Isolation and Characterization of the High Silicate and Phosphate Solubilizing Novel Strain Enterobacter ludwigii GAK2 that Promotes Growth in Rice Plants

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Screening of Bacteria
2.2. Evaluation of the Ability to Solubilize Insoluble Silicate and Phosphate
2.3. Preliminary Bioanalysis Test
2.3.1. Detection of Indole-3-Acetic Acid (IAA)
2.3.2. Production of Ammonia
2.3.3. Growth Promotion Test on Rice
2.4. Selection, Identification, and Phylogenetic Analysis of Microorganisms
2.5. Biochemical Analysis
2.5.1. Microorganism Culture
2.5.2. Quantification of Organic Acids
2.5.3. Analysis of IAA Production in E. ludwigii GAK2 Culture
2.5.4. Extraction and Quantification of Gibberellin in GAK2 Culture
2.6. Evaluation of the Effect of E. ludwigii GAK2 on Plant Growth
2.6.1. Pot Experiment
2.6.2. Analysis of Si Content of the Plant
2.7. Determination of Soil pH and Electrical Conductivity (EC)
2.8. Statistical Procedures for Data Analysis
3. Results
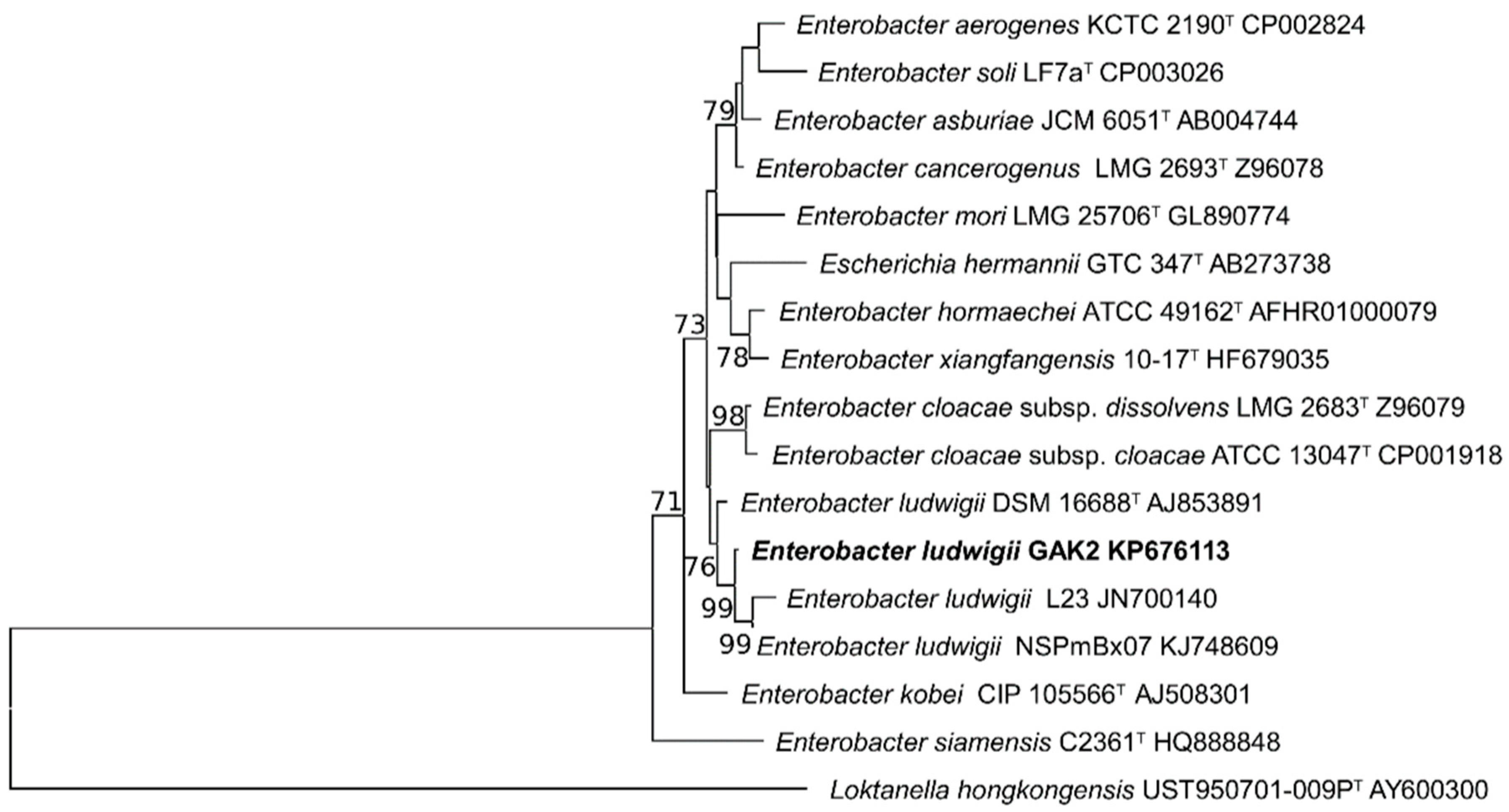
3.1. Isolation Selection and Identification of Silicate Solubilizing Bacteria
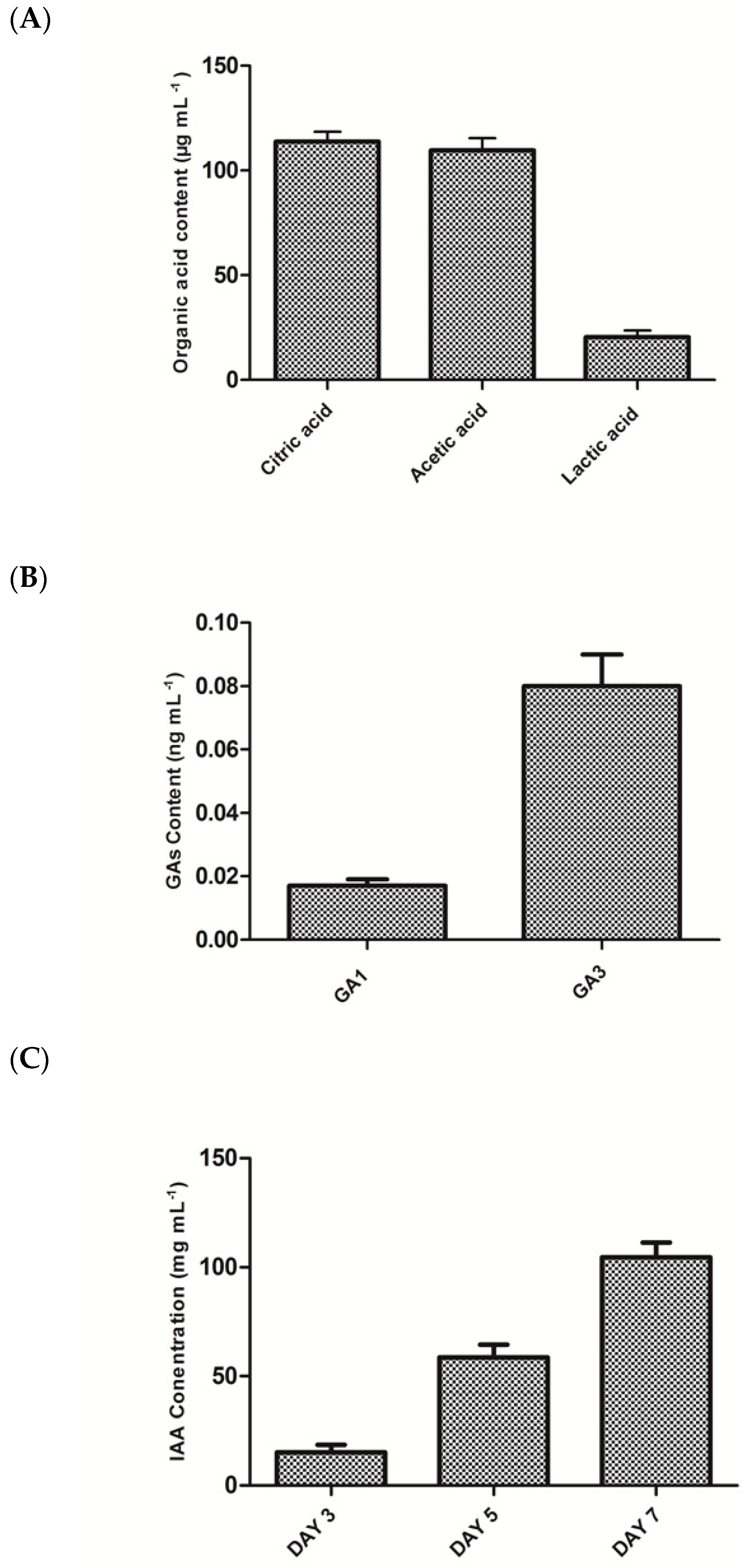
3.2. Biochemical Analysis of E. ludwigii GAK2 Culture
3.3. Plant Growth Promoting Characteristics
3.4. Silicon Content in Plant
3.5. Measurement of Electrical Conductivity and pH
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Zhao, F.J.; Moore, K.L.; Lombi, E.; Zhu, Y.G. Imaging element distribution and speciation in plant cells. Trends Plant Sci. 2014, 19, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, E.D.; Benfey, P.N. Regulation of plant root system architecture: Implications for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [Google Scholar] [CrossRef]
- Farooq, M.A.; Dietz, K.J. Silicon as Versatile Player in Plant and Human Biology: Overlooked and Poorly Understood. Front. Plant Sci. 2015, 6, 994. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef]
