Effects of Nanoparticles on Plant Growth-Promoting Bacteria in Indian Agricultural Soil


1
Caius Research Laboratory, St. Xavier’s College, Mumbai 400 001, India
2
Nanotechnology Research Group, Central Institute for Research on Cotton Technology, Mumbai 400 019, India
*
Author to whom correspondence should be addressed.
Agronomy 2019, 9(3), 140; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030140
Submission received: 2 February 2019
/
Revised: 11 March 2019
/
Accepted: 13 March 2019
/
Published: 18 March 2019
Abstract
:Soil bacteria are some of the key players affecting plant productivity. Soil today is exposed to emerging contaminants like metal engineered nanoparticles. The objective of this study was to evaluate the toxicological effects of silver and zinc oxide nanoparticles on bacteria classified as plant growth-promoting bacteria. Three types of bacteria—nitrogen fixers, phosphate solubilizers, and biofilm formers—were exposed to engineered nanoparticles. Initially, the effect of silver and zinc oxide nanoparticles was determined on pure cultures of the bacteria. These nanoparticles were then applied to soil to assess changes in composition of bacterial communities. Impacts of the nanoparticles were analyzed using Illumina MiSeq sequencing of 16S rRNA genes. In the soil used, relative abundances of the dominant and agriculturally significant phyla, namely, Proteobacteria, Actinobacteria, and Firmicutes, were altered in the presence of silver nanoparticles. Silver nanoparticles changed the abundance of the three phyla by 25 to 45%. Zinc oxide nanoparticles showed negligible effects at the phylum level. Thus, silver nanoparticles may impact bacterial communities in soil, and this in turn may influence processes carried out by soil bacteria.
1. Introduction
The use of engineered nanoparticles in a wide range of consumer and industrial products is increasing globally. Silver nanoparticles are being utilized for their antimicrobial properties in textiles, food packaging, wound dressings, medical devices like catheters, paints, water treatment processes, etc. They also find a use in optoelectronics, catalysis, surface-enhanced Raman scattering (SERS), and nanoparticle inks for printed electronics [1,2]. Zinc oxide nanoparticles have antimicrobial and UV protective properties and hence find a use in sunscreens, paints, cosmetics, food packaging, personal care products, etc. [3,4]. With the global production of nanoparticles expected to grow to over half a million tons by 2020, the release of nanoparticles into the environment is also bound to increase [5]. Data about nanoparticles entering the environment, and their concentrations therein, are limited. In the US, 94% of nanomaterials released into the environment from the use of personal care products are zinc oxide (1800–2100 t/year) and titanium dioxide (870–1000 t/year) [6]. In Singapore, the estimated range of release to the aquatic environment of zinc oxide and silver nanoparticles is 1.2–272 t/year and 26.7–27.5 t/year, respectively, from personal care products [7]. Predicted environmental concentrations range from 0.088–10,000 ng/L in surface water to 1.29–39 mg/kg in waste water treatment plant sludge for silver nanoparticles and 1–10,000 ng/L in surface water to 13.6–64.7 mg/kg in waste water treatment plant sludge for zinc oxide nanoparticles [5]. The concentration of zinc oxide nanoparticles in soil is predicted to be in the range of 0.026–100 µg/kg [8].
The field of nanotechnology is also being explored as a new source of key improvements in the agricultural sector. Silver and zinc oxide nanoparticles have been reported to have anti-phytopathogenic activity, increase germination of seeds, and improve plant growth [9]. Agriculture plays a significant role in the overall socio-economic fabric of India. Indian soils are zinc deficient and it has been recommended that 15 to 30 kg/ha of zinc be applied annually in order to maintain an adequate zinc nutritional status of crop plants in India [10]. Much of this zinc may be applied as nanoparticulate formulations [11,12]. Engineered nanoparticles will thereby gain entry into soil directly through nanoparticulate agrochemicals, and indirectly via the use of consumer and industrial products. This may lead to higher concentrations of zinc oxide nanoparticles in Indian soils, than what is currently predicted.
Therefore, monitoring the effects of engineered nanoparticles on ecologically important microbes in soil is the need of the day.
Soil microorganisms play an important role in agricultural productivity [13]. They recycle elements, break down organic matter, and promote plant growth [14]. A direct promotion of plant growth is brought about when these bacteria use any of the following mechanisms—nitrogen fixation, phosphate solubilization, and synthesis of phytohormones, siderophores, and antibiotics [15,16,17,18,19]. Biofilm formation is a property in plant growth-promoting bacteria that enables them to resist abiotic stresses [20]. Additionally, many bacteria promote plant growth indirectly by preventing the proliferation of plant pathogens [17]. Due to these properties, plant growth-promoting bacteria are used as soil inoculants to increase agricultural productivity [21,22,23,24]. With soil bacteria playing such a vital role, it is essential to study the impact of engineered nanoparticles on these bacteria. However, most studies about the impact of silver and zinc oxide nanoparticles have used pure cultures of common laboratory organisms such as Escherichia coli, Staphylococcus, Bacillus, and Pseudomonas, or clinically important bacteria [25,26,27,28].
Few studies have used pure cultures of ecologically important isolates to study the effects of silver and zinc oxide nanoparticles [29,30,31,32,33]. Similar disturbances in soil bacterial communities caused by the same nanoparticles have also been reported [34,35,36,37,38,39]. These disturbances were monitored by determining changes in microbial activity rates, microbial biomass, or microbial community composition. Of these, the analysis of microbial community composition provides more detailed information on phyla that may be affected. Sequencing and phylogenetic analysis of 16S rRNA genes using next generation sequencing (NGS) technologies represent one of the tools to determine the complexity of microbial communities in different environments [40]. This tool allows the faster generation of genetic profiles of culturable and unculturable bacteria from environmental samples [41].
Our study initially determined the effect of silver and zinc oxide nanoparticles on pure cultures of bacteria known to be agriculturally relevant—nitrogen fixers, phosphate solubilizers, and biofilm formers [42]. Silver and zinc oxide nanoparticles were then applied to soil microcosms, and preliminary bacterial community composition analysis was performed using Illumina MiSeq sequencing technology. This permitted a rapid means to explore the effects of these nanoparticles on the different bacteria in soil. The concentrations of nanoparticles that were sublethal for the pure cultures were added to the microcosms. Even though these concentrations were higher than the currently predicted environmental levels, the rationale was to use concentrations that would in all probability impact bacterial communities in soil.
There have been no studies in the Indian context on the impact of nanoparticles on bacterial community composition. Since soil characteristics are known to influence the toxicity of nanoparticles, this exploratory work is relevant due to the use of laterite soil commonly found in India [43,44]. It will add much needed data to the existing knowledge about the inhibitory effect of silver nanoparticles on bacteria in different soil types, enabling better environmental risk assessment.
2. Materials and Methods
2.1. Synthesis and Characterization of Nanoparticles
Reduction of silver nitrate using citrate was carried out to synthesize silver nanoparticles. Silver nitrate (10 mg) was dissolved in 100 mL deionized water and brought to boiling. A solution of 30 mM sodium citrate (10 mL) was added to the flask dropwise. Solution was boiled till it gradually turned yellow indicating the formation of silver nanoparticles [45]. The engineered nanoparticles were then analyzed using UV-Vis spectroscopy and transmission electron microscopy (Tecnai 20, Philips, Amsterdam, Netherlands). Zinc oxide nanoparticles were synthesized by wet chemical method [46]. A solution of 0.1 M zinc nitrate was added to 500 mL of 0.5% starch solution. Zinc nitrate was then completely dissolved by constantly stirring it using a magnetic stirrer. After complete dissolution of zinc nitrate, 20 mL of 0.2 M of sodium hydroxide were added dropwise under constant stirring. The reaction was allowed to proceed for 2 h after complete addition of sodium hydroxide. Once the reaction was complete, the solution was allowed to settle overnight and the supernatant solution then discarded. The remaining solution was centrifuged at 10,000× g for 10 min and the supernatant discarded. The nanoparticles were washed using distilled water. After washing, the nanoparticles were dried at 80 °C. During the drying process, complete conversion of zinc hydroxide into zinc oxide took place. The nanoparticles were then characterized by UV-Vis spectroscopy and X-ray diffraction (Philips® PW1710 X-ray diffractometer using Cu Kα radiation, λ = 1.54 Å, scan range 2θ = 20° to 80°, Philips, Amsterdam, Netherlands). The diffracted intensities were recorded from 20–80° 2θ angles. Scherrer’s formula, D = K λ/FWHM cosθ, was used for particle size determination.
2.2. Bacterial Strains
Agricultural soil and garden soil samples were collected from different parts of Maharashtra, India. Nitrogen fixers (symbiotic and free living), phosphate solubilizers (gram-positive and gram-negative), and biofilm formers (forming biofilm at air–liquid interface and forming adherent biofilm) were isolated from these samples using standard isolation methods. Although nitrogen fixing ability was not experimentally determined for the isolates, they were denoted as nitrogen fixers based on their phylogenetic affiliation [47].
Briefly, symbiotic nitrogen fixer was isolated from root nodules of Trigonella foenumgraecum on yeast extract mannitol medium (HiMedia, Mumbai, India). Free living nitrogen fixer was isolated using Azotobacter Mannitol medium (HiMedia). Pikovskaya’s (HiMedia) and NBRIP (National Botanical Research Institute’s phosphate) growth medium media [48] were used for isolation of phosphate solubilizers. Biofilm producers were screened on Congo red agar [49] and their adherent characteristic was studied by the microtiter plate method using crystal violet [50]. Bacteria-forming biofilms at air–liquid interface were identified by growing them in liquid medium. These isolates were then identified by 16S rDNA sequencing. Gen Bank accession numbers of the isolates are MH884596–MH884601.
Standard cultures of symbiotic nitrogen fixer, Rhizobium leguminosarum MTCC 10096, free living nitrogen fixer, Azotobacter chroococcum MTCC 3853, phosphate solubilizers, Arthrobacter sp. MTCC 8160 (gram-positive) and Serratia marcescens MTCC 7642 (gram-negative), and biofilm formers, Bacillus subtilis MTCC 441 (air–liquid interface biofilm former and Pseudomonas aeruginosa MTCC 7763 (adherent biofilm former), obtained from MTCC, Chandigarh, India, were also used in the study.
