Phenolic Composition and Antioxidant Activities of Soybean (Glycine max (L.) Merr.) Plant during Growth Cycle

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material and Growth Conditions
2.3. Extraction
2.4. Determination of Total Phenolic Content
2.5. HPLC Analysis
2.6. Trolox Equivalent Antioxidant Capacity
2.7. Ferric-Reducing Antioxidant Power
2.8. Photochemiluminescence Assay
2.9. Fe2+ Chelation Ability
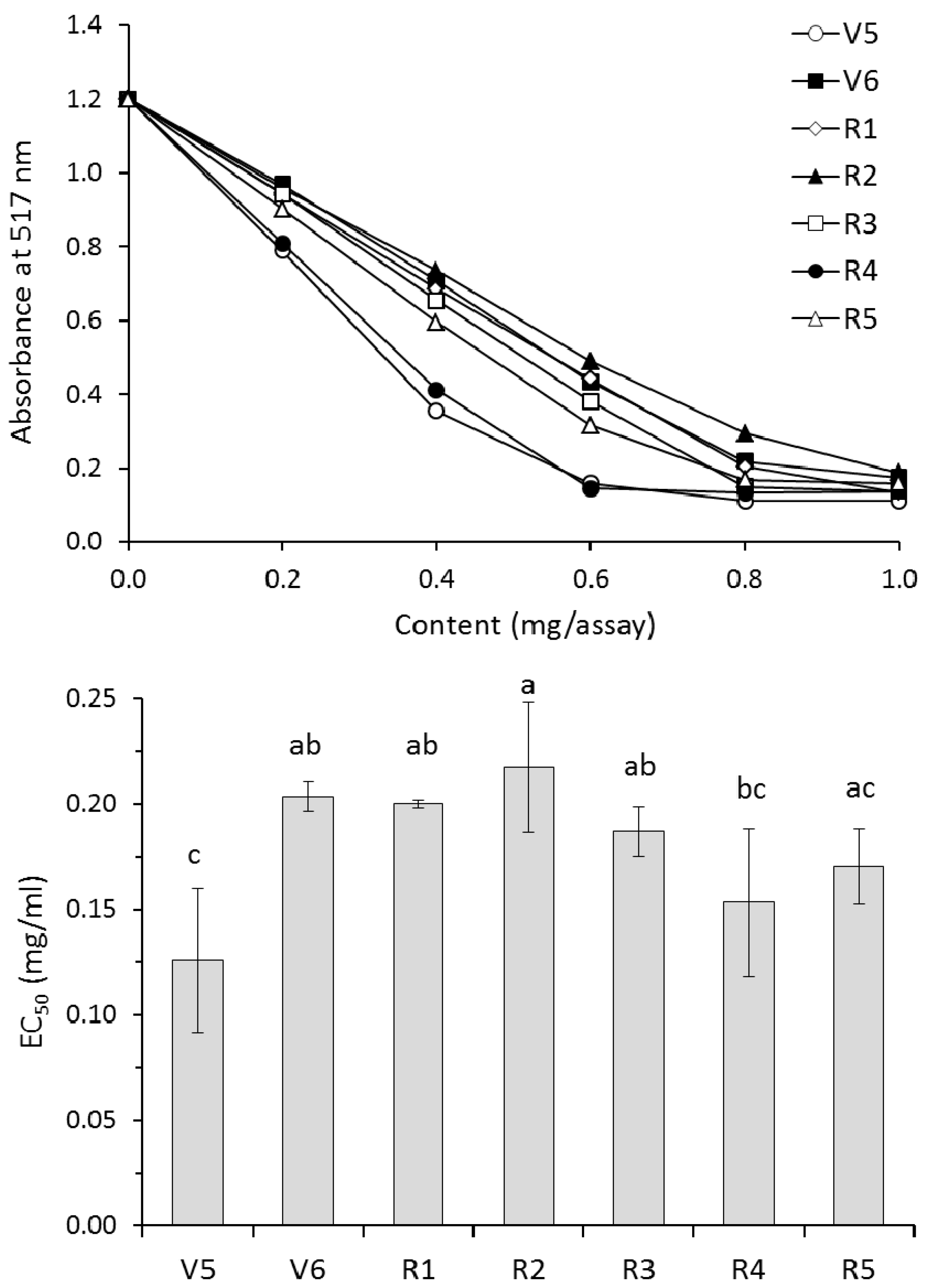
2.10. Scavenging of the DPPH Radical
2.11. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Peiretti, P.G.; Meineri, G.; Longato, E.; Tassone, S. Nutritive value and fatty acid content of soybean plant [Glycine max (L.) Merr.] during its growth cycle. Ital. J. Anim. Sci. 2018, 17, 347–352. [Google Scholar] [CrossRef]
- Seiter, S.; Altemose, C.E.; Davis, M.H. Forage soybean yield and quality responses to plant density and row distance. Agron. J. 2004, 96, 966–970. [Google Scholar] [CrossRef]
- Chang, S.R.; Lu, C.H.; Lur, H.S.; Hsu, F.H. Forage yield, chemical contents, and silage quality of manure soybean. Agron. J. 2012, 104, 130–136. [Google Scholar] [CrossRef]
- Touno, E.; Kaneko, M.; Uozumi, S.; Kawamoto, H.; Deguchi, S. Evaluation of feeding value of forage soybean silage as a substitute for wheat bran in sheep. Anim. Sci. J. 2014, 85, 46–52. [Google Scholar] [CrossRef]
- Đurović, S.; Nikolić, B.; Luković, N.; Jovanović, J.; Stefanović, A.; Šekuljica, N.; Mijin, D.; Knežević-Jugović, Z. The impact of high-power ultrasound and microwave on the phenolic acid profile and antioxidant activity of the extract from yellow soybean seeds. Ind. Crops Prod. 2018, 122, 223–231. [Google Scholar] [CrossRef]
- Lee, C.H.; Yang, L.; Xu, J.Z.; Yeung, S.Y.V.; Huang, Y.; Chen, Z.Y. Relative antioxidant activity of soybean isoflavones and their glycosides. Food Chem. 2005, 90, 735–741. [Google Scholar] [CrossRef]
- Mujić, I.; Šertović, E.; Jokić, S.; Sarić, Z.; Alibabić, V.; Vidović, S.; Živković, J. Isoflavone content and antioxidant properties of soybean seeds. Croat. J. Food Sci. Technol. 2011, 3, 16–20. [Google Scholar]
- Takahashi, R.; Ohmori, R.; Kiyose, C.; Momiyama, Y.; Ohsuzu, F.; Kondo, K. Antioxidant activities of black and yellow soybeans against low density lipoprotein oxidation. J. Agric. Food Chem. 2005, 53, 4578–4582. [Google Scholar] [CrossRef]
- Takahata, Y.; Ohnishi-Kameyama, M.; Furuta, S.; Takahashi, M.; Suda, I. Highly polymerized procyanidins in brown soybean seed coat with a high radical-scavenging activity. J. Agric. Food Chem. 2001, 49, 5843–5847. [Google Scholar] [CrossRef]
- Xu, B.; Chang, S.K. Characterization of phenolic substances and antioxidant properties of food soybeans grown in the North Dakota−Minnesota region. J. Agric. Food Chem. 2008, 56, 9102–9113. [Google Scholar] [CrossRef]
