Physiological and Growth Response of Pepper (Capsicum annum L.) Seedlings to Supplementary Red/Blue Light Revealed through Transcriptomic Analysis


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Treatments and Experimental Design
2.3. Measurement of Plant Growth and Physiological Parameters
2.4. RNA Extraction and RNA-Seq
2.5. Analysis of Differentially Expressed Genes (DEGs)
2.6. Validation of DEGs by Real-Time Quantitative PCR
2.7. Statistical Analysis
3. Results
3.1. Effect of Light Durations on Growth Indexes
3.2. Effect of Light Durations on Plant Physiological Indexes
3.3. Effects of Light Durations on the Transcriptome of Pepper Plantlets
3.3.1. Statistics and Sequencing Quality Assessment of the RNA-Seq Data
3.3.2. Results of the Screen for the DEGs between Treatments
3.3.3. GO Analysis of DEGs in Plants That Received Light Duration Treatments
3.3.4. KEGG Pathway Analysis of DEGs in the Light Duration Treatments
3.3.5. Venn Diagram Analysis and Cluster Analysis of DEGs among the Treatments
3.3.6. Confirmation of the RNA-Seq Results Using qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fukuda, N.; Ajima, C.; Yukawa, T.; Olsen, J.E. Antagonistic action of blue and red light on shoot elongation in petunia depends on gibberellin, but the effects on flowering are not generally linked to gibberellin. Environ. Exp. Bot. 2016, 121, 102–111. [Google Scholar] [CrossRef]
- Islam, M.A.; Kuwar, G.; Clarke, J.L.; Blystad, D.-R.; Gislerød, H.R.; Olsen, J.E.; Torre, S. Artificial light from light emitting diodes (LEDs) with a high portion of blue light results in shorter poinsettias compared to high pressure sodium (HPS) lamps. Sci. Hortic. 2012, 147, 136–143. [Google Scholar] [CrossRef]
