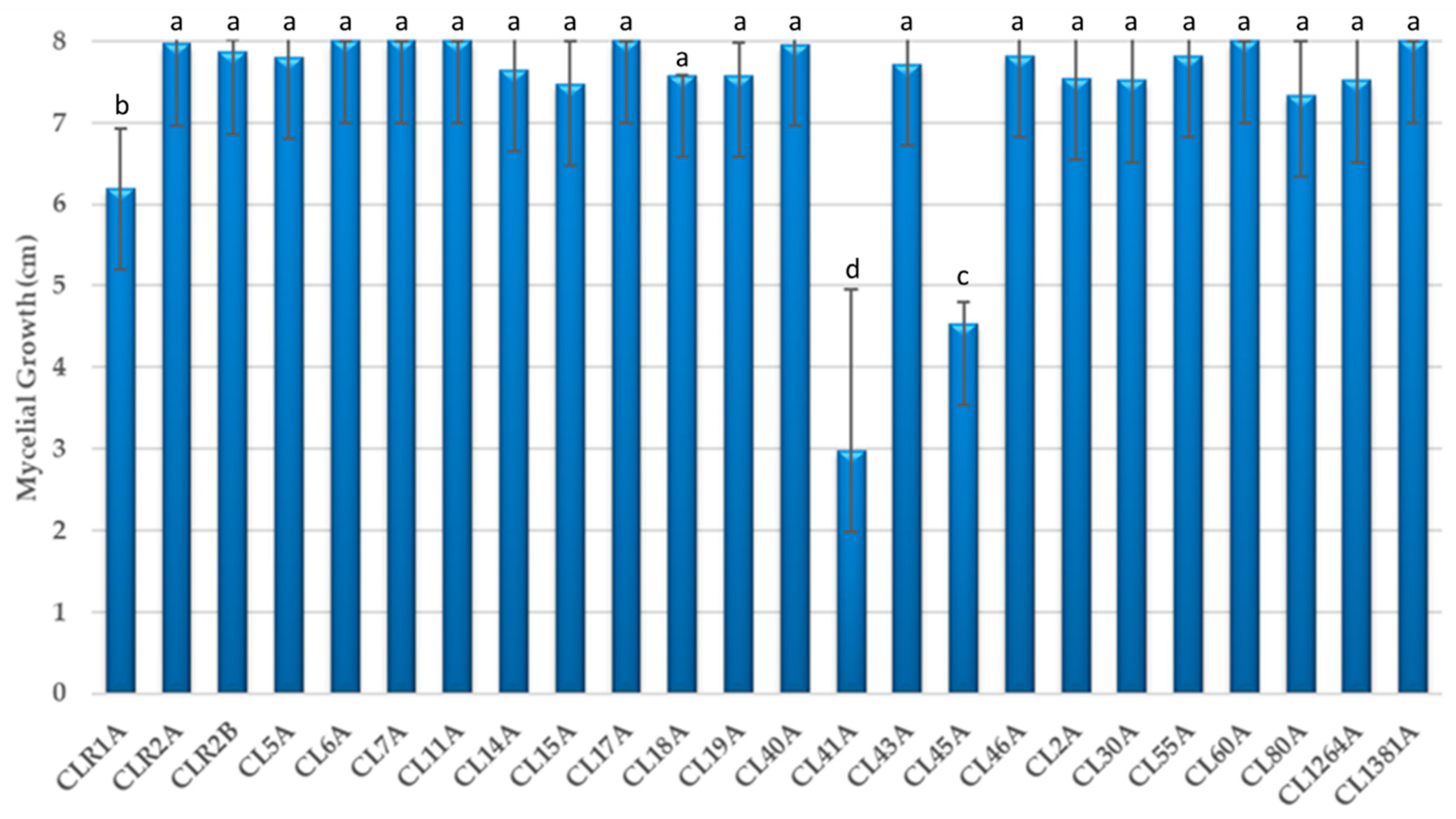
In Figure 1, the results obtained from the mycelial growth of 24 isolates of C. mycophilum are shown. The colony growth of most isolates was fast, reaching the opposite end of the Petri dish within 72–96 h. The colour of the colonies ranged from yellowish to pink and turned purple after a few days, except in isolate CL1381A, whose mycelium colour was greyish white. No isolates were discarded in determining the antagonistic activity of Cladobotryum against phytopathogens.
3.2. Effects of Cladobotryum Isolates on the Radial Growth of Phytopathogens
In Table 1, the results obtained from the microbial antagonism for all phytopathogens tested are observed. All C. mycophilum isolates have shown in vitro antagonistic activity against all phytopathogenic fungi tested. The highest inhibition percentages were detected for Phytophthora parasitica, which reached values higher than 90% for 12 Cladobotryum isolates. In contrast, the lowest inhibition values were detected for Pythium aphanidermatum, whose maximum failed to exceed 60% for the CL17A isolate. For the other phytopathogens, mycelial growth inhibition ranged from 30 to 85%. Three isolates were selected for the in vivo tests, CL60A, CL17A and CL18A, because they reached the highest values for the highest number of phytopathogens.