- Zhang, R.F.; Zhang, F.X.; Zhang, M.W.; Wei, Z.C.; Yang, C.Y.; Zhang, Y.; Tang, X.J.; Deng, Y.Y.; Chi, J.W. Phenolic composition and antioxidant activity in seed coats of 60 Chinese black soybean (Glycine max L. Merr.) varieties. J. Agric. Food Chem. 2011, 59, 5935–5944. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, S.S.; Jung, P.J.; Lee, M.H.I. Intake of fruits, vegetables, and soy foods in relation to breast cancer risk in Korean women: A case-control study. Nutr. Cancer 2007, 57, 20–27. [Google Scholar] [CrossRef]
- Inagaki, S.; Morimura, S.; Shigematsu, T.; Kida, K.; Akutagawa, H. Apoptosis induction by vinegar produced from boiled extract of black soybeans in human monoblastic leukemia U937 cells: Difference in sensitivity to cell toxicity compared to normal lymphocytes. Food Sci. Technol. Res. 2005, 11, 311–317. [Google Scholar] [CrossRef]
- Shinomiya, K.; Tokunaga, S.; Shigemoto, Y.; Kamei, C. Effects of seed coat extract of black soybeans on radial maze performance in rats. Clin. Exp. Pharmacol. Physiol. 2005, 32, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Giri, G.F.; Viarengo, G.; Furlán, R.L.; Suárez, A.G.; Véscovi, E.G.; Spanevello, R.A. Soybean hulls, an alternative source of bioactive compounds: Combining pyrolysis with bioguided fractionation. Ind. Crops Prod. 2017, 105, 113–123. [Google Scholar] [CrossRef]
- Astadi, I.R.; Astuti, M.A.; Santoso, U.; Nugraheni, P.S. In vitro antioxidant activity of anthocyanins of black soybean seed coat in human low density lipoprotein (LDL). Food Chem. 2009, 112, 659–663. [Google Scholar] [CrossRef]
- Wang, H.; Murphy, P.A. Isoflavone content in commercial soybean foods. J. Agric. Food Chem. 1994, 42, 1666–1673. [Google Scholar] [CrossRef]
- Coward, L.; Barnes, N.C.; Setchell, K.D.; Barnes, S. Genistein, daidzein, and their beta-glycoside conjugates: Antitumor isoflavones in soybean foods from American and Asian diets. J. Agric. Food Chem. 1993, 41, 1961–1967. [Google Scholar] [CrossRef]
- Ho, H.M.; Chen, R.Y.; Leung, L.K.; Chan, F.L.; Huang, Y.; Chen, Z.Y. Difference in flavonoid and isoflavone profile between soybean and soy leaf. Biomed. Pharmacother. 2002, 56, 289–295. [Google Scholar] [CrossRef]
- Bennett, J.O.; Yu, O.; Heatherly, L.G.; Krishnan, H.B. Accumulation of genistein and daidzein, soybean isoflavones implicated in promoting human health, is significantly elevated by irrigation. J. Agric. Food Chem. 2004, 52, 7574–7579. [Google Scholar] [CrossRef] [PubMed]
- Hoeck, J.; Fehr, W.; Murphy, P.; Welke, G. Influence of genotype and environment on isoflavone contents of soybean. Crop Sci. 2000, 40, 48–51. [Google Scholar] [CrossRef]
- Lee, S.; Ahn, J.; Kim, S.; Kim, J.; Han, S.; Jung, M.; Chung, I. Variation in isoflavone of soybean cultivars with location and storage duration. J. Agric. Food Chem. 2003, 51, 3382–3389. [Google Scholar] [CrossRef]