2.3. Determination of Antibacterial Activity of Nanoparticles
The toxicity of silver and zinc oxide nanoparticles was determined using pure cultures. Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) values are standard measures of toxicity in microbiology. Effects of antibacterial substances are often studied at sub-lethal (sub-MIC) concentrations.
2.3.1. MIC/MBC Determination
MIC and MBC of silver and zinc oxide nanoparticles were determined using broth micro dilution and tube dilution methods. Microorganisms in broth were exposed to serial dilutions of the nanoparticles and incubated for 24–96 h at room temperature under shaker conditions and the end points determined when no turbidity was observed. MBC concentrations were determined by agar plate method. Contents from tubes/wells that did not show growth were transferred to suitable agar media without nanoparticles and checked for growth. The lowest concentration that did not show growth on agar plates was noted as the MBC value.
2.3.2. Growth Curves
To determine growth behavior in the presence of the nanoparticles, the organisms were grown in broth with nanoparticles (sub-MIC) in 96-well microtiter plates. The isolates were grown for 24–48 h on solid medium and a saline-washed suspension of the isolate was prepared. The density was adjusted to 107 cells/mL. Broth containing nanoparticles (180 µL) was added to wells and these wells were inoculated with 20 µL of the culture suspension. Three types of control wells were maintained—only medium, medium with only cultures, and medium without cultures but containing the same concentration of nanoparticles (to check for any optical interference due to nanoparticles). The microtiter plates were kept at shaker conditions and optical density (OD) of the wells monitored every 30 min for 24–48 h using an EPOCH 2 Microplate Spectrophotometer (BioTek Instruments, Winooski, VT, USA).
2.4. Microcosm Study
Effect of nanoparticles on soil bacteria was studied in microcosms. The exposure doses were selected such that they were within the range of MIC values determined for pure cultures. For both types of nanoparticles, these were sub-lethal to lethal concentrations.
2.4.1. Microcosm Setup
The soil used for the study was obtained from agricultural land at Sarshi, Maharashtra, India (19.893252 N, 73.028924 E). This region has a tropical climate with an average annual temperature of 26.5 °C and average precipitation is 2101 mm. The soil type is classified as fine, montmorillonitic (calcareous), isohyperthermic, Vertic Halaquepts. These are soils with clay content greater than 30% [51]. Samples (maximum 10 cm depth) were obtained from five different points. Debris (plant matter, rocks, and wood chips) were removed manually and afterwards they were mixed into a single composite sample. pH of soil was found to be 6.9.
The soil was sieved using 4 mm mesh, and immediately conserved at 4 °C until use. Sieving achieved a homogenous mixture of soil particles to enable better contact with the nanoparticles.
Three types of microcosms, each of 50 g, were set up: (1) only soil (1C); (2) soil with silver nanoparticles added to give a final concentration of 15 µg of nanoparticles/g of soil (2A); and (3) soil with zinc oxide nanoparticles added to give a final concentration of 500 µg of nanoparticles/g of soil (3Z). Each type of microcosm was set up in triplicates (1Ci,1Cii,1Ciii; 2Ai,2Aii,2Aiii; 3Zi,3Zii,3Ziii) and incubated at room temperature for four days.
2.4.2. Soil DNA Extraction, PCR Amplification and Sequencing
After incubation, DNA was extracted from 500 mg of soil samples from each microcosm using HiPurA™ Soil DNA Purification Kit (HiMedia, India) as per the manufacturer’s instructions. The DNAs from microcosms 1Ci,1Cii,1Ciii were pooled and labeled as 1C. Similarly DNAs from the other microcosms were pooled and labeled as 2A and 3Z, respectively. All the pooled DNAs were quantified using a NanoDropTM 2000 Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA). DNA was amplified and sequenced on an Illumina MiSeq platform at AgriGenome (SciGenom) Labs Private Ltd., Cochin, Kerala, India. The V3 PCR primers used were 341F 5′CCTACGGGAGGCAGCAG 3′ and 518R 5′ATTACCGCGGCTGCTGG 3′ and PCR master mix contained 2 μL each of 10 pmol/uL forward and reverse primers, 0.5 μL of 40 mM dNTP, 5 μL of 5× Phusion HF reaction buffer, 0.2 μL of 2 U/μL F-540 Special Phusion HS DNA Polymerase, 5 ng input DNA and water to make up the total volume to 25 μL. Conditions for PCR were as follows: 98 °C for 30 s, 30 cycles of 98 °C for 10 s, 72 °C for 30 s, extension at 72 °C for 5 s followed by 4 °C hold. The PCR product was run on 2% Agarose gel with SYBR® Safe DNA gel stain (10 μL/100 mL). PCR products were purified using InvitrogenTM PureLinkTM Quick Gel Extraction and PCR Purification Combo Kit (Thermo Fisher Scientific, Waltham, MA, USA). The PCR products were quantified using the Qubit® 2.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA, USA). The NEBNext® Ultra® DNA Library Prep Kit for Illumina (New England Biolabs, Ipswich, MA, USA) was used to prepare the DNA library according to the manufacturer’s instructions. Briefly, the steps involved end repair and dA tailing followed by adapter ligation. Clean up of adaptor ligated DNA was then performed using AMPure XP beads. The adaptor-ligated DNA was then enriched by PCR (conditions described earlier). The PCR products were then cleaned using AMPure XP beads and size distribution of the library confirmed by running it on an Agilent Bioanalyzer® (Agilent, Santa Clara, CA, USA).
2.4.3. Data Analysis
Raw sequence data generated from Illumina sequencing were processed and analyzed using the Quantitative Insights into Microbial Ecology (QIIME) software package, version 1.9.0 (http://qiime.sourceforge.net/) [52]. All sequences have been submitted to Sequence Read Archive, NCBI (accession number SRP145628). After checking quality parameters, V3 regions were extracted from Illumina paired-end sequences. Chimeras were removed using the de novo chimera removal method UCHIME implemented in the tool USEARCH. Pre-processed reads from the sample were clustered into Operational Taxonomic Units (OTUs) based on their sequence similarity using the Uclust program (similarity cutoff = 0.97) [53,54]. After removal of singleton OTUs from the total number of OTUs picked, the OTUs were taken for further analysis. The QIIME program was used for the entire downstream analysis. Representative sequence was identified for each OTU and aligned against Greengenes core set of sequences using the PyNAST program [55,56,57]. Further, this representative sequence was aligned against reference chimeric datasets. Then, taxonomy classification was performed using the RDP classification system. The generated OTU table was rarefied to an even sampling depth (115,508 sequences per sample) using the single_rarefaction.py script in the QIIME pipeline, and this rarefied OTU table was used to calculate alpha diversity metrics, including OTU abundance, Chao1, phylogenetic tree (PD_whole tree), and Shannon’s index.
3. Results
3.1. Synthesis and Characterization of Nanoparticles
The silver nanoparticles synthesized were found to be 11.79 ± 4.74 nm in size and showed an absorption maximum at 416 nm (Supplementary Materials, Figure S1). The absorption spectrum between 200 to 800 nm showed maximum absorption at 360 nm for zinc oxide nanoparticles. X-ray diffraction pattern peaks at different crystal planes of nano-ZnO match exactly with that of bulk ZnO (Supplementary Materials, Figure S2). Crystallite size estimated from XRD data using Scherrer’s formula indicates the presence of nanoparticles of zinc oxide (Supplementary Materials, Table S1).
3.2. Identification of Bacterial Isolates and Determination of MIC/MBC of Nanoparticles for the Isolates
Bacterial strains isolated from soil were identified (Table 1). Gen Bank accession numbers of the isolates are MH884596–MH884601. Both standard cultures and soil isolates were used for MIC and MBC determination. Silver nanoparticles were found to have MIC/MBC in the range of 2 to 22 µg/mL and MIC of zinc oxide nanoparticles ranged from 80 µg/mL to greater than 3000 µg/mL. MBC values were higher, ranging from 150 µg/mL to greater than 3000 µg/mL (Table 1).
3.3. Effect of Silver and Zinc Oxide Nanoparticle Concentrations That Are Sub-MIC
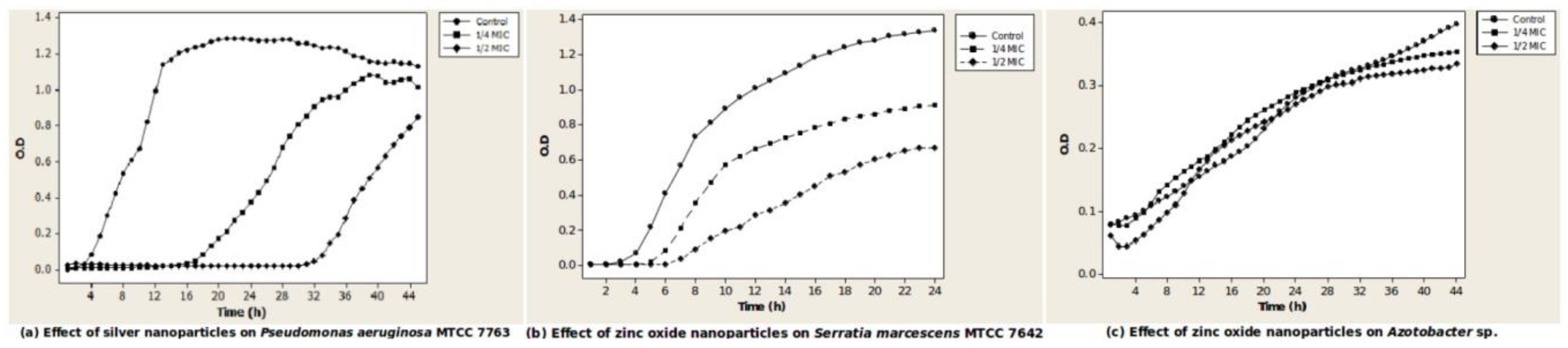
The bacteria showed different types of growth responses to concentrations of nanoparticles that were sub-MIC. At these values, there was either a long lag with some inhibition of growth, no lag with some inhibition of growth, or no change. Sinorhizobium meliloti clsxc_S_SNF and Pantoea dispersa clsxc_PSD showed a long lag of 14 h (1/4 MIC) to 30 h (1/2 MIC) in the presence of ZnO nanoparticles. Azotobacter chroococcum clsxc-a_FLNF, Azotobacter chroococcum MTCC 3853, Pseudomonas aeruginosa MTCC 7763 (Figure 1a), and Serratia marcescens MTCC 7642 showed a long lag of 16 h (1/4 MIC) to 32 h (1/2 MIC) in the presence of Ag nanoparticles. Bacillus subtilis MTCC 441 and Bacillus sp. clsxc_TYA showed a long lag in the presence of both Ag and ZnO nanoparticles.
