May the Inclusion of a Legume Crop Change Weed Composition in Cereal Fields? Example of Sainfoin in Aragon (Spain)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
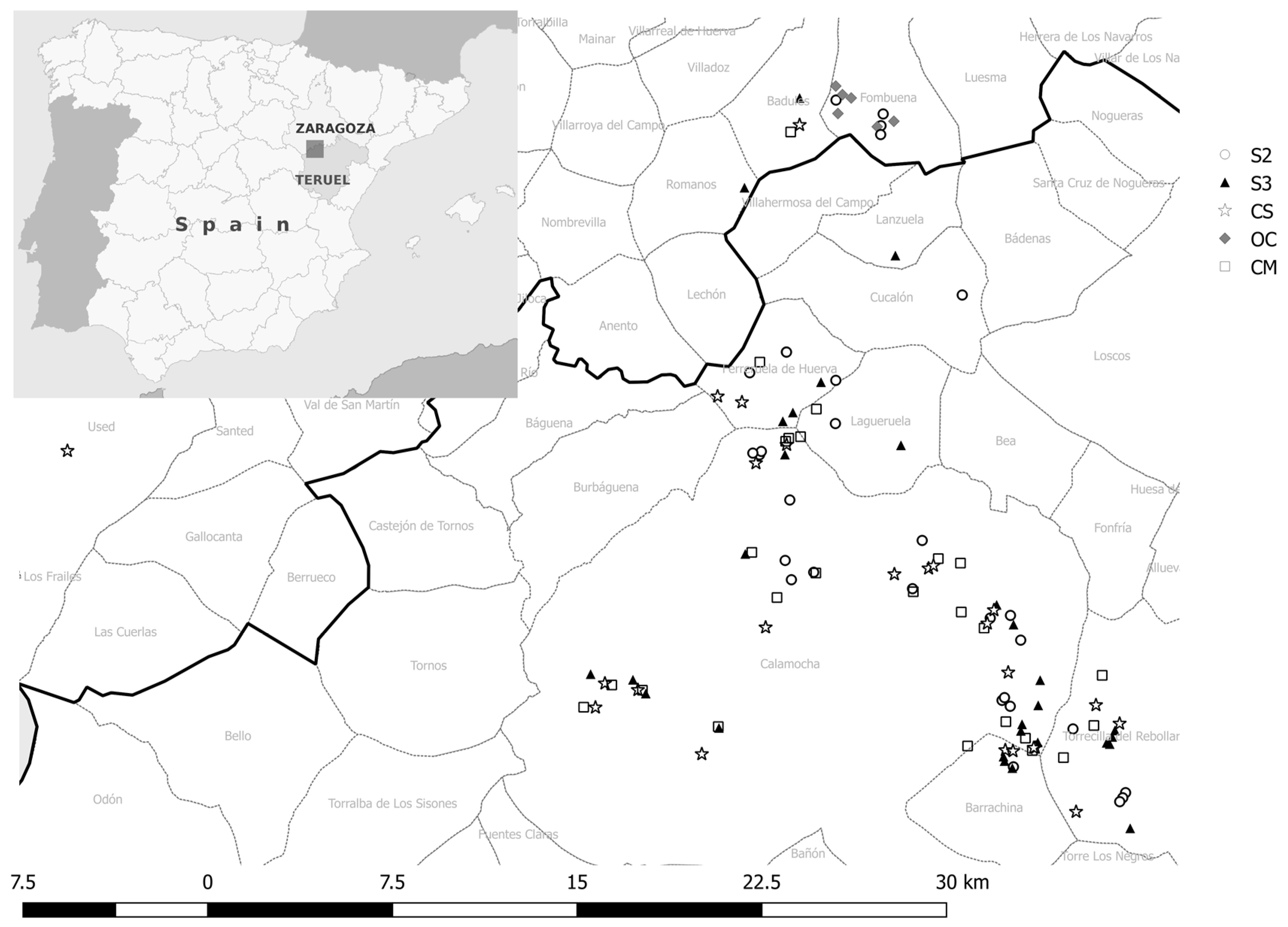
2.1. Field Surveys
2.2. Seed Bank Determination
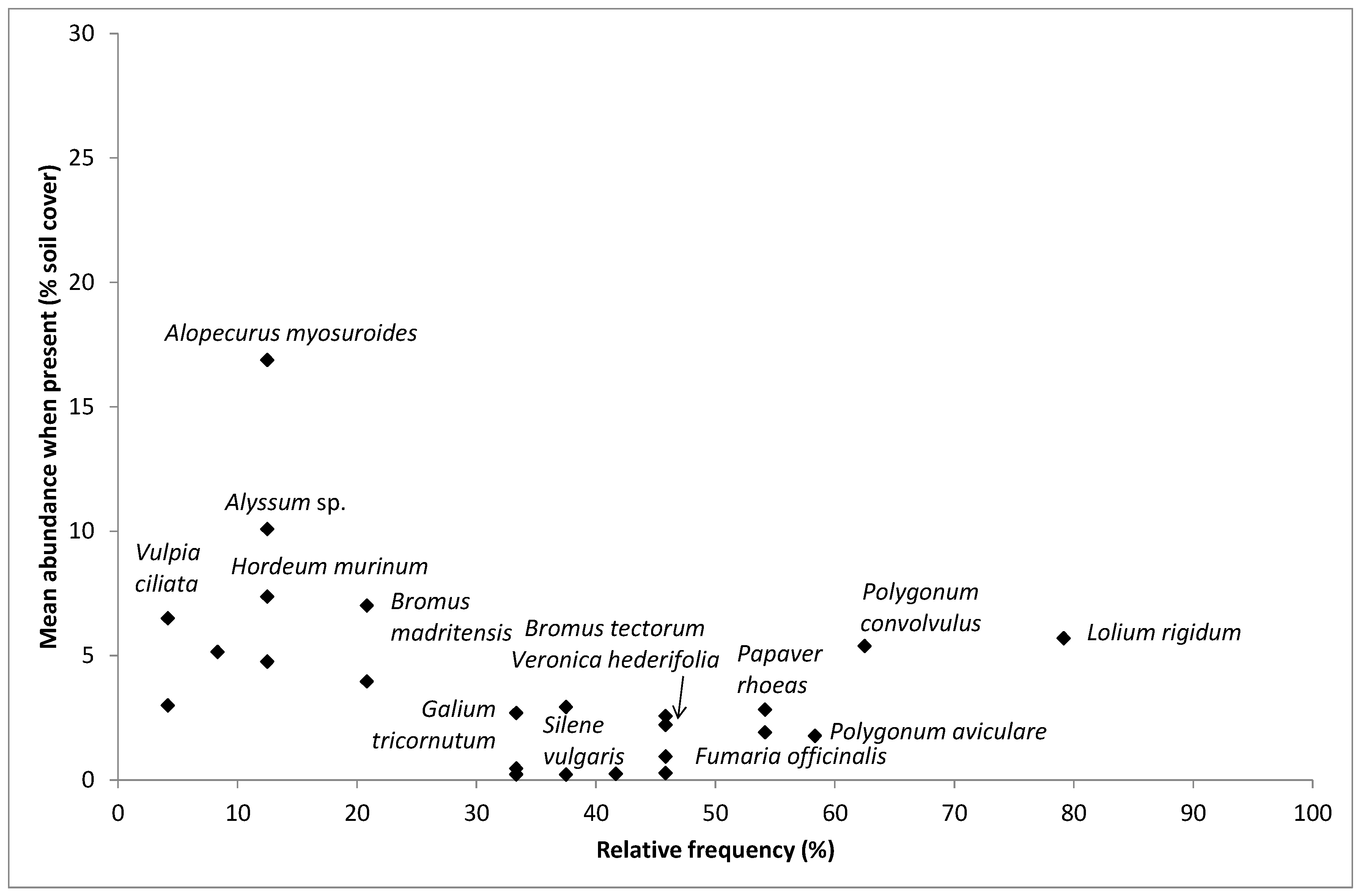
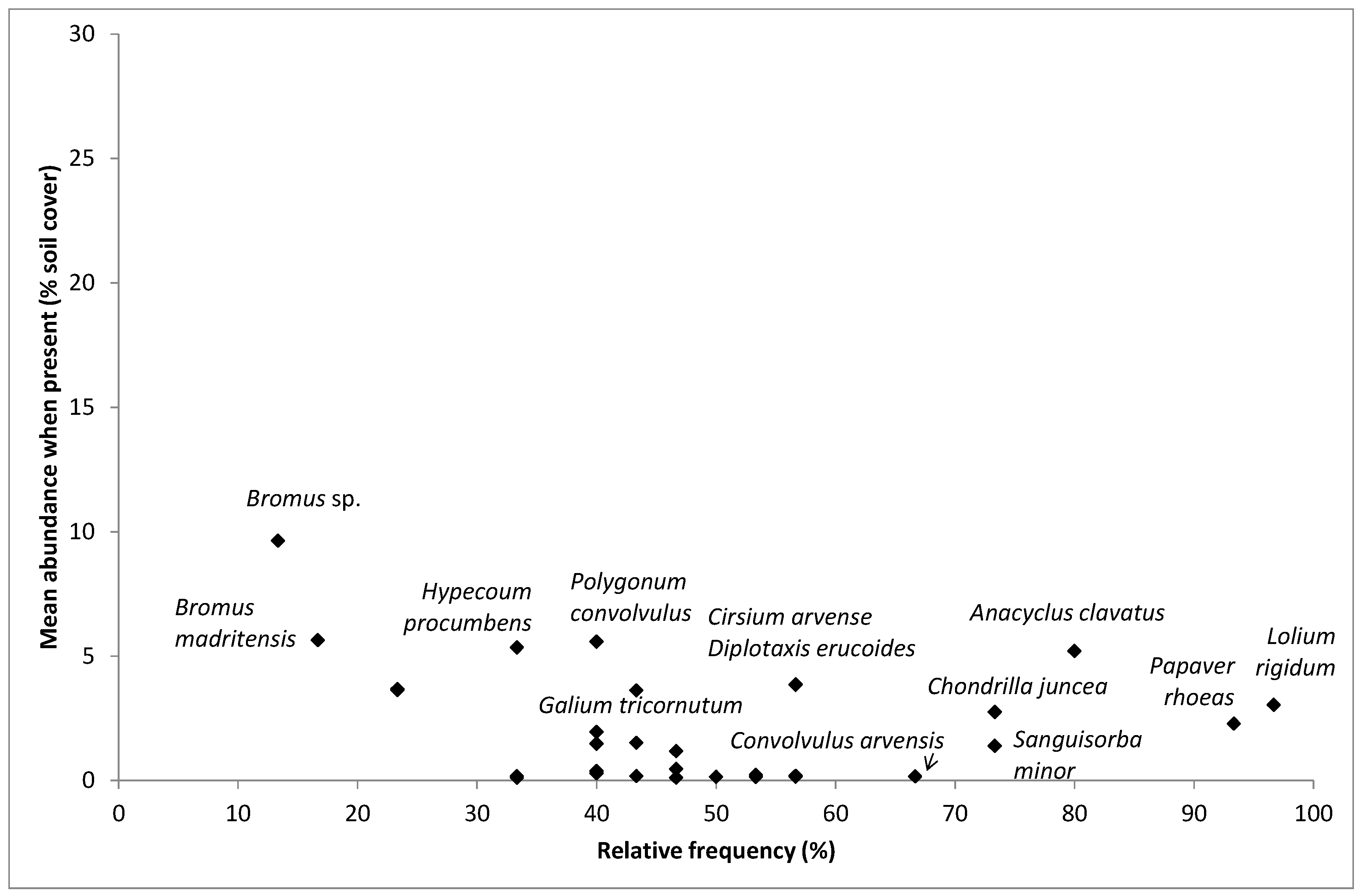
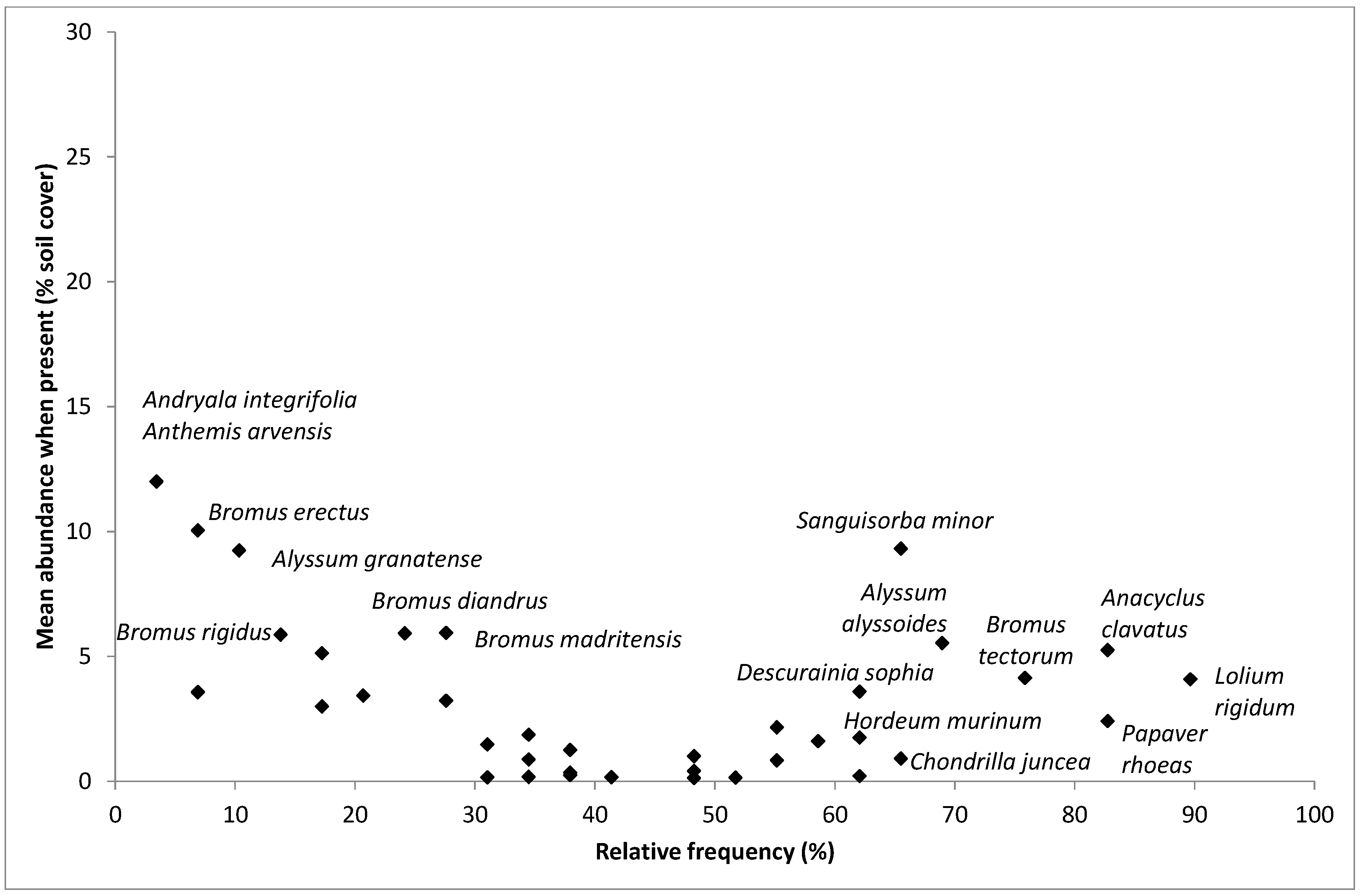
2.3. Weed Species Composition: Most Frequent and Most Abundant Weeds, Richness, Cover, Shannon Index and Multivariate Analysis
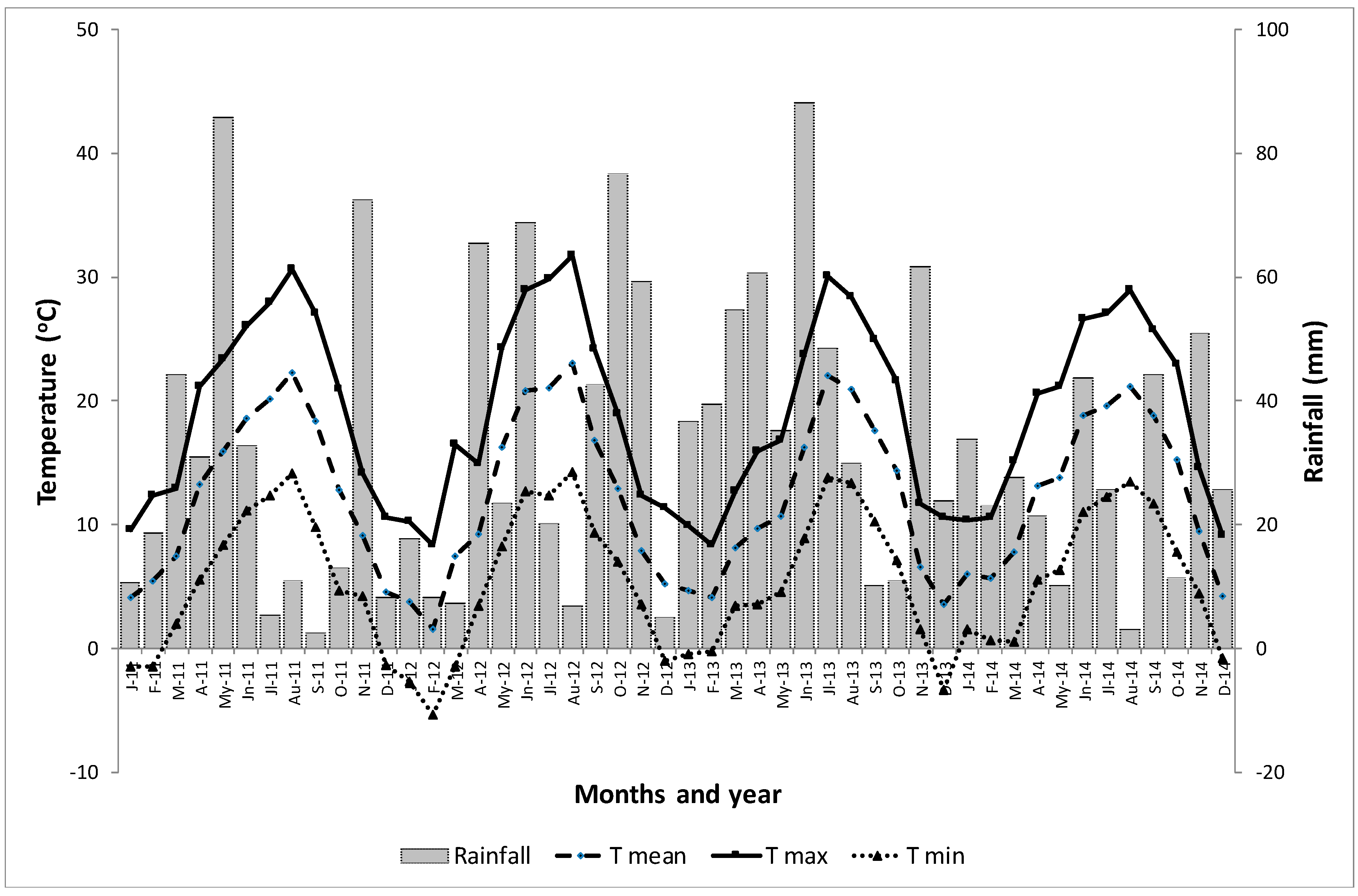
2.4. Climatic Data
3. Results
3.1. Species Richness, Diversity and Weed Cover