- Chennupati, P.; Seguin, P.; Liu, W. Effects of high temperature stress at different development stages on soybean isoflavone and tocopherol concentrations. J. Agric. Food Chem. 2011, 59, 13081–13088. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, C.; Shimada, S.; Igita, K.; Kudou, S.; Kokubun, M.; Okubo, K.; Kitamura, K. Factors affecting isoflavone content in soybean seeds: Changes in isoflavones, saponins, and composition of fatty acids at different temperatures during seed development. J. Agric. Food Chem. 1995, 43, 1184–1192. [Google Scholar] [CrossRef]
- Kumar, V.; Rani, A.; Dixit, A.K.; Bhatnagar, D.; Chauhan, G.S. Relative changes in tocopherols, isoflavones, total phenolic content, and antioxidative activity in soybean seeds at different reproductive stages. J. Agric. Food Chem. 2009, 57, 2705–2710. [Google Scholar] [CrossRef]
- Kumar, V.; Rani, A.; Dixit, A.K.; Pratap, D.; Bhatnagar, D. A comparative assessment of total phenolic content, ferric reducing-anti-oxidative power, free radical-scavenging activity, vitamin C and isoflavones content in soybean with varying seed coat colour. Food Res. Int. 2010, 43, 323–328. [Google Scholar] [CrossRef]
- Malenčić, D.; Maksimović, Z.; Popović, M.; Miladinović, J. Polyphenol contents and antioxidant activity of soybean seed extracts. Bioresour. Technol. 2008, 99, 6688–6691. [Google Scholar] [CrossRef] [PubMed]
- Whent, M.; Hao, J.; Slavin, M.; Zhou, M.; Song, J.; Kenworthy, W.; Yu, L. Effect of genotype, environment, and their interaction on chemical composition and antioxidant properties of low-linolenic soybeans grown in Maryland. J. Agric. Food Chem. 2009, 57, 10163–10174. [Google Scholar] [CrossRef] [PubMed]
- Gai, F.; Peiretti, P.G.; Karamać, M.; Amarowicz, R. Changes in the total polyphenolic content and antioxidant capacities of perilla (Perilla frutescens L.) plant extracts during the growth cycle. J. Food Qual. 2017, 2017, 7214747. [Google Scholar] [CrossRef]
- Gai, F.; Peiretti, P.G.; Karamać, M.; Amarowicz, R. Changes in antioxidative capacity and phenolic compounds in quinoa (Chenopodium quinoa Willd.) plant extracts during growth cycle. In Quinoa: Cultivation, Nutritional Properties and Effects on Health, 1st ed.; Peiretti, P.G., Gai, F., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2019. [Google Scholar]
- Peiretti, P.G.; Gai, F.; Karamać, M.; Amarowicz, R. Antioxidant activities and phenolic composition of safflower (Carthamus tinctorius L.) plant during their growth cycle. In Agricultural Research Updates; Gorawala, P., Mandhatri, S., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2017; Volume 19, pp. 23–39. [Google Scholar]
