Exogenous Foliar Application of Glycine Betaine to Alleviate Water Deficit Tolerance in Two Indica Rice Genotypes under Greenhouse Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Exogenous Glybet Foliar and Water Deficit Treatments
2.2. Proline Quantification
2.3. Plant Physiological Assay
2.4. Plant Morphological Characterization and Yield Traits
2.5. Statistical Analysis
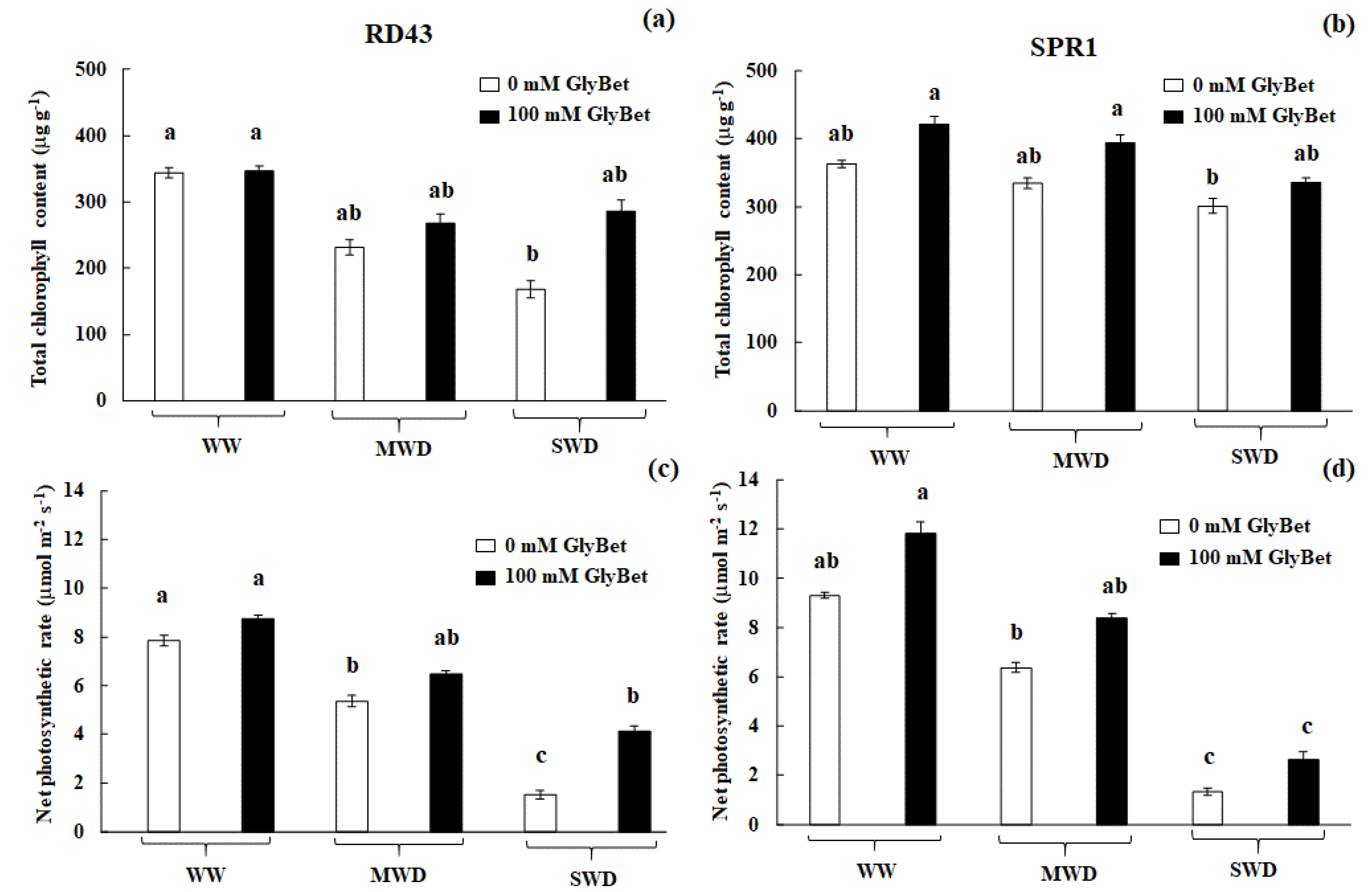
3. Results and Discussion
3.1. Free Proline Accumulation Pattern under Water Deficit Conditions
3.2. Overall Growth Performances and Yield Traits
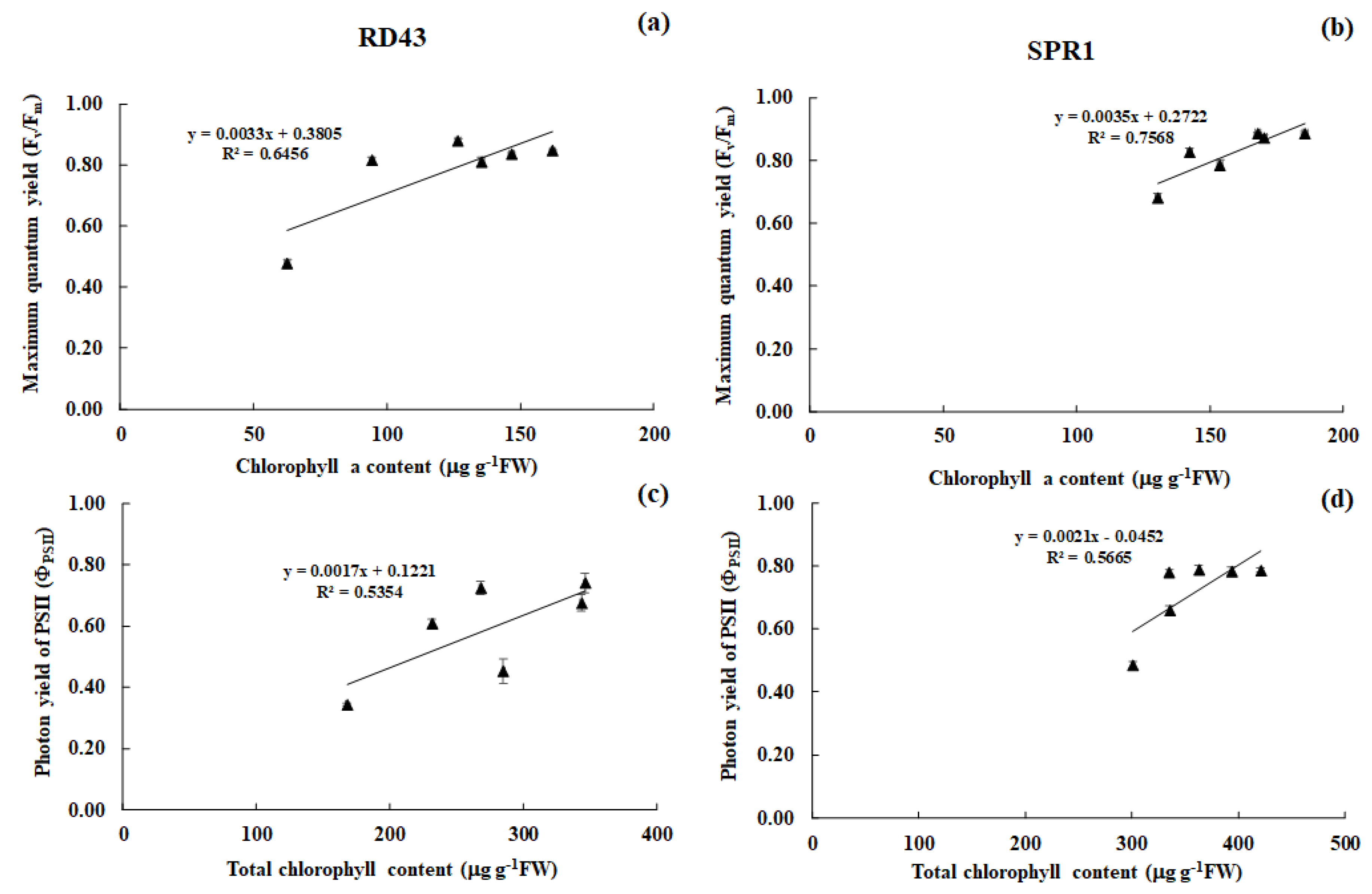
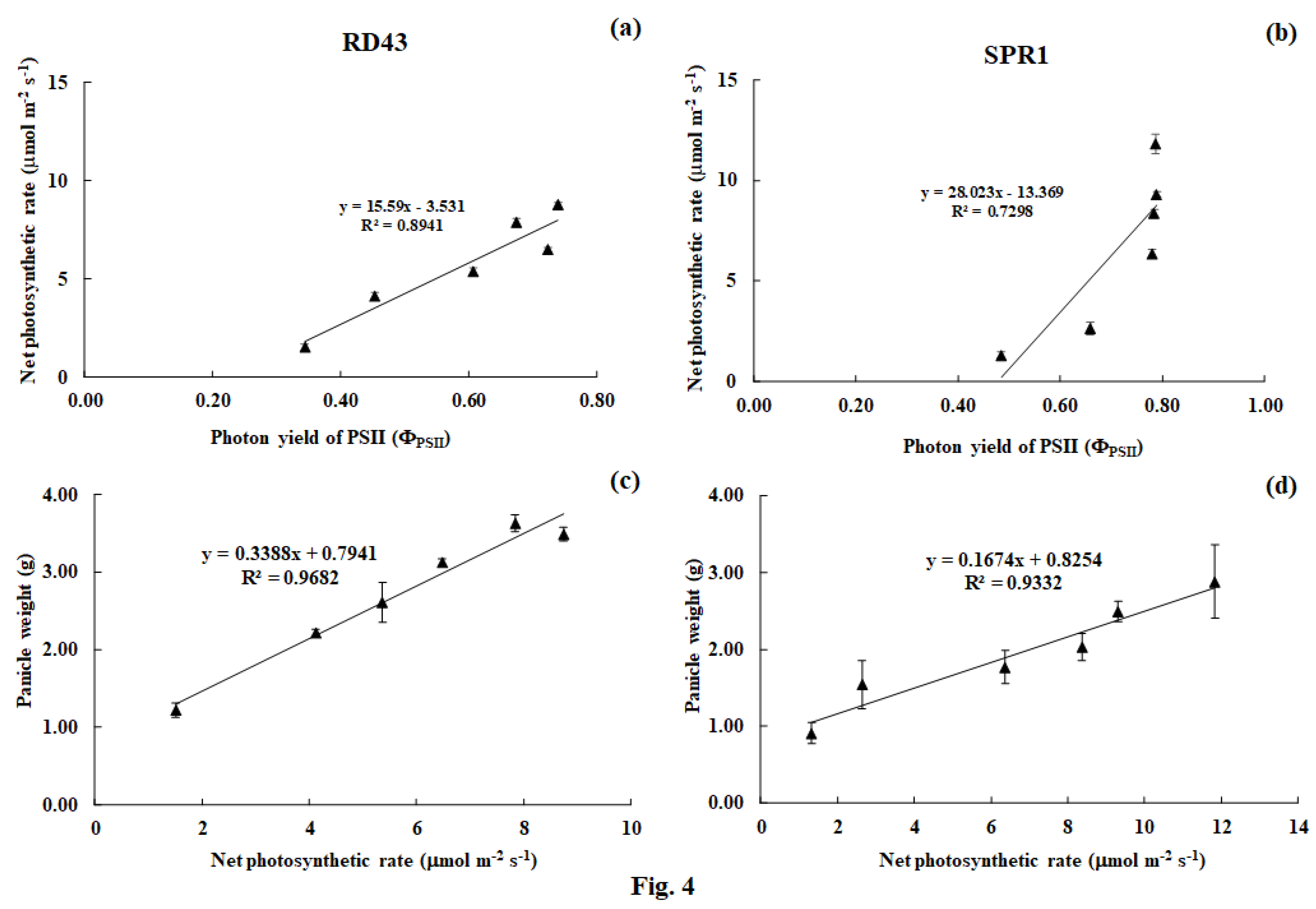
3.3. Physiological Changes and Relationships between Those Traits
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brar, D.S.; Singh, K.; Khush, G.S. Frontiers in Rice Breeding. In The Future Rice Strategy for India; Academic Press: Cambridge, MA, USA, 2017; pp. 137–160. [Google Scholar]
- Mottaleb, K.A.; Rejesus, R.M.; Murty, M.V.R.; Mohanty, S.; Li, T. Benefits of development and dissemination of climate-smart rice: Ex ante impact assessment of drought-tolerant rice in South Asia. Mitig. Adapt. Strat. Glob. Chang. 2017, 22, 879–901. [Google Scholar] [CrossRef]
- Bouman, B.A.M.; Humphreys, E.; Toung, T.P.; Barker, R. Rice and water. Adv. Agron. 2007, 92, 187–237. [Google Scholar]
- Smith, M.L.; Mohanty, R.K. Monsoon, rice production, and urban growth: The microscale management of ‘too much’ water. Holocene 2018, 28, 1325–1333. [Google Scholar] [CrossRef]
- Wu, S.J.; Chiueh, Y.W.; Hsu, C.T. Modeling risk analysis for rice production due to agro-climate change and uncertainty in irrigated water. Paddy Water Environ. 2018, 16, 35–53. [Google Scholar] [CrossRef]
- Maclean, J.L.; Dawe, D.; Hardy, B.; Hettle, G.P. Rice Almanac; International Rice Research Institute: Los Baños, Philippines, 2002; p. 253. [Google Scholar]
- Tuong, T.P.; Bouman, B.A.M.; Mortimer, M. More rice, less water–integrated approaches for increasing water productivity in irrigated rice-based system in Asia. Plant Prod. Sci. 2005, 8, 231–241. [Google Scholar] [CrossRef]
- Tuong, T.P.; Bouman, B.A.M. Rice production in water-scarce environments. In Water Productivity in Agriculture: Limits and Opportunities for Improvement; Kijne, J.W., Baker, R., Molden, D., Eds.; CAB International: Wallingford, UK, 2003; pp. 53–67. [Google Scholar]