3.3. Biological Control of Cladobotryum Isolates against Several Diseases
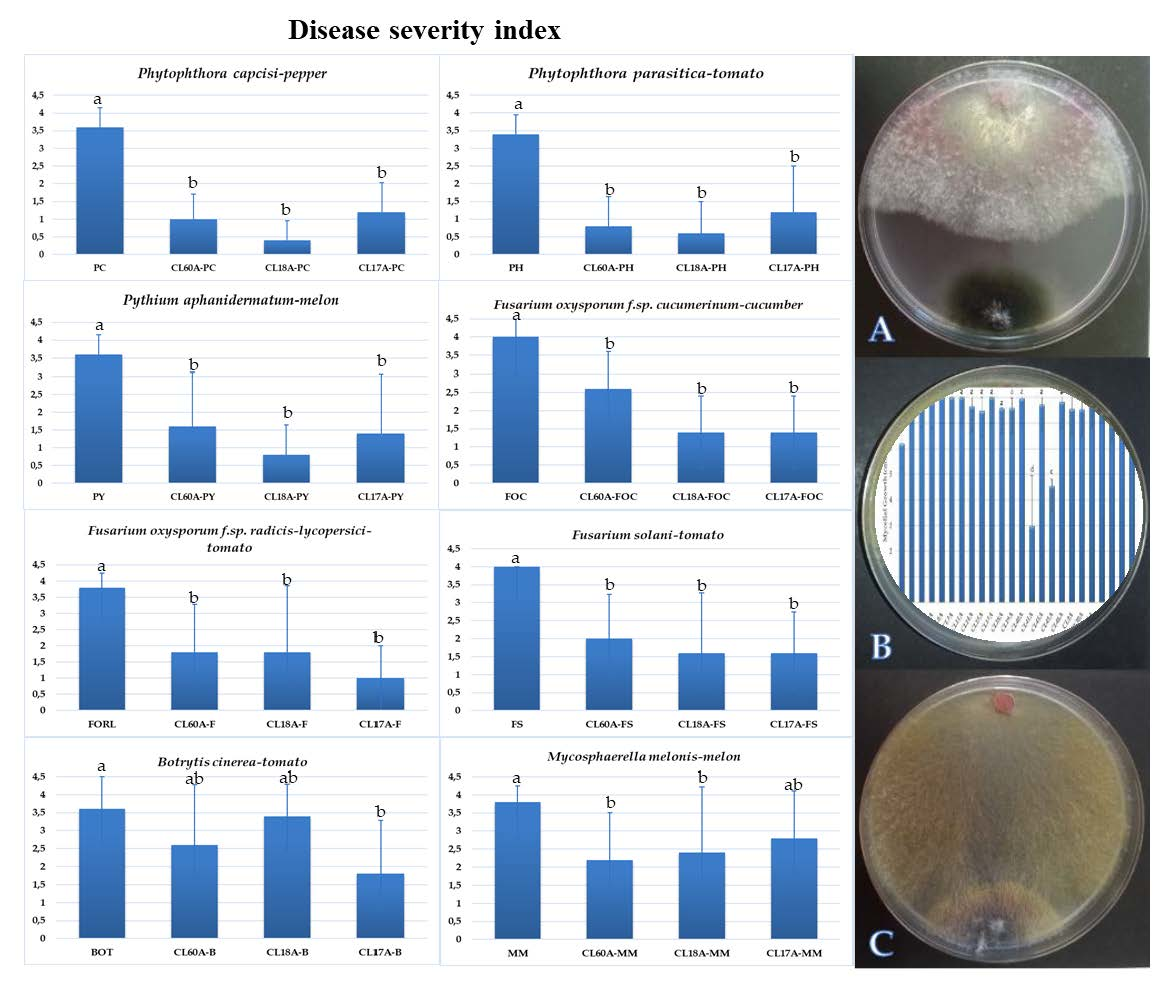
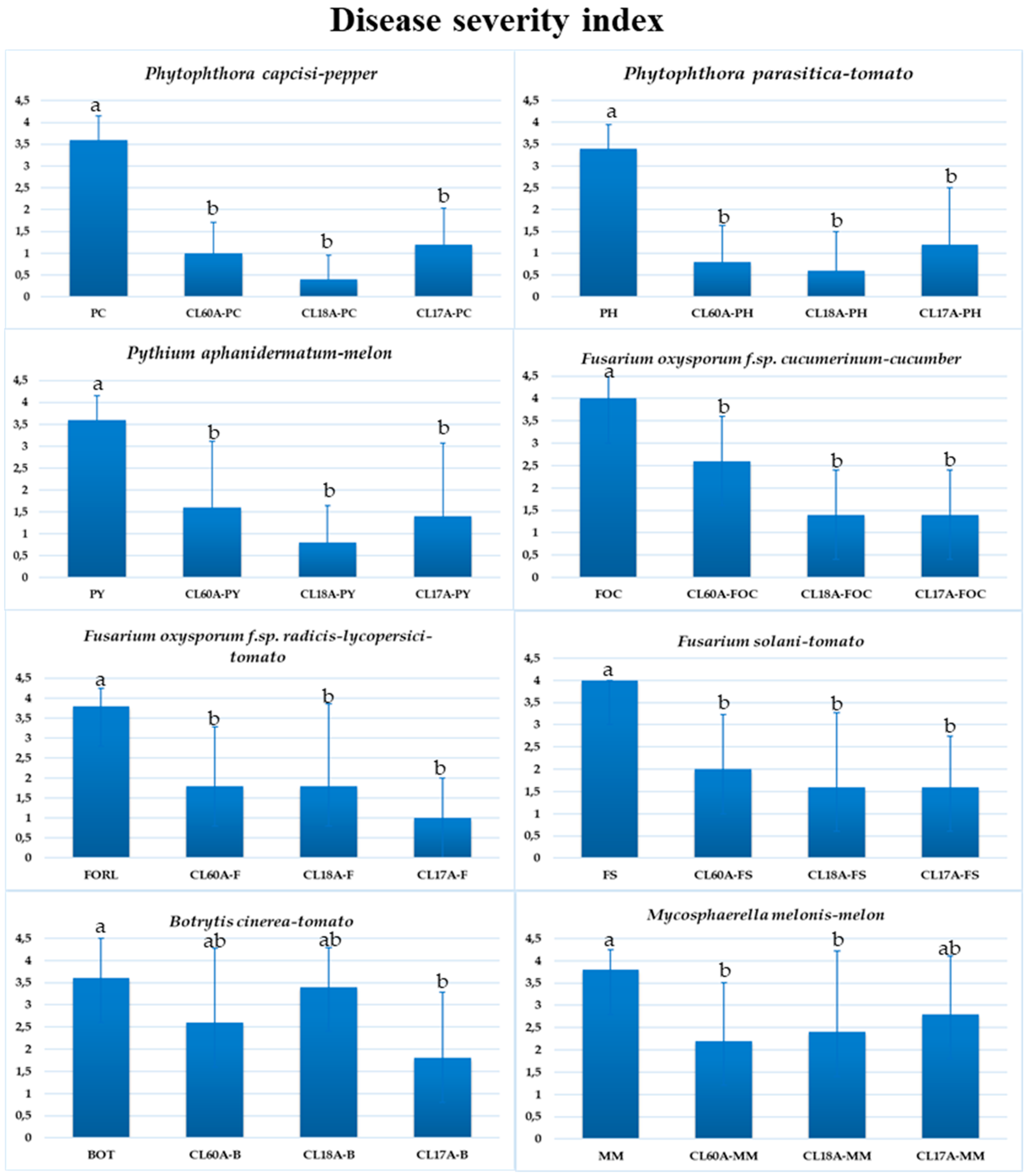
The severity of the disease caused by the different phytopathogens tested decreased after applying the three isolates, CL60A, CL17A and CL18A (Figure 2). Statistically significant differences between all treatments and experimental controls were observed (p < 0.05). No symptom was observed after applying CL60A, CL17A or CL18A in any plant tested (Figure 2), thus showing a plant growth-promoting effect for different parameters (data not shown).
Despite the lack of significant differences in the biological control of CL60A, CL17A and CL18A for different oomycetes, CL18A has reduced the disease by 77.8, 82.3, and 88.9% for Pythium aphanidermatum, Phytophthora capsici and Phytophthora parasitica, respectively. In the case of Fusaria, the disease reduction shown by the different isolates ranged from 35 to 60% for F. solani and F. oxysporum f.sp. cucumerinum, and from 52.7 to 73.7% for F. oxysporum f. sp. radicis-lycopersici. Lower values were detected after applying the three antagonistic isolates against B. cinerea and M. melonis. Thus, significant differences were only observed when applying CL17A, which reduced the disease severity by 50% for B. cinerea and by 37 to 42.1% in the control of Mycosphaerella melonis.
4. Discussion