- United States Department of Agriculture. Soil taxonomy: A basic system of soil classification for making and interpreting soil surveys. In Agriculture Handbook Number 436, 2nd ed.; USDA, Natural Resources Conservation Service: Washington, DC, USA, 1999. [Google Scholar]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Karamać, M.; Kosińska, A.; Estrella, I.; Hernández, T.; Dueñas, M. Antioxidant activity of phenolic compounds identified in sunflower seeds. Eur. Food Res. Technol. 2012, 235, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Janiak, M.A.; Slavova-Kazakova, A.; Kancheva, V.D.; Ivanova, M.; Tsrunchev, T.; Karamać, M. Effects of γ-Irradiation of wild thyme (Thymus serpyllum L.) on the phenolic compounds profile of its ethanolic extract. Pol. J. Food Nutr. Sci. 2017, 67, 309–316. [Google Scholar] [CrossRef]
- Benzie, I.E.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Meth. Enzymol. 1990, 299, 15–27. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Popov, I.; Lewin, G. Oxidants and antioxidants part B—Antioxidative homeostasis: Characterization by means of chemiluminescent technique. Methods Enzymol. 1999, 300, 437–456. [Google Scholar]
- Karamać, M.; Pegg, R.B. Limitations of the tetramethylmurexide assay for investigating the Fe(II) chelation activity of phenolic compounds. J. Agric. Food Chem. 2009, 57, 6425–6431. [Google Scholar] [CrossRef] [PubMed]
- Karamać, M.; Kosińska-Cagnazzo, A.; Kulczyk, A. Use of different proteases to obtain flaxseed protein hydrolysates with antioxidant activity. Int. J. Mol. Sci. 2016, 17, 1027. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free-radical method to evaluate antioxidant activity. Lebensm.-Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Riedl, K.M.; Lee, J.H.; Renita, M.; St Martin, S.K.; Schwartz, S.J.; Vodovotz, Y. Isoflavone profiles, phenol content, and antioxidant activity of soybean seeds as influenced by cultivar and growing location in Ohio. J. Sci. Food Agric. 2007, 87, 1197–1206. [Google Scholar] [CrossRef]
- Slavin, M.; Cheng, Z.; Luther, M.; Kenworthy, W.; Yu, L. Antioxidant properties and phenolic, isoflavone, tocopherol, and carotenoid composition of Maryland-grown soybean lines with altered fatty acid profiles. Food Chem. 2009, 114, 20–27. [Google Scholar] [CrossRef]
- Chung, H.; Hogan, S.; Zhang, L.; Rainey, K.; Zhou, K. Characterization and comparison of antioxidant properties and bioactive components of Virginia soybeans. J. Agric. Food Chem. 2008, 56, 11515–11519. [Google Scholar] [CrossRef] [PubMed]
- Šibul, F.; Orčić, D.; Vasić, M.; Anačkov, G.; Nađpal, J.; Savić, A.; Mimica-Dukić, N. Phenolic profile, antioxidant and anti-inflammatory potential of herb and root extracts of seven selected legumes. Ind. Crops Prod. 2016, 83, 641–653. [Google Scholar] [CrossRef]
- Malenčić, D.; Popović, M.; Miladinović, J. Phenolic content and antioxidant properties of soybean (Glycine max (L.) Merr.) seeds. Molecules 2007, 12, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Song, H.H.; Ryu, H.W.; Kyung Jun Lee, K.J.; Jeong, I.J.; Kim, D.S.; Sei-Ryang Oh, S.-R. Metabolomics investigation of flavonoid synthesis in soybean leaves depending on the growth stage. Metabolomics 2014, 10, 833–841. [Google Scholar] [CrossRef]