- Kummu, M.; Guillaume, J.H.A.; de Moel, H.; Eisner, S.; Flörke, M.; Porkka, M.; Siebert, S.; Veldkamp, T.I.E.; Ward, P.J. The world’s road to water scarcity: Shortage and stress in the 20th century and pathways towards sustainability. Sci. Rep. 2016, 6, 38495. [Google Scholar] [CrossRef]
- Jiang, S.; Wang, J.; Zhao, Y.; Shang, Y.; Gao, X.; Li, H.; Wang, Q.; Zhu, Y. Sustainability of water resources for agriculture considering grain production, trade and consumption in China from 2004 to 2013. J. Clean. Prod. 2017, 149, 1210–1218. [Google Scholar] [CrossRef]
- van Oort, P.A.J.; Zwart, S.J. Impacts of climate change on rice production in Africa and causes of simulated yield changes. Glob. Chang. Biol. 2017, 24, 1029–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabnakorn, S.; Maskey, S.; Suryadi, F.X.; de Fraiture, C. Rice yield in response to climate trends and drought index in the Mun River Basin, Thailand. Sci. Total Environ. 2018, 621, 108–119. [Google Scholar] [CrossRef]
- Bandara, J.S.; Cai, Y. The impact of climate change on food crop productivity, food prices and food security in South Asia. Econ. Anal. Pol. 2014, 44, 451–465. [Google Scholar] [CrossRef]
- Li, T.; Angeles, O.; Radanielson, A.; Marcaida, M.; Manalo, E. Drought stress impacts of climate change on rainfed rice in South Asia. Clim. Chang. 2015, 133, 709–720. [Google Scholar] [CrossRef]
- Hayashi, K.; Llorca, L.; Rustini, S.; Setyanto, P.; Zaini, Z. Reducing vulnerability of rainfed agriculture through seasonal climate predictions: A case study on the rainfed rice production in Southeast Asia. Agric. Syst. 2018, 162, 66–76. [Google Scholar] [CrossRef]
- Shao, H.; Chen, X.-Y.; Chu, L.-Y.; Zhao, X.-N.; Wu, G.; Yuan, Y.-B.; Zhao, C.-X.; Hu, Z.-M. Investigation on the relationship of proline with wheat anti-drought under soil water deficits. Colloids Surf. B Biointerfaces 2006, 53, 113–119. [Google Scholar] [Green Version]
- Zhang, L.; Li, S.; Liang, Z. Differential plant growth and osmotic effects of two maize (Zea mays L.) cultivars to exogenous glycinebetaine application under drought stress. Plant Growth Regul. 2009, 58, 297–305. [Google Scholar]