- Darzi-Naftchali, A.; Shahnazari, A.; Karandish, F. Phosphorus balance and risk assessment in the rice–canola cropping system under different drainage strategies. Arch. Agron. Soil Sci. 2016, 62, 1554–1566. [Google Scholar] [CrossRef]
- Mehmood, T.; Chaudhry, M.; Tufail, M.; Irfan, N. Heavy metal pollution from phosphate rock used for the production of fertilizer in Pakistan. Microchem. J. 2009, 91, 94–99. [Google Scholar]
- Uroz, S.; Calvaruso, C.; Turpault, M.P.; Frey-Klett, P. Mineral weathering by bacteria: Ecology, actors and mechanisms. Trends Microbiol. 2009, 17, 378–387. [Google Scholar] [CrossRef]
- Rogers, J.R.; Bennett, P.C. Mineral stimulation of subsurface microorganisms: Release of limiting nutrients from silicates. Chem. Geol. 2004, 203, 91–108. [Google Scholar] [CrossRef]
- Ahemad, M. Phosphate-solubilizing bacteria-assisted phytoremediation of metalliferous soils: A review. 3 Biotech 2015, 5, 111–121. [Google Scholar] [CrossRef]
- Taktek, S.; Trépanier, M.; Servin, P.M.; St-Arnaud, M.; Piché, Y.; Fortin, J.A.; Antoun, H. Trapping of phosphate solubilizing bacteria on hyphae of the arbuscular mycorrhizal fungus Rhizophagus irregularis DAOM 197198. Soil Biol. Biochem. 2015, 90, 1–9. [Google Scholar] [CrossRef]
- Kang, S.M.; Waqas, M.; Shahzad, R.; You, Y.H.; Asaf, S.; Khan, M.A.; Lee, K.E.; Joo, G.J.; Kim, S.J.; Lee, I.J. Isolation and characterization of a novel silicate-solubilizing bacterial strain Burkholderia eburnea CS4-2 that promotes growth of japonica rice (Oryza sativa L. cv. Dongjin). Soil Sci. Plant Nutr. 2017, 63, 233–241. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Sitepu, I.R.; Tang, S.Y.; Hashidoko, Y. Salkowski’s reagent test as a primary screening index for functionalities of rhizobacteria isolated from wild dipterocarp saplings growing naturally on medium-strongly acidic tropical peat soil. Biosci. Biotechnol. Biochem. 2010, 74, 2202–2208. [Google Scholar] [CrossRef]