Numerous studies have reported using mycopathogenic fungi to control pests and diseases. Thus, Ampelomyces quisqualis, a mycoparasite specific to Erysiphales, has been studied as a biocontrol agent [20], and Sphaerellopsis filum has been used for the control of plant rust disease caused by Puccinia graminis subsp. graminícola [21], among other examples. However, the aggressive fungicolous taxa most studied and used in agriculture is Trichoderma, the genus by excellence [22,23,24].
In this study, we analysed the capacity of Cladobotryum isolates to control different phytopathogens that cause severe damage in many crops in vitro and in vivo to assess whether we can consider Cladobotryum as a potential biological control agent. All 24 C. mycophilum isolates have shown high percentages of in vitro inhibition of the phytopathogens tested, as a result of high competition for nutrients and space because of the rapid growth of Cladobotryum (Figure 3). These values are lower in the case of P. aphanidermatum because this phytopathogen grows even faster, but antagonism exceeds 50% for nine of the isolates tested. No inhibition halos for the production of metabolites with fungistatic activity were observed, and the mycelium of Cladobotryum grew normally over that of the phytopathogens, albeit showing mycelial degradation under the microscope. The production of metabolites identified in these isolates has not been studied, but this genus is known to cause hyphal lysis by producing a wide variety of secondary metabolites with marked antifungal as well as antibacterial activities as well as repressive effects on cancer cells [25,26,27]. One of these fungal metabolites is cladobotryal, a Cladobotryum varium isolate with antifungal activity [28].