- Ahanger, M.A.; Tyagi, S.R.; Wani, M.R.; Ahmad, P. Drought tolerance: Role of organic osmolytes, growth regulators, and mineral nutrients. In Physiological Mechanisms and Adaptation Strategies in Plants under Changing Environment; Ahmad, P., Wani, M.R., Eds.; Springer: New York, NY, USA, 2014; pp. 25–55. [Google Scholar]
- Ahmad, P.; Hameed, A.; Abd-Allah, E.F.; Sheikh, S.A.; Wani, M.R.; Rasool, S.; Jamsheed, S.; Kumar, A. Biochemical and molecular approaches for drought tolerance in plants. In Physiological Mechanisms and Adaptation Strategies in Plants under Changing Environment; Ahmad, P., Wani, M.R., Eds.; Springer: New York, NY, USA, 2014; pp. 1–29. [Google Scholar]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Kurepin, L.; Ivanov, A.G.; Zaman, M.; Pharis, R.P.; Allakhverdiev, S.I.; Hurry, V.; Hüner, N.P.A. Stress-related hormones and glycinebetaine interplay in protection of photosynthesis under abiotic stress conditions. Photosynth. Res. 2015, 126, 221–235. [Google Scholar] [CrossRef]
- Rathinasabapathi, R.; Gage, D.A.; Mackill, D.J.; Hanson, A.W. Cultivated and wild rice do not accumulate glycinebetaine due to deficiencies in two biosynthetic steps. Crop Sci. 1993, 33, 534–538. [Google Scholar] [CrossRef]
- Kishitani, S.; Takanami, T.; Suzuki, M.; Oikawa, M.; Yokoi, S.; Ishitani, M.; Alvarez-Nakase, A.M.; Takabe, T. Compatibility of glycinebetaine in rice plants: Evaluation using transgenic rice plants with a gene for peroxisomal betaine aldehyde dehydrogenase from barley. Plant Cell Environ. 2000, 23, 107–114. [Google Scholar] [CrossRef]
- Shirasawa, K.; Takabe, T.; Takabe, T.; Kishitani, S. Accumulation of glycinebetaine in rice plants that overexpress choline monooxygenase from spinach and evaluation of their tolerance to abiotic stress. Ann. Bot. 2006, 98, 565–571. [Google Scholar] [CrossRef]
- Soren, K.R.; Ali, K.; Tyagi, V.; Tyagi, A. Recent advances in molecular breeding of drought tolerance in rice (Oryza sativa L.). Indian J. Biotechnol. 2010, 9, 233–251. [Google Scholar]
- Farooq, M.; Basra, S.M.A.; Wahid, A.; Cheema, Z.A.; Cheema, M.A.; Khaliq, A. Physiological role of exogenously applied glycinebetaine to improve drought tolerance in fine grain aromatic rice (Oryza sativa L.). J. Agron. Crop Sci. 2008, 194, 325–333. [Google Scholar] [CrossRef]