Shorter lags of 2 h (1/4 MIC) to 6 h (1/2 MIC) were seen in the case of Serratia marcescens MTCC 7642 (Figure 1b) and Pseudomonas aeruginosa MTCC 7763 in the presence of ZnO nanoparticles. In the presence of Ag nanoparticles, a lag of 6 h was seen in the case of Bacillus sp. clsxc_NPS and Pantoea dispersa clsxc_PSD.
Inhibition of growth to an extent of 50% was seen in Sinorhizobium meliloti clsxc_S_SNF, Arthrobacter sp. MTCC 8160, Bacillus sp. clsxc_NPS, Bacillus subtilis MTCC 441, Pantoea dispersa clsxc_PSD, and Serratia marcescens MTCC 7642 (Figure 1b) when exposed to ZnO nanoparticles. Most cultures showed some inhibition on exposure to Ag nanoparticles. Two cultures, Rhizobium leguminosarum MTCC 10096 and Azotobacter chroococcum clsxc_A-FLNF (Figure 1c), when exposed to ZnO nanoparticles, showed growth response similar to control. Growth response of all isolates to concentrations of Ag and ZnO nanoparticles that were sub-MIC is included in the Supplementary Materials, Figure S3.
3.4. Sequencing Summary and Bacterial Community Diversity
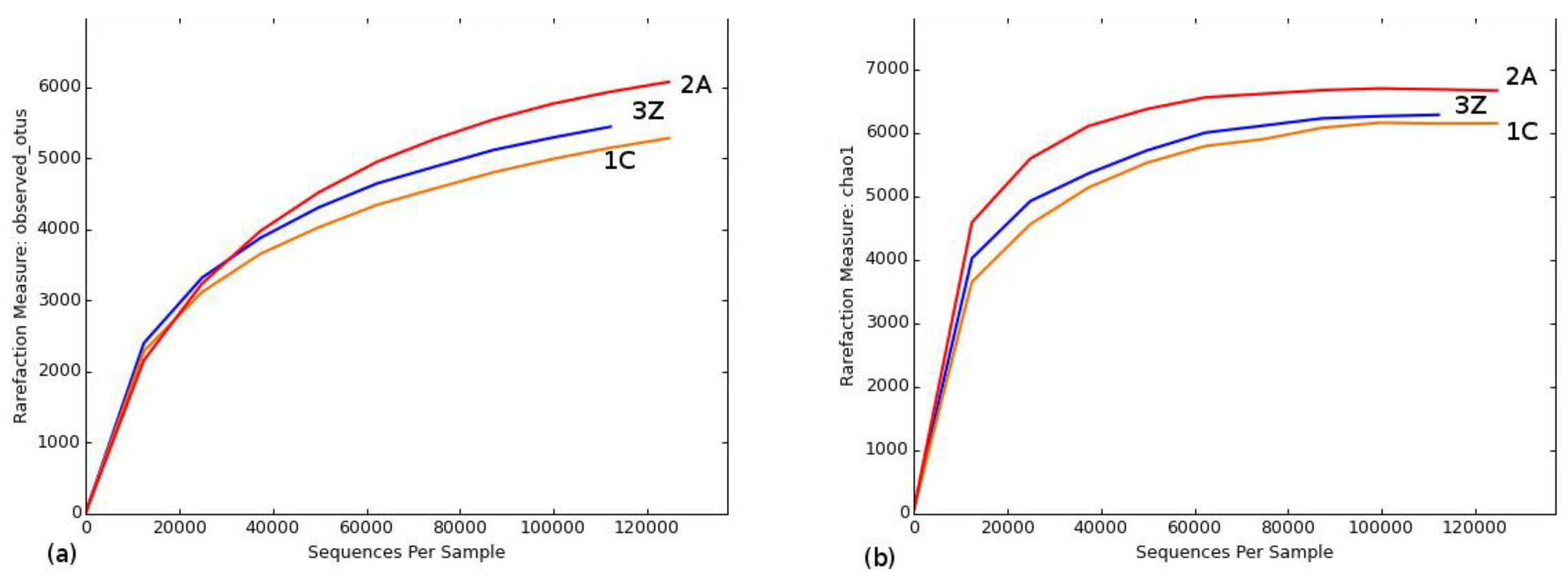
A total of 365,617 bacterial 16S rRNA gene sequences were obtained following assembly and quality filtering from the three soil samples, namely, control soil (1C), Ag nanoparticle-treated soil (2A), and ZnO nanoparticle-treated soil (3Z). The number of sequences obtained was sufficient to describe the bacterial diversity within these samples, as is evident from the fact that all rarefaction curves tended to approach a plateau (Figure 2 and Supplementary Materials, Figure S4). Table 2 shows that alpha diversity estimates of operational taxonomic unit (OTU) abundance increased in sample 2A (6077) as compared to sample 1C (5286). Chao richness estimates also increased in sample 2A (6153 to 6667) and sample 3Z (6153 to 6286). Phylogenetic diversity (PD_whole tree) increased for sample 2A (133 to 149). As can be seen, bacterial community in sample 2A is the most phylogenetically diverse. Shannon’s diversity estimate for sample 2A decreased (Table 2).
3.5. Bacterial Community Composition
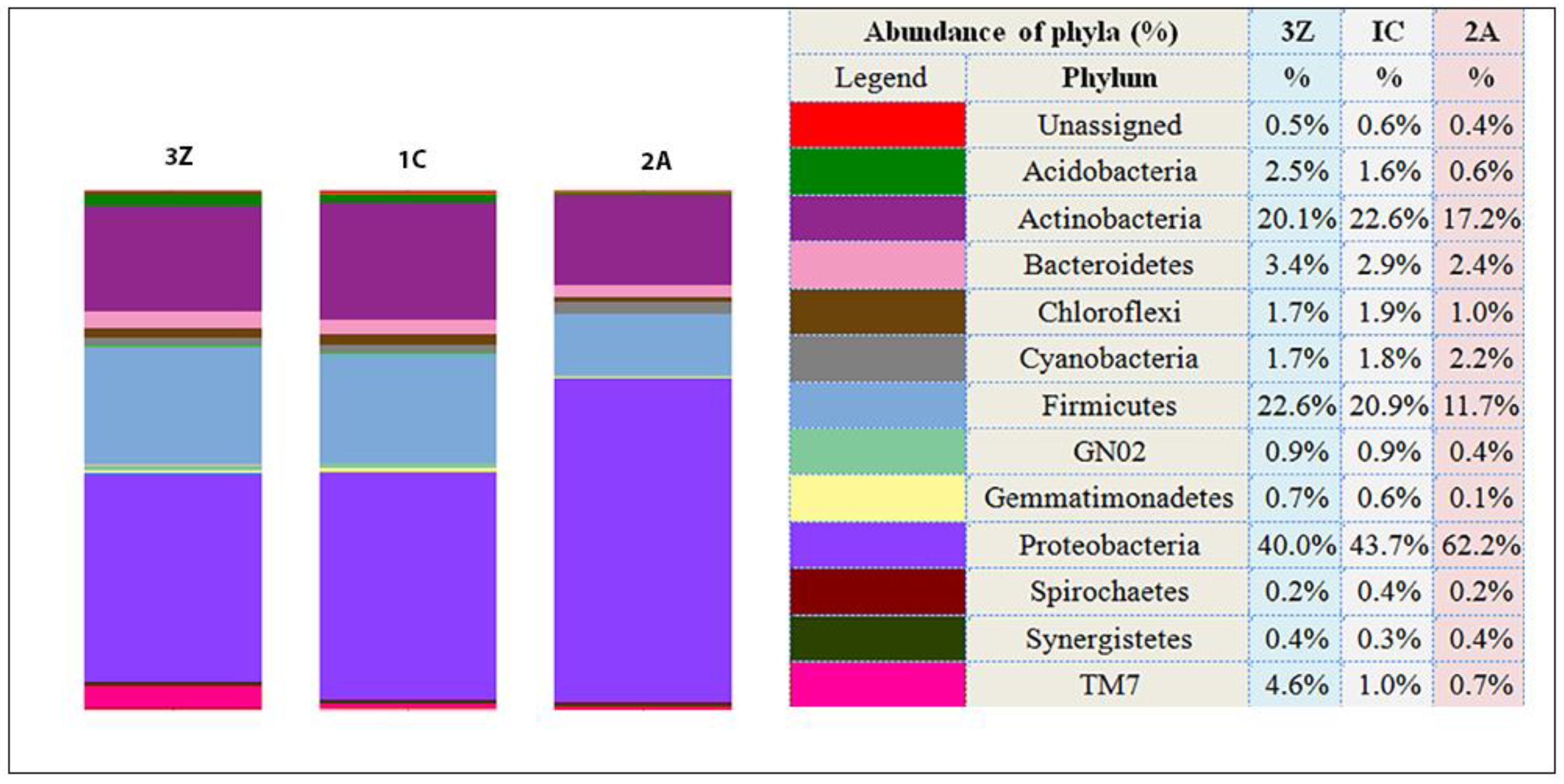
Proteobacteria was the most dominant phylum (greater than 40% of all sequences) in all samples, but change in this phylum was evident in the Ag nanoparticle-treated sample (2A) with abundance of sequences increasing from 43.7% in the control (1C) to 62.2% in sample 2A (30% increase). In the other two dominant phyla, Firmicutes and Actinobacteria, abundance of affiliated sequences decreased from 20.99% in sample 1C to 11.7% in sample 2A and from 22.6% in sample 1C to 17.2% in sample 2A, respectively (Figure 3).