- Nanya, K.; Ishigami, Y.; Hikosaka, S.; Goto, E. Effects of blue and red light on stem elongation and flowering of tomato seedlings. In Proceedings of the VII International Symposium on Light in Horticultural Systems, Wageningen, The Netherlands, 15–18 October 2012; pp. 261–266. [Google Scholar]
- Terfa, M.T.; Solhaug, K.A.; Gislerød, H.R.; Olsen, J.E.; Torre, S. A high proportion of blue light increases the photosynthesis capacity and leaf formation rate of Rosa× hybrida but does not affect time to flower opening. Physiol. Plant. 2013, 148, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-L.; Guo, W.-Z.; Xue, X.-Z.; Wang, L.-C.; Qiao, X.-J. Growth and quality responses of ‘Green Oak Leaf’ lettuce as affected by monochromic or mixed radiation provided by fluorescent lamp (FL) and light-emitting diode (LED). Sci. Hortic. 2014, 172, 168–175. [Google Scholar] [CrossRef]
- Brown, C.S.; Schuerger, A.C.; Sager, J.C. Growth and photomorphogenesis of pepper plants under red light-emitting diodes with supplemental blue or far-red lighting. J. Am. Soc. Hortic. Sci. 1995, 120, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Snowden, M.C. Effects of Blue and Green Light on Plant Growth and Development at Low and High Photosynthetic Photon Flux; Utah State University: Logan, UT, USA, 2015. [Google Scholar]
- Košvancová-Zitová, M.; Urban, O.; Navrátil, M.; Špunda, V.; Robson, T.; Marek, M. Blue radiation stimulates photosynthetic induction in Fagus sylvatica L. Photosynthetica 2009, 47, 388. [Google Scholar] [CrossRef]
- Son, K.-H.; Oh, M.-M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Li, X.; Lu, W.; Hu, G.; Wang, X.C.; Zhang, Y.; Sun, G.X.; Fang, Z. Effects of light-emitting diode supplementary lighting on the winter growth of greenhouse plants in the Yangtze River Delta of China. Bot. Stud. 2016, 57, 2. [Google Scholar] [CrossRef] [PubMed]