- Cha-um, S.; Samphumphuang, T.; Kirdmanee, C. Glycinebetaine alleviates water deficit stress in indica rice using proline accumulation, photosynthetic efficiencies, growth performances and yield attributes. Aust. J. Crop Sci. 2013, 7, 213–218. [Google Scholar]
- Jalal-ud-Din; Khan, S.U.; Khan, A.; Naveed, S. Effect of exogenously applied kinetin and glycinebetaine on metabolic and yield attributes of rice (Oryza sativa L.) under drought stress. Emir. J. Food Agric. 2015, 27, 75–81. [Google Scholar] [CrossRef]
- Mäkelä, P.; Peltonen-Sainio, P.; Jokinen, K.; Pehu, E.; Setälä, H.; Hinkkanen, R.; Somersalo, S. Uptake and translocation of foliar-applied glycinebetaine in crop plants. Plant Sci. 1996, 121, 221–223. [Google Scholar] [CrossRef]
- Agboma, P.C.; Sinclair, T.R.; Jokinen, K.; Peltonen-Sainio, P.; Pehu, E. An evaluation of the effect of exogenous glycinebetaine on the growth and yield of soybean: Timing of application, watering regimes and cultivars. Field Crop Res. 1997, 54, 51–64. [Google Scholar] [CrossRef]
- Xing, W.; Rajashekar, C.B. Alleviation of water stress in beans by exogenous glycine betaine. Plant Sci. 1999, 148, 185–195. [Google Scholar] [CrossRef]
- Cruz, F.J.R.; Castro, G.L.S.; Silva Júnior, D.D.; Festucci-Buselli, R.A.; Pinheiro, H.A. Exogenous glycine betaine modulates ascorbate peroxidase and catalase activities and prevent lipid peroxidation in mild water-stressed Carapa guianensis plants. Photosynthetica 2013, 51, 102–108. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Ivanov, A.G.; Zaman, M.; Pharis, R.P.; Hurry, V.; Hüner, N.P.A. Interaction of glycine betaine and plant hormones: Protection of the photosynthetic apparatus during abiotic stress. In Photosynthesis, Structures, Mechanisms and Applications; Hou, H.J.M., Najafpour, M.M., Moore, G.F., Allakhverdiev, S.I., Eds.; Springer International Publishing: New York, NY, USA, 2017; pp. 185–202. [Google Scholar]
- Wasusun, A.; Wongpiyachon, S.; Songchitsomboon, S.; Sukviwat, W.; Maneenin, P.; Pakkethaiti, S. Rice variety RD43: Medium glycemic index rice for niche market. Thai Rice Res. J. 2017, 8, 45–53. [Google Scholar]