- Kang, S.M.; Radhakrishnan, R.; You, Y.H.; Khan, A.L.; Park, J.M.; Lee, S.M.; Lee, I.J. Cucumber performance is improved by inoculation with plant growth-promoting microorganisms. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2015, 65, 36–44. [Google Scholar] [CrossRef]
- Lee, K.E.; Radhakrishnan, R.; Kang, S.M.; You, Y.H.; Joo, G.J.; Lee, I.J.; Ko, J.H.; Kim, J.H. Enterococcus faecium LKE12 Cell-Free Extract Accelerates Host Plant Growth via Gibberellin and Indole-3-Acetic Acid Secretion. J. Microbiol. Biotechnol. 2015, 25, 1467–1475. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis; Prentice-Hall Inc.: Englewood Cliffs, NJ, USA, 1958. [Google Scholar]
- Kalra, Y.P. Determination of pH of soils by different methods: Collaborative study. J. AOAC Int. 1995, 78, 310–324. [Google Scholar]
- Imtiaz, M.; Rizwan, M.S.; Mushtaq, M.A.; Ashraf, M.; Shahzad, S.M.; Yousaf, B.; Saeed, D.A.; Rizwan, M.; Nawaz, M.A.; Mehmood, S.; et al. Silicon occurrence, uptake, transport and mechanisms of heavy metals, minerals and salinity enhanced tolerance in plants with future prospects: A review. J. Environ. Manag. 2016, 183, 521–529. [Google Scholar] [CrossRef]
- Anand, K.; Kumari, B.; Mallick, M. Phosphate solubilizing microbes: An effective and alternative approach as biofertilizers. J. Pharm. Pharm Sci. 2016, 8, 37–40. [Google Scholar]
- Khalifa, A.Y.; Alsyeeh, A.-M.; Almalki, M.A.; Saleh, F.A. Characterization of the plant growth promoting bacterium, Enterobacter cloacae MSR1, isolated from roots of non-nodulating Medicago sativa. Saudi J. Biol. Sci. 2016, 23, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, P.; Ruppel, S.; Remus, R. Enterobacter radicincitans sp. nov., a plant growth promoting species of the family Enterobacteriaceae. Syst. Appl. Microbiol. 2005, 28, 213–221. [Google Scholar]
- Jha, C.K.; Saraf, M. Plant growth promoting rhizobacteria (PGPR): A review. J. Agric. Res. Dev. 2015, 5, 108–119. [Google Scholar]
- Park, Y.-G.; Mun, B.-G.; Kang, S.-M.; Hussain, A.; Shahzad, R.; Seo, C.-W.; Kim, A.-Y.; Lee, S.-U.; Oh, K.Y.; Lee, D.Y. Bacillus aryabhattai SRB02 tolerates oxidative and nitrosative stress and promotes the growth of soybean by modulating the production of phytohormones. PLoS ONE 2017, 12, e0173203. [Google Scholar]