In addition, very promising results have been obtained in in vivo tests. The best results have been achieved with the three oomycetes tested, with high percentages of inhibition of disease development. Bastos et al. [15] demonstrated that filtered cultures of C. amazonense are highly toxic to Phytophthora palmivora spores; they reduce the disease level under field conditions. Similarly, tests conducted by Bastos [29] confirm the inhibition of Crinipellis perniciosa, Microcyclus ulei and Hemileia vastatrix spore germination, as well as the growth of other fungi and of Pseudomonas solanacearum. In turn, Goh et al. [30] found similar results when using Cladobotryum semicirculare as a fungicolous antagonist, which inhibited Rigidoporus microporus mycelial growth by 79% in a direct antagonism test as well as heat stable antifungal Cladobotryum semicirculare filtrates. The authors also observed a reduction in disease severity of white root disease in rubber trees (47 to 50%) by the application of Cladobotryum semicirculare.
To our knowledge, no phytopathogenic fungal control test has been described, neither in vitro nor in vivo, for Cladobotryum mycophilum. Despite the good results, these studies must be continued to determine appropriate doses and modes of application to pathogens and culture environments, and to identify the type of bioactive molecules responsible for such control.
Currently, there is a greater awareness of food safety, and consumers demand more sustainable systems that do not harm the environment and biodiversity. One of the biggest challenges we face in agricultural production is the reduction of risks and negative impacts of phytosanitary products and fertilisers on human health and the environment, while maintaining agricultural productivity and profitability. This is only possible by using new technologies in integrated production systems that reduce waste and minimise environmental impacts. New mycopathogenic microorganisms that can be used as tools for fungal disease control and plant probiotic microorganisms are needed to design new control strategies that help meet present and future needs of agricultural production systems.
5. Conclusions
All Cladobotryum mycophilum isolates have shown the ability to inhibit the mycelial development of eight phytopathogens. In this study, three potential Cladobotryum isolates have been selected for their high capacity to control diseases caused by phytopathogens, both airborne and soil pathogens. This is the first report of this mycopathogen as a beneficial fungus for agriculture.
Author Contributions
F.D. and M.S. conceived and designed the experiments; B.S.-M. and A.M.-G. performed the experiments; F.J.G. and F.D. analysed the data; M.S. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
The present work benefited from the input of the project RTC-2017-6486-2 was supported by the Spanish Ministry of Science, Innovation and Universities.
Conflicts of Interest
The authors declare that there is no conflict of interests regarding the publication of this manuscript.
References
- Fletcher, J.T.; Gaze, R.H. Mushroom Pest and Disease Control; Manson Publishing: London, UK, 2008. [Google Scholar]
- Largeteau, M.L.; Savoie, J.M. Microbially induced diseases of Agaricus bisporus: Biochemical mechanisms and impact on commercial mushroom production. Appl. Microbiol. Biotechnol. 2010, 86, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, J.; Navarro, M.J.; Santos, M.; Diánez, F.; Gea, F.J. Incidence, identification and pathogenicity of Cladobotryum mycophilum, causal agent of cobweb disease on Agaricus bisporus mushroom crops in Spain. Ann. Appl. Biol. 2016, 168, 214–224. [Google Scholar] [CrossRef]
- Chakwiya, A.; Van der Linde, E.J.; Chidamba, L.; Korsten, L. Diversity of Cladobotryum mycophilum isolates associated with cobweb disease of Agaricus bisporus in the south African mushroom industry. Eur J. Plant. Pathol. 2019, 154, 767–776. [Google Scholar] [CrossRef]
- Adie, B.A.T. The Biology and Epidemiology of the Cobweb Disease Pathogen (Cladobotryum Spp.) Infecting the Cultivated Mushroom (Agaricus Bisporus). Ph.D. Thesis, Imperial College, University of London, London, UK, 2000. [Google Scholar]
- Carrasco, J.; Navarro, M.J.; Gea, F.J. Cobweb, a serious pathology in mushroom crops: A review. Span. J. Agric. Res. 2017, 15, e10R01. [Google Scholar] [CrossRef] [Green Version]
- Gea, F.J.; Navarro, M.J.; Suz, L.M. First report of Cladobotryum mycophilum causing cobweb on cultivated king oyster mushroom in Spain. Plant. Dis. 2011, 95, 1030. [Google Scholar] [CrossRef] [PubMed]
- Back, C.G.; Lee, C.Y.; Seo, G.S.; Jung, H.Y. Characterization of species of Cladobotryum which cause cobweb disease in edible mushrooms grown in Korea. Mycobiology 2012, 40, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.K.; Lee, Y.H.; Cho, K.M.; Lee, J.Y. First report of cobweb disease caused by Cladobotryum mycophilum on the edible mushroom Pleurotus eryngii in Korea. Plant. Dis. 2012, 96, 1374. [Google Scholar] [CrossRef]