- Suwannaporn, P.; Pitiphunpong, S.; Champangern, S. Classification of rice amylose content by discriminant analysis of physicochemical properties. Starch 2007, 59, 171–177. [Google Scholar] [CrossRef]
- Pengkumsri, N.; Chaiyasut, C.; Saenjum, C.; Sirilun, S.; Peerajan, S.; Suwannalert, P.; Sirisattha, S.; Sivamaruthi, B.S. Physicochemical and antioxidative properties of black, brown and red rice varieties of northern Thailand. Food Sci. Technol. 2015, 35, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Pongprayoon, W.; Tisarum, R.; Theerawitaya, C.; Cha-um, S. Evaluation and clustering on salt-tolerant ability in rice genotypes (Oryza sativa L. subsp. indica) using multivariate physiological indices. Physiol. Mol. Biol. Plant 2019. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Shabala, S.N.; Shabala, S.I.; Martynenko, A.I.; Babourina, O.; Newman, I.A. Salinity effect on bioelectric activity growth, Na+ accumulation and chlorophyll fluorescence of maize leaves: A comparative survey and prospects for screening. Aust. J. Plant Physiol. 1998, 25, 609–616. [Google Scholar] [CrossRef]
- Loggini, B.; Scartazza, A.; Brugnoli, E.; Navari-Izzo, F. Antioxidant defense system, pigment composition, and photosynthetic efficiency in two wheat cultivars subjected to drought. Plant Physiol. 1999, 119, 1091–1100. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Cha-um, S.; Supaibulwatana, K.; Kirdmanee, C. Glycinebetaine accumulation, physiological characterizations and growth efficiency in salt-tolerant and salt-sensitive lines of indica rice (Oryza sativa L. ssp. indica) in response to salt stress. J. Agron. Crop Sci. 2007, 193, 157–166. [Google Scholar]
- Harinasut, P.; Tsutsui, K.; Takabe, T.; Nomura, M.; Takabe, T.; Kishitani, S. Exogenous glycinebetaine accumulation and increased salt tolerance in rice seedlings. Biosci. Biotechnol. Biochem. 1996, 60, 366–368. [Google Scholar] [CrossRef]
- Demiral, T.; Türkan, I. Exogenous glycinebetaine affects growth and proline accumulation and retards senescence in two rice cultivars under NaCl stress. Environ. Exp. Bot. 2006, 56, 72–79. [Google Scholar] [CrossRef]
- Cha-um, S.; Kirdmanee, C. Effect of glycinebetaine on proline, water use and photosynthetic efficiencies, and growth of rice seedlings under salt stress. Turk. J. Agric. For. 2010, 34, 517–527. [Google Scholar]