ZnO nanoparticle treatment (3Z) seems to have brought about only a slight change, with members of Proteobacteria and Actinobacteria decreasing to 40% from 43.7% and 20.1% from 22.6%, respectively. Phylum Firmicutes members increased marginally from 20.99% to 22.6%.
Among the minor components of the bacterial communities in the samples, relative abundance of members of phyla Acidobacteria and Chloroflexi reduced in sample 2A and that of the TM7 (Candidatus Saccharibacteria) group increased over four times in sample 3Z (Figure 3).
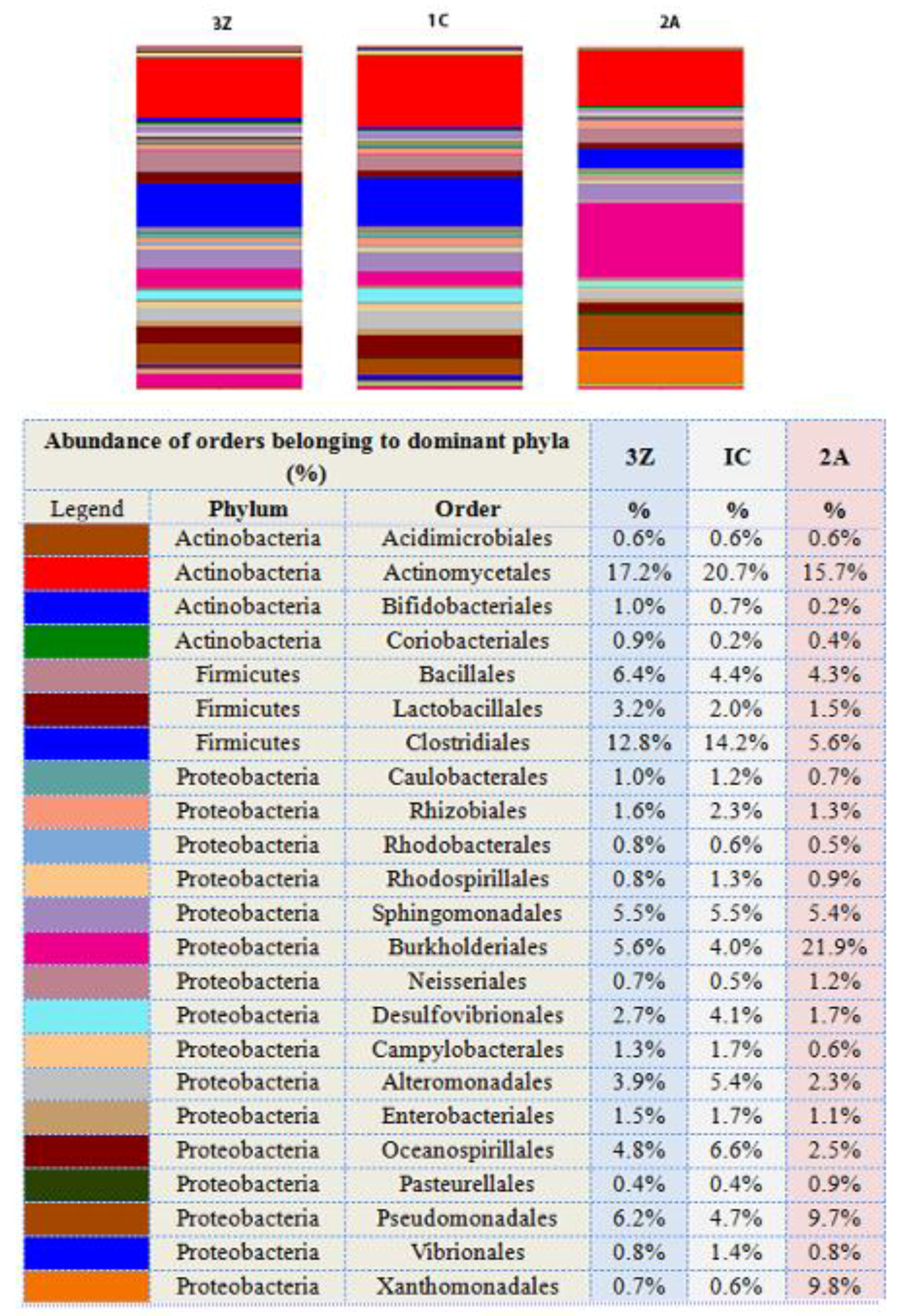
Bacterial community abundance analysis at lower taxonomic levels showed that in twelve out of twenty-three orders of the three dominant phyla, there was a concordant decrease in the number of the sequences affiliated with the three phyla with respect to control (1C) in both silver nanoparticle-treated soil (2A) and zinc oxide nanoparticle-treated soil (3Z) (Figure 4). The decrease in number of sequences in silver nanoparticle-treated soil was more than that in zinc oxide nanoparticle-treated soil.
Upon treatment with silver nanoparticles, the decrease in members of orders Clostridiales and Actinomycetales accounted for the reduction in abundance of the two phyla Firmicutes and Actinobacteria, respectively. On the other hand, although relative abundance decreased for most orders—Alteromonadales, Oceanospirillales, Enterobacteriales, Caulobacterales, Rhizobiales, Rhodospirillales, Desulfovibrionales, and Campylobacterales—belonging to Proteobacteria, the increase in overall abundance of members of this phylum was due to increases in only three orders, that is, Burkhoderiales, Pseudomonadales, and Xanthomonadales.
A look at specific genera showed an increase in numbers of Stenotrophomonas, uncultured Xanthomonadaceae, Pseudomonas, Commamonas, and Bacillus, and a decrease in Streptococcus, uncultured Lachnospiraceae, uncultured Ruminococcaceae, Oscillospira, and Halomonas in sample 2A (Supplementary Materials, Figure S5).
Zinc oxide nanoparticles may not have impacted dominant phyla as much as silver nanoparticles did, but relative abundance of members belonging to some orders decreased—Actinomycetales (16%), Desulfivibrionales (34%), Alteromonadales (27%), and Oceanospirillales (27%). In contrast, an increase was seen in members of orders Bacillales (45%), Lactobacillales (60%), and Pseudomonadales (31%). Specific genera Pseudomonas, Streptococcus, and Dialister increased, whereas Halomonas decreased (Supplementary Materials, Figure S5).
Out of the nine genera used in the pure culture study, only three were present in the soil used—Bacillus, Pseudomonas, and Serratia.
4. Discussion
Silver and zinc oxide nanoparticles were chosen for the study as they have antibacterial properties and, therefore, their release into ecosystems raise environmental concerns [58,59].
This study determined impacts of nanoparticles on pure cultures of agriculturally relevant bacteria. These nanoparticles were inhibitory to the isolates. Ag nanoparticles were more bactericidal as the MIC of Ag nanoparticles ranged from 2 to 22 µg/mL with almost similar MBCs, whereas zinc oxide nanoparticles were inhibitory at 80 to >3000 µg/mL. The MBCs of ZnO nanoparticles were higher, indicating the bacteriostatic nature of zinc oxide nanoparticles. The range of MIC/MBC values of Ag nanoparticles matched those from earlier studies [27,60,61].
The MIC/MBC values of ZnO nanoparticles reported in previous studies have ranged from as low as 8–18 µg/mL [62,63] to 0.5–1 mg/mL [64,65], whereas the MIC/MBC values as determined in this study are in the range of 0.08 to >3 mg/mL. The difference in values observed in this study from the earlier reported values could be due to the fact that many factors are known to influence MIC values. The growth medium, size, shape, and charge of nanoparticle, stabilizing agents, and the method used are some of these factors [66]. The time of incubation could also play a role; for example, the incubation time used by most studies to determine MIC is 24 h, whereas we followed the growth for 48 h. This allowed us to monitor growth of cultures in the presence of Ag nanoparticles, where Pseudomonas aeruginosa MTCC 7763 and Serratia marcescens MTCC 7642 (Supplementary Materials, Figure S3) showed a long lag of 16–32 h at sub-MIC values. The long lag shown by Serratia and Pseudomonas was not reported earlier. The cultures that showed a long lag were inhibited at higher nanoparticle concentrations. It has been suggested that bacteria probably adapt to the nanoparticles or modify them to make them less toxic during the lag period [31]. Such adaptation by some bacteria could give them a survival advantage and this could lead to disturbance in composition of bacterial populations in soil.
Exposure doses for the microcosm studies are higher than the currently predicted environmentally relevant concentrations. However, concentrations in soil may be more, especially at points of release and in soils where nanoparticulate formulations are used as agrochemicals.
With pure culture investigations, impacts of nanoparticles were determined on only a few genera of bacteria. Illumina MiSeq sequencing of 16S rRNA genes on the other hand offered a broader perspective of all bacteria present in soil, both culturable and non-culturable. Identity of bacteria determined by sequencing cannot always be equated to function. However, it could be indicative of bacterial contributions to soil health. Proteobacteria, Actinobacteria, and Firmicutes were found to be the three dominant phyla in the soil used, based on sequence data analysis. The Proteobacteria are a metabolically diverse group commonly found in soil with many members classified as plant growth-promoting bacteria that facilitate nutrient acquisition and provide protection against diseases [67,68]. Actinobacteria and Firmicutes are ubiquitous in terrestrial ecosystems and play a significant role in the formation of soil organic matter by mediating decomposition of plant litter. Actinobacteria are especially important in extreme environments [69]. Actinobacteria are plant growth promoters acting as biocontrol agents, nitrogen fixers, and activators of plant self-defense [70]. Aerobic endospore-formers belonging to the phylum Firmicutes are also important in soil being endophytic and having various plant growth-promoting characteristics like phosphate solubilization, siderophore and indole acetic acid production, and multiple enzyme activities [68,71]. All these three dominant phyla were found to be impacted by exposure to the two nanoparticles. Increase in Proteobacteria and decrease in Actinobacteria as seen in this study have also been observed in soil exposed to 10–100 µg of silver nanoparticles/g of soil for seven days [72].