- Lee, J.H.; Ha, T.J.; Baek, I.-Y.; Ko, J.M.; Cho, K.M.; Im, M.-H.; Choung, M.-G. Characterization of isoflavones accumulation in developing leaves of soybean (Glycine max) cultivars. J. Korean Soc. Appl. Biol. Chem. 2009, 52, 139–143. [Google Scholar] [CrossRef]
- Seo, W.D.; Jae Eun Kang, J.E.; Sik-Won Choi, S.-W.; Kwang-Sik Lee, K.-S.; Lee, M.-J.; Lee, J.H. Comparison of nutritional components (isoflavone, protein, oil, and fatty acid) and antioxidant properties at the growth stage of different parts of soybean [Glycine max (L.) Merrill]. Food Sci. Biotechnol. 2017, 26, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.P.; Strenge, D.; Miller, K.D. The role of glycosylation in flavonol-induced pollen germination. In Flavonoids in The Living System; Manthey, J.A., Buslig, B.S., Eds.; Plenum Press: New York, NY, USA, 1998; pp. 35–44. [Google Scholar]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef]
- Pourcel, L.; Grotewold, E. Participation of phytochemicals in plant development and growth. In Plant-Derived Natural Products; Osbourn, A.E., Lanzotti, V., Eds.; Springer: New York, NY, USA, 2009; pp. 269–279. [Google Scholar]
- Wang, Y.W.; Wang, W.C.; Jin, S.H.; Wang, J.; Wang, B.; Hou, B.K. Over-expression of a putative poplar glycosyltransferase gene, PtGT1, in tobacco increases lignin content and causes early flowering. J. Exp. Bot. 2012, 7, 2799–2808. [Google Scholar] [CrossRef]
- Yu, O.; Jung, W.; Shi, J.; Croes, R.A.; Fader, G.M.; Mc Gonigle, B.; Odell, J.T. Production of the isoflavones genistein and daidzein in non-legume dicot and monocot tissues. Plant Physiol. 2000, 124, 781–793. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Growth Stage | Days after Seeding | Extraction Yield | TPC | |
|---|---|---|---|---|
| (mg Catechin eq./g Extract) | (mg Catechin eq./g FM) | |||
| V5 | 34 | 19.2 ± 0.3 bc 1 | 47.7 ± 2.6 ab | 1.75 ± 0.09 ab |
| V6 | 41 | 21.0 ± 1.5 ab | 50.4 ± 1.1 a | 1.95 ± 0.002 a |
| R1 | 55 | 18.7 ± 1.4 bc | 44.3 ± 0.8 bc | 1.63 ± 0.002 bc |
| R2 | 62 | 19.2 ± 0.5 bc | 42.2 ± 2.3 c | 1.49 ± 0.09 cd |
| R3 | 69 | 17.6 ± 1.4 c | 43.8 ± 1.1 bc | 1.40 ± 0.04 d |
| R4 | 74 | 17.6 ± 1.1 c | 49.6 ± 3.7 a | 1.78 ± 0.10 ab |
| R5 | 78 | 18.1 ± 0.1 c | 50.4 ± 2.2 a | 1.82 ± 0.18 ab |
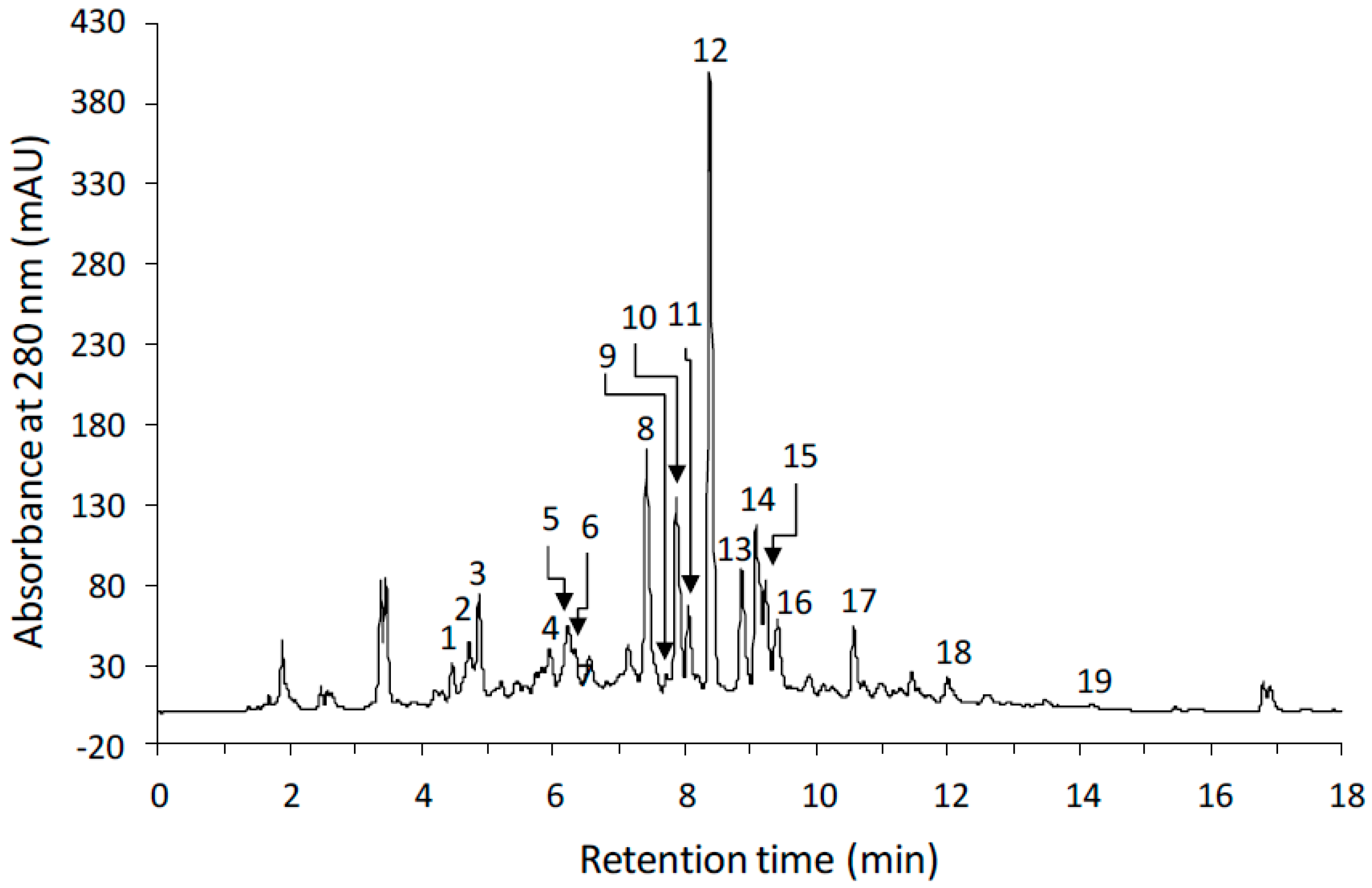
| No. | tR (min) | λMax (nm) | Compound | Content | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| V5 | V6 | R1 | R2 | R3 | R4 | R5 | ||||
| 1 | 4.46 | 328 | Hydroxycinnamic acid 1 | 0.34 ± 0.21 a | 0.47 ± 0.33 a | 0.39 ± 0.04 a | 0.50 ± 0.12 a | 0.45 ± 0.02 a | 0.44 ± 0.03 a | 0.45 ± 0.12 a |
| 2 | 4.73 | 328 | Hydroxycinnamic acid 1 | 1.23 ± 0.20 a | 1.37 ± 0.45 a | 1.08 ± 0.26 a | 1.38 ± 0.08 a | 1.27 ± 0.07 a | 0.98 ± 0.08 a | 1.21 ± 0.20 a |