- Bolger, M.E.; Arsova, B.; Usadel, B. Plant genome and transcriptome annotations: From misconceptions to simple solutions. Brief. Bioinform. 2017, 19, 437–449. [Google Scholar] [CrossRef]
- Li, C.-X.; Xu, Z.-G.; Dong, R.-Q.; Chang, S.-X.; Wang, L.-Z.; Khalil-Ur-Rehman, M.; Tao, J.-M. An RNA-seq analysis of grape plantlets grown in vitro reveals different responses to blue, green, red LED light, and white fluorescent light. Front. Plant Sci. 2017, 8, 78. [Google Scholar] [CrossRef]
- Deng, Y.; Yao, J.; Wang, X.; Guo, H.; Duan, D. Transcriptome sequencing and comparative analysis of Saccharina japonica (Laminariales, Phaeophyceae) under blue light induction. PLoS ONE 2012, 7, e39704. [Google Scholar] [CrossRef] [PubMed]
- Gunnlaugsson, B.; Adalsteinsson, S. Interlight and plant density in year-round production of tomato at northern latitudes. In Proceedings of the V International Symposium on Artificial Lighting in Horticulture 711, Lillehammer, Norway, 30 June 2006; pp. 71–76. [Google Scholar]
- Hovi-Pekkanen, T.; Tahvonen, R. Effects of interlighting on yield and external fruit quality in year-round cultivated cucumber. Sci. Hortic. 2008, 116, 152–161. [Google Scholar] [CrossRef]
- Pettersen, R.I.; Torre, S.; Gislerød, H.R. Effects of intracanopy lighting on photosynthetic characteristics in cucumber. Sci. Hortic. 2010, 125, 77–81. [Google Scholar] [CrossRef]
- Fierro, A.; Gosselin, A.; Tremblay, N. Supplemental carbon dioxide and light improved tomato and pepper seedling growth and yield. HortScience 1994, 29, 152–154. [Google Scholar] [CrossRef]
- Li, X.; Kang, S.; Li, F.; Zhang, X.; Huo, Z.; Ding, R.; Tong, L.; Du, T.; Li, S. Light supplement and carbon dioxide enrichment affect yield and quality of off-season pepper. Agron. J. 2017, 109, 2107–2118. [Google Scholar] [CrossRef]