Of the nine genera used in the pure culture study, three were found in the soil used. Effects of nanoparticles on pure cultures used in the study matched the results obtained by soil microcosm sequencing. Some of the isolates used in the study were sensitive to 15 µg/mL of silver nanoparticles and 500 µg/mL of zinc oxide nanoparticles. Bacterial community composition analysis showed that the abundance of bacterial taxa that these isolates belong to decreased in nanoparticle-treated soil microcosms. On the other hand, MICs of silver nanoparticles for Pseudomonas aeruginosa MTCC 7763, Bacillus subtilis MTCC 441, and Bacillus sp. clsxc TYA were greater than 15 µg/mL and, therefore, the abundance of the bacterial taxa that these isolates belong to remained unchanged.
The exception was Bacillus which was not affected by zinc oxide nanoparticles in the soil, whereas based on MIC/MBC values for pure cultures a reduction in members of the class Bacilli was expected. However, as can be seen from the MIC values of zinc oxide nanoparticles for the three Bacillus sp. used in the study, individual species respond differently to the same nanoparticles. Thus, extrapolating from effects seen for an individual species to a whole group should be done with caution.
Common nitrogen fixers in the soil such as Rhizobium, Azotobacter, Bradyrhizobium, and Beijerinckia were not influenced by the presence of nanoparticles. Since phosphate solubilization and biofilm formation abilities are found in many bacteria across different phyla, it was not possible to correlate bacterial groups based on sequence to these functions. However, among the specific genera that were impacted, it was seen that Stenotrophomonas sp. and Pseudomonas sp., with their multiple plant growth-promoting abilities including nitrogen fixation, phosphate solubilization, and production of extra polymeric substances (EPS), increased in the presence of silver and zinc oxide nanoparticles, respectively [73,74]. Abundance of Halomonas that are known to be nitrogen fixers, phosphate solubilizers and EPS producers decreased in the presence of both nanoparticles [75]. Thus, it is difficult to reach a conclusion about the effect of nanoparticles on functions of bacteria.
A closer look at the overall bacterial community in soil revealed a decrease in the relative abundance of members of most orders in response to both nanoparticles. A significant order that decreased was Rhizobiales. Bacteria belonging to this order are agriculturally significant since they oxidize methane to CO2 and also fix nitrogen. Relative abundance of the order Sphingomonadales did not change. Collins et al. [34] reported that members of Rhizobiales showed less sensitivity and Sphingomonadales showed high sensitivity to ZnO nanoparticles, whereas a decrease in Rhizobiales and an increase in abundance of Sphingomonadaceae were observed by Ge et al. [76] in response to ZnO nanoparticles.
Increase in relative abundance of sequences was seen only for the orders Pseudomonadales and Burkholderiales in both Ag and ZnO nanoparticle-treated soil, but increase was drastic for members of Burkholderiales in Ag nanoparticle-treated soil. Increase in members of Pseudomonadales and Burkholderiales could have both positive and negative connotations. Both orders include plant pathogens causing rots, necrosis, and galls, but strains of Burkholderia are also environmentally important as they are good biodegraders of poly-chlorinated biphenyls. Pseudomonas is known to promote plant growth by synthesizing plant hormones and suppressing plant pathogens [74].
Abundance of Xanthomonadales sequences increased only in Ag nanoparticle-treated soil. Xanthomonadales includes the largest group of phytopathogens and their increase in soil could be detrimental to plant productivity [77]. Increase in abundance of Pseudomonadales, Burkholderiales, and Xanthomonadales may not be due to an enhancement effect of specific nanoparticles per se, but more likely a result of decreased competition from the bacteria that were inhibited.
Members of two orders responded differently to the two nanoparticles. Relative abundance of Bacillales increased in response to treatment with ZnO nanoparticles with almost no change in Ag nanoparticle-treated soil. Lactobacillales also showed an increase in ZnO nanoparticle-treated soil and a slight decrease in Ag nanoparticle-treated soil.
Increase in members of the orders Burkholderiales and Bacillales in the presence of ZnO nanoparticles has also been reported by Meli et al. [78]. Tolerance of Bacilli to ZnO nanoparticles could be explained by their ability to form endospores as also the production of EPS [61,79]. Members of this class are also known to persist in metal-polluted soils [80,81].
Along with shifts in abundance, alpha diversity metrics seem to indicate bacterial diversity changes in silver nanoparticle-treated soil. The preliminary nature of the work does not permit definite conclusions to be drawn with respect to diversity. However, changes in soil bacterial diversity have been described in earlier studies in the presence of silver nanoparticles [38,82,83,84,85].
Overall, there was a concordant decrease in relative abundances of most orders of the three dominant phyla on exposure to both nanoparticles. The decrease in Ag nanoparticle-treated soil was more than that in ZnO nanoparticle-treated soil.
Both the pure culture study and the soil microcosm study confirm the inhibitory effects of silver nanoparticles on soil bacteria. Perturbations in microbial community structure due to these nanoparticles have also been reported in other soils [86,87,88]. These studies have used a wide range of concentrations of silver nanoparticles from 0.01 µg/g to 1815 µg/g of soil and varying exposure times, of one day to one year [39,72,82,83,84,85,89]. Two of these studies have also described bacterial communities recovering after 21 to 50 days following an initial shift [35,84]. Studies with longer exposure times have seen a definite impact on community compositions [82,89]. Resistant bacteria and persisters have been characterized in soil exposed to silver nanoparticles [82,85]. Nitrification, a key step in the nitrogen cycle, and activities of many soil enzymes were found to be affected in the presence of silver nanoparticles [72,83,85,90].
As compared to silver nanoparticles, effects of zinc oxide nanoparticles on soil bacteria are less studied [87,88]. Although the present study showed negligible effect of zinc oxide nanoparticles on soil bacteria, other studies have shown that they affect soil bacteria as well as soil enzymes at concentrations ranging from 0.1 to 2000 µg/g of soil [34,36,91,92]. Many factors are known to influence metal nanoparticle toxicity. Soil pH is thought to play an important role in zinc oxide nanoparticle toxicity, with toxicity enhanced at acidic pH [93]. Since this soil had a pH of 6.9, it could be the reason for the minimal effect seen.
While examining inferences based on such studies, it has to be kept in mind that there can be inherent bias in some of the procedures used. Sieving of soil can bring about change in microbial activity and community composition [94,95,96]. Bias associated with DNA extraction and PCR and sequencing techniques are also known [97,98,99,100]. Moreover, the choice of bioinformatics pipeline influences the analyses [101,102].
This is an exploratory study to examine the impact(s) of silver and zinc oxide nanoparticles on soil bacteria in an Indian soil type. In the presence of these nanoparticles, pure cultures of agriculturally relevant bacteria showed changes in growth patterns. When soil microcosms were exposed to the same nanoparticles, we observed changes in soil bacterial abundance and diversity. The effects seen on soil bacteria need to be supported with further comprehensive investigation. However, the inhibitory concentrations observed here may assist regulatory bodies determine hazardous levels of these emerging contaminants.
5. Conclusions
The study evaluated the effects of silver and zinc oxide nanoparticles on representatives of three groups of bacteria known to be agriculturally relevant—nitrogen fixers, phosphate solubilizers, and biofilm formers. Silver nanoparticles were bactericidal at relatively lower concentrations (2–22 µg/mL). Zinc oxide nanoparticles exhibited a bacteriostatic effect and the inhibitory concentrations in some cases were higher than 3000 μg/mL. Continuous monitoring of microbial growth in the presence of sub-lethal concentrations of the engineered nanoparticles enabled observation of finer changes in growth response. A few isolates showed a very long lag with decreased growth which may also occur in soil. In addition, soil bacterial community composition showed a pronounced change in the presence of silver nanoparticles. It may seem premature to correlate these changes in growth patterns and community composition to agricultural productivity, but it is a distinct possibility.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2073-4395/9/3/140/s1, Table S1: The XRD data of zinc oxide nanoparticles; Figure S1: Characterization of silver nanoparticles: (a) TEM of silver nanoparticles; (b) UV-visible spectrum of silver nanoparticles; Figure S2: Characterization of zinc oxide nanoparticles: (a) XRD patterns of ZnO nanoparticles; (b) UV-visible spectrum of zinc oxide nanoparticles; Figure S3: Growth response of isolates to Ag and ZnO nanoparticles (sub-MIC); Figure S4: Rarefaction curves of alpha diversity indices. (a) PD_whole_tree curve and (b) Shannon curve.; and Figure S5: Genus distribution in samples—1C, 2A, 3Z.
Author Contributions
Conceptualization and design, S.C. and V.N.; Methodology, S.C. and V.N.; Investigation, Analysis, S.C.; Writing—Original Draft Preparation, S.C.; Writing—Review and Editing, V.N.; Supervision, V.N.; Funding Acquisition, S.C.
Funding
This work was supported in part by funding from the University Grants Commission (UGC) of the Government of India (Minor research grant: 47-584/13(WRO)).