| 3 | 4.86 | 328 | Hydroxycinnamic acid 1 | 1.54 ± 0.23 a | 1.43 ± 1.08 a | 1.41 ± 0.01 a | 1.72 ± 0.41 a | 1.54 ± 0.09 a | 1.64 ± 0.02 a | 1.52 ± 0.43 a |
| 4 | 5.95 | 315 | Hydroxycinnamic acid 1 | 0.53 ± 0.08 a | 0.53 ± 0.36 a | 0.54 ± 0.01 a | 0.67 ± 0.10 a | 0.57 ± 0.06 a | 0.57 ± 0.02 a | 0.57 ± 0.08 a |
| 5 | 6.22 | 321 | Hydroxycinnamic acid 1 | 0.71 ± 0.09 bc | 1.21 ± 0.12 a | 0.76 ± 0.03 bc | 0.94 ± 0.22 ab | 0.73 ± 0.06 bc | 0.63 ± 0.10 c | 0.70 ± 0.07 bc |
| 6 | 6.33 | 325 | Hydroxycinnamic acid 1 | 0.52 ± 0.06 a | 0.52 ± 0.19 a | 0.47 ± 0.04 a | 0.56 ± 0.10 a | 0.54 ± 0.06 a | 0.53 ± 0.03 a | 0.53 ± 0.08 a |
| 7 | 6.54 | 327 | Hydroxycinnamic acid 1 | 0.82 ± 0.14 ab | 0.96 ± 0.13 a | 0.69 ± 0.08 b | 0.81 ± 0.03 ab | 0.76 ± 0.10 ab | 0.67 ± 0.07 b | 0.79 ± 0.10 ab |
| 8 | 7.42 | 256,353 | Flavonol 2 | 6.80 ± 0.33 ab | 3.80 ± 2.97 b | 5.62 ± 0.04 ab | 6.97 ± 0.52 a | 6.02 ± 0.88 ab | 7.00 ± 0.89 a | 6.32 ± 1.02 ab |
| 9 | 7.71 | 251,295 | Daidzin | 0.67 ± 0.17 ab | 0.91 ± 0.49 ab | 1.50 ± 0.89 a | 1.11 ± 0.02 ab | 0.75 ± 0.10 ab | 0.59 ± 0.01 ab | 0.54 ± 0.12 b |
| 10 | 7.87 | 265,346 | Flavonol 3 | 4.19 ± 0.33 a | 3.27 ± 0.70 a | 3.97 ± 0.45 a | 4.32 ± 0.10 a | 3.97 ± 0.70 a | 4.37 ± 0.29 a | 3.89 ± 0.57 a |
| 11 | 8.05 | 255,353 | Flavonol 2 | 2.50 ± 0.12 a | 1.34 ± 1.08 b | 2.14 ± 0.01 ab | 2.46 ± 0.10 a | 2.28 ± 0.28 ab | 2.51 ± 0.20 a | 2.19 ± 0.32 ab |
| 12 | 8.37 | 257,355 | Rutin | 32.89 ± 1.10 a | 23.12 ± 5.08 b | 24.45 ± 0.72 b | 28.09 ± 1.91 ab | 23.23 ± 2.69 b | 29.18 ± 4.32 ab | 25.55 ± 4.43 ab |
| 13 | 8.87 | 265,348 | Flavonol 3 | 3.40 ± 0.15 a | 2.23 ± 1.12 a | 3.07 ± 0.54 a | 3.13 ± 0.14 a | 2.70 ± 0.23 a | 3.09 ± 0.26 a | 2.78 ± 0.40 a |
| 14 | 9.08 | 263,349 | Flavonol 3 | 5.02 ± 0.17 a | 2.61 ± 1.81 b | 3.91 ± 0.30 ab | 4.24 ± 0.20 ab | 3.78 ± 0.46 ab | 4.06 ± 0.39 ab | 3.70 ± 0.40 ab |
| 15 | 9.23 | 259 | Isoflavone 4 | 1.49 ± 0.2 a | 1.37 ± 1.11 a | 2.48 ± 1.06 a | 2.16 ± 0.02 a | 1.53 ± 0.07 a | 1.24 ± 0.07 a | 1.11 ± 0.16 a |
| 16 | 9.41 | 268 | Isoflavone 4 | 0.57 ± 0.01 a | 0.62 ± 0.23 a | 0.56 ± 0.13 a | 0.56 ± 0.01 a | 0.46 ± 0.06 a | 0.52 ± 0.04 a | 0.48 ± 0.06 a |
| 17 | 10.56 | 260 | Isoflavone 4 | 0.68 ± 0.19 a | 0.67 ± 0.57 a | 1.44 ± 0.61 a | 1.30 ± 0.10 a | 0.96 ± 0.04 a | 0.90 ± 0.16 a | 0.85 ± 0.13 a |
| 18 | 11.99 | 259 | Daidzein | 0.10 ± 0.05 c | 0.25 ± 0.23 bc | 0.71 ± 0.33 a | 0.67 ± 0.03 a | 0.52 ± 0.01 ab | 0.62 ± 0.05 ab | 0.53 ± 0.08 ab |
| 19 | 14.10 | 260 | Genistein | 0.05 ± 0.01 a | 0.06 ± 0.05 a | 0.11 ± 0.10 a | 0.11 ± 0.01 a | 0.11 ± 0.01 a | 0.08 ± 0.01 a | 0.05 ± 0.01 a |
| Sum of compounds | 64.05 ± 3.80 a | 46.75 ± 17.13 a | 55.30 ± 5.46 a | 61.71 ± 3.91 a | 52.17 ± 5.56 a | 59.61 ± 7.02 a | 53.76 ± 8.75 a | |||