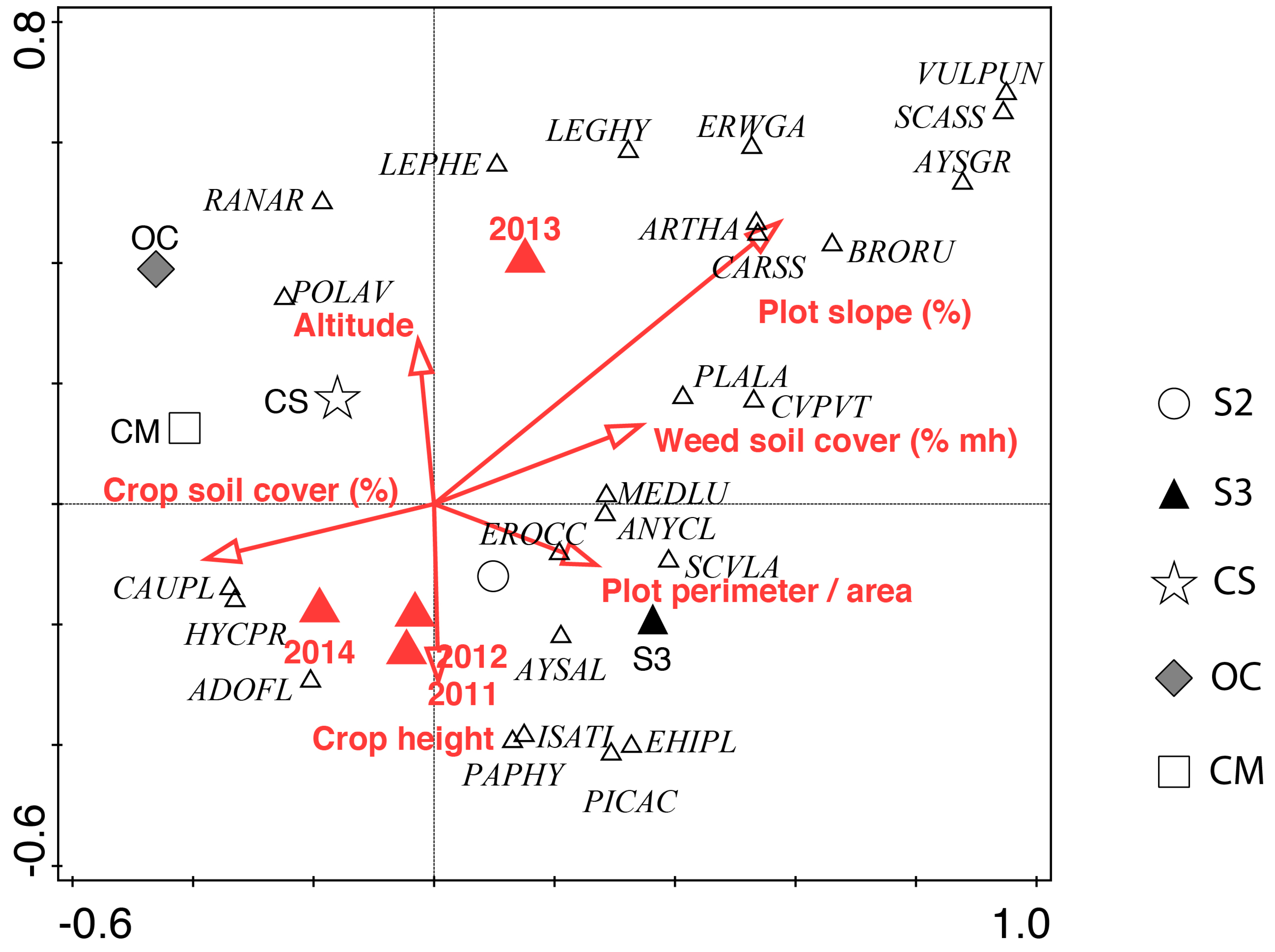
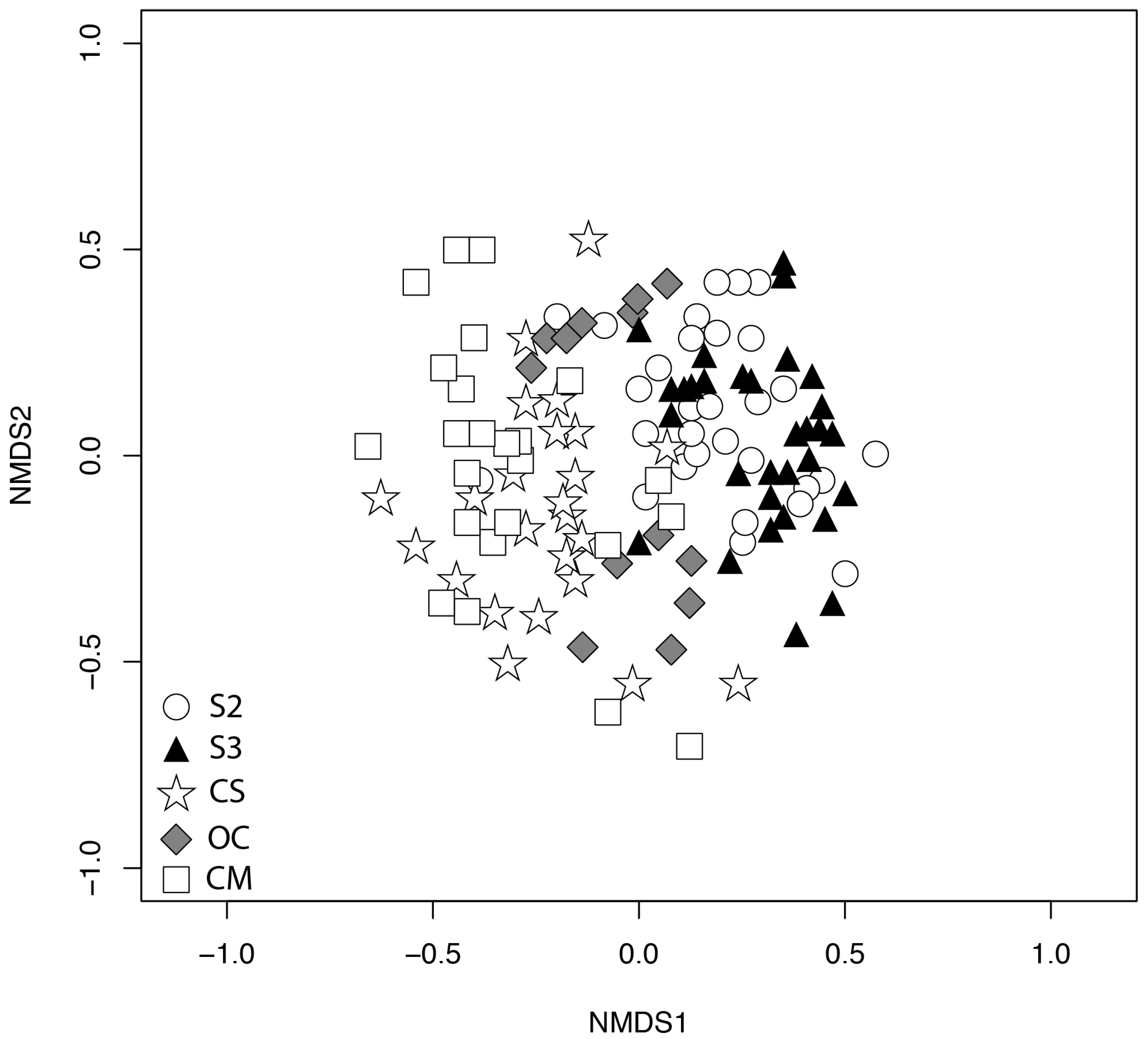
3.2. Weed Community Composition
3.3. Crop Soil Cover and Height
3.4. Soil Seedbank
4. Discussion
4.1. Species Richness and Weed Cover
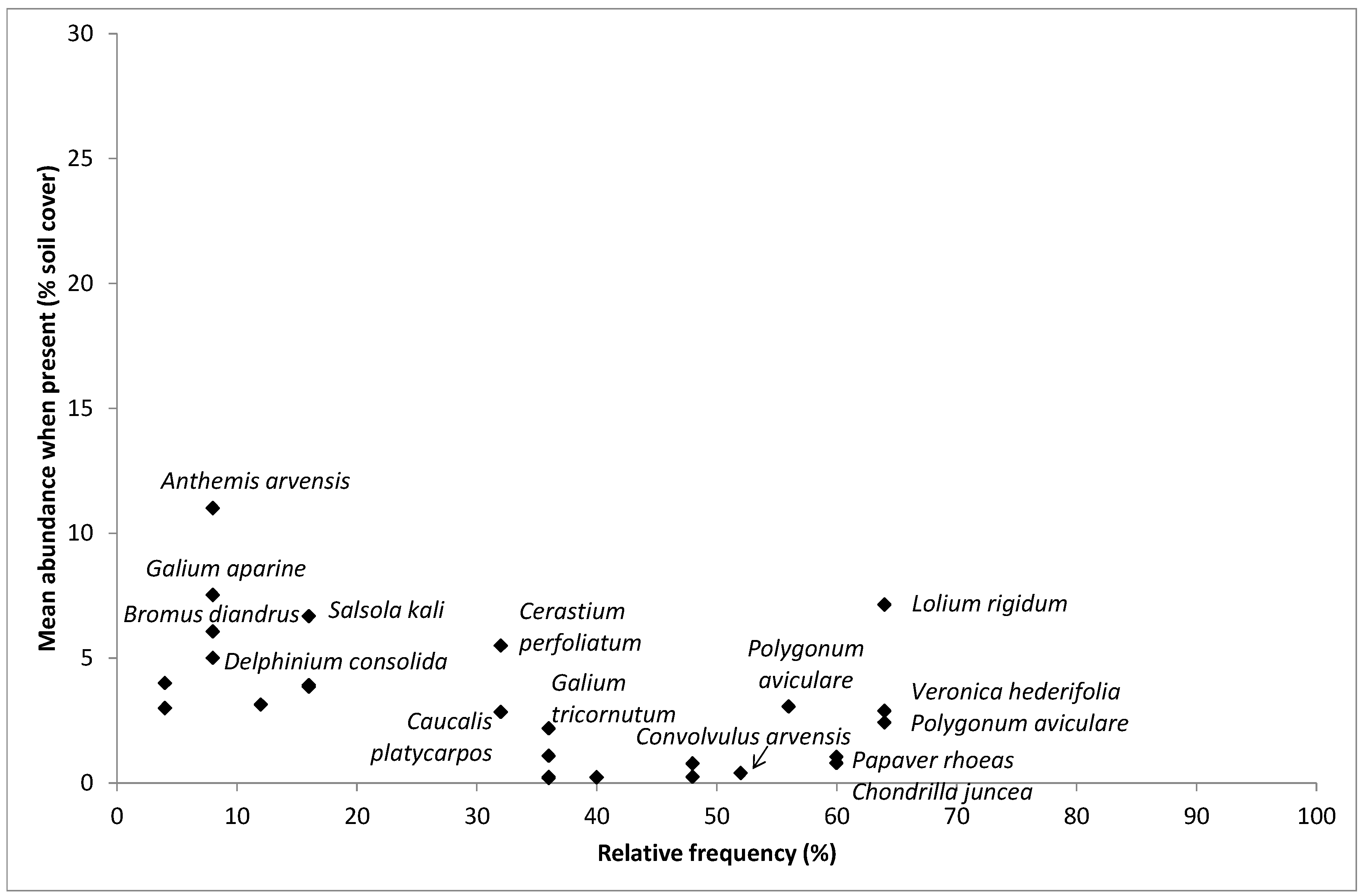
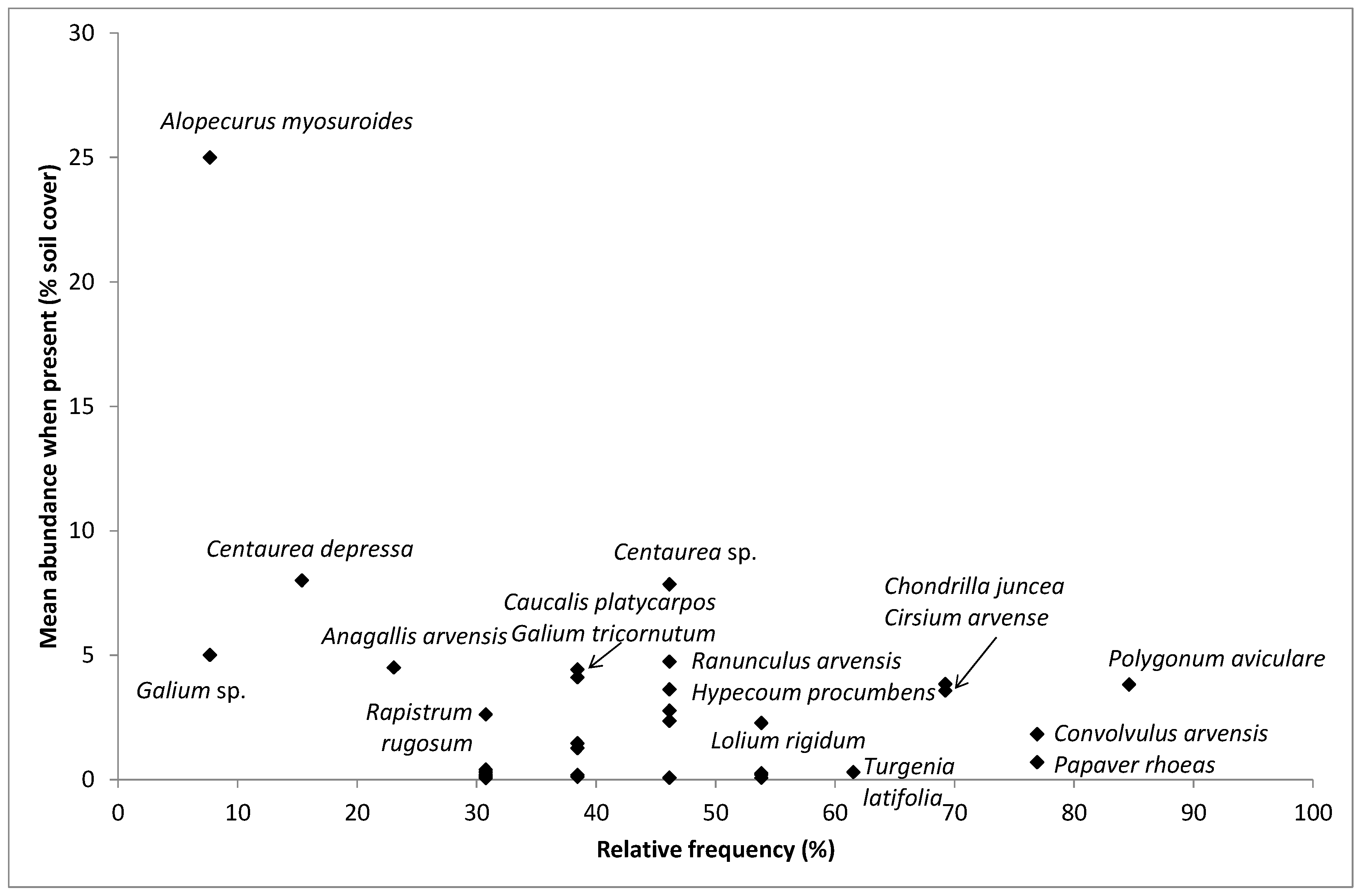
4.2. Most Frequent and Most Abundant Species
4.3. Species Composition
4.4. Soil Seedbank
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kallenbach, R.L.; Matches, A.G.; Mahan, J.R. Sainfoin regrowth declines as metabolic rate increases with temperature. Crop Sci. 1996, 36, 91–97. [Google Scholar] [CrossRef]