- El-Hendawy, S.E.; Kotab, M.A.; Al-Suhaibani, N.A.; Schmidhalter, U. Optimal coupling combinations between the irrigation rate and glycinebetaine levels for improving yield and water use efficiency of drip-irrigation maize grown under arid conditions. Agric. Water Manag. 2014, 140, 69–78. [Google Scholar] [CrossRef]
- Zhao, X.X.; Ma, Q.Q.; Liang, C.; Fang, Y.; Wang, Y.Q.; Wang, W. Effect of glycinebetaine on function of thylakoid membranes in wheat flag leaves under drought stress. Biol. Plant. 2007, 51, 584–588. [Google Scholar] [CrossRef]
- Ma, Q.Q.; Wang, W.; Li, Y.H.; Li, D.Q.; Zou, Q. Alleviation of photoinhibition in drought-stressed wheat (Triticumaestivum) by foliar-applied glycinebetaine. J. Plant Physiol. 2006, 163, 165–175. [Google Scholar] [CrossRef]
- Gupta, N.; Thind, S.K.; Bains, N.S. Glycine betaine application modifies biochemical attributes of osmotic adjustment in drought stressed wheat. Plant Growth Regul. 2014, 72, 221–228. [Google Scholar] [CrossRef]
- Ma, X.L.; Wang, Y.J.; Xie, S.L.; Wang, C.; Wang, W. Glycinebetaine application ameliorates negative effects of drought stress in tobacco. Russ. J. Plant Physiol. 2007, 54, 472–479. [Google Scholar] [CrossRef]
- Iqbal, N.; Ashraf, Y.; Ashraf, M. Modulation of endogenous levels of some key organic metabolites by exogenous application of glycine betaine in drought stressed plants of sunflower (Helianthus annuus L.). Plant Growth Regul. 2011, 63, 7–12. [Google Scholar] [CrossRef]
- Mahouachi, J.; Argamasilla, R.; Gómez-Cadenas, A. Influence of exogenous glycine betaine and abscisic acid on papaya in responses to water-deficit stress. J. Plant Growth Regul. 2012, 31, 1–10. [Google Scholar] [CrossRef]
- Hussain, M.; Malik, M.A.; Farooq, M.; Ashraf, M.Y.; Cheema, M.A. Improving drought tolerance by exogenous application of glycinebetaine and salicylic acid in sunflower. J. Agron. Crop Sci. 2008, 194, 193–199. [Google Scholar] [CrossRef]
- Osman, H.S. Enhancing antioxidant-yield relationship of pea plant under drought at different growth stages by exogenously applied glycine betaine and proline. Ann. Agric. Sci. 2015, 60, 389–402. [Google Scholar] [CrossRef]