- Naing, A.H.; Jeon, S.M.; Park, J.S.; Kim, C.K. Combined effects of supplementary light and CO2 on rose growth and the production of good quality cut flowers. Can. J. Plant Sci. 2016, 96, 503–510. [Google Scholar] [CrossRef]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef]
- Olle, M.; Viršile, A. The effects of light-emitting diode lighting on greenhouse plant growth and quality. Agric. Food Sci. 2013, 22, 223–234. [Google Scholar] [CrossRef]
- Taïbi, K.; Taïbi, F.; Abderrahim, L.A.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Elsevier: New York, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Chen, J.; Hou, K.; Qin, P.; Liu, H.; Yi, B.; Yang, W.; Wu, W. RNA-Seq for gene identification and transcript profiling of three Stevia rebaudiana genotypes. BMC Genom. 2014, 15, 571. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2009, 26, 136–138. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 289–300. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621. [Google Scholar] [CrossRef] [PubMed]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef]
- Evans, J.R. Acclimation by the thylakoid membranes to growth irradiance and the partitioning of nitrogen between soluble and thylakoid proteins. Funct. Plant Biol. 1988, 15, 93–106. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.G. Impact of LED irradiance on plant photosynthesis and action spectrum of plantlet. In Proceedings of the Optics and Photonics for Information Processing VIII, San Diego, CA, USA, 19–20 May 2014; p. 921602. [Google Scholar]
- Wellmer, F.; Kircher, S.; Rügner, A.; Frohnmeyer, H.; Schäfer, E.; Harter, K. Phosphorylation of the parsley bZIP transcription factor CPRF2 is regulated by light. J. Biol. Chem. 1999, 274, 29476–29482. [Google Scholar] [CrossRef]