- Meyer, D.W.; Badaruddin, M. Frost tolerance of ten seedling legume species at four growth stages. Crop Sci. 2001, 41, 1838–1842. [Google Scholar] [CrossRef]
- Molle, G.; Decandia, M.; Solter, U.; Greef, J.M.; Rochon, J.J.; Sitzia, M.; Hopkins, A.; Rook, A.J. The effect of different legume-based swards on intake and performance of grazing ruminants under Mediterranean and cool temperate conditions. Grass Forage Sci. 2008, 63, 513–530. [Google Scholar] [CrossRef]
- Mora-Ortiz, M.; Smith, L.M.J. Onobrychis viciifolia: A comprehensive literatura review of its histroy, etymology, taxonomy, genetics, agronomy and botany. Plant Genet. Resour.-C 2018, 16, 403–418. [Google Scholar] [CrossRef]
- Carbonero, C.H.; Mueller-Harvey, I.; Brown, T.A.; Smith, L. Sainfoin (Onobrychis viciifolia): A beneficial forage legume. Plant Genet. Resour.-C 2011, 9, 70–85. [Google Scholar] [CrossRef]
- Naseri, B.; Alizadeh, M.A. Climate, powdery mildew, sainfoin resistance and yield. J. Plant. Pathol. 2017, 99, 619–625. [Google Scholar]
- Hwang, D.F.; Gaudet, D.A. Effects of plant age and late-season hardening on development of resistance to winter crown rot in first-year sainfoin (Onobrychis viciifolia). J. Plant. Dis. Protect. 1999, 106, 188–197. [Google Scholar]
- Tozlu, G.; Hayat, R. Biology of Cholorophorus robustior Pic, 1900 (Coleoptera: Cerambycidae), a new record and a new sainfoin (Onobrychis sativa Lam.) pest for Turkey. Isr. J. Zool. 2000, 46, 201–206. [Google Scholar] [CrossRef]
- Sintim, H.Y.; Adjesiwor, A.T.; Zheljazkov, V.D.; Islam, M.A.; Obour, A.K. Nitrogen application in sainfoin under rain-fed conditions in Wyoming: Productivity and cost implications. Agron. J. 2016, 108, 294–300. [Google Scholar] [CrossRef]
- MAPAMA. Anuario de Estadísticas. Avance 2016. Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente. 2017. Available online: http://www.mapama.gob.es/estadistica/pags/anuario/2016-Avance/avance/AvAE16.pdf. (accessed on 26 June 2017).