- Gea, F.J.; Navarro, M.J.; Suz, L.M. Cobweb disease on oyster culinary medicinal mushroom (Pleurotus ostreatus) caused by the mycoparasite Cladobotryum mycophilum. J. Plant. Pathol. 2019, 101, 349–354. [Google Scholar] [CrossRef]
- Carrasco, J.; Navarro, M.J.; Santos, M.; Gea, F.J. Effect of five fungicides with different modes of action on mushroom cobweb disease (Cladobotryum mycophilum) and mushroom yield. Ann. Appl. Biol. 2017, 171, 62–69. [Google Scholar] [CrossRef]
- Barnett, H.L.; Binder, F.L. The fungal host-parasite relationship. Annu. Rev. Phytopathol. 1973, 11, 273–292. [Google Scholar] [CrossRef]
- Rudakov, O.L. Physiological groups in mycophilic fungi. Mycologia 1978, 70, 150–159. [Google Scholar] [CrossRef]
- Rosenheim, J.A.; Kaya, H.K.; Ehler, L.E.; Marois, J.J.; Jaffee, B.A. Intraguild predation among biological-control agents—Theoryand evidence. Biol. Control 1995, 5, 303–335. [Google Scholar] [CrossRef]
- Bastos, C.N.; Evans, H.C.; Samson, R.A. A new hyperparasitic fungus, Cladobotryum amazonense, with potential for control of fungal pathogens of cocoa. Trans. Br. Mycol. Soc. 1981, 77, 273–278. [Google Scholar] [CrossRef]
- Marzuki, N.F.; Goh, Y.K.; Tung, H.J.; Goh, Y.K.; Goh, K.J. Evaluation on the cultural characteristics and antagonistic activities of Cladobotryum semicirculare against Ganoderma boninense in vitro. J. Oil Palm Res. 2015, 27, 326–338. [Google Scholar]
- Ramos, B. Actividad fungicida de cepas de Cladobotryum spp., para el control de hongos patógenos presentes en modalidades productivas de la agricultura urbana. Agrotec. Cuba 2018, 42, 98–99. [Google Scholar]
- Santos, M.; Diánez, F.; González, M.; Tello, J.C. Grape marc compost: Microbial studies and suppression of soilborne mycosis in vegetable seedlings. World J. Microbiol. Biotechnol. 2008, 24, 1493–1505. [Google Scholar] [CrossRef]
- Diánez, F.; Santos, M.; Carretero, F.; Marín, F. Trichoderma saturnisporum, a new biological control agent. J. Sci. Food Agric. 2016, 96, 1934–1944. [Google Scholar] [CrossRef]
- Siozios, S.; Tosi, L.; Ferrarini, A.; Ferrari, A.; Tononi, P.; Bellin, D.; Maurhofer, M.; Gessler, C.; Delledonne, M.; Pertot, I. Transcriptional reprogramming of the mycoparasitic fungus Ampelomyces quisqualis during the powdery mildew host induced germination. Phytopathology 2015, 105, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Gordon, T.C.; Pfender, W.F. Effects of the Mycoparasite Sphaerellopsis filum on Overwintering Survival of Stem Rust in Perennial Ryegrass. Plant. Dis. 2012, 96, 1471–1481. [Google Scholar] [CrossRef] [Green Version]
- Gajera, H.; Domadiya, R.; Patel, S.; Kapopara, M.; Golakiya, B. Molecular mechanism of Trichoderma as bio-control agents against phytopathogen system—A review. Curr. Res. Microbiol. Biotechnol. 2013, 1, 133–142. [Google Scholar]
- Khan, M.R.; Mohiddin, F.A. Trichoderma: Its multifarious utility in crop improvement. In Crop Improvement through Microbial Biotechnology. New and Future Developments in Microbial Biotechnology and Bioengineering; Aligarh Muslim University: Aligarh, India; Srinagar, India, 2018; Chapter 13; pp. 263–291. [Google Scholar]
- Adnan, M.; Islam, W.; Shabbir, A.; Khan, K.A.; Ghramh, H.A.; Huang, Z.; Chen, H.Y.H.; Lu, G. Plant defense against fungal pathogens by antagonistic fungi with Trichoderma in focus. Microb. Pathog. 2019, 129, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Sakemi, S.; Bordner, J.; Decosta, D.L.; Dekker, K.A.; Hirai, H.; Inagaki, T.; Kim, Y.; Sugiura, A.; Sutcliffe, J.A.; Tachikawa, K.; et al. CJ-15,696 and its analogs, new furopyridine antibiotics from the fungus Cladobotryum varium: Fermentation, isolation, structural elucidation, biotransformation and antibacterial activities. J. Antibiot. 2002, 55, 6–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Blunt, J.W.; Cole, A.L.; Cannon, J.F.; Robinson, W.T.; Munro, M.H. Two novel cytotoxic cyclodepsipeptides from a mycoparasitic Cladobotryum sp. J. Org. Chem. 2003, 68, 2002–2005. [Google Scholar] [CrossRef] [PubMed]
- Mitova, M.I.; Lang, G.; Blunt, J.W.; Cummings, N.J.; Cole, A.L.; Robinson, W.T.; Munro, M.H. Cladobotric acids AF: New cytotoxic polyketides from a New Zealand Cladobotryum sp. J. Org. Chem. 2006, 71, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Breinholt, J.; Jensen, H.C.; Kjaer, A.; Olsen, C.E.; Rassing, B.R.; Søtofte, I. Cladobotryal: A fungal metabolite with a novel ring system. Acta Chem. Scand. 1998, 52, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Bastos, C.N. Effect of the culture filtrate of Cladobotryum amazonense on Crinipellis perniciosa (Stahel) Singer and other pathogens. Rev. Theobroma 1984, 14, 263–269. [Google Scholar]
- Goh, Y.K.; Marzuki, Y.N.; Liew, F.A.; Goh, Y.K.; Goh, K.J. Antagonistic effects of fungicolous ascomycetous Cladobotryum Semicirculare on Rigidoporus Microporus White Root Disease in Rubber Trees (Hevea Brasiliensis) under in vitro and Nursery Experiments. J. Rubber Res. 2018, 21, 62–72. [Google Scholar] [CrossRef]
Figure 1.