Acknowledgments
We thank Vishwas Sarangdhar for his valuable and constructive suggestions and useful critiques of this research work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bumbudsanpharoke, N.; Choi, J.; Ko, S. Applications of Nanomaterials in Food Packaging. J. Nanosci. Nanotechnol. 2015, 15, 6357–6372. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Li, Q.; Wang, M.; Ouyang, J.; Li, X.; Xing, M.M.Q. Nanosilver particles in medical applications: Synthesis, performance, and toxicity. Int. J. Nanomed. 2014, 9, 2399–2407. [Google Scholar] [CrossRef]
- Bangale, M.S.; Mitkare, S.S.; Gattani, S.G.; Sakarkar, D.M. Recent nanotechnological aspects in cosmetics and dermatological preparations. Int. J. Pharm. Pharm. Sci. 2012, 4, 88–97. [Google Scholar]
- Vance, M.E.; Kuiken, T.; Vejerano, E.P.; McGinnis, S.P.; Hochella, M.F.; Hull, D.R. Nanotechnology in the real world: Redeveloping the nanomaterial consumer products inventory. Beilstein J. Nanotechnol. 2015, 6, 1769–1780. [Google Scholar] [CrossRef]
- Maurer-Jones, M.; Gunsolus, I.; Murphy, C.; Haynes, C. Toxicity of Engineered Nanoparticles in the Environment. Anal. Chem. 2013, 85, 3036–3049. [Google Scholar] [CrossRef]
- Keller, A.A.; Vosti, W.; Wang, H.; Lazareva, A. Release of engineered nanomaterials from personal care products throughout their life cycle. J. Nanopart. Res. 2014, 16, 2489. [Google Scholar] [CrossRef]
- Zhang, Y.; Leu, Y.R.; Aitken, R.J.; Riediker, M. Inventory of engineered nanoparticle-containing consumer products available in the singapore retail market and likelihood of release into the aquatic environment. Int. J. Environ. Res. Public Health 2015, 12, 8717–8743. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.M.; Behal, A.; Sushkova, S.N.; Mandzhieva, S.; Singh, R.; Gorovtsov, A.; Tsitsuashvili, V.S.; Purvis, W.O.; Ghazaryan, K.A.; et al. Effects of zinc-oxide nanoparticles on soil, plants, animals and soil organisms: A review. Environ. Nanotechnol. Monit. Manag. 2018, 9, 76–84. [Google Scholar] [CrossRef]
- Gogos, A.; Knauer, K.; Bucheli, T.D. Nanomaterials in Plant Protection and Fertilization: Current State, Foreseen Applications, and Research Priorities Foreseen Applications, and Research Priorities. J. Agric. Food Chem. 2012, 60, 9781–9792. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Green, A. Importance of zinc in crops and human health. J. SAT Agric. Res. 2013, 11, 1–7. [Google Scholar]
- Prasad, T.N.V.K.V.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Reddy, K.R.; Sreeprasad, T.S.; Sajanlal, P.R.; Pradeep, T. Effect of Nanoscale Zinc Oxide Particles on the Germination, Growth and Yield of Peanut. J. Plant Nutr. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 15, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The Role of Soil Microorganisms in Plant Mineral Nutrition—Current Knowledge and Future Directions. Front. Plant Sci. 2017, 8, 1–19. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd-Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Plant GrowthPromoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability-A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- Bouskill, N.J.; Wood, T.E.; Baran, R.; Hao, Z.; Ye, Z.; Bowen, B.P.; Lim, H.C.; Nico, P.S.; Holman, H.Y.; Gilbert, B.; et al. Belowground response to drought in a tropical forest soil. II. Change in microbial function impacts carbon composition. Front. Microbiol. 2016, 7, 323. [Google Scholar] [CrossRef] [PubMed]
- Maurer, K.; Zachow, C.; Seefelder, S.; Berg, G. Initial Steps towards Biocontrol in Hops: Successful Colonization and Plant Growth Promotion by Four Bacterial Biocontrol Agents. Agronomy 2013, 3, 583–594. [Google Scholar] [CrossRef]
- De Souza, R.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef]
- Baez-Rogelio, A.; Morales-García, Y.E.; Quintero-Hernández, V.; Muñoz-Rojas, J. Next generation of microbial inoculants for agriculture and bioremediation. Microb. Biotechnol. 2017, 10, 19–21. [Google Scholar] [CrossRef]
- Finkel, O.M.; Castrillo, G.; Herrera Paredes, S.; Salas González, I.; Dangl, J.L. Understanding and exploiting plant beneficial microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef]
- Cavassin, E.D.; de Figueiredo, L.F.P.; Otoch, J.P.; Seckler, M.M.; de Oliveira, R.A.; Franco, F.F.; Marangoni, V.S.; Zucolotto, V.; Levin, A.S.S.; Costa, S.F. Comparison of methods to detect the in vitro activity of silver nanoparticles (AgNP) against multidrug resistant bacteria. J. Nanobiotechnol. 2015, 13, 64. [Google Scholar] [CrossRef]
- Radzig, M.A.; Nadtochenko, V.A.; Koksharova, O.A.; Kiwi, J.; Lipasova, V.A.; Khmel, I.A. Antibacterial effects of silver nanoparticles on gram-negative bacteria: Influence on the growth and biofilms formation, mechanisms of action. Colloids Surf. B Biointerfaces 2013, 102, 300–306. [Google Scholar] [CrossRef]
- Sondi, I.; Salopek-Sondi, B. Silver nanoparticles as antimicrobial agent: A case study on E. coli as a model for Gram-negative bacteria. J. Colloid Interface Sci. 2004, 275, 177–182. [Google Scholar] [CrossRef]
- Stanković, A.; Dimitrijević, S.; Uskoković, D. Influence of size scale and morphology on antibacterial properties of ZnO powders hydrothemally synthesized using different surface stabilizing agents. Colloids Surf. B Biointerfaces 2013, 102, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Peralta-Videa, J.R.; Plascencia-Villa, G.; José-Yacamán, M.; Gardea-Torresdey, J.L. Comparative toxicity assessment of CeO2 and ZnO nanoparticles towards Sinorhizobium meliloti, a symbiotic alfalfa associated bacterium: Use of advanced microscopic and spectroscopic techniques. J. Hazard. Mater. 2012, 241–242, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Beddow, J.; Stolpe, B.; Cole, P.; Lead, J.R.; Sapp, M.; Lyons, B.P.; Colbeck, I.; Whitby, C. Effects of engineered silver nanoparticles on the growth and activity of ecologically important microbes. Environ. Microbiol. Rep. 2014, 6, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Dhas, S.P.; Shiny, P.J.; Khan, S.; Mukherjee, A.; Chandrasekaran, N. Toxic behavior of silver and zinc oxide nanoparticles on environmental microorganisms. J. Basic Microbiol. 2014, 54, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Mirzajani, F.; Askari, H.; Hamzelou, S.; Farzaneh, M.; Ghassempour, A. Effect of silver nanoparticles on Oryza sativa L. And its rhizosphere bacteria. Ecotoxicol. Environ. Saf. 2013, 88, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Radniecki, T.S.; Stankus, D.P.; Neigh, A.; Nason, J.A.; Semprini, L. Influence of liberated silver from silver nanoparticles on nitrification inhibition of Nitrosomonas europaea. Chemosphere 2011, 85, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Collins, D.; Luxton, T.; Kumar, N.; Shah, S.; Walker, V.K.; Shah, V. Assessing the impact of copper and zinc oxide nanoparticles on soil: A field study. PLoS ONE 2012, 7, e42663. [Google Scholar] [CrossRef]
- Colman, B.P.; Arnaout, C.L.; Anciaux, S.; Gunsch, C.K.; Hochella, M.F.; Kim, B.; Lowry, G.V.; McGill, B.M.; Reinsch, B.C.; Richardson, C.J.; et al. Low Concentrations of Silver Nanoparticles in Biosolids Cause Adverse Ecosystem Responses under Realistic Field Scenario. PLoS ONE 2013, 8, e57189. [Google Scholar] [CrossRef]
- Ge, Y.; Schimel, J.P.; Holden, P.A. Evidence for negative effects of TiO2 and ZnO nanoparticles on soil bacterial communities. Env. Sci. Technol. 2011, 45, 1659–1664. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Shah, V.; Walker, V.K. Influence of a nanoparticle mixture on an arctic soil community. Environ. Toxicol. Chem. 2012, 31, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Pallavi; Mehta, C.M.; Srivastava, R.; Arora, S.; Sharma, A.K. Impact assessment of silver nanoparticles on plant growth and soil bacterial diversity. 3 Biotech 2016, 6, 254. [Google Scholar] [CrossRef]
- Sillen, W.M.A.; Thijs, S.; Abbamondi, G.R.; Janssen, J.; Weyens, N.; White, J.C.; Vangronsveld, J. Effects of silver nanoparticles on soil microorganisms and maize biomass are linked in the rhizosphere. Soil Biol. Biochem. 2015, 91, 14–22. [Google Scholar] [CrossRef]
- Rastogi, G.; Sani, R.S. Molecular Techniques to Assess Microbial Community Structure, Function, and Dynamics in the Environment. In Microbes and Microbial Technology: Agricultural and Environmental Applications; Ahmad, I., Ahmad, F., Pichtel, J., Eds.; Springer: New York, NY, USA, 2011; pp. 29–57. ISBN 9781441979308. [Google Scholar]
- Lynch, M.D.J.; Bartram, A.K.; Neufeld, J.D. Targeted recovery of novel phylogenetic diversity from next-generation sequence data. ISME J. 2012, 6, 2067–2077. [Google Scholar] [CrossRef]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef] [PubMed]
- Simonin, M.; Guyonnet, J.P.; Martins, J.M.F.; Ginot, M.; Richaume, A. Influence of soil properties on the toxicity of TiO2 nanoparticles on carbon mineralization and bacterial abundance. J. Hazard. Mater. 2015, 283, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Frenk, S.; Ben-Moshe, T.; Dror, I.; Berkowitz, B.; Minz, D. Effect of metal oxide nanoparticles on microbial community structure and function in two different soil types. PLoS ONE 2013, 8, e84441. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C.; Meisel, D. Adsorption and surface-enhanced Raman of dyes on silver and gold sols. J. Phys. Chem. 1982, 86, 3391–3395. [Google Scholar] [CrossRef]
- Yadav, A.; Prasad, V.; Kathe, A.A.; Raj, S.; Yadav, D.; Sundaramoorthy, C.; Vigneshwaran, N. Functional finishing in cotton fabrics using zinc oxide nanoparticles. Bull. Mater. Sci. 2006, 29, 641–645. [Google Scholar] [CrossRef]
- Bergey, D.; Whitman, W.B.; De Vos, P.; Garrity, G.M.; Jones, D. Bergey’s Manual of Systematic Bacteriology; Springer: New York, NY, USA, 2009. [Google Scholar]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method for detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar]
- Challa, O.; Gajbhiye, K.S.; Velayatham, M. Soil Series Of Maharashtra; NBBS Publ. No 79; NBBS & LUP: Nagpur, India, 2008. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high- throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; Desantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Hristozov, D.; Malsch, I. Hazards and Risks of engineered nanoparticles for the environment and human health. Sustainability 2009, 1, 1161–1194. [Google Scholar] [CrossRef]