- Delgado, I.; Andrés, C.; Sin, E.; Ochoa, M.J. Estado actual del cultivo de la esparceta (Onobrychis viciifolia Scop.). Encuesta realizada a agricultores productores de semilla. Pastos 2002, 32, 235–247. [Google Scholar]
- BOA. Boletín Oficial de Aragon. 2015. Available online: http://www.boa.aragon.es/cgi-bin/EBOA/BRSCGI?CMD=VEROBJ&MLKOB=840111824848 (accessed on 12 September 2016).
- Marí, A.I.; Cirujeda, A.; Murillo, S.; Pardo, G.; Aibar, J. Resultado de las encuestas realizadas sobre el cultivo de esparceta (Onobrychis viciifolia Scop.) y su efecto sobre las malas hierbas en la zona de Teruel. In Proceedings of the XV Congress of the Spanish Weed Science Society (SEMh), Sevilla, Spain, 19–22 October 2015. [Google Scholar]
- Legendre, H.; Hoste, H.; Gidenne, T. Nutritive value and anthelmintic effect of sainfoin pellets fed to experimentally infected growing rabbits. Animal 2017, 11, 1464–1471. [Google Scholar] [CrossRef]
- Bhattarai, S.; Coulman, B.; Biligetu, B. Sainfoin (Onobrychis viciifolia Scop.): Renewed interest as a forage legume for western Canada. Can. J. Plant Sci. 2016, 96, 748–756. [Google Scholar] [CrossRef]
- Huyen, N.T.; Desrues, O.; Alferink, S.J.J.; Zandstra, T.; Verstegen, M.W.A.; Hendriks, W.H.; Pellikaan, W.F. Inclusion of sainfoin (Onobrychis viciifolia) silage in dairy cow rations affects nutrient digestibility, nitrogen utilization, energy balance, and methane emissions. J. Dairy Sci. 2016, 99, 3566–3577. [Google Scholar] [CrossRef]
- Heckendorn, F.; Werne, S.; Maurer, V.; Perler, E.; Amsler, Z.; Probst, J.; Zaugg, C.; Krenmeyr, I. Sainfoin—New data on anthelmintic effects and production in sheep and goats. Planta Med. 2013, 79, 1125. [Google Scholar] [CrossRef]
- Saratsis, A.; Voutzourakis, N.; Theodosiou, T.; Stefanakis, A.; Sotiraki, S. The effect of sainfoin (Onobrychis viciifolia) and carob pods (Ceratonia siliqua) feeding regimes on the control of lamb coccidiosis. Parasitol. Res. 2016, 115, 2233–2242. [Google Scholar] [CrossRef]
- Baldinger, L.; Hagmuller, W.; Minihuber, U.; Matzner, M.; Zollitsch, W. Sainfoin seeds in organic diets for weaned piglets utilizing the protein-rich grains of a long-known forage legume. Ren. Agric. Food Syst. 2016, 31, 12–21. [Google Scholar] [CrossRef]
- Kashevarov, N.I.; Polyudina, R.I.; Rozhanskaya, O.A.; Jeleznov, A.V. Sainfoin (Onobrychis Mill.) breeding for forage production in Siberia. Kormoproizvodstvo 2013, 9, 22–24. [Google Scholar]
- Aufrere, J.; Theodoridou, K.; Baumont, R.; Baumont, R. Agronomic and nutritional value of sainfoin. Fourrages 2013, 213, 63–75. [Google Scholar]
- Marinov-Serafimov, P.; Golubinova, I.; Vasileva, V. Dynamics and distribution of weed species in weed associations. Indian J. Agric. Sci. 2019, 89, 105–110. [Google Scholar]
- Liu, Z.; Baines, R.N.; Lane, G.P.F.; Davies, W.P. Survival of plants of common sainfoin (Onobrychis viciifolia Scop.) in competition with two companion grass species. Grass Forage Sci. 2010, 65, 11–14. [Google Scholar] [CrossRef]
- Kempf, K.; Grieder, C.; Walter, A.; Widmer, F.; Reinhard, S.; Kolliker, R. Evidence and consequences of self-fertilisation in the predominantly outbreeding forage legume Onobrychis viciifolia. BCM Genet. 2015, 16. [Google Scholar] [CrossRef]
- Delgado, I.; Salvia, J.; Buil, I.; Andrés, C. The agronomic varibaility of a collection of sainfoin accessions. Span. J. Agric. Res. 2008, 63, 401–407. [Google Scholar] [CrossRef]
- Sebastià, M.; Palero, N.; De Bello, F. Changes in Management modify agro-diversity in sainfoin swards in the Eastern Pyrenees. Agron. Sustain. Dev. 2011, 341, 533–540. [Google Scholar]
- Cirujeda, A.; Marí, A.; Murillo, S.; Aibar, J. La flora arvense en el cultivo de la esparceta (Onobrychis viciifolia L.) aumenta la biodiversidad vegetal. In Proceedings of the XIV Congress of the Spanish Weed Science Society (SEMh), Valencia, Spain, 5–7 November 2013; Osca, J.M., Gómez de Barreda, D., Castell, V., Pascual, N., Eds.; Universitat Politécnica de València: Valencia, Spain, 2013; pp. 31–35. [Google Scholar]
- Anderson, R.L. Managing weeds with a dualistic approach of prevention and control. A review. Agron Sustain. Dev. 2007, 27, 13–18. [Google Scholar] [CrossRef]
- BBCH Working Group. Compendium of Growth Stage Identification Keys for Mono- and Dicotyledonous Plants. Extended BBCH Scale; BBA, BSA, IGZ, IVA, AGREVO, BASF, Bayer, Novartis, Eds.; Allcomm Business Communication: Bezug, Germany, 1997. [Google Scholar]
- Walker, K.J.; Critchley, C.N.R.; Sherwood, A.J.; Large, R.; Nuttall, P.; Hulmes, S.; Rose, R.; Mountford, J.O. The conservation on arable plants on cereal field margins: An assessment of new agri-environment scheme options in England, UK. Biol. Conserv. 2007, 136, 260–270. [Google Scholar] [CrossRef]
- Marnotte, P. Influence des facteurs agroécologiques sur le developpement des mauvaises herbes en climat tropical humide. In Proceedings of the 1984 7ème Colloque International Ecologie, Biologie et Systematique des Mauvaises Herbes, Paris, France, 9–11 October 1984; pp. 183–188. [Google Scholar]
- Cirujeda, A.; Aibar, J.; Zaragoza, C. Remarkable changes of weed species in Spanish cereal fields from 1976 to 2007. Agron. Sustain. Dev. 2011, 31, 675–688. [Google Scholar] [CrossRef]
- SigPac database, 3.2. Available online: http://sigpac.mapama.gob.es/fega/visor/ (accessed on 28 August 2014).