- Anjum, S.A.; Farooq, M.; Wang, L.C.; Xue, L.L.; Wang, S.G.; Wang, L.; Zhang, S.; Chen, M. Gas exchange and chlorophyll synthesis of maize cultivars are enhanced by exogenously-applied glycinebetaine under drought conditions. Plant Soil Environ. 2011, 57, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Rezaei, M.A.; Jokar, I.; Ghorbanli, M.; Kaviani, B.; Kharabian-Masouleh, A. Morpho-physiological improving effects of exogenous glycine betaine on tomato (Lycopersicum esculentum Mill.) cv. PS under drought stress conditions. Plant Omic 2012, 5, 79–86. [Google Scholar]
- Gupta, N.; Thind, S.K. Improving photosynthetic performance of bread wheat under field drought stress by foliar applied glycine betaine. J. Agric. Sci. Technol. 2015, 17, 75–86. [Google Scholar]
- Cha-um, S.; Supaibulwatana, K.; Kirdmanee, C. Water relation, photosynthetic ability and growth of Thai jasmine rice (Oryza sativa L. ssp. indica cv. KDML105) to salt stress by application of exogenous glycinebetaine and choline. J. Agron. Crop Sci. 2006, 192, 25–36. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Regimes | GlyBet | RD43 | SPR1 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SH (cm) | NL | NT | FLA (mm2) | SH (cm) | NL | NT | FLA (mm2) | ||
| WW | 0 | 95.47a | 3.67a | 11.00a | 8158ab | 107.67a | 3.33a | 7.67a | 8647a |
| 100 | 96.28a | 3.50a | 10.00ab | 8865a | 105.05a | 3.17a | 7.67a | 10176a | |
| MWD | 0 | 92.10b | 3.33a | 9.33b | 7183bc | 105.70a | 3.20a | 6.00b | 5314b |
| 100 | 93.17ab | 3.50a | 9.50b | 9393a | 107.88a | 3.27a | 6.33ab | 8695a | |
| SWD | 0 | 90.48b | 2.00b | 9.00b | 3857d | 92.13c | 1.60b | 5.80b | 3848b |
| 100 | 92.13b | 2.67b | 9.33b | 6287c | 106.84a | 1.83b | 6.00b | 5962b | |
| Significant level | |||||||||
| Water | * | ** | ** | ** | * | ns | ns | ** | |
| GlyBet | ns | ns | ns | ** | ** | ** | ** | ** | |
| Water×GlyBet | ns | ns | ns | ns | * | * | * | ** | |
| Water Regimes | GlyBet | RD43 | SPR1 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| PW (g) | PL (cm) | SF (%) | HGW(g) | PW (g) | PL (cm) | SF (%) | HGW (g) | ||
| WW | 0 | 3.63a | 27.91a | 66.18ab | 2.61a | 2.49a | 27.08ab | 70.34a | 2.66a |
| 100 | 3.49ab | 27.85a | 72.08a | 2.58a | 2.88a | 28.03a | 70.27a | 2.69a | |
| MWD | 0 | 2.61c | 25.32b | 65.58ab | 2.56a | 1.77bc | 24.80cd | 48.98b | 2.58a |