- Huq, E.; Tepperman, J.M.; Quail, P.H. GIGANTEA is a nuclear protein involved in phytochrome signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2000, 97, 9789–9794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harari-Steinberg, O.; Ohad, I.; Chamovitz, D.A. Dissection of the light signal transduction pathways regulating the two early light-induced protein genes in Arabidopsis. Plant Physiol. 2001, 127, 986–997. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Nakazawa, M.; Takase, T.; Manabe, K.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Matsui, M. Overexpression of LSH1, a member of an uncharacterised gene family, causes enhanced light regulation of seedling development. Plant J. 2004, 37, 694–706. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Li, Y.; Liu, T.; Duan, W.; Huang, Z.; Wang, L.; Tan, H.; Hou, X. Genes associated with agronomic traits in non-heading Chinese cabbage identified by expression profiling. BMC Plant Biol. 2014, 14, 71. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, M.; Spetea, C.; Hundal, T.; Oppenheim, A.B.; Adam, Z.; Andersson, B. The thylakoid FtsH protease plays a role in the light-induced turnover of the photosystem II D1 protein. Plant Cell 2000, 12, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Berry-Lowe, S.L.; Meagher, R.B. Transcriptional regulation of a gene encoding the small subunit of ribulose-1, 5-bisphosphate carboxylase in soybean tissue is linked to the phytochrome response. Mol. Cell. Biol. 1985, 5, 1910–1917. [Google Scholar] [CrossRef] [PubMed]