Mycelial growth of 24 isolates of C. mycophilum in vitro after 4 days. Mean standard deviation is expressed in error bar (n = 5). For each isolate, columns marked with different letters indicate a significant difference according to analyses of variance (p < 0.05).
Figure 1.
Mycelial growth of 24 isolates of C. mycophilum in vitro after 4 days. Mean standard deviation is expressed in error bar (n = 5). For each isolate, columns marked with different letters indicate a significant difference according to analyses of variance (p < 0.05).

Figure 2.
Disease severity of phytopathogens in plants was rated 30–60 days after inoculation based on a 0–4 scale: where 0 = no visible disease symptoms and 4 = plant dead. Mean standard deviation is expressed in error bar (24 plants/repetition). Means with the same letter are not significantly different (LSD) according to ANOVA test (p < 0.05).
Figure 2.
Disease severity of phytopathogens in plants was rated 30–60 days after inoculation based on a 0–4 scale: where 0 = no visible disease symptoms and 4 = plant dead. Mean standard deviation is expressed in error bar (24 plants/repetition). Means with the same letter are not significantly different (LSD) according to ANOVA test (p < 0.05).

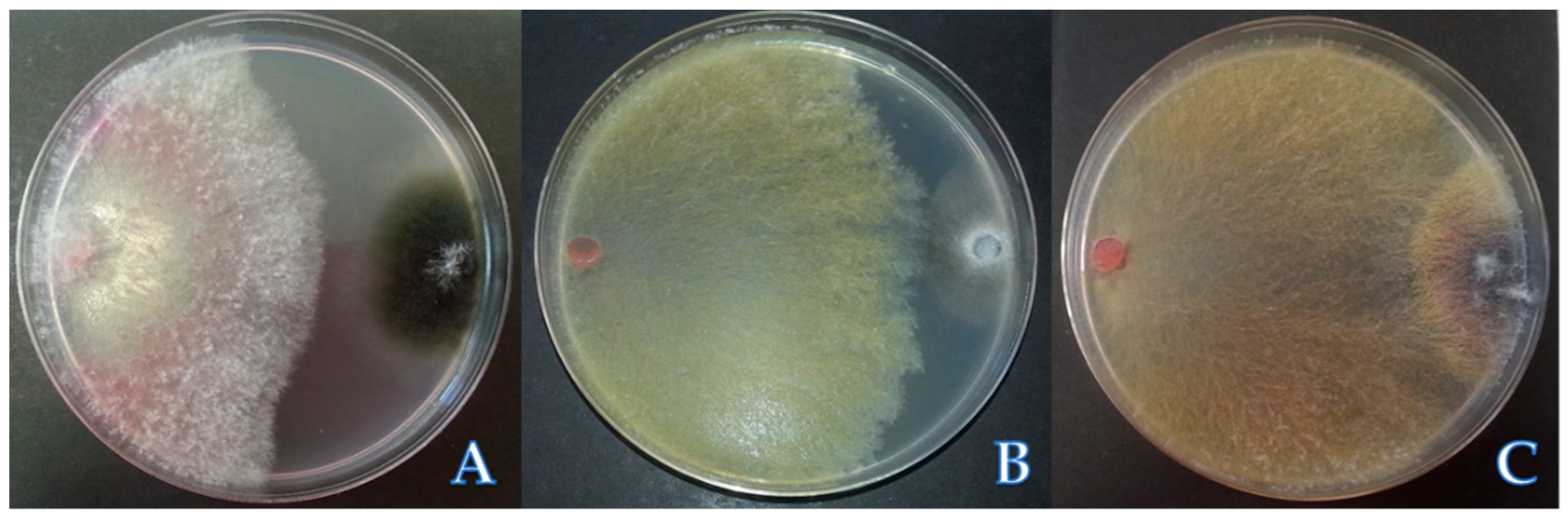
Figure 3.