- Lazo, D.E.; Dyer, L.G.; Alorro, R.D. Silicate, phosphate and carbonate mineral dissolution behaviour in the presence of organic acids: A review. Miner. Eng. 2017, 100, 115–123. [Google Scholar] [CrossRef]
- Patra, P.K.; Neue, H.U. Dynamics of water soluble silica and silicon nutrition of rice in relation to changes in iron and phosphorus in soil solution due to soil drying and reflooding. Arch. Agron. Soil Sci. 2010, 56, 605–622. [Google Scholar] [CrossRef]
- Jones, D.L.; Dennis, P.G.; Owen, A.G.; van Hees, P.A.W. Organic acid behavior in soils—misconceptions and knowledge gaps. Plant Soil 2003, 248, 31–41. [Google Scholar] [CrossRef]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 2009, 9, 174. [Google Scholar] [CrossRef]
- Liu, W.; Xu, X.; Wu, X.; Yang, Q.; Luo, Y.; Christie, P. Decomposition of silicate minerals by Bacillus mucilaginosus in liquid culture. Environ. Geochem. Health 2006, 28, 133–140. [Google Scholar] [CrossRef]
- Bennett, P.C.; Rogers, J.R.; Choi, W.J.; Hiebert, F.K. Silicates, silicate weathering, and microbial ecology. Geomicrobiol. J. 2001, 18, 3–19. [Google Scholar] [CrossRef]
- Shoebitz, M.; Ribaudo, C.M.; Pardo, M.A.; Cantore, M.L.; Ciampi, L.; Curá, J.A. Plant growth promoting properties of a strain of Enterobacter ludwigii isolated from Lolium perenne rhizosphere. Soil Biol. Biochem. 2009, 41, 1768–1774. [Google Scholar] [CrossRef]
- Mathesius, U. Goldacre paper: Auxin: At the root of nodule development? Funct. Plant Biol. 2008, 35, 651–668. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedden, P.; Sponsel, V. A Century of Gibberellin Research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottini, R.; Cassan, F.; Piccoli, P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase. Appl. Microbiol. Biotechnol. 2004, 65, 497–503. [Google Scholar] [CrossRef]