- Montané, M.-H.; Kloppstech, K. The family of light-harvesting-related proteins (LHCs, ELIPs, HLIPs): Was the harvesting of light their primary function? Gene 2000, 258, 1–8. [Google Scholar] [CrossRef]






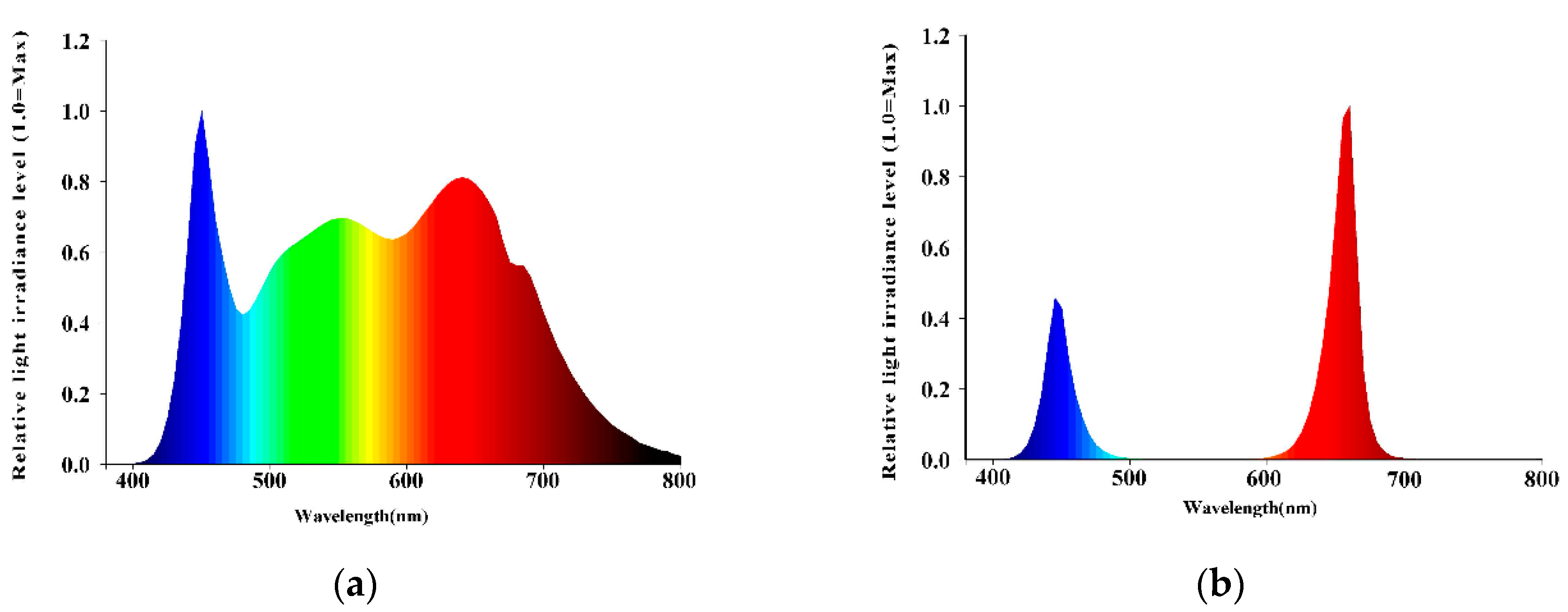{kind=link}
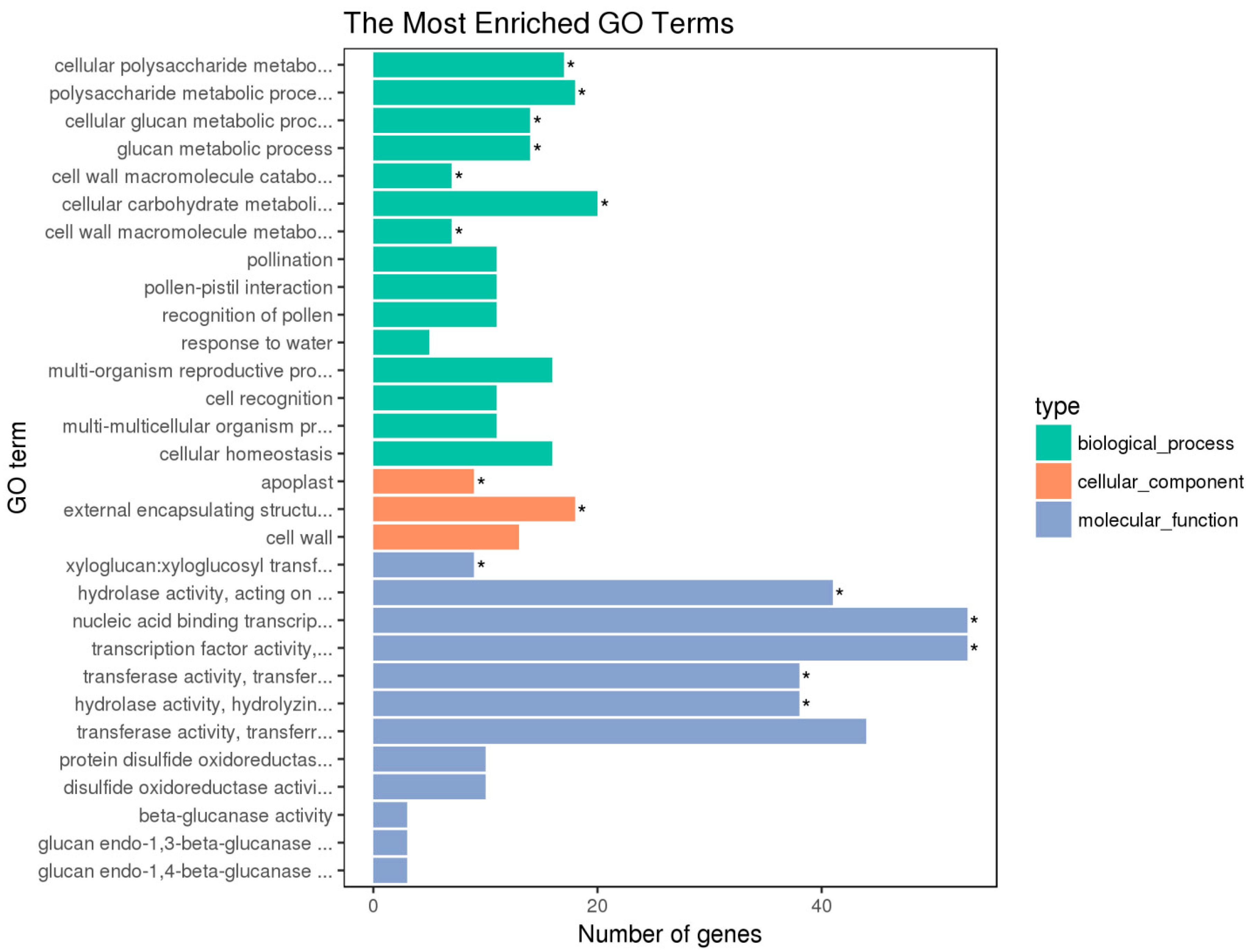{kind=link}
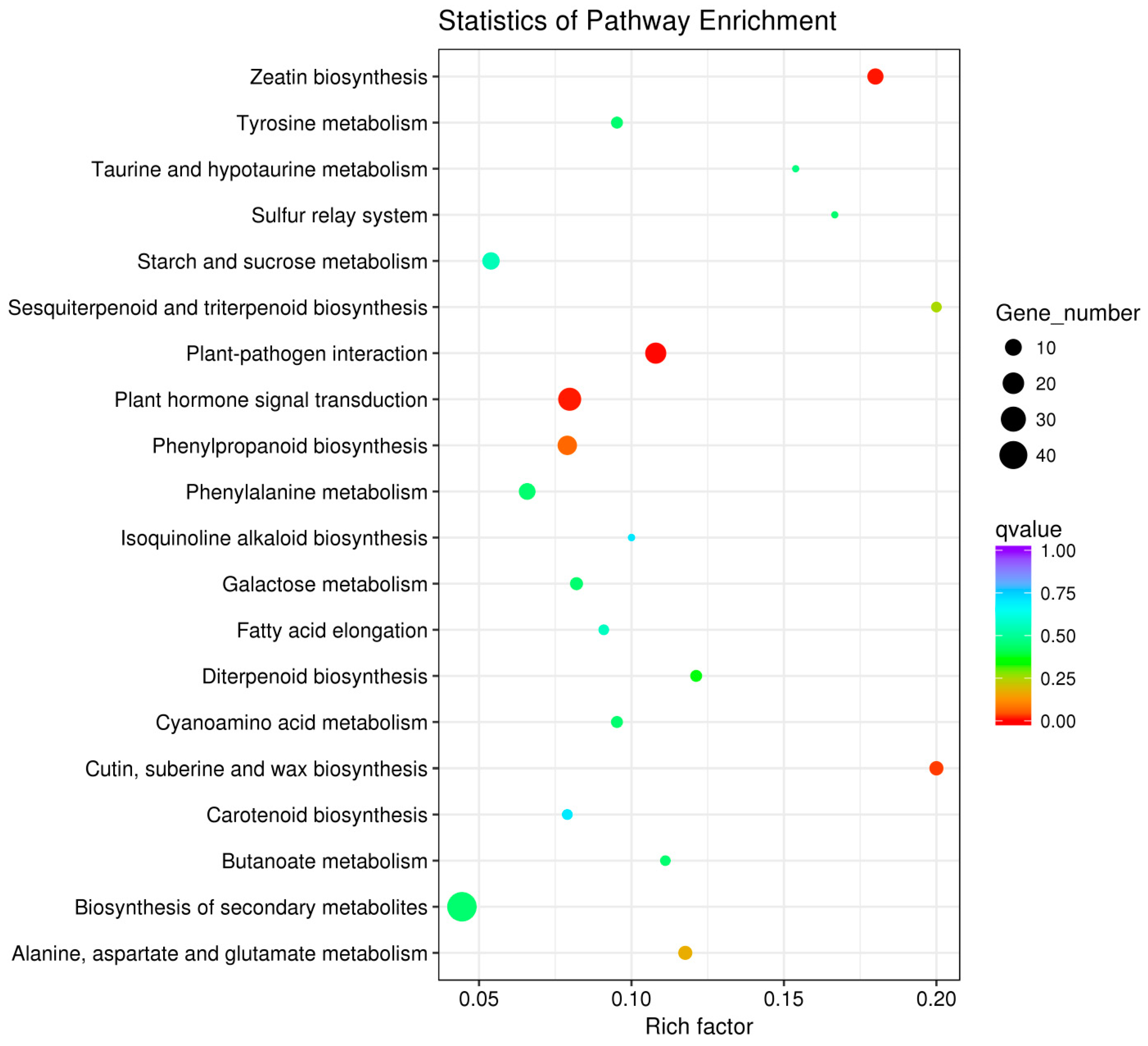{kind=link}
{kind=link}
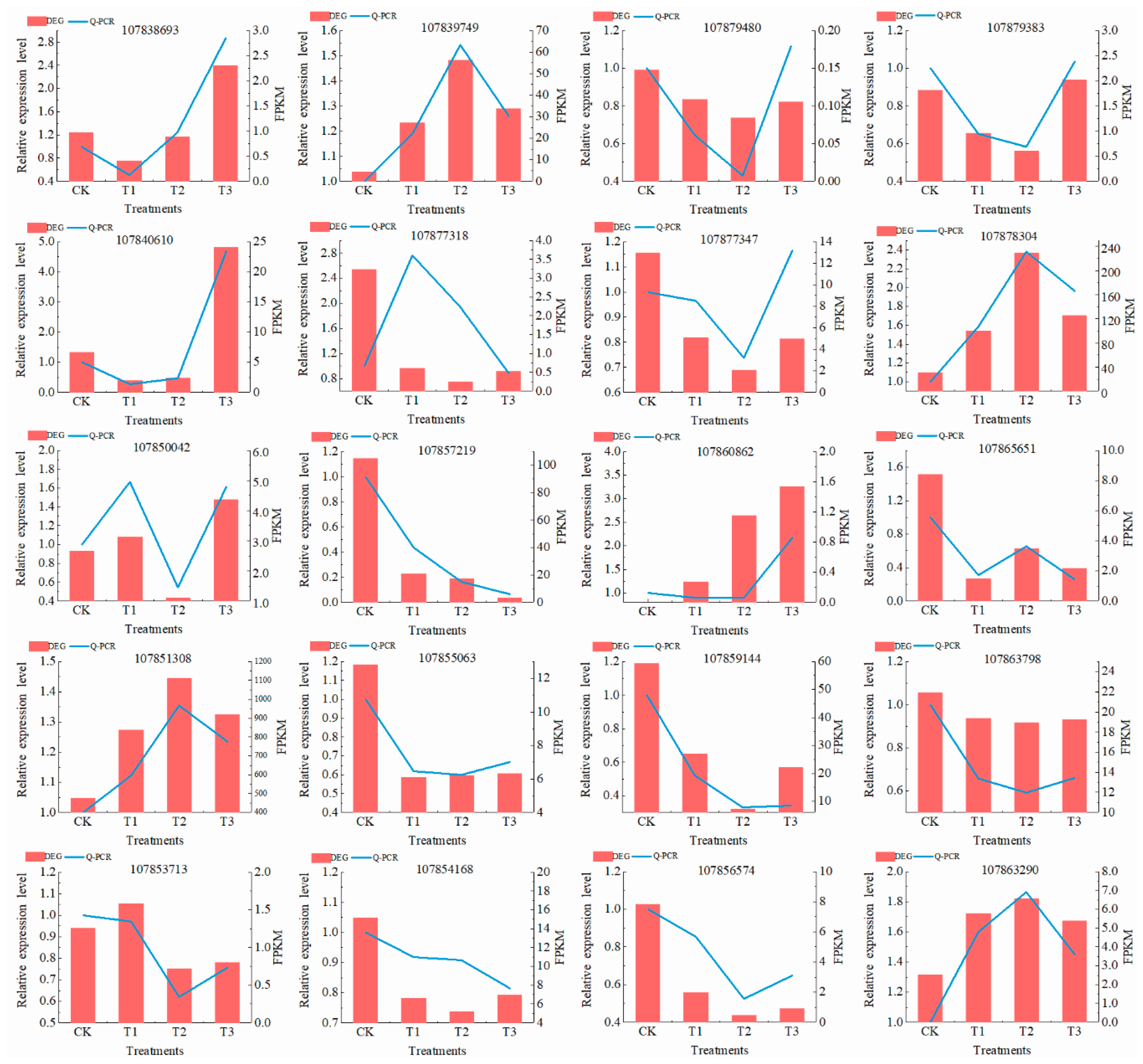{kind=link}
| Treatments | Description |
|---|---|
| CK (Control) | 7 h white LED + no supplementary R4B1 |
| T1 | 7 h white LED + 1 h supplementary R4B1 |
| T2 | 7 h white LED + 3 h supplementary R4B1 |
| T3 | 7 h white LED + 5 h supplementary R4B1 |
| Light Treatment | Plant Height (cm) | Stem Diameter (mm) | Leaf Area (cm2) | Total Length of Root (cm) | Root Surface Area (cm2) | Root Volume (cm3) | Fresh Biomass (g·plant−1) | Dry Weight of Root (g·plant−1) | Dry Weight of Stem (g·plant−1) | Dry Weight of Leaf (g·plant−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| CK | 10.33 ± 0.19 c | 3.45 ± 0.11 a | 33.48 ± 1.52 b | 444.51 ± 32.49 c | 67.81 ± 4.69 b | 0.83 ± 0.10 b | 9.62 ± 0.30 b | 0.14 ± 0.02 b | 0.10 ± 0.01 b | 0.49 ± 0.08 b |
| T1 | 14.97 ± 0.29b | 3.54 ± 0.08 a | 39.81 ± 2.56 a | 502.40 ± 32.28 c | 80.37 ± 7.04 b | 1.00 ± 0.06 b | 11.12 ± 0.15 a | 0.20 ± 0.01 ab | 0.17 ± 0.01 a | 0.71 ± 0.01 a |
| T2 | 17.37 ± 0.91 a | 3.65 ± 0.10 a | 43.79 ± 0.79 a | 963.94 ± 21.90 a | 155.76 ± 1.81 a | 2.01 ± 0.06 a | 12.48 ± 0.61 a | 0.21 ± 0.01 ab | 0.22 ± 0.03 a | 0.79 ± 0.02 a |
| T3 | 16.73 ± 0.96 ab | 3.78 ± 0.10 a | 33.67 ± 0.96 b | 803.07 ± 27.87 b | 137.23 ± 13.67 a | 1.87 ± 0.37 a | 12.03 ± 0.51 a | 0.27 ± 0.01 a | 0.19 ± 0.02 a | 0.74 ± 0.03 a |
| Light Treatment | Net Photosynthetic Rate (Pn/μmol·m–2·s–1) | Stomatal Conductance (Gs/mmol·m−2·s−1) | Intercellular CO2 Concentration (Ci/μmol·mol−1) | Transpiration Rate (Tr/mmol·m−2·s−1) | Chlorophyll a Content (mg·gFW−1) | Chlorophyll b Content (mg·gFW−1) | Carotenoid Content (mg·gFW−1) | Total Chlorophyll Content (mg·gFW−1) |