- Bundschuh, M.; Filser, J.; Lüderwald, S.; McKee, M.S.; Metreveli, G.; Schaumann, G.E.; Schulz, R.; Wagner, S. Nanoparticles in the environment: Where do we come from, where do we go to? Environ. Sci. Eur. 2018, 30, 6. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Khan, H.M.; Khan, A.A.; Ahmad, M.K.; Mahdi, A.A.; Pal, R.; Cameotra, S.S. Interaction of silver nanoparticles with Escherichia coli and their cell envelope biomolecules. J. Basic Microbiol. 2013, 54, 905–915. [Google Scholar] [CrossRef]
- Hsueh, Y.H.; Lin, K.S.; Ke, W.J.; Hsieh, C.T.; Chiang, C.L.; Tzou, D.Y.; Liu, S.T. The antimicrobial properties of silver nanoparticles in bacillus subtilis are mediated by released Ag+ ions. PLoS ONE 2015, 10, e144306. [Google Scholar] [CrossRef] [PubMed]
- Azam, A.; Ahmed, A.S.; Oves, M.; Khan, M.S.; Habib, S.S.; Adnan, M. Antimicrobial activity of metal oxide nanoparticles against Gram-positive and Gram-negative bacteria: A comparative study. Int. J. Nanomed. 2012, 7, 6003–6009. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.F.; Ansari, A.H.; Hameedullah, M.; Ahmad, E.; Husain, F.M.; Zia, Q.; Baig, U.; Zaheer, M.R.; Alam, M.M.; Khan, A.M.; et al. Sol-gel synthesis of thorn-like ZnO nanoparticles endorsing mechanical stirring effect and their antimicrobial activities: Potential role as nano-antibiotics. Sci. Rep. 2016, 6, 27689. [Google Scholar] [CrossRef]
- Ansari, M.A.; Khan, H.M.; Khan, A.A.; Sultan, A.; Azam, A. Synthesis and characterization of the antibacterial potential of ZnO nanoparticles against extended-spectrum β-lactamases-producing escherichia coli and klebsiella pneumoniae isolated from a tertiary care hospital of North India. Appl. Microbiol. Biotechnol. 2012, 94, 467–477. [Google Scholar] [CrossRef]
- Li, M.; Zhu, L.; Lin, D. Toxicity of ZnO nanoparticles to Escherichia Coli: Mechanism and the influence of medium components. Environ. Sci. Technol. 2011, 45, 1977–1983. [Google Scholar] [CrossRef] [PubMed]
- Suresh, A.K.; Pelletier, D.A.; Doktycz, M. Relating nanomaterial properties and microbial toxicity. Nanoscale 2013, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef]
- Yadav, A.N.; Verma, P.; Singh, B.; Chauahan, V.S.; Suman, A.; Saxena, A.K. Plant Growth Promoting Bacteria: Biodiversity and Multifunctional Attributes for Sustainable Agriculture. Adv. Biotechnol. Microbiol. 2017, 5, 1–16. [Google Scholar] [CrossRef]
- Qin, S.; Li, W.; Dastager, S.G.; Hozzein, W.N. Actinobacteria in Special and Extreme Habitats: Diversity, Function Roles and Environmental Adaptations. Front. Microbiol. 2016, 7, 1415. [Google Scholar] [CrossRef]
- Barka, A.B.; Vatsa, P.; Sanchez, L.; Gaveau-Vailant, N.; Jacquard, C.; Klenk, H.-P.; Clement, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef]
- Kumar, G.; Kanaujia, N.; Bafana, A. Functional and phylogenetic diversity of root-associated bacteria of Ajuga bracteosa in Kangra valley. Microbiol. Res. 2012, 167, 220–225. [Google Scholar] [CrossRef]
- Wang, J.; Shu, K.; Zhang, L.; SI, Y. Effects of Silver Nanoparticles on Soil Microbial Communities and Bacterial Nitrification in Suburban Vegetable Soils. Pedosphere 2017, 27, 482–490. [Google Scholar] [CrossRef]
- Ryan, R.P.; Monchy, S.; Cardinale, M.; Taghavi, S.; Crossman, L.; Avison, M.B.; Berg, G.; van der Lelie, D.; Dow, J.M. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat. Rev. Microbiol. 2009, 7, 514–525. [Google Scholar] [CrossRef]
- Preston, G.M. Plant perceptions of plant growth-promoting Pseudomonas. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Llamas, I.; del Moral, A.; Martínez-Checa, F.; Arco, Y.; Arias, S.; Quesada, E. Halomonas maura is a physiologically versatile bacterium of both ecological and biotechnological interest. Antonie van Leeuwenhoek 2006, 89, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Schimel, J.P.; Holdena, P.A. Identification of soil bacteria susceptible to TiO2 and ZnO nanoparticles. Appl. Environ. Microbiol. 2012, 78, 6749–6758. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.P.; Vorhölter, F.-J.; Potnis, N.; Jones, J.B.; Van Sluys, M.-A.; Bogdanove, A.J.; Dow, J.M. Pathogenomics of Xanthomonas: Understanding bacterium–plant interactions. Nat. Rev. Microbiol. 2011, 9, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Meli, K.; Kamika, I.; Keshri, J.; Momba, M.N.B. The impact of zinc oxide nanoparticles on the bacterial microbiome of activated sludge systems. Sci. Rep. 2016, 6, 39176. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, C. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef]
- Desai, C.; Jain, K.; Madamwar, D. Evaluation of In vitro Cr(VI) reduction potential in cytosolic extracts of three indigenous Bacillus sp. isolated from Cr(VI) polluted industrial landfill. Bioresour. Technol. 2008, 99, 6059–6069. [Google Scholar] [CrossRef]
- Mandic-Mulec, I.; Prosser, J.I. Diversity of Endospore-forming Bacteria in Soil: Characterization and Driving Mechanism. In Endospore-forming Soil Bacteria; Logan, N.A., Vos, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 27, pp. 31–59. [Google Scholar] [CrossRef]
- Vasileiadis, S.; Puglisi, E.; Trevisan, M.; Scheckel, K.G.; Langdon, K.; McLaughlin, M.J.; Lombi, E.; Donner, E. Changes in soil Bacterial communities and diversity in response to long-term silver exposure. FEMS Microbiol. Ecol. 2015, 91, fiv114. [Google Scholar] [CrossRef]
- Asadishad, B.; Chahal, S.; Akbari, A.; Cianciarelli, V.; Azodi, M.; Ghoshal, S.; Tufenkji, N. Amendment of Agricultural Soil with Metal Nanoparticles: Effects on Soil Enzyme Activity and Microbial Community Composition. Environ. Sci. Technol. 2018, 52, 1908–1918. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, M.; Jin, Y.; Fan, X.; Xu, J.; Zhu, Y.; Fu, Z.; Pan, X.; Qian, H. The Effects of Low Concentrations of Silver Nanoparticles on Wheat Growth, Seed Quality, and Soil Microbial Communities. Water. Air. Soil Pollut. 2017, 228, 348. [Google Scholar] [CrossRef]
- Samarajeewa, A.D.; Velicogna, J.R.; Princz, J.I.; Subasinghe, R.M.; Scroggins, R.P.; Beaudette, L.A. Effect of silver nano-particles on soil microbial growth, activity and community diversity in a sandy loam soil. Environ. Pollut. 2017, 220, 504–513. [Google Scholar] [CrossRef]
- Dimkpa, C.O. Can nanotechnology deliver the promised benefits without negatively impacting soil microbial life? J. Basic Microbiol. 2014, 54, 889–904. [Google Scholar] [CrossRef]
- McKee, M.S.; Filser, J. Impacts of metal-based engineered nanomaterials on soil communities. Environ. Sci. Nano 2016, 3, 506–533. [Google Scholar] [CrossRef]
- Simonin, M.; Richaume, A. Impact of engineered nanoparticles on the activity, abundance, and diversity of soil microbial communities: A review. Environ. Sci. Pollut. Res. 2015, 22, 13710–13723. [Google Scholar] [CrossRef]
- Grün, A.L.; Emmerling, C. Long-term effects of environmentally relevant concentrations of silver nanoparticles on major soil bacterial phyla of a loamy soil. Environ. Sci. Eur. 2018, 30, 31. [Google Scholar] [CrossRef]
- Peyrot, C.; Wilkinson, K.J.; Desrosiers, M.; Sauvé, S. Effects of silver nanoparticles on soil enzyme activities with and without added organic matter. Environ. Toxicol. Chem. 2014, 33, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Santimano, M.C.; Kowshik, M. Altered growth and enzyme expression profile of ZnO nanoparticles exposed non-target environmentally beneficial bacteria. Environ. Monit. Assess. 2013, 185, 7205–7214. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, J.; Lee, I. Effects of Zn and ZnO nanoparticles and Zn2+ on soil enzyme activity and bioaccumulation of zinc in Cucumis sativus. Chem. Ecol. 2011, 21, 49–55. [Google Scholar] [CrossRef]
- García-Gómez, C.; Fernández, D.; García, S.; Obrador, A.F.; Letón, M.; Babín, M. Soil pH effects on the toxicity of zinc oxide nanoparticles to soil microbial community. Environ. Sci. Pollut. Res. 2018, 25, 28140–28152. [Google Scholar] [CrossRef] [PubMed]
- Thomson, B.C.; Ostle, N.J.; McNamara, N.P.; Whiteley, A.S.; Griffiths, R.I. Effects of sieving, drying and rewetting upon soil bacterial community structure and respiration rates. J. Microbiol. Methods 2010, 83, 69–73. [Google Scholar] [CrossRef]
- Blaud, A.; Menon, M.; van der Zaan, B.; Lair, G.J.; Banwart, S.A. Effects of Dry and Wet Sieving of Soil on Identification and Interpretation of Microbial Community Composition. In Advances in Agronomy; Banwart, S.A., Sparks, D.L., Eds.; Burlington Academic Press: Cambridge, MA, USA, 2017; Volume 142, ISBN 9780128122228. [Google Scholar]
- Datta, R.; Vranová, V.; Pavelka, M.; Rejšek, K.; Formánek, P. Effect of soil sieving on respiration induced by low-molecular-weight substrates. Int. Agrophys. 2014, 28, 119–124. [Google Scholar] [CrossRef]
- Zielińska, S.; Radkowski, P.; Blendowska, A.; Ludwig-Gałęzowska, A.; Łoś, J.M.; Łoś, M. The choice of the DNA extraction method may influence the outcome of the soil microbial community structure analysis. Microbiologyopen 2017, 6, e00453. [Google Scholar] [CrossRef] [PubMed]
- Kebschull, J.M.; Zador, A.M. Sources of PCR-induced distortions in high-throughput sequencing data sets. Nucleic Acids Res. 2015, 43, e143. [Google Scholar] [CrossRef]
- Pfeiffer, F.; Gröber, C.; Blank, M.; Händler, K.; Beyer, M.; Schultze, J.L.; Mayer, G. Systematic evaluation of error rates and causes in short samples in next-generation sequencing. Sci. Rep. 2018, 8, 10950. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.G.; Russ, C.; Costello, M.; Hollinger, A.; Lennon, N.J.; Hegarty, R.; Nusbaum, C.; Jaffe, D.B. Characterizing and measuring bias in sequence data. Genome Biol. 2013, 14, R51. [Google Scholar] [CrossRef]
- Allali, I.; Arnold, J.W.; Roach, J.; Cadenas, M.B.; Butz, N.; Hassan, H.M.; Koci, M.; Ballou, A.; Mendoza, M.; Ali, R.; et al. A comparison of sequencing platforms and bioinformatics pipelines for compositional analysis of the gut microbiome. BMC Microbiol. 2017, 17, 1–16. [Google Scholar] [CrossRef]
- Siegwald, L.; Touzet, H.; Lemoine, Y.; Hot, D.; Audebert, C.; Caboche, S. Assessment of common and emerging bioinformatics pipelines for targeted metagenomics. PLoS ONE 2017, 12, e0169563. [Google Scholar] [CrossRef]
Figure 1.