| Sum of hydroxycinnamic acids | 5.69 ± 1.00 a | 6.49 ± 2.15 a | 5.33 ± 0.39 a | 6.59 ± 1.05 a | 5.86 ± 0.46 a | 5.46 ± 0.36 a | 5.76 ± 1.07 a | |||
| Sum of flavonols | 54.8 ± 2.19 a | 36.4 ± 12.76 b | 43.2 ± 1.96 ab | 49.2 ± 2.98 ab | 42.0 ± 5.24 ab | 50.2 ± 6.34 ab | 44.4 ± 7.14 ab | |||
| Sum of isoflavones | 3.56 ± 0.62 a | 3.88 ± 2.22 a | 6.80 ± 3.12 a | 5.90 ± 0.12 a | 4.33 ± 0.14 a | 3.95 ± 0.32 a | 3.56 ± 0.54 a | |||
| No. | tR (min) | λMax (nm) | Compound | Content | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| V5 | V6 | R1 | R2 | R3 | R4 | R5 | ||||
| 1 | 4.46 | 328 | Hydroxycinnamic acid 1 | 12.5 ± 7.7 a | 18.1 ± 12.3 a | 14.3 ± 1.1 a | 17.7 ± 4.2 a | 14.3 ± 0.7 a | 16.0 ± 0.9 a | 16.5 ± 5.1 a |
| 2 | 4.73 | 328 | Hydroxycinnamic acid 1 | 45.2 ± 7.1 a | 53.1 ± 16.2 a | 39.7 ± 8.8 a | 48.8 ± 2.9 a | 40.5 ± 2.3 a | 35.2 ± 2.2 a | 43.8 ± 9.7 a |
| 3 | 4.86 | 328 | Hydroxycinnamic acid 1 | 56.2 ± 8.5 a | 55.0 ± 40.4 a | 52.1 ± 1.5 a | 60.8 ± 14.7 a | 49.4 ± 3.0 a | 59.0 ± 0.3 a | 55.4 ± 18.5 a |
| 4 | 5.95 | 315 | Hydroxycinnamic acid 1 | 19.4 ± 2.9 a | 20.5 ± 13.6 a | 19.8 ± 0.0 a | 23.5 ± 3.7 a | 18.4 ± 2.0 a | 20.5 ± 0.4 a | 20.6 ± 4.2 a |
| 5 | 6.22 | 321 | Hydroxycinnamic acid 1 | 26.0 ± 3.1 c | 47.0 ± 5.8 a | 27.9 ± 1.5 bc | 33.3 ± 7.9 b | 23.2 ± 2.2 bc | 22.6 ± 3.2 c | 25.2 ± 3.8 bc |
| 6 | 6.33 | 325 | Hydroxycinnamic acid 1 | 18.9 ± 2.0 a | 19.9 ± 7.0 a | 17.3 ± 1.0 a | 19.9 ± 3.7 a | 17.2 ± 2.0 a | 18.9 ± 0.9 a | 19.2 ± 3.8 a |
| 7 | 6.54 | 327 | Hydroxycinnamic acid 1 | 30.1 ± 4.9 ab | 37.3 ± 6.1 a | 25.5 ± 2.5 b | 28.6 ± 1.0 ab | 24.4 ± 3.4 b | 24.3 ± 2.1 b | 28.5 ± 5.0 ab |
| 8 | 7.42 | 256,353 | Flavonol 2 | 249 ± 12 a | 146 ± 112 a | 207 ± 6 a | 246 ± 20 a | 193 ± 29 a | 252 ± 27 a | 229 ± 50 a |
| 9 | 7.71 | 251,295 | Daidzin | 24.5 ± 6.3 ab | 35.2 ± 18.1 ab | 54.8 ± 31.8 a | 39.0 ± 0.9 ab | 24.1 ± 3.0 ab | 21.2 ± 0.7 b | 19.7 ± 5.3 b |
| 10 | 7.87 | 265,346 | Flavonol 3 | 153 ± 12 a | 127 ± 24 a | 146 ± 14 a | 153 ± 4 a | 127 ± 23 a | 157 ± 8 a | 141 ± 28 a |
| 11 | 8.05 | 255,353 | Flavonol 2 | 91.5 ± 4.2 a | 51.6 ± 40.5 a | 78.8 ± 1.9 a | 86.9 ± 4.0 a | 72.9 ± 9.4 a | 90.5 ± 5.4 a | 79.4 ± 15.9 a |
| 12 | 8.37 | 257,355 | Rutin | 1203 ± 39 a | 895 ± 176 bc | 902 ± 8 bc | 991 ± 72 abc | 743 ± 90 c | 1050 ± 136 ab | 928 ± 211 abc |
| 13 | 8.87 | 265,348 | Flavonol 3 | 124 ± 5 a | 86 ± 42 a | 113 ± 17 a | 110 ± 6 a | 87 ± 8 a | 111 ± 7 a | 101 ± 20 a |
| 14 | 9.08 | 263,349 | Flavonol 3 | 184 ± 6 a | 101 ± 68 b | 144 ± 8 ab | 150 ± 8 ab | 121 ± 15 ab | 146 ± 11 ab | 134 ± 22 ab |
| 15 | 9.23 | 259 | Isoflavone 4 | 54.5 ± 7.5 a | 52.5 ± 41.7 a | 91.1 ± 37.2 a | 76.2 ± 0.3 a | 48.9 ± 2.0 a | 44.5 ± 1.7 a | 40.4 ± 8.1 a |
| 16 | 9.41 | 268 | Isoflavone 4 | 21.0 ± 0.4 a | 24.3 ± 9.4 a | 20.6 ± 4.2 a | 19.6 ± 0.3 a | 14.8 ± 2.0 a | 18.5 ± 1.2 a | 17.4 ± 3.2 a |