Antagonistic effects of Cladobotryum isolates against M. melonis (A), P. capsici (B) and F. oxysporum f.sp. lycopersici (C) in dual culture. C show overgrowth of Cladobotryum covering the Fusarium oxysporum f. sp. radicis-lycopersici (FORL) colony after 7 days of inoculation in dual culture.
Figure 3.
Antagonistic effects of Cladobotryum isolates against M. melonis (A), P. capsici (B) and F. oxysporum f.sp. lycopersici (C) in dual culture. C show overgrowth of Cladobotryum covering the Fusarium oxysporum f. sp. radicis-lycopersici (FORL) colony after 7 days of inoculation in dual culture.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Antagonistic potential of Cladobotryum isolates against eight phytopathogens in dual culture on PDA medium. % mycelial inhibition was calculated as PIRG = (R1 − R2) ÷ R1 × 100, where: PIRG: percentage inhibition of radial mycelia growth of the pathogen, R1: radial growth of pathogen in control plates, R2: radial growth of pathogen in dual culture plates. Means with the same letter are not significantly different (LSD) according to ANOVA test (p < 0.05).
Table 1.
Antagonistic potential of Cladobotryum isolates against eight phytopathogens in dual culture on PDA medium. % mycelial inhibition was calculated as PIRG = (R1 − R2) ÷ R1 × 100, where: PIRG: percentage inhibition of radial mycelia growth of the pathogen, R1: radial growth of pathogen in control plates, R2: radial growth of pathogen in dual culture plates. Means with the same letter are not significantly different (LSD) according to ANOVA test (p < 0.05).
| Isolates | % Inhibition of Mycelial Growth | |||||||
|---|---|---|---|---|---|---|---|---|
| Plant Pathogens | ||||||||
| Botrytis cinerea | FORL | Fusarium solani | FOC | Phytophthora parasitica | Mycosphaerella melonis | Phytophthora capsici | Pythium aphanidermatum | |
| CLR1A | 68.25 ± 1.89 cdefg | 72 ± 2.44 bc | 76 ± 2.05 cdefg | 73.75 ± 3.19 abc | 81.25 ± 10.72 cdefg | 86.5 ± 7.68 ab | 66.5 ± 15.72 efg | 49.25 ± 1.42 defg |
| CLR2A | 68.5 ± 1.63 cdefg | 71.25 ± 3.75 bc | 77.5 ± 0.88 cdef | 72.75 ± 2.05 abc | 89.25 ± 0.68 abcdef | 80.5 ± 3.38 abc | 77 ± 2.88 abcd | 48 ± 4.29 efghi |
| CLR2B | 68.75 ± 3.53 cdefg | 71.5 ± 1.05 bc | 79.25 ± 1.12 bcd | 72.25 ± 2.40 abc | 89.5 ± 2.09 abcde | 72 ± 1.68 cd | 74 ± 1.85 bcde | 51.5 ± 2.05 cdef |
| CL5A | 58.75 ± 12.37 hi | 47.75 ± 6.09 ef | 70.25 ± 6.52 h | 46.5 ± 7.97 g | 71.75 ± 11.20 h | 60.62 ± 11.39 e | 63 ± 14.19 fgh | 45.75 ± 3.25 ghijk |
| CL6A | 73.5 ± 3.23 abc | 76.25 ± 2.34 ab | 82.5 ± 2.17 ab | 78.25 ± 4.20 a | 91 ± 2.85 ab | 79.25 ± 3.26 abc | 77.25 ± 1.63 abcd | 53.25 ± 5.19 bcde |
| CL7A | 76.25 ± 1.97 ab | 73.25 ± 1.12 ab | 80 ± 1.25 abc | 76 ± 1.63 ab | 91 ± 1.05 ab | 78.25 ± 4.11 bc | 79.75 ± 3.24 ab | 53.5 ± 2.40 bcd |
| CL11A | 74.5 ± 2.87 abc | 75.75 ± 1.90 ab | 79.75 ± 1.05 abcd | 73 ± 2.44 abc | 92 ± 4.73 ab | 87.5 ± 4.76 ab | 77.75 ± 2.24 abc | 55.25 ± 4.54 abc |
| CL14A | 79.25 ± 12.00 a | 73.75 ± 3.95 ab | 74.75 ± 3.69 efg | 75.75 ± 1.68 ab | 91 ± 2.71 ab | 86.75 ± 1.90 ab | 80.75 ± 1.43 ab | 49 ± 9.28 defg |
| CL15A | 62.75 ± 6.33 gh | 46.75 ± 9.04 ef | 74.75 ± 4.37 efg | 59.75 ± 4.28 f | 80.31 ± 14.10 bcdef | 54 ± 16.52 ef | 65.25 ± 14.43 fg | 43.25 ± 4.72 ijkl |