- Reinhardt, T.; Leon, R.G. Extractable and germinable seedbank methods provide different quantifications of weed communities. Weed Sci. 2018, 66, 715–720. [Google Scholar] [CrossRef]
- Hanf, M. The Arable Weeds of Europe with Their Seedlings and Seeds; BASF: Ludwigshafen, Germany, 1983; pp. 1–494. [Google Scholar]
- Holm-Nielsen, C. Frø fra det Dyrkede Land. 258 Plantearter; Forskningscenter Flakkebjerg: Slagelse, Denmark, 1998; pp. 1–178. ISBN 87-984996-0-2. [Google Scholar]
- Viaggiani, P. Erbe Spontanee e Infestanti: Technique di Risonoscimento: Dicotiledoni; Bayer: Milano, Italy, 1990; pp. 1–271. [Google Scholar]
- MAPAMA (Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente). Guía de Gestión Integrada de Plagas. Cereales de Invierno. 2015. Available online: http://www.magrama.gob.es/es/agricultura/temas/sanidad-vegetal/productos-fitosanitarios/guias-gestion-plagas/ (accessed on 12 September 2016).
- Lloveras, J.; Melines, M.A. La calidad en la alfalfa. Posibles clasificaciones. Vida Rural 2015, 212, 36–40. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; pp. 145–149. ISBN 978-0691084916. [Google Scholar]
- SAS Institute. SAS Systems for Linear Models; SAS Series in Statistical Applications; SAS Institute: Cary, NC, USA, 1991. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 20 February 2019).
- Smilauer, P.; Leps, J. Multivariate Analysis of Ecological Data using Canoco 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; pp. 1–362. ISBN 978-0-521-89108-0. [Google Scholar]
- La garita del Jiloca. Available online: http://tiempocalamocha.blogspot.com.es (accessed on 5 June 2017).
- Storkey, J.; Neve, P. What good is weed diversity? Weed Res. 2018, 58, 239–243. [Google Scholar] [CrossRef]
- Mayerová, M.; Mikulka, J.; Soukup, J. Effects of selective herbicide treatment on weed community in cereal crop rotation. Plant Soil Environ. 2018, 64, 413–420. [Google Scholar] [CrossRef]
- Stevovic, V.; Stanisavljevic, R.; Djukic, D.; Djurovic, D. Effect of row spacing on seed and forage yield in sainfoin (Onobrychis viciifolia Scop.) cultivars. Turk. J. Agric. For. 2012, 36, 35–44. [Google Scholar]
- Chamorro, L.; Masalles, R.M.; Sans, F.X. Arable weed decline in Northeast Spain: Does organic farming recover functional biodiversity. Agric. Ecosyst. Environ. 2016, 223, 1–9. [Google Scholar] [CrossRef]
- Romero, A.; Chamorro, L.; Sans, F.X. Weed diversity in crop edges and inner fields of organic and conventional dryland Winter cereal crops in NE Spain. Agric. Ecosyst. Environ. 2008, 124, 97–104. [Google Scholar] [CrossRef]
- Sopeña, J.M.; Zaragoza, C. Principales problemas de malas hierbas y herbicidas plantados durante 1997 en Aragon. In Proceedings of the XVII Meeting of the Spanish Weed Working Group, Santa Cruz de Tenerife, Spain, 3–6 March 1998; p. 118. [Google Scholar]
- Cirujeda, A.; Taberner, A. Cultural control of herbicide-resistant Lolium rigidum Gaud. populations in winter cereal in Northeastern Spain. Span. J. Agric. Res. 2009, 7, 146–154. [Google Scholar] [CrossRef]
- Cirujeda, A.; Recasens, J.; Taberner, A. Effect of ploughing and harrowing on a herbicide resistant corn poppy (Papaver rhoeas) population. Biol. Agric. Hortic. 2003, 21, 231–246. [Google Scholar] [CrossRef]
- Navarrete, L.; Sánchez, M.J.; Alarcón, R.; Hernanz, J.L.; Sánchez-Girón, V. Respuesta de los cultivos y la vegetación arvense a la reducción de la fertilización y al tipo de laboreo en sistemas cerealistas de secano. In Proceedings of the XV Congress of the Spanish Weed Science Society (SEMh), Sevilla, Spain, 19–22 October 2015. [Google Scholar]
- Gerowitt, B.; Heitefuss, R. Weed economic thresholds in cereals in the Federal Republic of Germany. Crop Prot. 1990, 9, 323–331. [Google Scholar] [CrossRef]
- Solé-Senan, X.O.; Juárez-Escario, A.; Conesa, J.A.; Torra, J.; Royo-Esnal, A.; Recasens, J. Plant diversity in mediterranean cereal fields: Unraveling the effect of landscape complexity on rare arable plants. Agric. Ecosyst. Environ. 2014, 185, 221–230. [Google Scholar] [CrossRef]
- Solé-Senan, X.O.; Juárez-Escario, A.; Robleño, I.; Conesa, J.A.; Recasens, J. Using the response-effect trait framework to disentangle the effects of agricultural intensification on the provision of ecosystem services by mediterranean arable plants. Agric. Ecosyst. Environ. 2017, 247, 255–264. [Google Scholar] [CrossRef]
- Pinke, G.; Gunton, R. Refining rare weed trait syndromes along arable intensification gradients. J. Veg. Sci. 2014, 25, 978–989. [Google Scholar] [CrossRef]
- Solé-Senan, X.O.; Juárez-Escario, A.; Conesa, J.A.; Recasens, J. Plant species, functional assemblages and partitioning of diversity in a mediterranean agricultural mosaic landscape. Agric. Ecosyst. Environ. 2018, 256, 163–172. [Google Scholar] [CrossRef]
- Burnside, O.C.; Wilson, R.G.; Weisberg, S.; Hubbard, K.G. Seed longevity of 41 weed species buried 17 years in Eastern and Western Nebraska. Weed Sci. 1996, 44, 74–86. [Google Scholar]
- Conn, J.S.; Beattle, K.L.; Blanchard, A. Seed viability and dormancy of 17 weed species after 19.7 years of burial in Alaska. Weed Sci. 2006, 54, 464–470. [Google Scholar] [CrossRef]
- Baraibar, B.; Torra, J.; Westermann, P. Harvester ant (Messor barbarus (L.)) density as related to soil properties, topography and management in semi-arid cereals. Appl. Soil Ecol. 2011, 51, 60–65. [Google Scholar] [CrossRef]