| 17 | 10.56 | 260 | Isoflavone 4 | 24.9 ± 7.0 a | 25.8 ± 21.5 a | 52.9 ± 21.5 a | 45.7 ± 3.3 a | 30.5 ± 1.1 a | 32.4 ± 5.3 a | 30.9 ± 6.4 a |
| 18 | 11.99 | 259 | Daidzein | 3.7 ± 1.6 c | 9.7 ± 8.9 bc | 26.0 ± 11.6 a | 23.6 ± 0.9 a | 16.7 ± 0.1 abc | 22.4 ± 1.3 ab | 19.1 ± 3.8 ab |
| 19 | 14.10 | 260 | Genistein | 1.7 ± 0.4 a | 2.1 ± 2.0 a | 4.2 ± 3.5 a | 4.0 ± 0.1 a | 3.5 ± 0.3 a | 3.0 ± 0.2 a | 1.8 ± 0.1 a |
| Sum of compounds | 2343 ± 136 a | 1806 ± 622 a | 2038 ± 160 a | 2177 ± 148 a | 1669 ± 187 a | 2145 ± 213 a | 1951 ± 424 a | |||
| Sum of hydroxycinnamic acids | 208 ± 30 a | 251 ± 78 a | 197 ± 10 a | 233 ± 38 a | 187 ± 16 a | 197 ± 9 a | 209 ± 50 a | |||
| Sum of flavonols | 2005 ± 77 a | 1406 ± 462 b | 1592 ± 40 ab | 1736 ± 113 ab | 1343 ± 175 b | 1806 ± 195 ab | 1612 ± 347 ab | |||
| Sum of isoflavones | 130 ± 22 a | 150 ± 83 a | 250 ± 110 a | 208 ± 3 a | 139 ± 4 a | 142 ± 9 a | 129 ± 27 a | |||
| Growth Stage | TEAC 1 | FRAP 2 | PCL-ACL 3 | Fe2+ Chelating Ability | |||
|---|---|---|---|---|---|---|---|
| (μmol Trolox eq./g Extract) | (μmol Trolox eq./g FM) | (μmol Fe2+/g Extract) | (μmol Fe2+/g FM) | (μmol Trolox eq./g Extract) | (μmol Trolox eq./g FM) | (%) | |
| V5 | 190 ± 20 c | 6.93 ± 0.71 ab | 780 ± 19 a | 28.5 ± 0.7 a | 552 ± 1 a | 20.2 ± 0.1 ab | 49.1 ± 5.2 ab |
| V6 | 201 ± 5 bc | 7.82 ± 0.03 ab | 694 ± 10 ab | 26.9 ± 1.0 a | 560 ± 38 a | 21.8 ± 2.0 a | 41.6 ± 6.4 bcd |
| R1 | 229 ± 39 bc | 8.43 ± 1.28 a | 651 ± 61 bc | 24.0 ± 1.8 ab | 517 ± 5 a | 19.1 ± 0.2 ab | 39.9 ± 4.7 cd |
| R2 | 177 ± 11 c | 6.26 ± 0.41 b | 623 ± 3 bc | 22.0 ± 0.2 b | 516 ± 36 a | 18.2 ± 1.4 ab | 46.4 ± 1.3 bc |
| R3 | 245 ± 21 ab | 7.82 ± 0.62 ab | 670 ± 77 ab | 21.4 ± 2.6 b | 552 ± 2 a | 17.6 ± 0.2 b | 51.7 ± 2.7 a |
| R4 | 220 ± 12 bc | 7.93 ± 0.29 ab | 712 ± 77 ab | 25.6 ± 2.3 ab | 534 ± 30 a | 19.2 ± 0.7 ab | 39.2 ± 2.2 cd |
| R5 | 219 ± 26 bc | 7.90 ± 0.95 ab | 666 ± 56 ab | 24.1 ± 3.3 ab | 531 ± 61 a | 19.3 ± 3.3 ab | 36.5 ± 0.8 d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peiretti, P.G.; Karamać, M.; Janiak, M.; Longato, E.; Meineri, G.; Amarowicz, R.; Gai, F. Phenolic Composition and Antioxidant Activities of Soybean (Glycine max (L.) Merr.) Plant during Growth Cycle. Agronomy 2019, 9, 153. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030153
Peiretti PG, Karamać M, Janiak M, Longato E, Meineri G, Amarowicz R, Gai F. Phenolic Composition and Antioxidant Activities of Soybean (Glycine max (L.) Merr.) Plant during Growth Cycle. Agronomy. 2019; 9(3):153. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030153
Chicago/Turabian StylePeiretti, Pier Giorgio, Magdalena Karamać, Michał Janiak, Erica Longato, Giorgia Meineri, Ryszard Amarowicz, and Francesco Gai. 2019. "Phenolic Composition and Antioxidant Activities of Soybean (Glycine max (L.) Merr.) Plant during Growth Cycle" Agronomy 9, no. 3: 153. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030153