- Kang, S.M.; Khan, A.L.; Waqas, M.; You, Y.H.; Hamayun, M.; Joo, G.J.; Shahzad, R.; Choi, K.S.; Lee, I.J. Gibberellin-producing Serratia nematodiphila PEJ1011 ameliorates low temperature stress in Capsicum annuum L. Eur. J. Soil Biol. 2015, 68, 85–93. [Google Scholar] [CrossRef]
- Ullah, I.; Khan, A.R.; Jung, B.K.; Khan, A.L.; Lee, I.J.; Shin, J.H. Gibberellins synthesized by the entomopathogenic bacterium, photorhabdus temperata M1021 as one of the factors of rice plant growth promotion. J. Plant Interact. 2014, 9, 775–782. [Google Scholar] [CrossRef]
- Kang, S.M.; Khan, A.L.; Hamayun, M.; Hussain, J.; Joo, G.J.; You, Y.H.; Kim, J.G.; Lee, I.J. Gibberellin-producing Promicromonospora sp. SE188 improves Solanum lycopersicum plant growth and influences endogenous plant hormones. J. Microbiol. 2012, 50, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Joo, G.J.; Kim, Y.M.; Kim, J.T.; Rhee, I.K.; Kim, J.H.; Lee, I.J. Gibberellins-producing rhizobacteria increase endogenous gibberellins content and promote growth of red peppers. J. Microbiol. 2005, 43, 510–515. [Google Scholar]
- Xiao, L.; Lian, B.; Hao, J.; Liu, C.; Wang, S. Effect of carbonic anhydrase on silicate weathering and carbonate formation at present day CO2 concentrations compared to primordial values. Sci. Rep. 2015, 5, 7733. [Google Scholar] [CrossRef]
- Javaheri, N.; Dries, R.; Burson, A.; Stal, L.J.; Sloot, P.M.A.; Kaandorp, J.A. Temperature affects the silicate morphology in a diatom. Sci. Rep. 2015, 5, 11652. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Root Length (cm) | Shoot Length (cm) | Total Fresh Weight (cm) | |
|---|---|---|---|
| Control | 3.63 ± 0.4 b | 4.63 ± 0.50 b | 0.59 ± 0.16 b |
| GAK2 | 5.86 ± 0.4 a | 6.36 ± 0.56 a | 1.01 ± 0.21 a |
| Treatment | F.RW (g plant−1) | F.SW (g plant−1) | RL (cm) | SL (cm) | Chl (SPAD) |
|---|---|---|---|---|---|
| Control | 0.47 ± 0.04 c | 2.20 ± 0.28 c | 8.85 ± 0.21 c | 46.50 ± 0.7 b | 44.05 ± 0.77 ab |
| IS | 0.66 ± 0.18 bc | 2.40 ± 0.28 bc | 9.95 ± 0.35 bc | 45.8 ± 0.21 b | 41.65 ± 5.58 b |