- Baraibar, B.; Carrión, E.; Recasens, J.; Westermann, P.R. Unravelling the process of weed seed predation: Developing options for better weed control. Biol. Control 2011, 56, 85–90. [Google Scholar] [CrossRef]
- Harradine, A.R. Seed longevity and seedling establishment of Bromus diandrus Roth. Weed Res. 1986, 26, 173–180. [Google Scholar] [CrossRef]
- Haring, S.C.; Flessner, M.L. Improving soil seed bank management. Pest Manag. Sci. 2018, 74, 2412–2418. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | p-Value (Species Richness) | p-Value (Shannon) |
|---|---|---|
| Year | 0.0227 | 0.3997 |
| Crop | <0.0001 | <0.0001 |
| Year*Crop | 0.4582 | 0.203 |
| Crop | CM | CS | OC | S2 | S3 |
|---|---|---|---|---|---|
| Shannon Index | 1.26 bc | 1.19 c | 1.23 c | 1.38 a | 1.35 ab |
| Species richness | 13.9 c | 17.7 c | 24.0 b | 31.6 a | 33.1 a |
| Year | 2011 | 2012 | 2013 | 2014 | |
| Species richness | 25.1 xy | 21.4 y | 26.9 x | 21.8 y |
| Crop | 2011 | 2012 | 2013 | 2014 |
|---|---|---|---|---|
| CM | 20.7 a | 24.3 c | 33.3 a | 14.5 a |
| CS | 16.8 a | 41.7 bc | 41.7 a | 20.4 a |
| OC | - | - | 46.7 a | 23.6 a |
| S2 | 42.5 a | 59.2 ab | 41.7 a | 11.4 a |
| S3 | 44.4 a | 71.7 a | 55.7 a | 27.5 a |
| Crop | Number of Species with Frequency >30% | Number of Species with Soil Cover >3% | Number of Species Fulfilling Both Conditions |
|---|---|---|---|
| CM | 17 | 13 | 3 |
| CS | 17 | 13 | 2 |
| OC | 35 | 12 | 8 |
| S2 | 30 | 11 | 6 |
| S3 | 26 | 19 | 6 |
| Species | Crops Where the Species is Within the Most Abundant or Most Frequent Ones |
|---|---|
| Lolium rigidum Gaud. | CM, CS, S2, S3 |
| Polygonum aviculare L. | CM, OC |
| Cerastium perfoliatum L. | CM |
| Polygonum convolvulus L. | CS, S2 |
| Chondrilla juncea L. | OC |
| Cirsium arvense L. | OC, S2 |
| Centaurea sp. | OC |
| Ranunculus arvensis L. | OC |
| Hypecoum procumbens L. | OC, S2 |
| Caucalis platycarpos L. | OC |
| Galium tricornutum Dandy | OC, S2 |
| Diplotaxis erucoides (L.) DC | S2 |
| Anacyclus clavatus (Desf.) Pers. | S3 |
| Bromus tectorum L. | S3 |
| Alyssum alyssoides (L.) L. | S3 |
| Sanguisorba minor Scop. | S3 |
| Descurainia sophia (L.) Webb ex Prantl | S3 |
| Name | Contribution % | p |
|---|---|---|
| S3 | 10.5 | <0.001 |
| OC | 9.3 | <0.001 |
| S2 | 9.2 | <0.001 |
| CM | 5.2 | 0.226 |
| CS | 5.2 | 0.226 |
| 2013 | 10.4 | <0.001 |
| 2012 | 8.6 | <0.001 |
| 2011 | 6.8 | <0.001 |
| 2014 | 6.8 | <0.001 |
| Slope | 7.7 | <0.001 |
| Altitude | 7.4 | <0.001 |
| Crop height | 6.8 | 0.007 |
| Crop soil cover (%) | 6.4 | 0.007 |
| Field perimeter/area | 6.1 | 0.027 |
| Weed soil cover (%) | 5.8 | 0.028 |
| Year/Crop | 2011 | 2012 | 2013 | 2014 | ||||
|---|---|---|---|---|---|---|---|---|
| Crop Cover (%) | Crop Height (cm) | Crop Cover (%) | Crop Height (cm) | Crop Cover (%) | Crop Height (cm) | Crop Cover (%) | Crop Height (cm) | |
| CM | 81.4 a | 53.6 a | 77.9 a | 41.4 a | 78.3 a | 50.8 a | 72.0 a | 56.0 a |
| CS | 80.8 a | 48.3 a | 76.7 a | 46.7 a | 78.0 a | 41.7 ab | 62.5 a | 58.3 a |
| OC | - | - | - | - | 51.7 a | 26.7 b | 66.4 a | 35.0 b |
| S2 | 74.4 a | 53.1 a | 73.3 a | 53.3 a | 81.7 a | 53.3 a | 80.7 a | 57.9 a |
| S3 | 68.3 a | 50.6 a | 46.7 b | 43.3 a | 58.7 a | 42.9 a | 69.2 a | 50.8 a |
| Species Richness | Total Seed Number (Mean) * | Total Seed Number (Mean) | Shannon Index * | Shannon Index | |
|---|---|---|---|---|---|
| CM | 8 a | 201 a | 202 a | 1.11 a | 1.11 a |
| S3 | 10 a | 226 a | 239 a | 1.18 a | 1.34 a |
| Rank of the Species | CM | S3 |
|---|---|---|
| Most abundant species | Polygonum spp.: 85 ± 26.5 | Polygonum spp.: 99 ± 16.0 |
| 2nd most abundant species | Amaranthus spp.: 62 ± 18.6 | Amaranthus spp.: 55 ± 25.0 |
| 3rd most abundant species | Heliotropium europaeum Pall: 11 ± 10.9 | Anthemis sp.: 18 ± 15.9 |
| 4th most abundant species | Nonidentified forb: 8 ± 5.3 | O. viciifolia: 13 ± 3.0 |
| Rank of the Species | CM | S3 |
|---|---|---|
| Most frequent species | Polygonum spp.: 100 | Polygonum spp.: 100 |
| 2nd most frequent species | Amaranthus spp.: 90 | Amaranthus spp., O. viciifolia, Veronica spp.: 100 |
| 3rd most frequent species | Veronica spp: 70 | Portulaca oleracea Lolium rigidum: 46 |
| 4th most frequent species | Lolium rigidum, Heliotropium europaeum, Fumaria spp.: 40 | Lamium amplexicaule: 39 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cirujeda, A.; Marí, A.I.; Murillo, S.; Aibar, J.; Pardo, G.; Solé-Senan, X.-O. May the Inclusion of a Legume Crop Change Weed Composition in Cereal Fields? Example of Sainfoin in Aragon (Spain). Agronomy 2019, 9, 134. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030134
Cirujeda A, Marí AI, Murillo S, Aibar J, Pardo G, Solé-Senan X-O. May the Inclusion of a Legume Crop Change Weed Composition in Cereal Fields? Example of Sainfoin in Aragon (Spain). Agronomy. 2019; 9(3):134. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030134
Chicago/Turabian StyleCirujeda, Alicia, Ana Isabel Marí, Sonia Murillo, Joaquín Aibar, Gabriel Pardo, and Xavier-Oriol Solé-Senan. 2019. "May the Inclusion of a Legume Crop Change Weed Composition in Cereal Fields? Example of Sainfoin in Aragon (Spain)" Agronomy 9, no. 3: 134. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030134