| 100 | 3.13b | 26.95ab | 69.25ab | 2.48a | 2.03b | 25.78bc | 55.83a | 2.55a | |
| SWD | 0 | 1.22e | 20.82c | 26.84d | 2.23b | 0.91d | 22.01e | 17.61c | 2.13b |
| 100 | 2.22d | 25.50b | 48.19c | 2.54a | 1.54c | 23.25de | 31.86b | 2.58a | |
| Significant level | |||||||||
| Water | ** | ** | ** | ** | ** | ** | ** | ** | |
| GlyBet | ** | ** | ns | ** | ** | * | ** | ** | |
| Water×GlyBet | ** | ** | ** | ** | ns | ns | ** | ** | |
| Water Regimes | GlyBet | RD43 | SPR1 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Chla | Chlb | Fv/Fm | ΦPSII | Chla | Chlb | Fv/Fm | ΦPSII | ||
| WW | 0 | 146.76a | 197.09a | 0.836a | 0.675b | 167.92ab | 195.04ab | 0.877a | 0.789a |
| 100 | 162.15a | 184.27a | 0.845a | 0.744a | 185.44a | 235.51a | 0.888a | 0.787a | |
| MWD | 0 | 94.41b | 137.34ab | 0.813ab | 0.607b | 142.35b | 192.57ab | 0.827a | 0.780a |
| 100 | 126.62ab | 141.35ab | 0.879a | 0.724a | 170.18ab | 223.35a | 0.847a | 0.784a | |
| SWD | 0 | 62.59c | 105.44b | 0.477b | 0.344d | 130.38b | 170.63b | 0.681c | 0.458b |
| 100 | 135.45ab | 149.70ab | 0.809ab | 0.453c | 153.79ab | 181.65ab | 0.786b | 0.786a | |
| Significant level | |||||||||
| Water | ** | ** | ** | ** | ** | ** | ** | ** | |
| GlyBet | ns | ns | ns | ** | ns | ns | * | ns | |
| Water×GlyBet | * | * | * | ** | * | * | * | * | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tisarum, R.; Theerawitaya, C.; Samphumphung, T.; Takabe, T.; Cha-um, S. Exogenous Foliar Application of Glycine Betaine to Alleviate Water Deficit Tolerance in Two Indica Rice Genotypes under Greenhouse Conditions. Agronomy 2019, 9, 138. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030138
Tisarum R, Theerawitaya C, Samphumphung T, Takabe T, Cha-um S. Exogenous Foliar Application of Glycine Betaine to Alleviate Water Deficit Tolerance in Two Indica Rice Genotypes under Greenhouse Conditions. Agronomy. 2019; 9(3):138. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030138
Chicago/Turabian StyleTisarum, Rujira, Cattarin Theerawitaya, Thapanee Samphumphung, Teruhiro Takabe, and Suriyan Cha-um. 2019. "Exogenous Foliar Application of Glycine Betaine to Alleviate Water Deficit Tolerance in Two Indica Rice Genotypes under Greenhouse Conditions" Agronomy 9, no. 3: 138. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030138