| IP | 0.84 ± 0.11 bc | 2.25 ± 0.21 c | 10 ± 0.28 abc | 46.10 ± 2.9 b | 44.65 ± 1.62 ab |
| GAK2 | 0.88 ± 0.34 bc | 3.07 ± 0.38 ab | 10.75 ± 0.35 ab | 52.75 ± 1.06 a | 42.72 ± 0.73 ab |
| GAK2 + IS | 1.43 ± 0.09 a | 3.25 ± 0.25 a | 11 ± 0.35 a | 53.25 ± 0.35 a | 47.9 ± 1.55 a |
| GAK2 + IP | 0.99 ± 0.13 b | 3.21 ± 0.35 a | 11.25 ± 1.06 a | 52.76 ± 1.07 a | 43.65 ± 0.63 ab |
| Treatment | EC (ds m−1) | pH | P (mg Kg−1 D.W.) | Si (mg Kg−1 D.W.) |
|---|---|---|---|---|
| Control | 0.72 ± 0.02 c | 5.08 ± 0.14 de | 4063 ± 4.33 c | 156.28 ± 8.06 c |
| IS | 0.83 ± 0.04 c | 6.55 ± 0.07 a | 4030 ± 189.54 c | 128.08 ± 7.37 d |
| IP | 1.05 ± 0.07 b | 5.45 ± 0.07 c | 4931 ± 57.34 b | 115.68 ± 6.56 d |
| GAK2 | 1.07 ± 0.07 b | 4.96 ± 0.05 e | 4133 ± 211.26 c | 194.06 ± 4.67 b |
| GAK2 + IS | 1.27 ± 0.07 a | 6.16 ± 0.07 b | 3884.9 ± 67.08 c | 232.41 ± 11.18 a |
| GAK2 + IP | 1.11 ± 0.02 b | 5.31 ± 0.15 cd | 5549.7 ± 263.46 a | 127.46 ± 8.62 d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.-E.; Adhikari, A.; Kang, S.-M.; You, Y.-H.; Joo, G.-J.; Kim, J.-H.; Kim, S.-J.; Lee, I.-J. Isolation and Characterization of the High Silicate and Phosphate Solubilizing Novel Strain Enterobacter ludwigii GAK2 that Promotes Growth in Rice Plants. Agronomy 2019, 9, 144. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030144
Lee K-E, Adhikari A, Kang S-M, You Y-H, Joo G-J, Kim J-H, Kim S-J, Lee I-J. Isolation and Characterization of the High Silicate and Phosphate Solubilizing Novel Strain Enterobacter ludwigii GAK2 that Promotes Growth in Rice Plants. Agronomy. 2019; 9(3):144. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030144
Chicago/Turabian StyleLee, Ko-Eun, Arjun Adhikari, Sang-Mo Kang, Young-Hyun You, Gil-Jae Joo, Jin-Ho Kim, Sang-Jun Kim, and In-Jung Lee. 2019. "Isolation and Characterization of the High Silicate and Phosphate Solubilizing Novel Strain Enterobacter ludwigii GAK2 that Promotes Growth in Rice Plants" Agronomy 9, no. 3: 144. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030144