|---|---|---|---|---|---|---|---|---|
| CK | 8.5 ± 0.31 a | 402 ± 8.50 b | 396 ± 3.10 a | 3.07 ± 0.11 b | 1.79 ± 0.03 b | 0.69 ± 0.02 a | 0.24 ± 0.01 a | 2.48 ± 0.05 c |
| T1 | 9.1 ± 0.22 a | 473 ± 8.07 a | 389 ± 0.45 a | 3.4 ± 0.19 ab | 1.89 ± 0.05 b | 0.74 ± 0.05 a | 0.31 ± 0.04 a | 2.63 ± 0.04 c |
| T2 | 9.8 ± 0.47 a | 488 ± 9.62 a | 365 ± 8.06 b | 3.8 ± 0.13 a | 2.38 ± 0.09 a | 0.81 ± 0.09 a | 0.31 ± 0.04 a | 3.19 ± 0.08 b |
| T3 | 6.6 ± 0.21 b | 332 ± 19.74 c | 393 ± 2.93 a | 3.2 ± 0.18 ab | 2.49 ± 0.09 a | 0.90 ± 0.09 a | 0.33 ± 0.04 a | 3.39 ± 0.03 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Z.; Yu, J.; Xie, J.; Lyu, J.; Feng, Z.; Dawuda, M.M.; Liao, W.; Wu, Y.; Hu, L. Physiological and Growth Response of Pepper (Capsicum annum L.) Seedlings to Supplementary Red/Blue Light Revealed through Transcriptomic Analysis. Agronomy 2019, 9, 139. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030139
Tang Z, Yu J, Xie J, Lyu J, Feng Z, Dawuda MM, Liao W, Wu Y, Hu L. Physiological and Growth Response of Pepper (Capsicum annum L.) Seedlings to Supplementary Red/Blue Light Revealed through Transcriptomic Analysis. Agronomy. 2019; 9(3):139. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030139
Chicago/Turabian StyleTang, Zhongqi, Jihua Yu, Jianming Xie, Jian Lyu, Zhi Feng, Mohammed Mujitaba Dawuda, Weibiao Liao, Yue Wu, and Linli Hu. 2019. "Physiological and Growth Response of Pepper (Capsicum annum L.) Seedlings to Supplementary Red/Blue Light Revealed through Transcriptomic Analysis" Agronomy 9, no. 3: 139. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030139