| CL17A | 73.25 ± 0.68 abc | 80 ± 0.88 a | 83 ± 2.09 ab | 79 ± 0.56 a | 93.75 ± 1.25 a | 85.25 ± 1.37 ab | 83.25 ± 3.60 a | 59.25 ± 2.43 a |
| CL18A | 72.5 ± 1.76 bcd | 76.5 ± 1.63 ab | 79.5 ± 4.11 abcd | 76.25 ± 1.98 a | 91.75 ± 0.68 ab | 85.5 ± 2.59 ab | 81.25 ± 1.53 ab | 58.25 ± 1.42 ab |
| CL19A | 64.25 ± 1.67 fgh | 32.18 ± 2.13 h | 74.5 ± 2.88 efg | 30.83 ± 1.91 h | 72.5 ± 5.00 gh | 38.25 ± 2.44 g | 55.25 ± 10.73 h | 36.25 ± 8.14 m |
| CL40A | 71.5 ± 1.04 bcde | 48.5 ± 2.40 ef | 79.75 ± 2.24 abcd | 75.25 ± 1.63 ab | 90 ± 3.75 abcd | 62.5 ± 5.80 de | 74.25 ± 1.12 bcde | 48.5 ± 2.40 defghi |
| CL41A | 47.25 ± 10.24 j | 42 ± 12.39 fg | 72 ± 3.60 gh | 32.75 ± 1.63 h | 66.67 ± 18.76 fgh | 46.75 ± 14.16 fg | 62.75 ± 8.45 fgh | 21 ± 1.62 n |
| CL43A | 70 ± 2.16 bcdef | 61.25 ± 18.54 d | 79 ± 3.58 bcd | 60 ± 20.54 ef | 92 ± 2.27 ab | 80.25 ± 3.79 abc | 76 ± 6.93 abcd | 53.5 ± 7.72 bcd |
| CL45A | 53 ± 5.49 ij | 51 ± 3.79 e | 70.25 ± 2.71 h | 67 ± 6.47 cde | 80.75 ± 5.90 defgh | 64 ± 5.89 de | 67 ± 3.81 efg | 38.5 ± 4.08 lm |
| CL46A | 65.75 ± 3.37 efg | 53.5 ± 9.82 e | 76.5 ± 5.89 cdef | 64 ± 2.85 def | 86.25 ± 9.80 abcdef | 54.75 ± 10.66 ef | 60 ± 11.28 gh | 40.5 ± 4.80 klm |
| CL2A | 71.5 ± 3.46 bcde | 72 ± 1.43 bc | 77.75 ± 1.37 cdef | 74 ± 1.85 ab | 90.75 ± 2.74 ab | 88.75 ± 4.59 a | 79.25 ± 3.49 abc | 48.75 ± 4.05 defgh |
| CL30A | 68 ± 1.89 cdefg | 75 ± 1.25 ab | 79.25 ± 1.43 bcd | 75.5 ± 2.09 ab | 90.5 ± 0.68 abc | 82 ± 3.38 abc | 78.75 ± 1.53 abc | 51.5 ± 1.04 cdef |
| CL55A | 70.5 ± 8.50 bcdef | 70.25 ± 2.85 bc | 78.25 ± 6.29 cde | 72.75 ± 3.35 abc | 87.75 ± 1.05 abcdef | 77.75 ± 3.99 bc | 76 ± 2.05 abcd | 47 ± 3.81 fghij |
| CL60A | 76.25 ± 1.97 ab | 76.5 ± 3.47 ab | 83.5 ± 4.18 a | 76.75 ± 1.12 a | 91.5 ± 1.85 ab | 86.75 ± 1.90 ab | 80 ± 2.34 ab | 57.75 ± 1.85 ab |
| CL80A | 68.75 ± 4.33 cdefg | 35.5 ± 5.20 gh | 75.75 ± 1.43 defg | 69 ± 6.34 bcd | 88.75 ± 11.39 abcdef | 61 ± 22.73 e | 71 ± 1.63 cdef | 46.25 ± 4.23 fghij |
| CL1264 | 70.25 ± 1.85 bcdef | 74 ± 1.85 ab | 73.75 ± 4.15 fgh | 75.25 ± 1.37 ab | 93.25 ± 2.44 ab | 80.75 ± 3.71 abc | 77 ± 1.90 abcd | 42.25 ± 4.08 jkl |
| CL1381 | 66.25 ± 1.25 defg | 65 ± 2.05 cd | 74.75 ± 1.05 efg | 46.75 ± 2.05 g | 80.25 ± 3.47 efgh | 62.25 ± 2.05 de | 69 ± 2.05 def | 43.5 ± 2.23 hijkl |
| p | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Santos, M.; Diánez, F.; Moreno-Gavíra, A.; Sánchez-Montesinos, B.; Gea, F.J. Cladobotryum mycophilum as Potential Biocontrol Agent. Agronomy 2019, 9, 891. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120891
AMA Style
Santos M, Diánez F, Moreno-Gavíra A, Sánchez-Montesinos B, Gea FJ. Cladobotryum mycophilum as Potential Biocontrol Agent. Agronomy. 2019; 9(12):891. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120891
Chicago/Turabian StyleSantos, Mila, Fernando Diánez, Alejandro Moreno-Gavíra, Brenda Sánchez-Montesinos, and Francisco J. Gea. 2019. "Cladobotryum mycophilum as Potential Biocontrol Agent" Agronomy 9, no. 12: 891. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120891
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.