Effect of silver and zinc oxide nanoparticles on growth of pure cultures. The graphs show growth curves of isolates in the presence and absence of nanoparticles. The isolates were inoculated in broth and optical density (OD) recorded every hour for 24–48 h. (a) Effect of silver nanoparticles on growth of Pseudomonas aeruginosa MTCC 7763; (b) Effect of zinc oxide nanoparticles on growth of Serratia marcescens MTCC 7642; (c) Effect of zinc oxide nanoparticles on Azotobacter chroococcum clsxc_A_FLNF. Each point represents mean of triplicate values of absorbance adjusted against data from sterile controls. All growth curve experiments were repeated at least thrice. MIC: Minimum Inhibitory Concentration.
Figure 1.
Effect of silver and zinc oxide nanoparticles on growth of pure cultures. The graphs show growth curves of isolates in the presence and absence of nanoparticles. The isolates were inoculated in broth and optical density (OD) recorded every hour for 24–48 h. (a) Effect of silver nanoparticles on growth of Pseudomonas aeruginosa MTCC 7763; (b) Effect of zinc oxide nanoparticles on growth of Serratia marcescens MTCC 7642; (c) Effect of zinc oxide nanoparticles on Azotobacter chroococcum clsxc_A_FLNF. Each point represents mean of triplicate values of absorbance adjusted against data from sterile controls. All growth curve experiments were repeated at least thrice. MIC: Minimum Inhibitory Concentration.

Figure 2.
Rarefaction curves of alpha diversity indices. (a) Observed species curves and (b) Chao 1 curves, obtained for the samples. Alpha diversity was computed using observed species and Chao 1 metrics, respectively, with rarefied OTU table size of 100. 1C—control (untreated soil); 2A—Ag nanoparticle-treated soil; and 3Z—ZnO nanoparticle-treated soil.
Figure 2.
Rarefaction curves of alpha diversity indices. (a) Observed species curves and (b) Chao 1 curves, obtained for the samples. Alpha diversity was computed using observed species and Chao 1 metrics, respectively, with rarefied OTU table size of 100. 1C—control (untreated soil); 2A—Ag nanoparticle-treated soil; and 3Z—ZnO nanoparticle-treated soil.

Figure 3.
Relative abundance of bacterial phyla. The figure and the accompanying table indicate relative abundance (%) of bacterial phyla in treated and untreated soil samples. 1C—control (untreated soil); 2A—silver nanoparticle-treated soil; and 3Z—zinc oxide nanoparticle-treated soil.
Figure 3.
Relative abundance of bacterial phyla. The figure and the accompanying table indicate relative abundance (%) of bacterial phyla in treated and untreated soil samples. 1C—control (untreated soil); 2A—silver nanoparticle-treated soil; and 3Z—zinc oxide nanoparticle-treated soil.

Figure 4.
Relative abundance of bacterial orders of dominant phyla in treated and untreated soil samples. The figure and accompanying table indicate relative abundance (%) of bacterial orders belonging to only the three dominant phyla—Actinobacteria, Firmicutes, and Proteobacteria. The table also shows the phyla to which these orders belong. Since only the most dominant phyla have been presented at the order level, the total of percentages do not add up to 100. The orders have been chosen based on their relative abundance within the phylum and the most prominent ones have been tabulated. 1C—control (untreated soil); 2A—silver nanoparticle-treated soil; and 3Z—zinc oxide nanoparticle-treated soil.
Figure 4.
Relative abundance of bacterial orders of dominant phyla in treated and untreated soil samples. The figure and accompanying table indicate relative abundance (%) of bacterial orders belonging to only the three dominant phyla—Actinobacteria, Firmicutes, and Proteobacteria. The table also shows the phyla to which these orders belong. Since only the most dominant phyla have been presented at the order level, the total of percentages do not add up to 100. The orders have been chosen based on their relative abundance within the phylum and the most prominent ones have been tabulated. 1C—control (untreated soil); 2A—silver nanoparticle-treated soil; and 3Z—zinc oxide nanoparticle-treated soil.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial isolates and the MIC/MBC of Ag and ZnO nanoparticles for the isolates. The MIC and MBC values are average of five replicates, and have been rounded to the nearest significant figure.
Table 1.
Bacterial isolates and the MIC/MBC of Ag and ZnO nanoparticles for the isolates. The MIC and MBC values are average of five replicates, and have been rounded to the nearest significant figure.
| Cultures | Gram Nature | Characteristics | Ag Nanoparticles | ZnO Nanoparticles | ||
|---|---|---|---|---|---|---|
| MIC µg/mL | MBC µg/mL | MIC µg/mL | MBC µg/mL | |||
| Nitrogen Fixers | ||||||
| Rhizobium leguminosarum MTCC 10096 | Negative | Known symbiotic nitrogen fixer, biofilm former | 2 | 2 | 104 | 2000 |
| Sinorhizobium meliloti clsxc_S_SNF (MH884597) | Negative | Known symbiotic nitrogen fixer | 4 | 4 | 144 | 600 |
| Azotobacter chroococcum MTCC 3853 | Negative | Known free living nitrogen fixer, biofilm former | 4.9 | 4.9 | 2000 | >3000 |
| Azotobacter chroococcum clsxc_A_FLNF (MH884596) | Negative | Known free living nitrogen fixer | 3.9 | 3.9 | 197 | 400 |
| Phosphate solubilizers | ||||||
| Arthrobacter sp. MTCC 8160 | Positive | Phosphate solubilizer | 3 | 3 | 99 | 500 |
| Bacillus sp. clsxc_ NPS (MH884601) | Positive | Phosphate solubilizer, air–liquid interface biofilm former, amylase producer | 8 | 8 | 100 | 147 |
| Serratia marcescens MTCC 7642 | Negative | Phosphate solubilizer, adherent biofilm former | 20 | 22 | 1480 | 2000 |
| Pantoea dispersa clsxc_PSD (MH884599) | Negative | Phosphate solubilizer | 3 | 3 | 680 | 1000 |
| Biofilm formers | ||||||
| Bacillus subtilis MTCC 441 | Positive | Air–liquid interface biofilm former, amylase producer | 14.8 | 15 | 79 | 460 |
| Bacillus sp. clsxc_TYA (MH884600) | Positive | Air–liquid interface biofilm former, amylase producer | 17.5 | 18 | 104 | 510 |
| Pseudomonas aeruginosa MTCC 7763 | Negative | Adherent biofilm former, Phosphate solubilizer | 20.6 | 22 | >3000 | >3000 |
| Klebsiella pneumoniae clsxc_AZ2 (MH884598) | Negative | Adherent biofilm former, free living nitrogen fixer, phosphate solubilizer | 7.5 | 9 | 840 | 1600 |
Table 2.
Alpha diversity metrics for the samples. 1C—control (untreated soil); 2A—Ag nanoparticle-treated soil; and 3Z—ZnO nanoparticle-treated soil.
Table 2.
Alpha diversity metrics for the samples. 1C—control (untreated soil); 2A—Ag nanoparticle-treated soil; and 3Z—ZnO nanoparticle-treated soil.
| Sample | Observed OTUs | Chao 1 Richness Estimate | PD_Whole Tree a | Shannon’s Index b |
|---|---|---|---|---|
| 1C | 5286 | 6153 | 133 | 9.271 |
| 2A | 6077 | 6667 | 149 | 8.280 |
| 3Z | 5448 | 6286 | 136 | 9.444 |
a,b (Supplementary Materials, Figure S4). (OTU: Operational Taxonomic Unit; Chao 1: Abundance-based estimator of species richness; PD_Whole Tree: a measure of phylogenetic diversity).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chavan, S.; Nadanathangam, V. Effects of Nanoparticles on Plant Growth-Promoting Bacteria in Indian Agricultural Soil. Agronomy 2019, 9, 140. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030140
AMA Style
Chavan S, Nadanathangam V. Effects of Nanoparticles on Plant Growth-Promoting Bacteria in Indian Agricultural Soil. Agronomy. 2019; 9(3):140. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030140
Chicago/Turabian StyleChavan, Sangeeta, and Vigneshwaran Nadanathangam. 2019. "Effects of Nanoparticles on Plant Growth-Promoting Bacteria in Indian Agricultural Soil" Agronomy 9, no. 3: 140. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030140
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.